Effects of Water Velocity and Specific Surface Area on Filamentous Periphyton Biomass in an Artificial Stream Mesocosm

Abstract
:1. Introduction
2. Materials and Methods
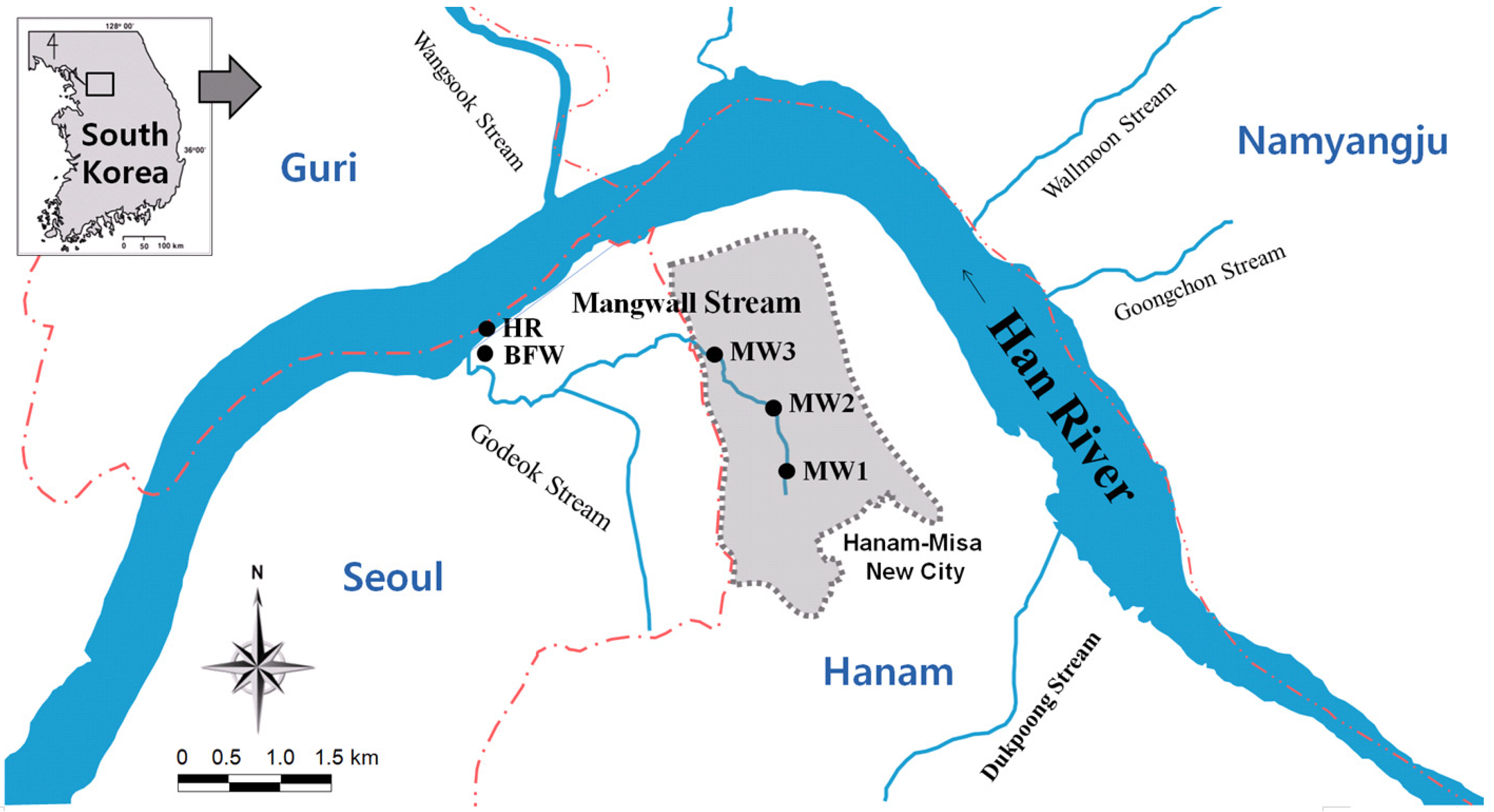
2.1. Research Site Overview
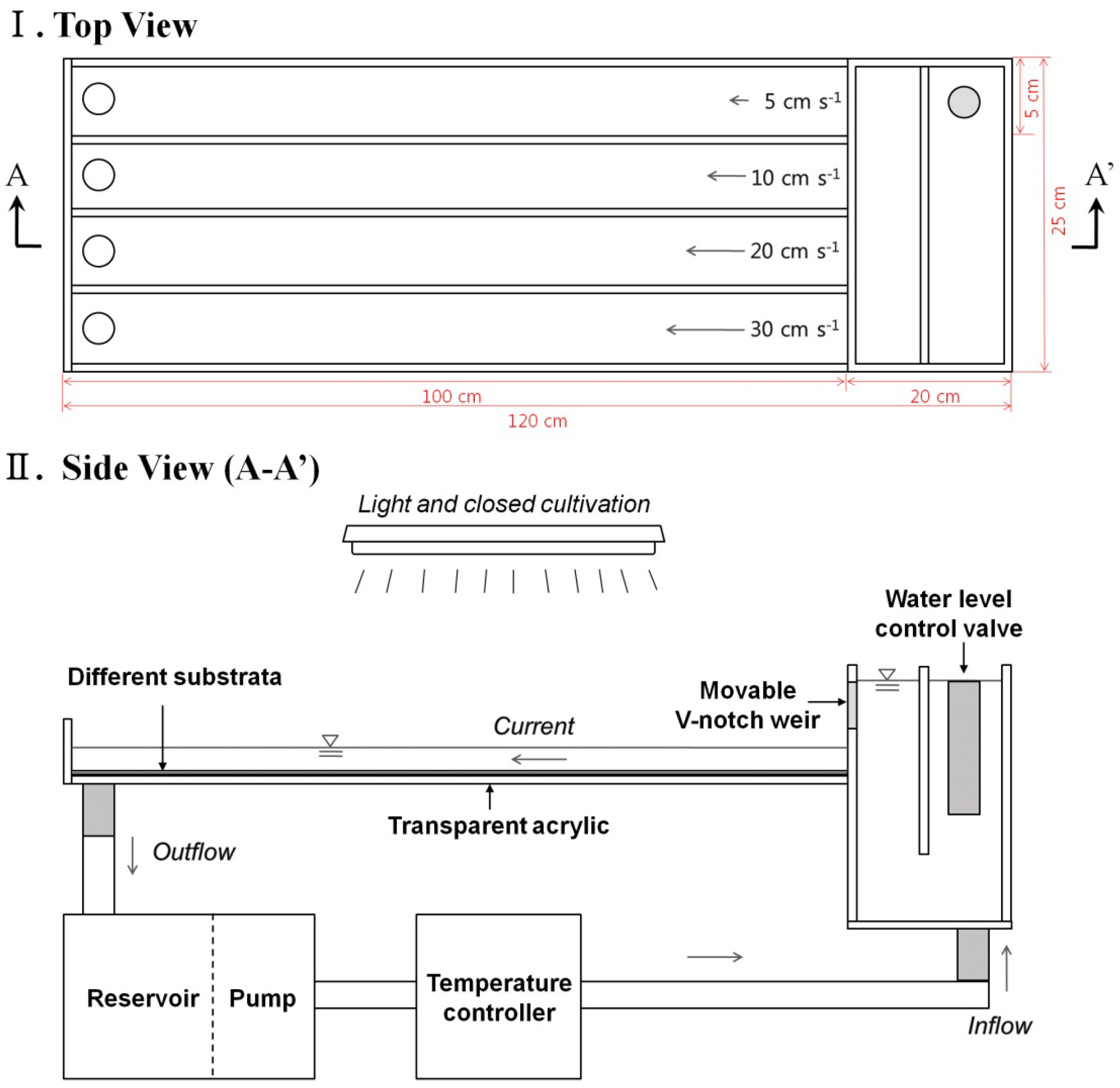
2.2. Artificial Stream Mesocosm Apparatus
2.3. Experimental Procedures
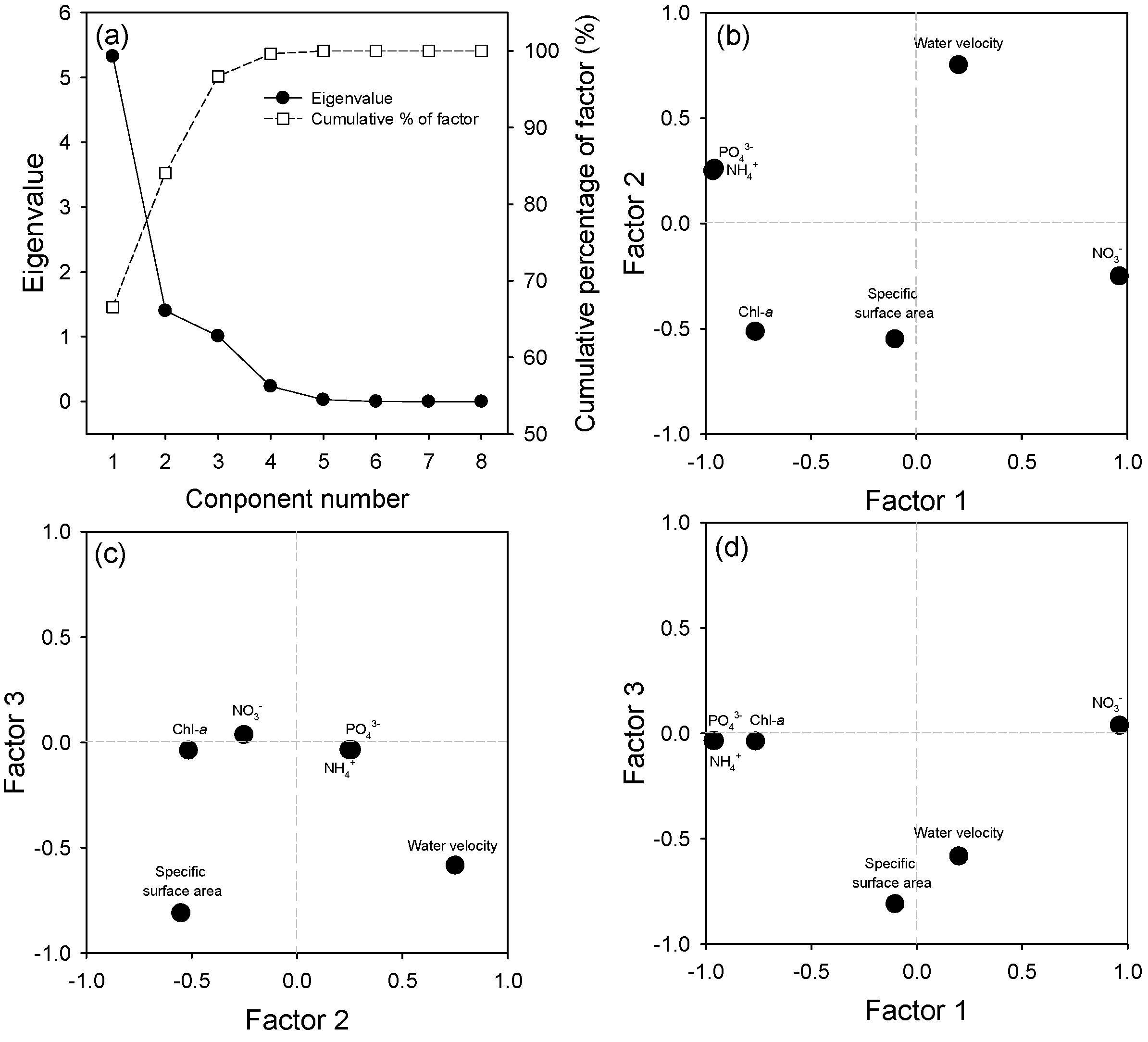
2.4. Principal Components Analysis (PCA)
3. Results and Discussion
3.1. Water Quality

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | MWa | HRb | BFWc | ||
|---|---|---|---|---|---|
| MW1 | MW2 | MW3 | |||
| Water depth (cm) | 6.3 ± 1.5d | 20.5 ± 1.7 | 31.5 ± 1.9 | 85.3 ± 9.1 | N.De |
| Water velocity (cm s−1) | 1 ± 1 | 6 ± 4 | 7 ± 4 | 16 ± 4 | N.D |
| Temperature (°C) | 24.4 ± 6.8 | 24.4 ± 6.9 | 24.4 ± 6.5 | 21.5 ± 5.5 | 17.3 ± 3.8 |
| pH | 7.6 ± 0.4 | 7.6 ± 0.4 | 7.6 ± 0.3 | 7.8 ± 0.3 | 7.4 ± 0.5 |
| DO (mg O2 L−1) | 6.5 ± 1.3 | 6.7 ± 1.3 | 6.5 ± 1.3 | 8.1 ± 0.4 | 7.0 ± 0.1 |
| Conductivity (µS cm−1) | 674 ± 93 | 670 ± 95 | 722 ± 74 | 167 ± 34 | 156 ± 29 |
| Salinity (‰) | 0.5 ± 0.1 | 0.5 ± 0.0 | 0.5 ± 0.0 | 0.1 ± 0.0 | 0.1 ± 0.0 |
| SS (mg L−1) | 4.3 ± 1.2 | 5.3 ± 1.5 | 5.2 ± 1.2 | 5.2 ± 1.7 | N.D |
| BOD (mg O2 L−1) | 6.3 ± 1.9 | 6.4 ± 2.0 | 6.6 ± 1.4 | 2.0 ± 0.7 | 0.9 ± 0.6 |
| CODMn (mg O2 L−1) | 7.3 ± 1.1 | 6.9 ± 1.1 | 7.0 ± 1.1 | 3.1 ± 0.6 | 2.1 ± 0.6 |
| TN (mg N L−1) | 6.9±1.5 | 7.1±1.6 | 7.0±1.5 | 2.6±0.7 | 2.2 ± 0.6 |
| NH4+ (µg N L−1) | 1241 ± 121 | 1384 ± 98 | 1395 ± 96 | 258 ± 55 | 183 ± 52 |
| NO3− (µg N L−1) | 716 ± 121 | 720 ± 128 | 707 ± 150 | 1644 ± 142 | 1649 ± 56 |
| TP (µg P L−1) | 632 ± 160 | 605 ± 192 | 620 ± 188 | 74 ± 7.7 | 59 ± 12.7 |
| PO43− (µg P L−1) | 425 ± 43 | 417 ± 50 | 413 ± 40 | 34 ± 8 | 28 ± 4 |
| Phytoplankton (µg chl-a L−1) | 2.0 ± 0.0 | 4.1 ± 1.8 | 5.6 ± 1.5 | 9.9 ± 0.0 | N.D |
| Periphyton (mg chl-a cm−2) | 7.0 ± 1.6 | 6.7 ± 1.3 | 6.5 ± 1.7 | 1.2 ± 8.3 | N.D |
| Description | MWa | HRb | BFWc | ||
|---|---|---|---|---|---|
| MW1* | MW2 | MW3 | |||
| Water depth (cm) | 7 | 22 | 34 | 85 | N.Dd |
| Water velocity (cm s−1) | 0 | 7 | 5 | 15 | N.D |
| Temperature (°C) | 19.2 | 18.9 | 19.1 | 18.7 | 13.3 |
| pH | 8.1 | 7.9 | 7.9 | 8.7 | 8.4 |
| DO (mg O2 L−1) | 7.3 | 7.2 | 7.7 | 8.3 | 7.2 |
| Conductivity (µS cm−1) | 925 | 1161 | 1079 | 220 | 147 |
| Salinity (‰) | 0.5 | 0.5 | 0.6 | 0.1 | 0.1 |
| SS (mg L−1) | 10.3 | 13.0 | 6.1 | 5.4 | N.D |
| BOD (mg O2 L−1) | 4.0 | 4.6 | 6.1 | 2.4 | 0.5 |
| CODMn (mg O2 L−1) | 6.3 | 6.7 | 7.0 | 3.2 | 0.9 |
| TN (mg N L−1) | 9.7 | 10.5 | 9.6 | 2.3 | 1.8 |
| NH4+ (µg N L−1) | 1512 | 1750 | 1664 | 302 | 189 |
| NO3− (µg N L−1) | 771 | 420 | 502 | 1550 | 1610 |
| TP (µg P L−1) | 823 | 842 | 790 | 52 | 45 |
| PO43− (µg P L−1) | 614 | 688 | 652 | 21 | 16 |
| Phytoplankton (µg chl-a L−1) | 2.5 | 5.2 | 6.0 | 9.9 | N.D |
| Periphyton (mg chl-a cm−2) | 8.8 | 8.3 | 8.2 | 2.3 | N.D |
3.2. Species Composition
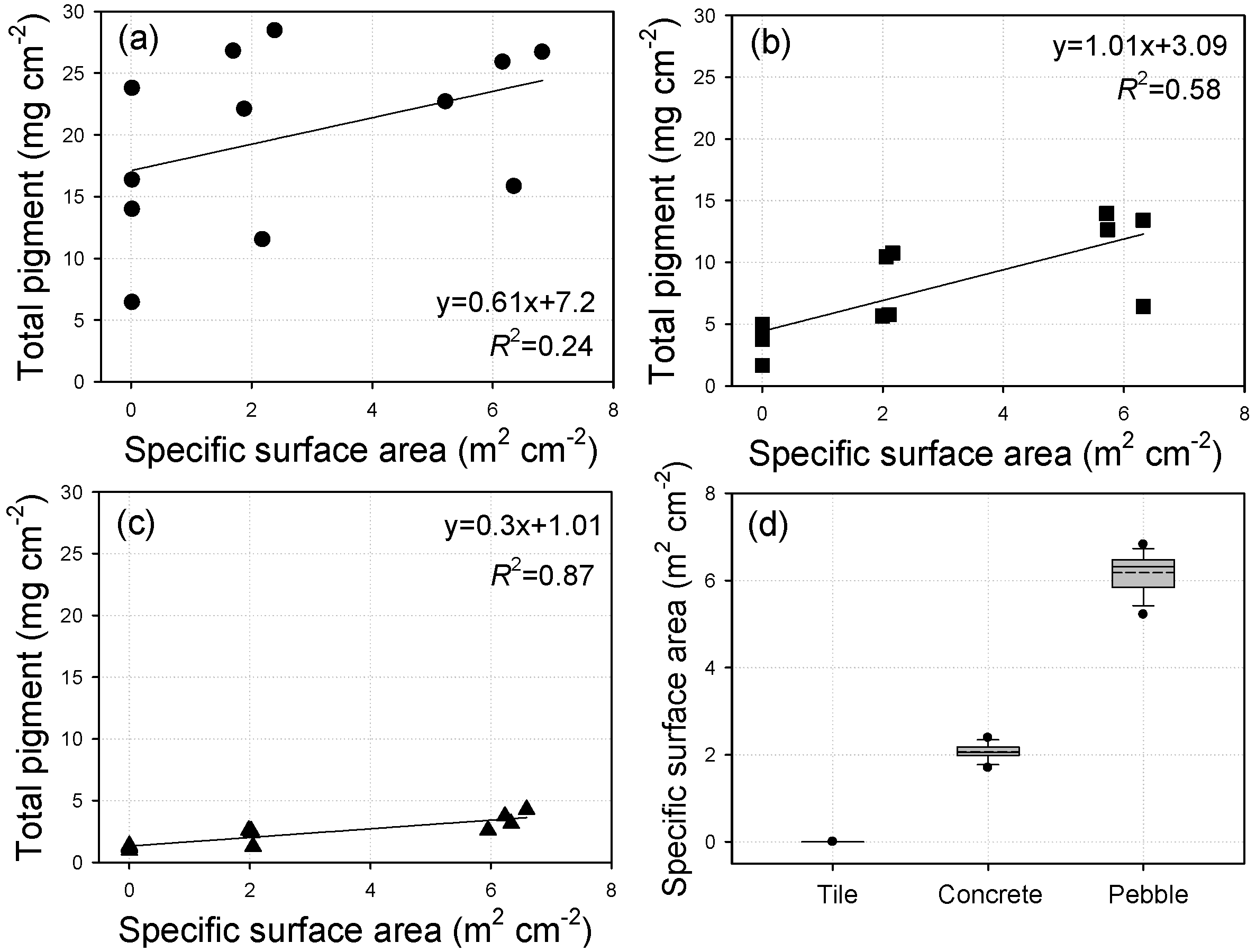
3.3. Surface Characteristics of Artificial Substrata
| Class | Species | MWa | HRb | BFWc | ||
|---|---|---|---|---|---|---|
| MW1* | MW2 | MW3 | ||||
| Cyanophyceae | Microcystis aeruginosa | N.Dd | +e | + | + | N.D |
| Phormidium sp. | 92.0 | 85.0 | 77.0 | N.D | ||
| Chlorophyceae | Closterium sp. | N.D | N.D | 3.0 | 4.0 | N.D |
| Oedogonium sp. | N.D | N.D | 4.0 | + | N.D | |
| Scenedesmus acuminatus | 5.0 | 5.0 | + | 13.0 | N.D | |
| Bacillariophyceae | Achnanthidium sp. | N.D | N.D | N.D | + | N.D |
| Asterionella formosa | N.D | N.D | 5.0 | 23.5 | N.D | |
| Cymbella minuta | N.D | N.D | N.D | 5.5 | N.D | |
| Fragilaria rumpens | N.D | N.D | N.D | 9.0 | N.D | |
| Melosira varians | N.D | N.D | + | 5.2 | N.D | |
| Navicula cryptocephala | N.D | N.D | N.D | + | N.D | |
| Nitzschia amphibia | + | 10.0 | 4.0 | 22.5 | N.D | |
| Stephanodiscus hantzschii | N.D | N.D | 4.0 | 6.3 | N.D | |
| Synedra ulna Ehrenberg | 3.0 | + | 3.0 | 11.0 | N.D | |
| Total (%) | 100.0 | 100.0 | 100.0 | 100.0 | N.D | |

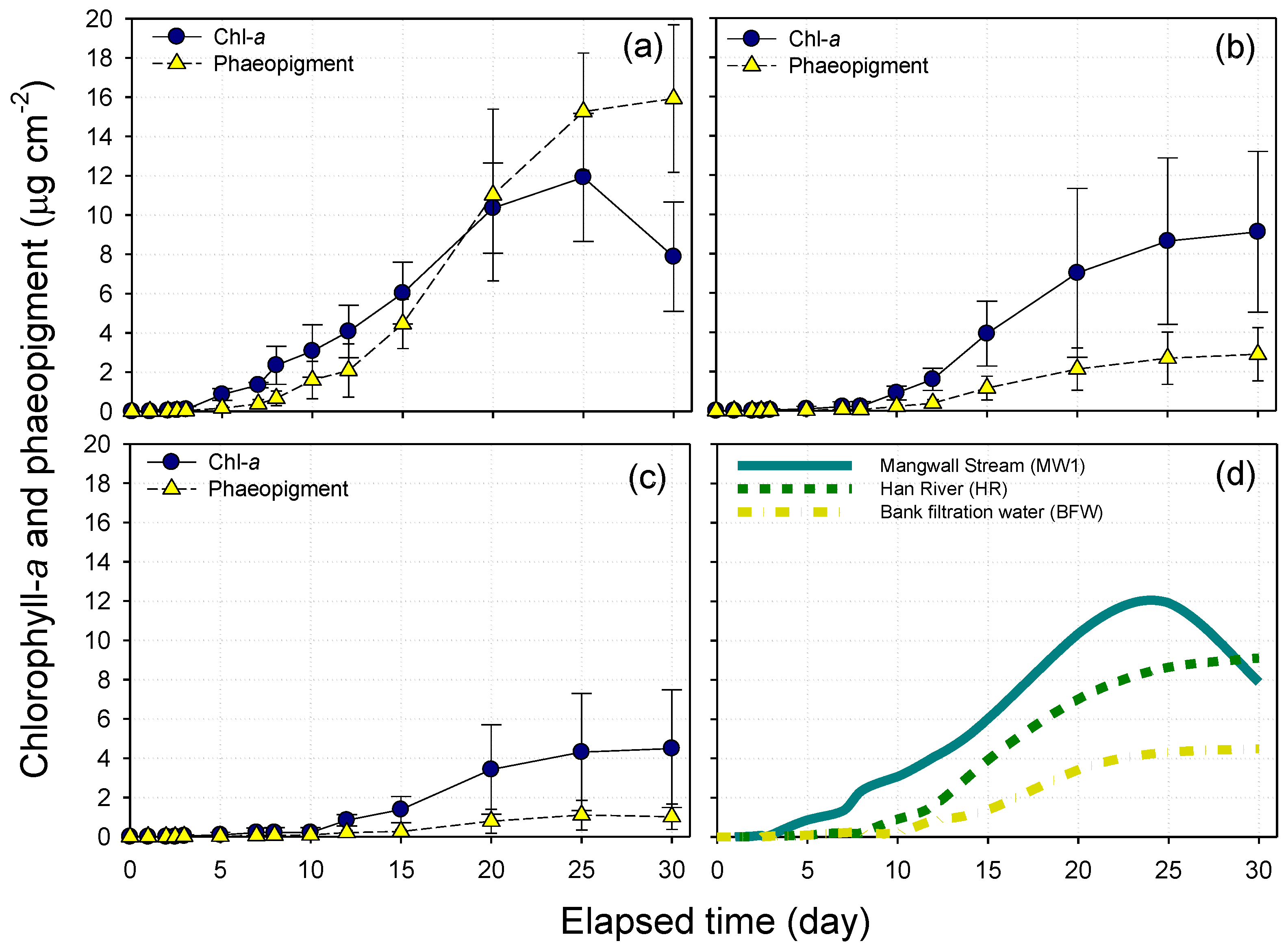
3.4. Periphyton Biomass Accrual Curves

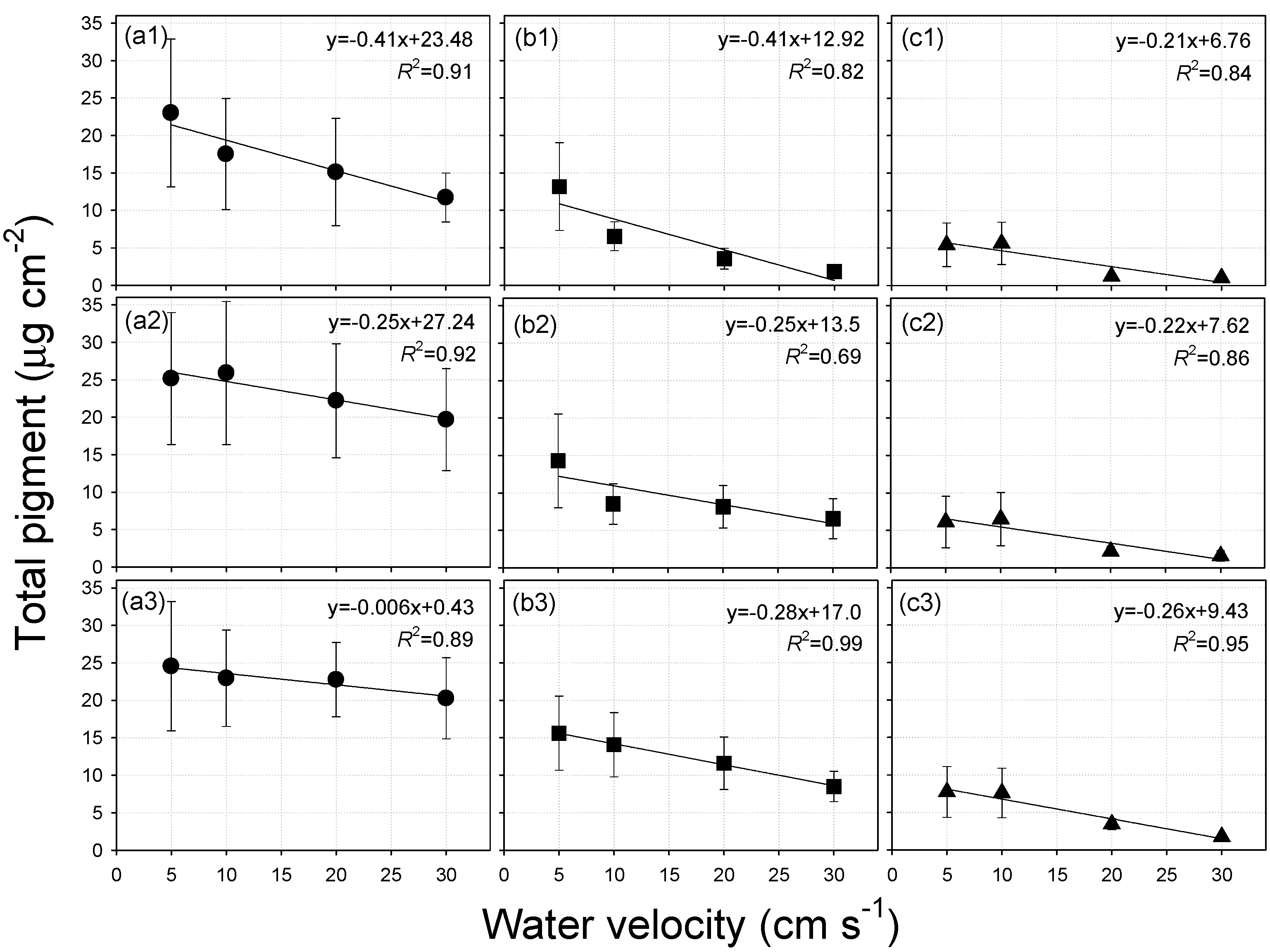
3.5. Relationships between Water Velocity and Periphyton Biomass

3.6. Relationships between Specific Surface Area of Substratum and Periphyton Biomass
3.7. Species Composition of Periphyton after the Artificial Stream Mesocosm Experiment
3.8. PCA Results

4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Sun, C.C.; Wang, Y.S.; Wu, M.L.; Dong, J.D.; Wang, Y.T.; Sun, F.L.; Zhang, Y.Y. Seasonal variation of water quality and phytoplakton response patterns in Daya Bay, China. Int. J. Environ. Res. Public Health 2011, 8, 2951–2966. [Google Scholar] [CrossRef]
- McCormick, P.V. Soil and periphyton indicators of anthropogenic water-quality changes in a rainfall-driven wetland. Wetl. Ecol. Manag. 2011, 19, 19–34. [Google Scholar] [CrossRef]
- McIntire, C.D. Periphyton dynamics in laboratory streams: A simulation model and its implications. Ecol. Mon. 1973, 43, 399–420. [Google Scholar] [CrossRef]
- Dos Santos, T.R.; Ferragut, C.; Bicudo, C.E.M. Does macrophyte architecture influence periphyton? Relationships among Utricularia foliosa, periphyton assemblage structure and its nutrient (C, N, P) status. Hydrobiologia 2013, 714, 71–83. [Google Scholar] [CrossRef]
- Ishida, C.K.; Arnon, S.; Peterson, C.G.; Kelly, J.J.; Gray, K.A. Influence of algal community structure on denitrification rates in periphyton cultivated on artificial substrata. Microb. Ecol. 2008, 56, 140–152. [Google Scholar] [CrossRef]
- Ghosh, M.; Gaur, J.P. Current velocity and the establishment of stream algal periphyton communities. Aquat. Bot. 1998, 60, 1–10. [Google Scholar] [CrossRef]
- Montuelle, B.; Dorigo, U.; Bérard, A.; Volat, B.; Bouchez, A.; Tlili, A.; Gouy, V.; Pesce, S. The periphyton as a multimetric bioindicator for assessing the impact of land use on rivers: An overview of the Ardières-Morcille experimental watershed. Hydrobiologia 2010, 659, 123–141. (in France). [Google Scholar]
- Dempster, P.W.; Beveridge, M.C.M.; Baird, D.J. Herbivory in tilapia Oreochromis niloticus: acomparison of feeding rates on phytoplankton and periphyton. J. Fish Biol. 1993, 43, 385–392. [Google Scholar] [CrossRef]
- Kersters, I.; Vooren, L.V.; Huys, G.; Janssen, P.; Kersters, K.; Verstraete, W. Influence of temperature and process technology on the occurrence of Aeromonas species and hygienic Indicator organisms in drinking water production plants. Microb. Ecol. 1995, 30, 203–218. [Google Scholar]
- Serra, A.; Guasch, H.; Admiraal, W.; van der Geest, H.G.V.; van Beusekom, S.A.M. Influence of phosphorus on copper sensitivity of fluvial periphyton: The role of chemical, physiological and community-related factors. Ecotoxicology 2010, 19, 770–780. [Google Scholar] [CrossRef]
- Baffico, G.D. Variations in the periphytic community structure and dynamics of Lake Nahuel Huapi (Patagonia, Argentina). Hydrobiologia 2001, 455, 79–85. [Google Scholar] [CrossRef]
- Flum, T.; Huxel, G.L.; LaRue, C.S.; Hardison, B.; Duncan, J.R.; Drake, J.A. A closed artificial stream for conducting experiments requiring a controlled species pool. Hydrobiologia 1993, 271, 75–85. [Google Scholar] [CrossRef]
- Horner, R.R.; Welch, E.B.; Seeley, M.R.; Jacoby, J.M. Responses of periphyton to changes in current velocity, suspended sediment and phosphorus concentration. Freshw. Biol. 1990, 24, 215–232. [Google Scholar] [CrossRef]
- Biggs, B.J.F.; Goring, D.G.; Nikora, V.I. Subsidy and stress responses of stream periphyton to gradients in current velocity as a function of community growth form. J. Phycol. 1998, 34, 598–607. [Google Scholar]
- Sekar, R.; Nandakumar, K.; Venugopalan, V.P.; Nair, K.V.K.; Rao, N.R. Spatial variation in microalgal colonization on hard surfaces in a lentic freshwater environment. Biofouling 1998, 13, 177–195. [Google Scholar] [CrossRef]
- Godillot, R.; Caussade, B.; Ameziane, T.; Capblancq, J. Interplay between turbulence and periphyton in rough open-channel flow. J. Hydraul. Res. 2001, 39, 227–239. [Google Scholar]
- Horner, R.R.; Welch, E.B. Stream periphyton development in relation to current velocity and nutrients. Can. J. Fish. Aqut. Sci. 1981, 38, 449–457. [Google Scholar] [CrossRef]
- McIntire, C.D. Some effects of current velocity on periphyton communities in laboratory streams. Hydrobiologia 1966, 27, 559–570. [Google Scholar] [CrossRef]
- Labiod, C.; Godillot, R.; Caussade, B. The relationship between stream periphyton dynamics and near-bed turbulence in rough open-channel flow. Ecol. Model. 2007, 209, 78–96. [Google Scholar] [CrossRef]
- Dodds, W.K.; Biggs, B.J.F. Water velocity attenuation by stream periphyton and macrophytes in relation to growth form and architecture. J. North. Am. Benthol. Soc. 2002, 21, 2–15. [Google Scholar] [CrossRef]
- Herder-Brouwer, S.J. The development of periphyton on artificial substrates. Hydrobiol. Bull. 1975, 9, 81–86. [Google Scholar]
- Cullinane, J.; Maguire, D.; Whelan, P. The importance of substrate type in colonization by Cryptonemia hibernica Guiry et Irvine and its associated algae. Hydrobiologia 1984, 116, 438–442. [Google Scholar] [CrossRef]
- Zhang, N.; Li, H.; Jeppesen, E.; Li, W. Influence of substrate type on periphyton biomass and nutrient state at contrasting high nutrient levels in a subtropical shallow lake. Hydrobiologia 2013, 710, 129–141. [Google Scholar] [CrossRef]
- Lee, J.S. Measuring the economic benefits of residential water quality improvement in Ulsan, Korea using a contingent valuation. Urban Water J. 2013, 10, 1–8. [Google Scholar] [CrossRef]
- Jeong, D.I. Trends and multi-decadal variability of annual maximum precipitation for Seoul, South Korea. Urban Water J. 2009, 9, 431–439. [Google Scholar] [CrossRef]
- Clinton, B.D.; Vose, J.M. Variation in stream water quality in an urban headwater stream in the southern Appalachians. Water Air Soil Pollut. 2006, 169, 331–353. [Google Scholar] [CrossRef]
- Paul, M.J.; Meyer, J.L. Streams in the urban landscape. Ann. Rev. Ecol. Syst. 2001, 32, 333–365. [Google Scholar] [CrossRef]
- Magoulick, D.D. Spatial and temporal variation in fish assemblages of drying stream pools: The role of abiotic and biotic factors. Aquat. Ecol. 2000, 34, 29–41. [Google Scholar] [CrossRef]
- Kim, B.-H. Ecology of a cyanobacterial mat community in a Korean thermal wastewater stream. Aquat. Ecol. 1999, 33, 331–338. [Google Scholar] [CrossRef]
- Harada, S.; Wagatsuma, R.; Koseki, T.; Aoki, T.; Hashimoto, T. Water quality criteria for water bodies in urban areas and accompanying changes in surrounding and in-situ vegetation: considerations from the landscape aspect of planning water recreational areas. J. Water Resour. Prot. 2013, 5, 156–163. [Google Scholar] [CrossRef]
- Kumar, S.; Tripathi, V.R.; Garg, S.K. Physicochemical and microbiological assessment of recreational and drinking waters. Environ. Monit. Assess. 2012, 184, 2691–2698. [Google Scholar] [CrossRef]
- Massoud, M.A. Assessment of water quality along a recreational section of the Damour River in Lebanon using the water quality index. Environ. Monit. Assess. 2012, 184, 4151–4160. [Google Scholar] [CrossRef]
- Kindsvater, C.E.; Carter, R.W.C. Discharge characteristics of rectangular thin-plate weirs. J. Hydro. Div. 1957, 83, 1–36. [Google Scholar]
- Wetzel, R.G.; Likens, G.E. Limnological Analysis, 3rd ed.; Springer-Verlag: New York, NY, USA, 2000; pp. 1–429. [Google Scholar]
- American Public Health Association (APHA). Standard Methods for the Examination of Water and Wastewater, 21th ed.; APHA: Washington, DC, USA, 2005; pp. 9–72. [Google Scholar]
- LeBouf, R.F.; Ku, B.K.; Chen, B.T.; Frazer, D.G.; Cumpston, J.L.; Stefaniak, A.B. Measuring surface area of airborne titanium dioxide powder agglomerates: Relationships between gas adsorption, diffusion and mobility-based methods. J. Nanopart. Res. 2011, 13, 7029–7039. [Google Scholar] [CrossRef]
- Talbot, P.; de la Noüe, J. Tertiary treatment of wastewater with Phormidium bohneri (Schmidle) under various light and temperature conditions. Water Res. 1993, 27, 153–159. [Google Scholar] [CrossRef]
- Biggs, B.J.F. New Zealand Periphyton Guidelines: Detecting, Monitoring and Managing the Enrichment of Rivers. Background and Guidelines; Ministry for the Environment: Wellington, New Zealand, 2000; pp. 1–122. [Google Scholar]
- Scheffer, M.; Rinaldi, S.; Gragnani, A.; Mur, L.R.; Nes, E.H.V. On the dominance of filamentous cyanobacteria in shallow, Turbid Lakes. Ecology 1997, 78, 272–282. [Google Scholar] [CrossRef]
- Azim, M.E.; Milstein, A.; Wahab, M.A.; Verdegama, M.C.J. Periphyton-water quality relationships in fertilized fishponds with artificial substrates. Aquaculture 2003, 228, 169–187. [Google Scholar] [CrossRef]
- Yentsch, C.S. Distribution of chlorophyll and phaeophytin in the open ocean. Deep Sea Res. 1965, 12, 653–666. [Google Scholar]
- Carpenter, S.R.; Leavitt, P.R.; Elser, J.J.; Elser, M.M. Chlorophyll budgets: Response to food web manipulation. Biogeochemistry 1988, 272, 79–90. [Google Scholar]
- Alfi, R.; Guiral, D. Chlorophyll budget in a productive tropical pond: Algal production, sedimentation, and grazing by microzooplankton and rotifers. Hydrobiologia 1994, 272, 239–249. [Google Scholar] [CrossRef]
- Fovet, O.; Belaud, G.; Litrico, X.; Charpentier, S.; Bertrand, C.; Dauta, A.; Hugodot, C. Modelling periphyton in irrigation canals. Ecol. Model. 2010, 221, 1153–1161. [Google Scholar] [CrossRef]
- Whitford, L.A.; Schumacher, G.J. Effect of current respiration and mineral uptake in Spirogyra and Oedogonium. Ecology 1964, 45, 168–170. [Google Scholar] [CrossRef]
- Borchardt, M.A.; Hoffman, P.; Cook, P.W. Phosphorus uptake kinetics of Spirogyra fluviatilis (Charophyseae) in flowing water. J. Phycol. 1994, 30, 403–417. [Google Scholar]
- Stevenson, R.J. Effects of current and conditions simulating autogenically changing microhabitats on benthic diatom immigration. Ecology 1983, 64, 1514–1281. [Google Scholar] [CrossRef]
- Sumina, E.L. Behavior of filamentous cyanobacteria in laboratory culture. Microbiology 2006, 75, 459–464. [Google Scholar] [CrossRef]
- Smith, V.H. Low nitrogen to phosphorus ratios favor dominance by blue-green algae in lake phytoplankton. Science 1983, 221, 669–671. [Google Scholar]
- Laliberté, G.; Lessard, P.; Noüe, J.; Sylvestre, S. Effect of phosphorus addition on nutrient removal from wastewater with the cyanobacteriom Phormidium bohneri. Bioresour. Technol. 1997, 59, 227–233. [Google Scholar] [CrossRef]
- Maria, J.B.; Graca, C. Identification of similar environmental areas in Tagus Estuary by using multivariate analysis. Ecol. Ind. 2006, 6, 508–515. [Google Scholar] [CrossRef]
- Liu, C.W.; Lin, K.H.; Kuo, Y.N. Application of factor analysis in the assessment of groundwater quality in a blackpoot disease area in Taiwan. Sci. Tot. Environ. 2003, 313, 77–89. [Google Scholar] [CrossRef]
- Yerel, S.; Ankara, H. Application of multivariate statistical techniques in the assessment of water quality in Sakarya River, Turkey. J Geol Soc India 2012, 79, 89–93. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ahn, C.H.; Song, H.M.; Lee, S.; Oh, J.H.; Ahn, H.; Park, J.-R.; Lee, J.M.; Joo, J.C. Effects of Water Velocity and Specific Surface Area on Filamentous Periphyton Biomass in an Artificial Stream Mesocosm. Water 2013, 5, 1723-1740. https://doi.org/10.3390/w5041723
Ahn CH, Song HM, Lee S, Oh JH, Ahn H, Park J-R, Lee JM, Joo JC. Effects of Water Velocity and Specific Surface Area on Filamentous Periphyton Biomass in an Artificial Stream Mesocosm. Water. 2013; 5(4):1723-1740. https://doi.org/10.3390/w5041723
Chicago/Turabian StyleAhn, Chang Hyuk, Ho Myeon Song, Saeromi Lee, Ju Hyun Oh, Hosang Ahn, Jae-Roh Park, Jung Min Lee, and Jin Chul Joo. 2013. "Effects of Water Velocity and Specific Surface Area on Filamentous Periphyton Biomass in an Artificial Stream Mesocosm" Water 5, no. 4: 1723-1740. https://doi.org/10.3390/w5041723
APA StyleAhn, C. H., Song, H. M., Lee, S., Oh, J. H., Ahn, H., Park, J.-R., Lee, J. M., & Joo, J. C. (2013). Effects of Water Velocity and Specific Surface Area on Filamentous Periphyton Biomass in an Artificial Stream Mesocosm. Water, 5(4), 1723-1740. https://doi.org/10.3390/w5041723