
Widespread Geographical Disparities in Phytoplankton Ecology Research in the Face of Climate Change: A Review

1
Africa Centre of Excellence in Coastal Resilience (ACECoR), University of Cape Coast, Cape Coast 00223, Ghana
2
Department of Fisheries and Aquatic Sciences, School of Biological Sciences, University of Cape Coast, Cape Coast 00223, Ghana
3
National Commission for Science, Technology, and Innovation, Nairobi 30623-00100, Kenya
4
Institute for Water Research, Rhodes University, Grahamstown 6140, South Africa
*
Author to whom correspondence should be addressed.
Water 2023, 15(24), 4288; https://doi.org/10.3390/w15244288
Submission received: 4 September 2023
/
Revised: 26 October 2023
/
Accepted: 31 October 2023
/
Published: 15 December 2023
(This article belongs to the Special Issue Biodiversity and Functionality of Plankton Communities)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Climate change is a global threat that is better understood through global research spanning many regions, countries, and life-supporting ecosystems. This review focused on the bibliometric analysis of the distribution of a global research compendium on phytoplankton, an aquatic community that accounts for half of the carbon fixation in the global carbon cycle. A total of 716 primary research papers published in the immediate past decade (2012–2022) were reviewed. The articles were sampled from Web of Science and described field and laboratory experiments quantifying the impact of global climate change on phytoplankton from different climate zones (tropical, subtropical, temperate, subpolar, and polar) and ecosystems (freshwater, brackish, and marine). Analyses of these studies suggest that the bulk of the global research effort (80%) focused on high-latitude areas, and only a few (17%) were carried out in the tropics, largely led by four countries (Australia, Brazil, India, and Saudi Arabia), while Africa’s contribution was minuscule (<1%). In terms of ecosystems, the majority (76%) of the research was in marine waters, irrespective of the climate zone. Analyses of these studies also highlighted widespread disparities in phytoplankton research in tropical aquatic ecosystems, particularly in Africa. Strategic investment in terms of targeted funding, institutional networks, and partnerships between the global north and global south are necessary to increase phytoplankton research across different ecosystems in the tropics.
1. Introduction
Aquatic ecosystems cover about 71% of the Earth’s surface and comprise diverse habitats ranging from freshwater to brackish and marine environments [1]. These ecosystems provide critical functions important for achieving the sustainable development goals related to biodiversity conservation and livelihoods [2]. Now, it is common knowledge that these aquatic ecosystems and habitats in freshwater, brackish, and marine environments are under pressure from multiple stressors and factors related to pollution, global climate change, and other human activities [3,4]. Only 13% of the world’s marine environments are currently considered pristine or untouched by human impacts [5].
For coastal ecosystems in particular, estimates suggest that about 50% of salt marshes, 35% of mangroves, 30% of coral reefs, and 29% of seagrasses are already lost or degraded worldwide due to human activities [6]. This degradation is expected to be even more accelerated with the increasing urbanization of riparian/coastal communities and changes in land use, particularly for agriculture and other industrial activities [7]. This threatens the objectives of the global ocean decade as well as the sustainable development goals (SDG) to achieve food security (SDG 2) and decent economic growth (SDG 8) through sustainable management of life below water (SDG 14) [8]. There is therefore an urgent call for concerted, global action to understand and possibly predict the response(s) of aquatic ecosystems and organisms to the combination of multiple environmental stressor factors.
1.1. Climate Change and Its Impacts on Aquatic Ecosystems
Climate change represents one of the most important sources of environmental change in aquatic ecosystems [9,10]. Climate change has resulted in five critical global environmental changes: the warming temperature of the Earth’s surface and the oceans, changes in the global water cycle (“hydrologic” cycle), declining glaciers and snowpack, sea level rise, and ocean acidification. These changes in the environment are primarily driven by the emissions of greenhouse gases in the atmosphere from anthropogenic sources, such as the combustion of fossil fuels, coupled with the resultant warming of the Earth’s surface. Reports from the Intergovernmental Panel for Climate Change (IPCC) have shown that the world has been warming gradually, with some studies estimating that these temperature increases have doubled over the last 50 years alone [9,10,11]. This warming will potentially have disproportional catastrophic effects on various ecosystems by altering their structures and functions, particularly those adapted to specific climatic conditions [10,11,12,13].
Many aquatic organisms, particularly within the tropics and polar regions, have evolved to survive within specific ranges of environmental conditions [14,15]. Climate change impacts may create physical and biological conditions that the majority of these organisms have previously not experienced within their evolutionary history [16], thereby pushing them towards extinction [13,15]. In particular, research has shown that many of the species found in tropical marine and coastal ecosystems are already living on the edge of their thermal tolerances [13,17]. In addition, climate change impacts such as heatwaves are also more prevalent in tropical regions, thereby altering the biodiversity, productivity, and potential for aquatic ecosystems to provide functions and services [13]. There is therefore a need to increase our understanding of how climate change impacts aquatic biodiversity and productivity, particularly in tropical ecosystems [15,18].
Regarding the impacts of global climate change in particular, phytoplankton play a key role in regulating the balance of carbon in the atmosphere. With a turnover rate of about one week [19], phytoplankton account for half of the carbon fixation in the global carbon cycle; they release a large fraction (>90%) of the organic matter that fuels ocean carbon sequestration and burial [20,21]. The sum of these processes controls more carbon than the amount available in the atmosphere [22,23]. Phytoplankton research is therefore considered imperative for ocean science [24] and requires insight into the evolution of phytoplankton communities in different climate regions of the world [25]. This is because phytoplankton species diversity, function (e.g., carbon fixation), and climate sensitivity differ among species from different climate zones [19,25,26]. Anthropogenic (e.g., pollution) and natural forces (e.g., upwelling) that impact phytoplankton biology are also not equally distributed around the world [25].
The purpose of this report is to present the trends in global phytoplankton research in the last decade (2012–2022), with a special focus on work evaluating impacts associated with global climate change. The objective is to provide a reference point for interventions to strengthen phytoplankton research in tropical areas.
1.2. Phytoplankton as Indicators of Climate Change
Phytoplankton represent a taxonomically diverse group of photosynthetic, mostly single-celled aquatic organisms that drift with the current. There are approximately 20,000 species of phytoplankton that are distributed among eight phyla. Phytoplankton can be divided into three distinct groups: diatoms, dinoflagellates, and coccolithophorids and microflagellates. Unlike terrestrial plants, which have more than 250,000 recorded species, phytoplankton are poor in species diversity but are phylogenetically diverse [27].
Apart from bacteria, phytoplankton are the most abundant life form in pelagic ecosystems [28]. They have short life cycles and are amenable to subtle variations in the environment [29,30,31]. Diatoms, for example, can adapt to warming over a period of about three weeks after 300 generations [19,32]. Dinoflagellates, including species (Prorocentrum micans) common in cold temperate to tropical waters, are also able to adapt to elevated pH levels over short cycles of 34–126 generations [33]. The calcification of coccolithophorids such as Emiliania huxleyi is also adaptable to elevated temperatures and carbon dioxide levels after seven generations [34]. This ability to respond quickly to changes in the environment qualifies microalgae as good reference indicators for assessing the impact of global climate change [30,35].
Current efforts to evaluate the response of aquatic ecosystems to climate change and other anthropogenic factors involve the use of water quality criteria, usually derived from studies testing the response of sensitive organisms [36]. These analyses typically consider the response of keystone organisms with large populations distributed across different ecosystem zones [29,37]. Microalgae are a good example of these keystone organisms because of the significant role they play in aquatic ecosystems [30]. These organisms modulate the efficiency of aquatic food webs with consequences for the global carbon cycle, food security, and livelihood opportunities (e.g., fisheries) in many communities [21,38,39].
In addition to primary production, phytoplankton also play a significant role in the carbon (C), nitrogen (N), phosphorus (P), iron (Fe), and silicon (Si) biochemical cycles. They utilise these compounds for their vital processes, and in these processes, they reintroduce them back to the environment as either particulate or dissolved organic matter, which is either remineralised by heterotrophs and transferred to higher trophic levels or sinks to form elemental compositions in deeper waters. Studies have highlighted that understanding how phytoplankton take up these inorganic nutrients and allocate their resources to undertake physiological processes is important in understanding the present, past, and future linkages to these important biogeochemical cycles [40].
Phytoplankton also play an important role in influencing the Earth’s climate and the functioning of the biological carbon pump. The biological carbon pump is a major component of the global carbon cycle and refers to the process by which atmospheric CO2 is transferred by primary producers (mostly phytoplankton) from the eutrophic zone of the ocean to the underlying sediments [41]. Marine ecosystems provide the major sink for atmospheric CO2 and account for the removal of approximately one-third of all anthropogenic CO2 emissions from the atmosphere [42]. The conversion of atmospheric CO2 to ocean sediment is a direct result of the combined effect of solubility and the biological pump [41]. In addition, the effectiveness of the biological pump depends highly on phytoplankton physiology and community structure. Phytoplankton primarily drive the biological pump through primary production, where they convert inorganic carbon into organic matter. As a result, after their consumers and bacteria that feed on their waste die, they are transported down into sediments, where they are locked out of circulation for centuries.
Despite their considerable importance, some phytoplankton species may have direct, devastating impacts on humans and animals through the production of toxic algal blooms. Algal blooms refer to the above-average outbreak of phytoplankton cells within a given body of water, which occur during peaks in the annual cycle of phytoplankton biomass and chlorophyll concentrations [43]. These blooms occur when the rate of phytoplankton assemblage exceeds their normal mortality rates, often facilitated by the occurrence of excess growth-limiting nutrients such as nitrogen and phosphorus in the environment. As a result, the phytoplankton assemblages accumulate in the water column until the limiting nutrients are depleted. Out of the thousands of phytoplankton species, only less than 5% can form algal blooms [43]. When they occur, HABs pose significant effects to human and biodiversity health, recreation, and aquaculture. It is projected that stressors such as pollution and climate change will influence aquatic planktonic systems, thereby increasing the frequency and intensity of harmful algal blooms [44].
The diversity, growth, and development of plankton communities are greatly tied to environmental parameters such as nutrient availability, light regimes, temperature, alkalinity, and pH [21,45]. Climate change and its associated impacts on aquatic environments alter these environmental parameters and, subsequently, phytoplankton community structures [20]. As a result, studies in phytoplankton ecology have always sought to understand how these organisms respond to environmental stressors emanating from anthropogenic activities [19,46,47,48,49,50,51]. In this review, we highlight the efforts being directed towards understanding the links between climate change and global phytoplankton communities while also highlighting the existing regional biases in research.
2. Materials and Methods
Data Sources, Search Criteria, Analyses
Publications reporting primary research results spanning the years 2012–2022 were considered. They were sampled from Web of Science (WoS), a database of credible academic publications [52]. WoS is widely used as an authoritative data source for assessing trends in global research activities [39,53,54,55,56]. The sampling approach in this report is therefore consistent with the practice in the scientific community. Research papers were obtained using the following search terms on WoS: “climate change”, “phytoplankton”, “chlorophyll-a concentration”, “growth rate”, and “dry weight”. These search terms were selected based on the authors’ own research interests and on the basis of their broad relevance to phytoplankton biomass (chlorophyll-a concentration), species fitness (growth rate), and carbon content (dry weight).
An initial search in Web of Science generated 2204 articles, which were reduced to 716 after omitting duplicates and papers beyond the scope of the search criteria. Merging of the records and deletion of duplicates were carried out in the R console (Version 4.2.2). The publications were thereafter categorised based on study location (countries), ecosystem type (freshwater, brackish, or marine), and climate region (polar, temperate, subtropical, or tropical). Information for each of these categories was gleaned from materials and method sections of the sampled reports. In terms of climate regions, the categories were defined based on the approach used in previous reviews [57,58,59]. Studies were classified as a tropical region when research organisms were collected from areas between 23.4° north and south of the Equator; research involving organisms from areas between 23.5° and 35° north and south of the Equator were placed in the subtropical region. Studies were categorised as temperate when they involved organisms from regions between 36° and 60° north and south of the Equator. Polar studies involved organisms from regions between latitudes 60° and 90° north and south of the Equator.
A two-way ANOVA comparison and a Chi-square analysis of publications investigating the impact of climate change on phytoplankton from different climate regions and ecosystems (marine, fresh, and brackish water) were carried out in the R console. The probability densities for the number of papers focused on ecosystems from different climate zones were estimated using Bayesian modelling, following previous bibliometric investigations [54]. A generalised linear mixed model that provides Bayesian estimates via the Markov chain Monte Carlo (MCMC) method was used to analyse the data on the number of papers [60]. Models for describing the number of papers (NP) on phytoplankton from ecosystems (Eco) of different climate regions (CR) were selected based on the Akaike Information Criterion, AIC (Supplementary Table S3A). The model included negative binomial distribution of errors with prior probabilities at 1, assuming research opportunities were the same for each climate zone and ecosystem type. The binomial model was verified statistically by a significant, positive linear relationship between the variance and the mean of the grouped data (slope = 1 at alpha = 0.05). The probability densities were estimated using stan_glm, a package in the R software (version 4.1.2) for statistical modelling.
3. Results and Discussion
3.1. Under-Representation of Phytoplankton Research in the Tropics
The majority (53.2%) of the publications reported research conducted in the temperate region; this was followed by work performed in polar areas (26.7%) (Figure 1). Tropical and subtropical regions were the least-researched areas; they contributed 11.6% and 8.5% to the total publications, respectively. On average, the number of papers from the temperate, polar, tropical, and subtropical regions was 35, 17, 8, and 6 per year, respectively. A Chi-square analysis using a 4 × 3 design (four climate zones and three aquatic ecosystems) at alpha = 0.05 suggested that the number of publications was contingent on the climate region (X-squared = 258.33, df = 6, p-value < 2.2 × 10−16, range of expected values = 4–200). The contribution of various countries (based on the study area and not authorship) to phytoplankton research is summarised in Figure 2.
The number of publications on different ecosystems in different climate zones was compared using a two-way analysis of variance (ANOVA at alpha < 0.05) for an unequal sample size; the research year and the ecosystem type were the fixed factors in this analysis. The results indicated significant differences in phytoplankton research across different ecosystems, independent of the year (Figure 3; Supplemental Table S2). The bulk (≈65%) of the research was carried out in marine systems; about 26% was conducted in freshwater systems; only about 9% of the efforts were devoted to brackish ecosystems. This trend occurred in all climate regions (Figure 3).
Posterior probability density estimates for the number of papers generally reflected the raw data on the distribution of research among different climate zones and ecosystems (Figure 4). The probability densities were estimated using the Bayesian Monte Carlo simulation with 1000 iterations, assuming research opportunities were the same for different climate zones and aquatic ecosystems (Supplemental Table S3A,B). The present analysis therefore spotlights the incontrovertible regional bias in phytoplankton research related to global climate change in the last decade.
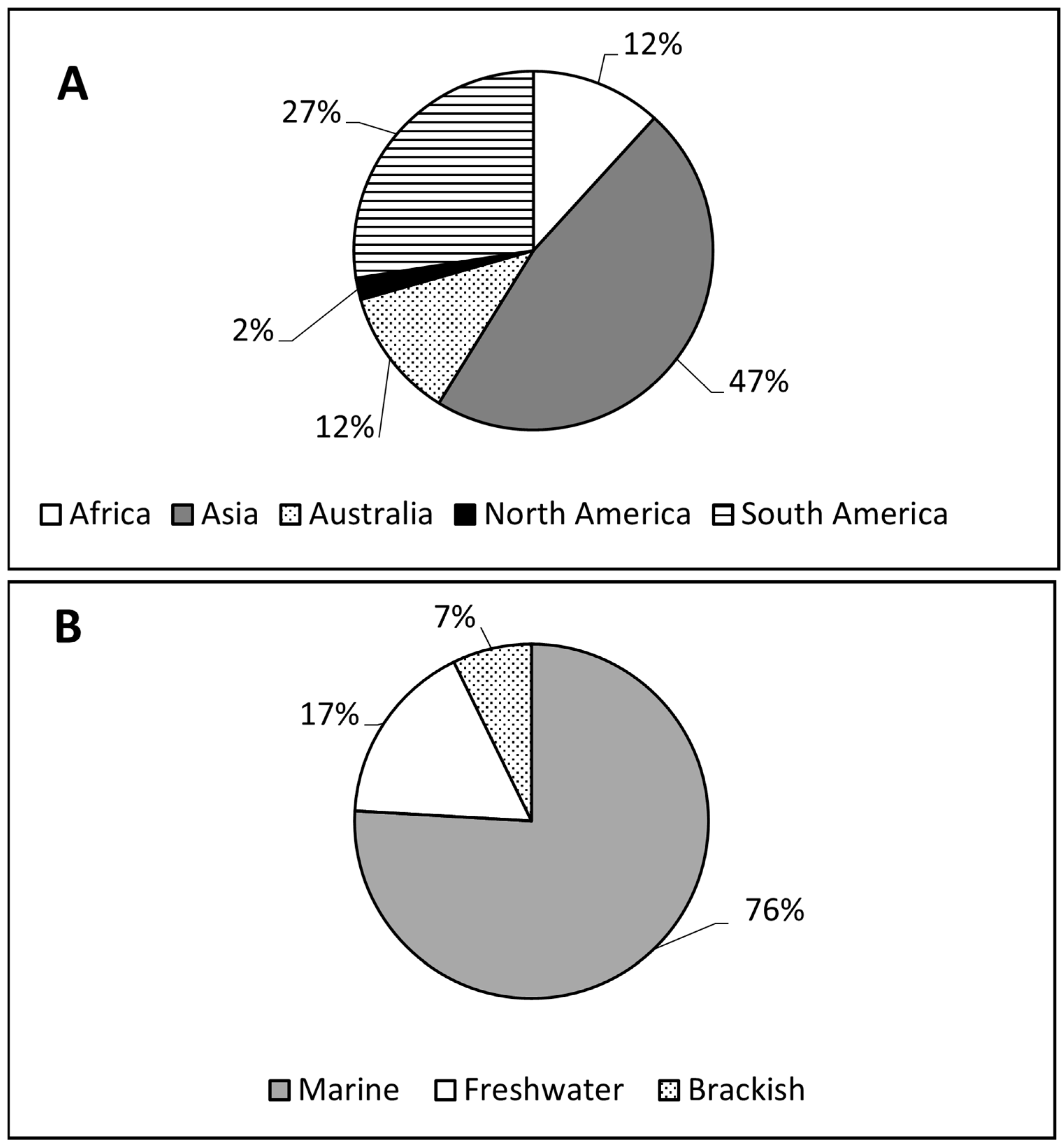
Particularly in the tropics, almost half (47.1%) of the phytoplankton research was from Asia, followed by South America (27.5%); Africa’s contribution was only about 12% (Figure 5). Only four tropical countries—India, Brazil, Australia, and Saudi Arabia—were dominant in phytoplankton research. Together, these countries provided six out of every ten publications from the tropics, consistent with reports on the general state of research in the tropics [61]. Approximately 22, 18, 12, and 10% of all the publications from the tropics were from India, Brazil, Australia, and Saudi Arabia, respectively (Supplemental Table S1).
Approximately 80% of the research effort was focused on temperate and polar regions (Figure 1 and Figure 2), which have large economies capable of supporting knowledge development [62,63]. Europe, North America, and Antarctica alone maintain more than half (54%) of all field stations and laboratories capable of supporting phytoplankton research in the world [15,64]. In contrast, only about 12% of global spending on research and development is undertaken in the tropics [65]. So, it is possible that the current state of phytoplankton research reflects global economic/social structures. Researchers such as [39], who assessed the trends in phytoplankton scientific literature, obtained similar trends, where significantly higher proportions of research publications were undertaken in developed countries, i.e., the USA, Canada, Germany, and France.
The low count of phytoplankton research in the tropics may also be attributed to taxonomic bias related to the choice of study organisms [54,66]. This bias arises when organisms from a particular taxonomic group are disproportionately targeted for research [54]. It occurs in other research areas, for example, on invasive species [67], biodiversity conservation [68], and community assessment [69]. The bias is more prevalent in areas with limited taxonomic expertise and funding opportunities [39,66,70]. Hence, it is possible that a lack of expertise and funding opportunities compel tropical countries to prioritise research efforts in areas such as fisheries, agriculture, and biomedical science, which are directly related to socioeconomic development. Phytoplankton are mostly microscopic and indirectly related to humans through complex trophic relationships and other indicators [71]; hence, they may not attract much attention in countries with limited research opportunities.
The lack of investment in phytoplankton research across the tropics is worrying on several levels. It is one of the major sources of uncertainty in Earth system models for projecting global primary production under climate change [72]. Currently, the criteria for assessing global primary production rely heavily on extrapolations from temperate data [38,72,73]. This surrogate approach introduces significant (≈20%) uncertainty in model predictions [72]; it underestimates the growth of pico-eukaryotic phytoplankton adapted to oligotrophic waters in tropical and subtropical areas [38,73]. This problem exists because, in terms of phytoplankton diversity and other essential variables, tropical communities are largely undescribed [74]. Hence, efforts should be made to increase phytoplankton research in the tropics to help improve future climate projections [73]. It is also likely that the low level of phytoplankton research in the tropics limits our ability to identify community- and ecosystem-level processes that mediate the risk of climate change and other human-induced impacts. Sea surface warming and nutrient pollution, for example, may enhance primary production, thus shifting algal communities toward bloom-forming, toxin-producing species [75]. Particularly in the tropics, such blooms of phytoplankton, even at moderate levels, lead to the redistribution of heat in surface waters, with potential effects on weather conditions (drought, flooding, etc.) directly related to the health of many communities [76]. Toxins from the bloom may also accumulate in the food web, resulting in massive fish kills and other hazards [77]. Research on harmful algal blooms (HABs) is dominated by Europe and North America [78]. Data from these regions alone may not address the risk of HABs around the world. Hence, research to (i) establish how climate change enhances toxin production by toxic HAB species, (ii) evaluate genetic controls on toxin production among HAB species, and (iii) provide long-term (seasonal-to-decadal scale) datasets to help evaluate the proliferation of HABs in all climate regions of the world is needed [44]. Currently, the majority (76%) of the phytoplankton research effort is focused on marine ecosystems. There is a need to extend this effort into freshwater and brackish ecosystems, particularly as marine resources are becoming increasingly challenged by environmental problems (effluent discharge and pollution) embedded in freshwater and brackish ecosystems [79,80].
3.2. Addressing the Under-Representation of Phytoplankton Research in the Tropics
The results presented in this report call for a fundamental reassessment of the framework for phytoplankton research in the tropics. Currently, only about 12% of the global research on phytoplankton is focused on species in tropical ecosystems (Figure 1). Yet, there are many compelling reasons for increased phytoplankton research in the tropics. The common one is that tropical areas are biodiversity powerhouses populated by species with limited tolerance to the impacts of global climate change (warming, acidification, altered nutrient concentrations, etc.) [9,81,82,83,84]. Another reason is that, in terms of organic matter production and carbon cycling, tropical aquatic ecosystems are generally more productive than those in other climate zones [85,86,87]. So, in order to better understand the magnitude of the ongoing global climate change problem, deliberate efforts to drive investment in aquatic tropical ecosystems and organisms are needed. The United Nations and other authorities have identified manpower development, technical infrastructure, and funding support as key areas that require sustained investment in global ocean science. We focus on these areas and suggest practical strategies to enhance phytoplankton research in the tropics.
Manpower development: This strategy should aim at developing skilled phycologists and research support groups through formal education and training. It should involve graduate-level programmes to produce professional phycologists as well as the specialised vocational training of plankton research technicians and citizen scientists [88]. These can include funding programmes aimed at providing scholarships for Ph.D./M.S. degree programs in phytoplankton research as well as targeted short-term capacity-building training. International development aid agencies can be tapped to provide the financial resources that will increase the capacities of developing countries to undertake research. For instance, in the last decade, the World Bank has invested more than USD 500 million in increasing the capacity of regional centres of excellence in Africa to undertake specialised research, particularly in science-related disciplines such as plant breeding, infectious diseases, and coastal resilience [89]. Collectively, the initiative has trained thousands of graduate students, equipping them with the skills to transform the continent. However, critics of the programme worry that the continued intervention from such programmes may be harmful in the long run, especially when governments do not make substantive national investments in research [89]. Another challenge associated with these interventions is the possibility of brain drain that may occur when local institutions do not have the capacity to retain the highly skilled researchers that these centres produce [90]. In this case, these countries stand to lose out even further because momentary paybacks cannot be compared with service paybacks. One way to remedy the situation is for governments within the tropics to increase their capacities to develop world-class institutions that can accommodate and retain the skilled workforce after their respective training.
The majority of the countries located in tropical climates have economic challenges, with limited funding for education [65]. Hence, it may be difficult for such countries to provide students with field-based experience in phytoplankton biology. Hence, help from Western centres of excellence will be needed for the training of phytoplankton ecologists in the tropics. This can be achieved, for example, through international partnership programmes such as those offered by the Nippon Foundation Partnerships for Observation of the Global Ocean (NF-POGO) shipboard training programmes; https://pogo-ocean.org/capacity-development/shipboard-training/ (accessed on 27 October 2023).
Technical infrastructure: Physical facilities for phytoplankton surveys, fieldwork, and experimental studies are similar to those needed in other areas of plankton research. They include research vessels for fieldwork, wet laboratories for culture experiments, chemical laboratories for sample analysis, and facilities for the acquisition and analysis of remote sensing data. Research vessels are expensive and costly to maintain. So, for sampling in deep, oceanic waters, tropical countries may rely on international assistance and industry collaboration to access field stations. One successful model of international assistance is provided by the Norwegian Agency for Development Cooperation (NORAD) through the UN’s Food and Agriculture Organisation (FAO) and the Institute of Marine Research of Bergen, Norway; https://www.fao.org/in-action/eaf-nansen/en (accessed on 15 February 2023). Shipping and offshore oil and gas industries may also support transport to offshore sampling stations along their route of operation. For example, British Petroleum has funded plankton research across the Great Australian Bight in southern Australia, a region of developing oil and gas interests; Nexen, an oil and gas exploration company, also provides travel support for plankton research on routes passing close to their drilling platforms in the North Atlantic [91]. In contrast, phytoplankton experiments and sampling from nearshore, coastal areas, and inland waters could be carried out using small vessels and canoes provided by local fishers, in academia–industry collaborations. Such collaborations have proven useful in plankton studies in Africa, e.g., [13,92]. They can be utilised to increase phytoplankton fieldwork and experimental studies in resource-limited countries that do not have research vessels.
Another strategy that can be used to increase phytoplankton field studies in tropical countries is citizen science, i.e., science undertaken by volunteers from the general public under the direction of professional scientists [88]. Such volunteers are useful for studies aiming to understand processes occurring at broad geographic scales and on private properties (e.g., aquaculture farms), which are impossible to sample extensively with traditional field research models. They can be trained, for example, to collect and preserve samples to help expand the reach of long-term monitoring programmes in marine and freshwater systems [93,94].
The cost of importing scientific equipment is another factor limiting scientific research in low-income countries [95]. Due to a lack of funds, equipment is imported without essential follow-up procedures to maintain and repair it. As a consequence, research facilities in low-income countries have become “graveyards” of equipment that require minimal repairs [96]. So, for research equipment, developing countries may need a model slightly different from the approach used by Western research institutions. For fieldwork, a good number of the needed equipment—including plankton net and water samplers such as Nansen bottles—is designed to be simple to use and make. So, countries can rely on local manufacturing and science workshops to construct such low-technology equipment whilst respecting patent and trademark arrangements on the equipment. High-tech equipment, on the other hand, uses advanced features that may be difficult to reproduce without compromising the quality of research outcomes. For such equipment, two strategies are advised. One is that countries can use alternative, low-cost methods to achieve results comparable to those produced by high-tech equipment. For example, an ultraviolet light-emitting diode can be used in place of an autoclave machine for sterilising culture media and other laboratory supplies [97] and manual microscopy methods (Sedgewick–Rafter counting chamber) can be used to count phytoplankton cells in the absence of a flow cytometer [98]. The second strategy is that countries can rely on international partners for high-tech laboratory supplies and consumables. It is advised that such collaborations should be arranged carefully to ensure equal publishing opportunities for all participating institutions and persons [61,99].
Financial support—The majority of the funding for research and development in most tropical countries is currently provided by international organisations such as the UN and the World Bank [89]. Such funding arrangements are provided based on the country’s development agenda and the interests of donor institutions. Therefore, funding may be secured for such programmes if phytoplankton research is included in national policy priorities, for example, on fisheries management and nature conservation. This may require educating governments and citizens to appreciate the importance of phytoplankton because the general public has limited knowledge of aquatic ecosystems and communities [100]. Funding by national governments and international donors should be supplemented by international competitive grants, which mostly fund centres of excellence [101]. For phytoplankton research, the present analysis suggests that the majority of the centres of excellence are likely found in Europe and North America (Figure 2). So tropical countries are likely to secure grants for phytoplankton research if they co-develop proposals with research centres in Europe and North America.
The challenges derailing research within the tropics, particularly in Africa, are complex and multifaceted and require concerted global efforts to remedy them. The under-representation of the tropics is not limited to only phytoplankton research; it occurs in nearly all fields of research in the tropics [54,66,102,103,104,105]. There is therefore an urgent need for collaborative efforts at national, regional, and international levels to increase high-quality research production within the tropics. We suggest a strategy that seeks to improve phytoplankton research in the tropics through targeted training of talented scientists, international collaboration, and funding mechanisms specifically engineered to promote phytoplankton fieldwork and experiments in the tropics [104]. This calls for the strengthening of legal frameworks within the tropics to strengthen their institutions to undertake research and protect their intellectual properties. Lastly, the formulation of sound policy and institutional frameworks will enable countries in the tropics to set research priorities that will drive their developmental agendas, thereby transforming their societies.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w15244288/s1, Table S1: Studies sampled from the Web of Science, Table S2: Two-way ANOVA comparison of publications investigating the impact of climate change on phytoplankton from different ecosystems (marine, fresh- and brackish water) from 2012–2022, Table S3A: Bayesian model selection testing the impact of the interaction between climate region (CR) and sampling year (Yr) on the number of publications (NP) on phytoplankton from different ecosystem, Table S3B: Parameter coefficient estimates from the Bayesian model on the number of publications by ecosystem.
Author Contributions
The study was conceptualised and executed by R.A., P.K.M. and E.A. Data collection was performed by R.A. Statistical analyses were undertaken by E.A. and R.A. The manuscript was revised critically for intellectual content by P.K.M. and E.A. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the World Bank through the African Centre of Excellence in Coastal Resilience (ACECoR) (Credit No: 6389-GH). Acheampong E was supported by the United Kingdom Research and Innovation (UKRI) Global Challenges Research Fund (GCRF) through the One Ocean Hub Project (Grant number: NE/S008950/1).
Data Availability Statement
The authors confirm that the data that support the findings of this study are available in the Supplementary Materials of this article.
Acknowledgments
This paper is part of a Ph.D. thesis under the Africa Centre of Excellence in Coastal Resilience (ACECoR), the University of Cape Coast, with the support of the World Bank and the Government of Ghana. The authors are grateful for the support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bashir, I.; Lone, F.A.; Bhat, R.A.; Mir, S.A.; Dar, Z.A.; Dar, S.A. Concerns and Threats of Contamination on Aquatic Ecosystems. In Bioremediation and Biotechnology; Hakeem, K.R., Bhat, R.A., Qadri, H., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–26. ISBN 978-3-030-35690-3. [Google Scholar]
- Grizzetti, B.; Lanzanova, D.; Liquete, C.; Reynaud, A.; Cardoso, A.C. Assessing water ecosystem services for water resource management. Environ. Sci. Policy 2016, 61, 194–203. [Google Scholar] [CrossRef]
- Chapman, P.M. Assessing and managing stressors in a changing marine environment. Mar. Pollut. Bull. 2017, 124, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Ormerod, S.J.; Dobson, M.; Hildrew, A.G.; Townsend, C.R. Multiple stressors in freshwater ecosystems. Freshw. Biol. 2010, 55, 1–4. [Google Scholar] [CrossRef]
- Jones, K.R.; Klein, C.J.; Halpern, B.S.; Venter, O.; Grantham, H.; Kuempel, C.D.; Shumway, N.; Friedlander, A.M.; Possingham, H.P.; Watson, J.E.M. The Location and Protection Status of Earth’s Diminishing Marine Wilderness. Curr. Biol. 2018, 28, 2506–2512.e3. [Google Scholar] [CrossRef] [PubMed]
- Barbier, E.B. Marine ecosystem services. Curr. Biol. 2017, 27, R507–R510. [Google Scholar] [CrossRef]
- Freeman, L.A.; Corbett, D.R.; Fitzgerald, A.M.; Lemley, D.A.; Quigg, A.; Steppe, C.N. Impacts of Urbanization and Development on Estuarine Ecosystems and Water Quality. Estuaries Coasts 2019, 42, 1821–1838. [Google Scholar] [CrossRef]
- WIOMSA/IOC UNESCO. United Nations Ocean Decade for Africa: The Science We Need for the Ocean We Want in Africa; WIOMSA: Zanzibar, Tanzania, 2022. [Google Scholar]
- IPCC. The Ocean and Cryosphere in a Changing Climate: Special Report of the Intergovernmental Panel on Climate Change, 1st ed.; Cambridge University Press: Cambridge, UK, 2022; ISBN 978-1-00-915796-4. [Google Scholar]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2023; p. 2391. [Google Scholar] [CrossRef]
- Emori, S.; Taylor, K.; Hewitson, B.; Zermoglio, F.; Juckes, M.; Lautenschlager, M.; Stockhause, M. CMIP5 Data Provided at the IPCC Data Distribution Centre. Fact Sheet of the Task Group on Data and Scenario Support for Impact and Climate Analysis (TGICA) of the Intergovernmental Panel on Climate Change (IPCC), 8p. 2016. Available online: https://www.ipcc.ch/site/assets/uploads/2020/11/TGICA_Fact_Sheet_CMIP5_data_provided_at_the_IPCC_DDC_Ver_1_2016.pdf (accessed on 30 October 2023).
- Gattuso, J.-P.; Magnan, A.; Billé, R.; Cheung, W.W.L.; Howes, E.L.; Joos, F.; Allemand, D.; Bopp, L.; Cooley, S.R.; Eakin, C.M.; et al. Contrasting futures for ocean and society from different anthropogenic CO2 emissions scenarios. Science 2015, 349, aac4722. [Google Scholar] [CrossRef]
- Hernández Ruiz, L.; Ekumah, B.; Asiedu, D.A.; Albani, G.; Acheampong, E.; Jónasdóttir, S.H.; Koski, M.; Nielsen, T.G. Climate change and oil pollution: A dangerous cocktail for tropical zooplankton. Aquat. Toxicol. 2021, 231, 105718. [Google Scholar] [CrossRef]
- Prakash, S. Impact of Climate Change on Aquatic Ecosystem and Its Biodiversity: An Overview. Int. J. Biol. Innov. 2021, 3, 312–317. [Google Scholar] [CrossRef]
- Wernberg, T.; Smale, D.A.; Thomsen, M.S. A decade of climate change experiments on marine organisms: Procedures, patterns and problems. Glob. Chang. Biol. 2012, 18, 1491–1498. [Google Scholar] [CrossRef]
- Kordas, R.L.; Harley, C.D.G.; O’Connor, M.I. Community ecology in a warming world: The influence of temperature on interspecific interactions in marine systems. J. Exp. Mar. Biol. Ecol. 2011, 400, 218–226. [Google Scholar] [CrossRef]
- Nguyen, K.D.T.; Morley, S.A.; Lai, C.-H.; Clark, M.S.; Tan, K.S.; Bates, A.E.; Peck, L.S. Upper Temperature Limits of Tropical Marine Ectotherms: Global Warming Implications. PLoS ONE 2011, 6, e29340. [Google Scholar] [CrossRef] [PubMed]
- Harley, C.D.G.; Randall Hughes, A.; Hultgren, K.M.; Miner, B.G.; Sorte, C.J.B.; Thornber, C.S.; Rodriguez, L.F.; Tomanek, L.; Williams, S.L. The impacts of climate change in coastal marine systems: Climate change in coastal marine systems. Ecol. Lett. 2006, 9, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Schaum, C.-E.; Buckling, A.; Smirnoff, N.; Studholme, D.J.; Yvon-Durocher, G. Environmental fluctuations accelerate molecular evolution of thermal tolerance in a marine diatom. Nat. Commun. 2018, 9, 1719. [Google Scholar] [CrossRef]
- Dunne, J.P. Fall and rise of the phytoplankton. Nat. Clim. Chang. 2022, 12, 708–709. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Laws, E.A.; Barber, R.T.; Murray, J.W. Phytoplankton and Their Role in Primary, New, and Export Production. In Ocean Biogeochemistry; Fasham, M.J.R., Ed.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 99–121. ISBN 978-3-642-62691-3. [Google Scholar]
- Hedges, J.I. Why Dissolved Organics Matter. In Biogeochemistry of Marine Dissolved Organic Matter; Elsevier: Amsterdam, The Netherlands, 2002; pp. 1–33. ISBN 978-0-12-323841-2. [Google Scholar]
- Nebbioso, A.; Piccolo, A. Molecular characterization of dissolved organic matter (DOM): A critical review. Anal. Bioanal. Chem. 2013, 405, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P. Ocean Science: The power of plankton. Nature 2012, 483, S17–S20. [Google Scholar] [CrossRef]
- Thomas, M.K.; Kremer, C.T.; Klausmeier, C.A.; Litchman, E. A Global Pattern of Thermal Adaptation in Marine Phytoplankton. Science 2012, 338, 1085–1088. [Google Scholar] [CrossRef]
- Kwok, K.W.; Leung, K.M.; Lui, G.S.; Chu, V.K.; Lam, P.K.; Morritt, D.; Maltby, L.; Brock, T.C.; Van den Brink, P.J.; Warne, M.S.J.; et al. Comparison of tropical and temperate freshwater animal species’ acute sensitivities to chemicals: Implications for deriving safe extrapolation factors: Tropical versus Temperate Species Sensitivity. Integr. Environ. Assess. Manag. 2007, 3, 49–67. [Google Scholar] [CrossRef]
- Falkowski, P.G. Evolution of the nitrogen cycle and its influence on the biological sequestration of CO2 in the ocean. Nature 1997, 387, 272–275. [Google Scholar] [CrossRef]
- Kirchman, D.L.; Morán, X.A.G.; Ducklow, H. Microbial growth in the polar oceans—Role of temperature and potential impact of climate change. Nat. Rev. Microbiol. 2009, 7, 451–459. [Google Scholar] [CrossRef]
- Beaugrand, G. Monitoring pelagic ecosystems using plankton indicators. ICES J. Mar. Sci. 2005, 62, 333–338. [Google Scholar] [CrossRef]
- Guinder, V.A.; Popovich, C.A.; Molinero, J.C.; Marcovecchio, J. Phytoplankton summer bloom dynamics in the Bahía Blanca Estuary in relation to changing environmental conditions. Cont. Shelf Res. 2013, 52, 150–158. [Google Scholar] [CrossRef]
- Irwin, A.J.; Finkel, Z.V.; Müller-Karger, F.E.; Troccoli Ghinaglia, L. Phytoplankton adapt to changing ocean environments. Proc. Natl. Acad. Sci. USA 2015, 112, 5762–5766. [Google Scholar] [CrossRef] [PubMed]
- Walter, B.; Peters, J.; van Beusekom, J.E.E.; St. John, M.A. Interactive effects of temperature and light during deep convection: A case study on growth and condition of the diatom Thalassiosira weissflogii. ICES J. Mar. Sci. 2015, 72, 2061–2071. [Google Scholar] [CrossRef]
- Collins, S.; Rost, B.; Rynearson, T.A. Evolutionary potential of marine phytoplankton under ocean acidification. Evol. Appl. 2014, 7, 140–155. [Google Scholar] [CrossRef]
- Feng, Y.; Warner, M.E.; Zhang, Y.; Sun, J.; Fu, F.-X.; Rose, J.M.; Hutchins, D.A. Interactive effects of increased pCO2, temperature and irradiance on the marine coccolithophore Emiliania huxleyi (Prymnesiophyceae). Eur. J. Phycol. 2008, 43, 87–98. [Google Scholar] [CrossRef]
- Murphy, G.E.P.; Romanuk, T.N.; Worm, B. Cascading effects of climate change on plankton community structure. Ecol. Evol. 2020, 10, 2170–2181. [Google Scholar] [CrossRef]
- Adams, S.M.; Greeley, M.S. Ecotoxicological Indicators of Water Quality: Using Multi-response Indicators to Assess the Health of Aquatic Ecosystems. Water Air Soil Pollut. 2000, 123, 103–115. [Google Scholar] [CrossRef]
- Parmar, T.K.; Rawtani, D.; Agrawal, Y.K. Bioindicators: The natural indicator of environmental pollution. Front. Life Sci. 2016, 9, 110–118. [Google Scholar] [CrossRef]
- Van De Waal, D.B.; Litchman, E. Multiple global change stressor effects on phytoplankton nutrient acquisition in a future ocean. Phil. Trans. R. Soc. B 2020, 375, 20190706. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, F.M.; Nabout, J.C.; Bini, L.M. Trends in the scientific literature on phytoplankton. Limnology 2008, 9, 153–158. [Google Scholar] [CrossRef]
- Bonachela, J.A.; Klausmeier, C.A.; Edwards, K.F.; Litchman, E.; Levin, S.A. The role of phytoplankton diversity in the emergent oceanic stoichiometry. J. Plankton Res. 2016, 38, 1021–1035. [Google Scholar] [CrossRef]
- Hülse, D.; Arndt, S.; Wilson, J.D.; Munhoven, G.; Ridgwell, A. Understanding the causes and consequences of past marine carbon cycling variability through models. Earth-Sci. Rev. 2017, 171, 349–382. [Google Scholar] [CrossRef]
- Haeder, D.-P.; Villafane, V.E.; Helbling, E.W. Productivity of aquatic primary producers under global climate change. Photochem. Photobiol. Sci. 2014, 13, 1370–1392. [Google Scholar] [CrossRef] [PubMed]
- Assmy, P.; Smetacek, V. Encyclopedia of Microbiology; Elsevier: Amsterdam, The Netherlands, 2009. [Google Scholar]
- Wells, M.L.; Karlson, B.; Wulff, A.; Kudela, R.; Trick, C.; Asnaghi, V.; Berdalet, E.; Cochlan, W.; Davidson, K.; De Rijcke, M.; et al. Future HAB science: Directions and challenges in a changing climate. Harmful Algae 2020, 91, 101632. [Google Scholar] [CrossRef] [PubMed]
- Abdul, W.O.; Adekoya, E.O.; Ademolu, K.O.; Omoniyi, I.T.; Odulate, D.O.; Akindokun, T.E.; Olajide, A.E. The effects of environmental parameters on zooplankton assemblages in tropical coastal estuary, South-west, Nigeria. Egypt. J. Aquat. Res. 2016, 42, 281–287. [Google Scholar] [CrossRef]
- Bopp, S.K.; Lettieri, T. Gene regulation in the marine diatom Thalassiosira pseudonana upon exposure to polycyclic aromatic hydrocarbons (PAHs). Gene 2007, 396, 293–302. [Google Scholar] [CrossRef]
- Ishida, Y.; Hiragushi, N.; Kitaguchi, H.; Mitsutani, A.; Nagai, S.; Yoshimura, M. A highly CO2-tolerant diatom, Thalassiosira weissflogii H1, enriched from coastal sea, and its fatty acid composition. Fish. Sci. 2000, 66, 655–659. [Google Scholar] [CrossRef]
- Vincent, F.; Bowler, C. Diatoms Are Selective Segregators in Global Ocean Planktonic Communities. mSystems 2020, 5, e00444-19. [Google Scholar] [CrossRef]
- Wu, Y.; Gao, K.; Riebesell, U. CO2-induced seawater acidification affects physiological performance of the marine diatom Phaeodactylum tricornutum. Biogeosciences 2010, 7, 2915–2923. [Google Scholar] [CrossRef]
- Boyce, D.G.; Dowd, M.; Lewis, M.R.; Worm, B. Estimating global chlorophyll changes over the past century. Prog. Oceanogr. 2014, 122, 163–173. [Google Scholar] [CrossRef]
- Lancelot, C.; Muylaert, K. 7.02-trends in estuarine phytoplankton ecology. In Treatise on Estuarine and Coastal Science; Academic Press: Waltham, MA, USA, 2011; pp. 5–15. [Google Scholar]
- Birkle, C.; Pendlebury, D.A.; Schnell, J.; Adams, J. Web of Science as a data source for research on scientific and scholarly activity. Quant. Sci. Stud. 2020, 1, 363–376. [Google Scholar] [CrossRef]
- Conrad, E.; Christie, M.; Fazey, I. Is research keeping up with changes in landscape policy? A review of the literature. J. Environ. Manag. 2011, 92, 2097–2108. [Google Scholar] [CrossRef]
- Culumber, Z.W.; Anaya-Rojas, J.M.; Booker, W.W.; Hooks, A.P.; Lange, E.C.; Pluer, B.; Ramírez-Bullón, N.; Travis, J. Widespread Biases in Ecological and Evolutionary Studies. BioScience 2019, 69, 631–640. [Google Scholar] [CrossRef]
- Felton, A.; Fischer, J.; Lindenmayer, D.B.; Montague-Drake, R.; Lowe, A.R.; Saunders, D.; Felton, A.M.; Steffen, W.; Munro, N.T.; Youngentob, K.; et al. Climate change, conservation and management: An assessment of the peer-reviewed scientific journal literature. Biodivers. Conserv. 2009, 18, 2243–2253. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Y.; Li, X.; Lai, Z.; Tackx, M.; Lek, S. A bibliometric analysis of scientific trends in phytoplankton research. Ann. Limnol.-Int. J. Limnol. 2015, 51, 249–259. [Google Scholar] [CrossRef]
- Clarke, D.A.; York, P.H.; Rasheed, M.A.; Northfield, T.D. Does Biodiversity–Ecosystem Function Literature Neglect Tropical Ecosystems? Trends Ecol. Evol. 2017, 32, 320–323. [Google Scholar] [CrossRef]
- Stroud, J.T.; Feeley, K.J. Neglect of the Tropics Is Widespread in Ecology and Evolution: A Comment on Clarke et al. Trends Ecol. Evol. 2017, 32, 626–628. [Google Scholar] [CrossRef] [PubMed]
- Karagulle, D. World Climate Regions. Available online: https://storymaps.arcgis.com/stories/61a5d4e9494f46c2b520a984b2398f3b (accessed on 10 January 2023).
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: Cambridge, UK, 2006; ISBN 1-139-46093-5. [Google Scholar]
- Edelman, A.; Gelding, A.; Konovalov, E.; McComiskie, R.; Penny, A.; Roberts, N.; Templeman, S.; Trewin, D.; Ziembicki, M.; Trewin, B. State of the Tropics 2014 Report; James Cook University: Townsville, Australia, 2014. [Google Scholar]
- Bloch, C.; Sorensen, M.P. The size of research funding: Trends and implications. Sci. Public Policy 2015, 42, 30–43. [Google Scholar] [CrossRef]
- Titley, M.A.; Snaddon, J.L.; Turner, E.C. Scientific research on animal biodiversity is systematically biased towards vertebrates and temperate regions. PLoS ONE 2017, 12, e0189577. [Google Scholar] [CrossRef]
- Valdés, L. Global Ocean Science Report: The Current Status of Ocean Science around the World; UNESCO: Paris, France, 2017. [Google Scholar]
- State of the Tropics. State of the Tropics 2020 Report; James Cook University: Townsville, Australia, 2020. [Google Scholar]
- Santos, J.W.; Correia, R.A.; Malhado, A.C.M.; Campos-Silva, J.V.; Teles, D.; Jepson, P.; Ladle, R.J. Drivers of taxonomic bias in conservation research: A global analysis of terrestrial mammals. Anim. Conserv. 2020, 23, 679–688. [Google Scholar] [CrossRef]
- Burian, R.M. How the Choice of Experimental Organism Matters: Epistemological Reflections on an Aspect of Biological Practice. J. Hist. Biol. 1993, 26, 351–367. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, M.R.; Burnett, N.J.; Braun, D.C.; Suski, C.D.; Hinch, S.G.; Cooke, S.J.; Kerr, J.T. Taxonomic bias and international biodiversity conservation research. FACETS 2017, 1, 105–113. [Google Scholar] [CrossRef]
- Kovalenko, K.E.; Reavie, E.D. Taxonomic bias in freshwater phytoplankton communities and its effect on environmental assessment metrics. Phycologia 2022, 61, 436–443. [Google Scholar] [CrossRef]
- Tyree, M.A.; Bishop, I.W.; Hawkins, C.P.; Mitchell, R.; Spaulding, S.A. Reduction of taxonomic bias in diatom species data. Limnol. Oceanogr. Methods 2020, 18, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef]
- Kwiatkowski, L.; Bopp, L.; Aumont, O.; Ciais, P.; Cox, P.M.; Laufkötter, C.; Li, Y.; Séférian, R. Emergent constraints on projections of declining primary production in the tropical oceans. Nat. Clim. Chang. 2017, 7, 355–358. [Google Scholar] [CrossRef]
- Flombaum, P.; Wang, W.-L.; Primeau, F.W.; Martiny, A.C. Global picophytoplankton niche partitioning predicts overall positive response to ocean warming. Nat. Geosci. 2020, 13, 116–120. [Google Scholar] [CrossRef]
- Estrada, M.; Delgado, M.; Blasco, D.; Latasa, M.; Cabello, A.M.; Benítez-Barrios, V.; Fraile-Nuez, E.; Mozetič, P.; Vidal, M. Phytoplankton across Tropical and Subtropical Regions of the Atlantic, Indian and Pacific Oceans. PLoS ONE 2016, 11, e0151699. [Google Scholar] [CrossRef]
- Amorim, C.A.; Moura, A. do N. Ecological impacts of freshwater algal blooms on water quality, plankton biodiversity, structure, and ecosystem functioning. Sci. Total Environ. 2021, 758, 143605. [Google Scholar] [CrossRef]
- Timmermann, A.; Jin, F.-F. Phytoplankton influences on tropical climate: Phytoplankton Influences on Tropical Climate. Geophys. Res. Lett. 2002, 29, 19-1–19-4. [Google Scholar] [CrossRef]
- Gorham, T.; Dowling Root, E.; Jia, Y.; Shum, C.K.; Lee, J. Relationship between cyanobacterial bloom impacted drinking water sources and hepatocellular carcinoma incidence rates. Harmful Algae 2020, 95, 101801. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, Q.; Zhang, Y.; Li, J.; Shi, K. Research Trends in the Remote Sensing of Phytoplankton Blooms: Results from Bibliometrics. Remote Sens. 2021, 13, 4414. [Google Scholar] [CrossRef]
- Schlüter, M.; Mcallister, R.R.J.; Arlinghaus, R.; Bunnefeld, N.; Eisenack, K.; Hölker, F.; Milner-Gulland, E.J.; Müller, B.; Nicholson, E.; Quaas, M.; et al. New horizons for managing the environment: A review of coupled social-ecological systems modelling: Social-ecological systems modelling. Nat. Resour. Model. 2012, 25, 219–272. [Google Scholar] [CrossRef]
- Vári, Á.; Podschun, S.A.; Erős, T.; Hein, T.; Pataki, B.; Iojă, I.-C.; Adamescu, C.M.; Gerhardt, A.; Gruber, T.; Dedić, A.; et al. Freshwater systems and ecosystem services: Challenges and chances for cross-fertilization of disciplines. Ambio 2022, 51, 135–151. [Google Scholar] [CrossRef]
- Jacobs, Z.L.; Yool, A.; Jebri, F.; Srokosz, M.; van Gennip, S.; Kelly, S.J.; Roberts, M.; Sauer, W.; Queirós, A.M.; Osuka, K.E.; et al. Key climate change stressors of marine ecosystems along the path of the East African coastal current. Ocean Coast. Manag. 2021, 208, 105627. [Google Scholar] [CrossRef]
- Saatchi, S.; Longo, M.; Xu, L.; Yang, Y.; Abe, H.; André, M.; Aukema, J.E.; Carvalhais, N.; Cadillo-Quiroz, H.; Cerbu, G.A.; et al. Detecting vulnerability of humid tropical forests to multiple stressors. One Earth 2021, 4, 988–1003. [Google Scholar] [CrossRef]
- Frölicher, T.L.; Rodgers, K.B.; Stock, C.A.; Cheung, W.W.L. Sources of uncertainties in 21st century projections of potential ocean ecosystem stressors. Glob. Biogeochem. Cycles 2016, 30, 1224–1243. [Google Scholar] [CrossRef]
- Rodgers, K.B.; Lin, J.; Frölicher, T.L. Emergence of multiple ocean ecosystem drivers in a large ensemble suite with an Earth system model. Biogeosciences 2015, 12, 3301–3320. [Google Scholar] [CrossRef]
- Arnosti, C.; Steen, A.D.; Ziervogel, K.; Ghobrial, S.; Jeffrey, W.H. Latitudinal Gradients in Degradation of Marine Dissolved Organic Carbon. PLoS ONE 2011, 6, e28900. [Google Scholar] [CrossRef]
- Brunskill, G.J. Tropical Margins. In Carbon and Nutrient Fluxes in Continental Margins; Liu, K.-K., Atkinson, L., Quiñones, R., Talaue-McManus, L., Eds.; Global Change–The IGBP Series; Springer: Berlin/Heidelberg, Germany, 2010; pp. 423–493. ISBN 978-3-540-92734-1. [Google Scholar]
- Lønborg, C.; Müller, M.; Butler, E.C.V.; Jiang, S.; Ooi, S.K.; Trinh, D.H.; Wong, P.Y.; Ali, S.M.; Cui, C.; Siong, W.B.; et al. Nutrient cycling in tropical and temperate coastal waters: Is latitude making a difference? Estuar. Coast. Shelf Sci. 2021, 262, 107571. [Google Scholar] [CrossRef]
- Dickinson, J.L.; Zuckerberg, B.; Bonter, D.N. Citizen Science as an Ecological Research Tool: Challenges and Benefits. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 149–172. [Google Scholar] [CrossRef]
- Nordling, L. World Bank pours hundreds of millions into African science. Nature 2018, 561, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Berger, S. Brain Drain, Brain Gain and Its Net Effect; Global Knowledge Partnerships on Migration and Development (KNOMAD): Washington, DC, USA, 2022. [Google Scholar]
- Batten, S.D.; Abu-Alhaija, R.; Chiba, S.; Edwards, M.; Graham, G.; Jyothibabu, R.; Kitchener, J.A.; Koubbi, P.; McQuatters-Gollop, A.; Muxagata, E.; et al. A Global Plankton Diversity Monitoring Program. Front. Mar. Sci. 2019, 6, 321. [Google Scholar] [CrossRef]
- Asiedu, D.A. Copepod Abundance and Impact of Selected Human Induced Stressors on Calanoid Copepod in the Coastal Sea of Ghana. Ph.D. Thesis, University of Cape Coast, Cape Coast, Ghana, 2020. [Google Scholar]
- Castilla, E.P.; Cunha, D.G.F.; Lee, F.W.F.; Loiselle, S.; Ho, K.C.; Hall, C. Quantification of phytoplankton bloom dynamics by citizen scientists in urban and peri-urban environments. Environ. Monit. Assess. 2015, 187, 690. [Google Scholar] [CrossRef]
- Esenkulova, S.; Suchy, K.D.; Pawlowicz, R.; Costa, M.; Pearsall, I.A. Harmful Algae and Oceanographic Conditions in the Strait of Georgia, Canada Based on Citizen Science Monitoring. Front. Mar. Sci. 2021, 8, 725092. [Google Scholar] [CrossRef]
- Vose, P.B.; Cervellini, A. Problems of scientific research in developing countries. IAEA Bull. 1983, 25, 37–40. [Google Scholar]
- Musar, A. Equipment for Science Education Constraints and Opportunities; World Bank: Washington, DC, USA, 1993. [Google Scholar]
- Song, K.; Mohseni, M.; Taghipour, F. Application of ultraviolet light-emitting diodes (UV-LEDs) for water disinfection: A review. Water Res. 2016, 94, 341–349. [Google Scholar] [CrossRef]
- Steinberg, M.K.; First, M.R.; Lemieux, E.J.; Drake, L.A.; Nelson, B.N.; Kulis, D.M.; Anderson, D.M.; Welschmeyer, N.A.; Herring, P.R. Comparison of techniques used to count single-celled viable phytoplankton. J. Appl. Phycol. 2012, 24, 751–758. [Google Scholar] [CrossRef]
- Jones, M.L.; Blunt, P. ‘Twinning’as a method of sustainable institutional capacity building. Public Adm. Dev. 1999, 19, 381–402. [Google Scholar] [CrossRef]
- Clare, B. Knowledge exchange: Plankton education and outreach activities. In Proceedings of the Book of Abstracts; VLIZ Special Publication 60–Flanders Marine Institute: Bruges, Belgium, 2012. [Google Scholar]
- Auranen, O.; Nieminen, M. University research funding and publication performance—An international comparison. Res. Policy 2010, 39, 822–834. [Google Scholar] [CrossRef]
- Deikumah, J.P.; Mcalpine, C.A.; Maron, M. Biogeographical and Taxonomic Biases in Tropical Forest Fragmentation Research: Biases in Forest Fragmentation Research. Conserv. Biol. 2014, 28, 1522–1531. [Google Scholar] [CrossRef] [PubMed]
- Trimble, M.J.; van Aarde, R.J. Geographical and taxonomic biases in research on biodiversity in human-modified landscapes. Ecosphere 2012, 3, art119. [Google Scholar] [CrossRef]
- Whitworth, J.A.; Kokwaro, G.; Kinyanjui, S.; Snewin, V.A.; Tanner, M.; Walport, M.; Sewankambo, N. Strengthening capacity for health research in Africa. Lancet 2008, 372, 1590–1593. [Google Scholar] [CrossRef]
- Winter, M.; Fiedler, W.; Hochachka, W.M.; Koehncke, A.; Meiri, S.; De la Riva, I. Patterns and biases in climate change research on amphibians and reptiles: A systematic review. R. Soc. Open Sci. 2016, 3, 160158. [Google Scholar] [CrossRef]
Figure 1.
Total number of phytoplankton research studies across different climate zones from 2012 to 2022.
Figure 1.
Total number of phytoplankton research studies across different climate zones from 2012 to 2022.
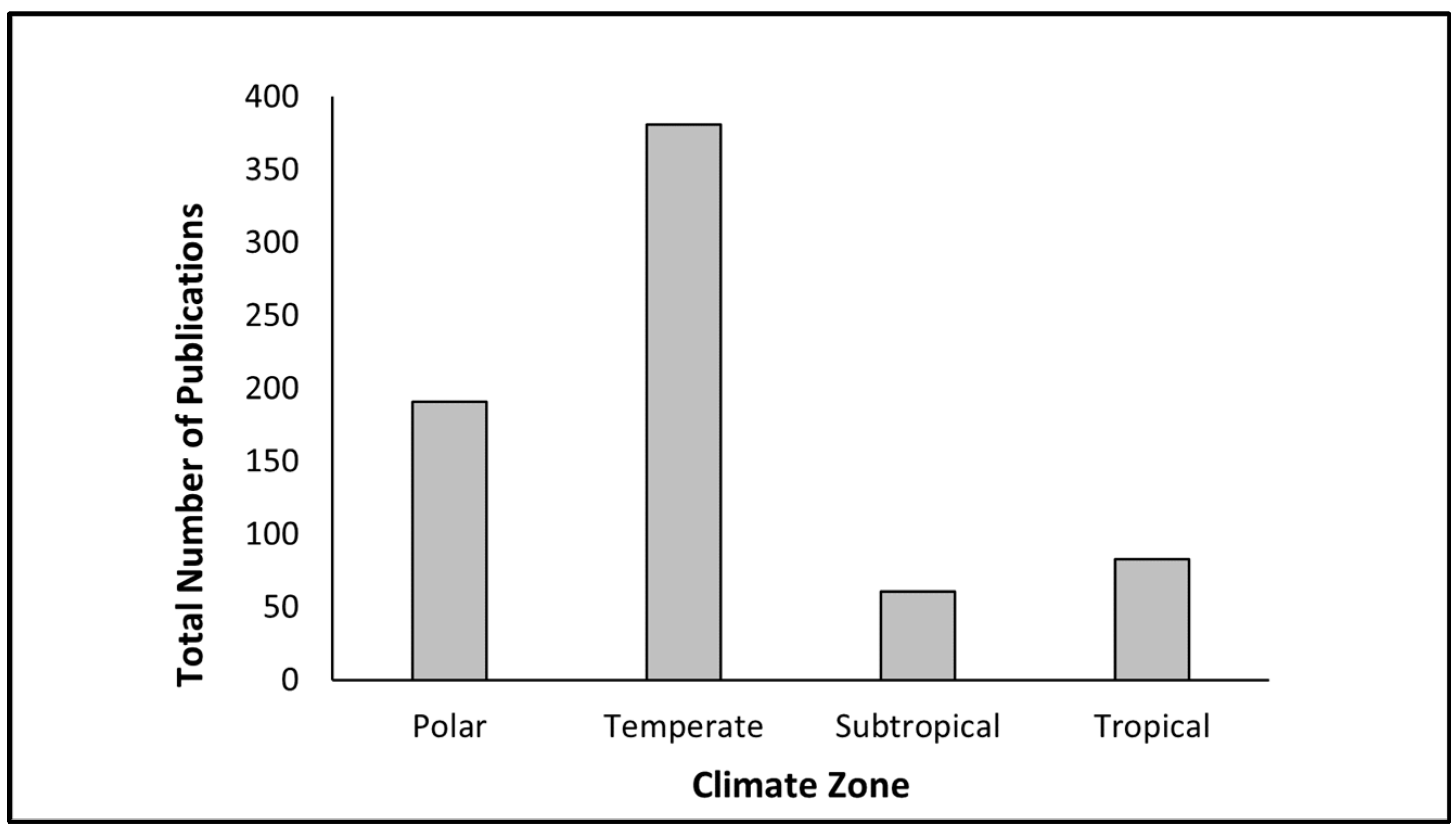
Figure 2.
Global distributions of scientific publications from 2012 to 2022 on phytoplankton. NB. This was based on the study area (and not the country of publication).
Figure 2.
Global distributions of scientific publications from 2012 to 2022 on phytoplankton. NB. This was based on the study area (and not the country of publication).
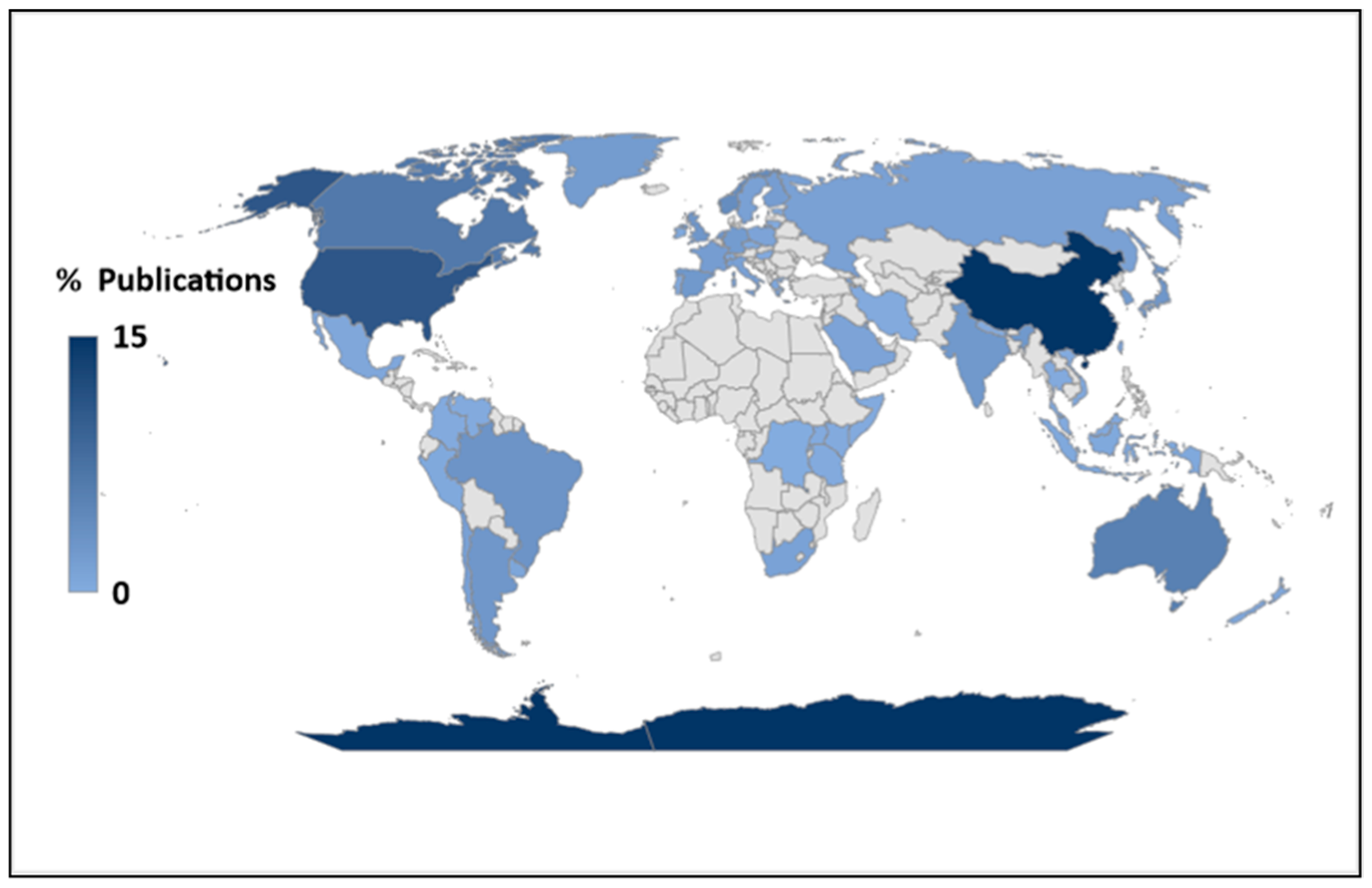
Figure 3.
Total number of scientific reports from 2012 to 2022 on phytoplankton in different aquatic ecosystems from different climate zones.
Figure 3.
Total number of scientific reports from 2012 to 2022 on phytoplankton in different aquatic ecosystems from different climate zones.
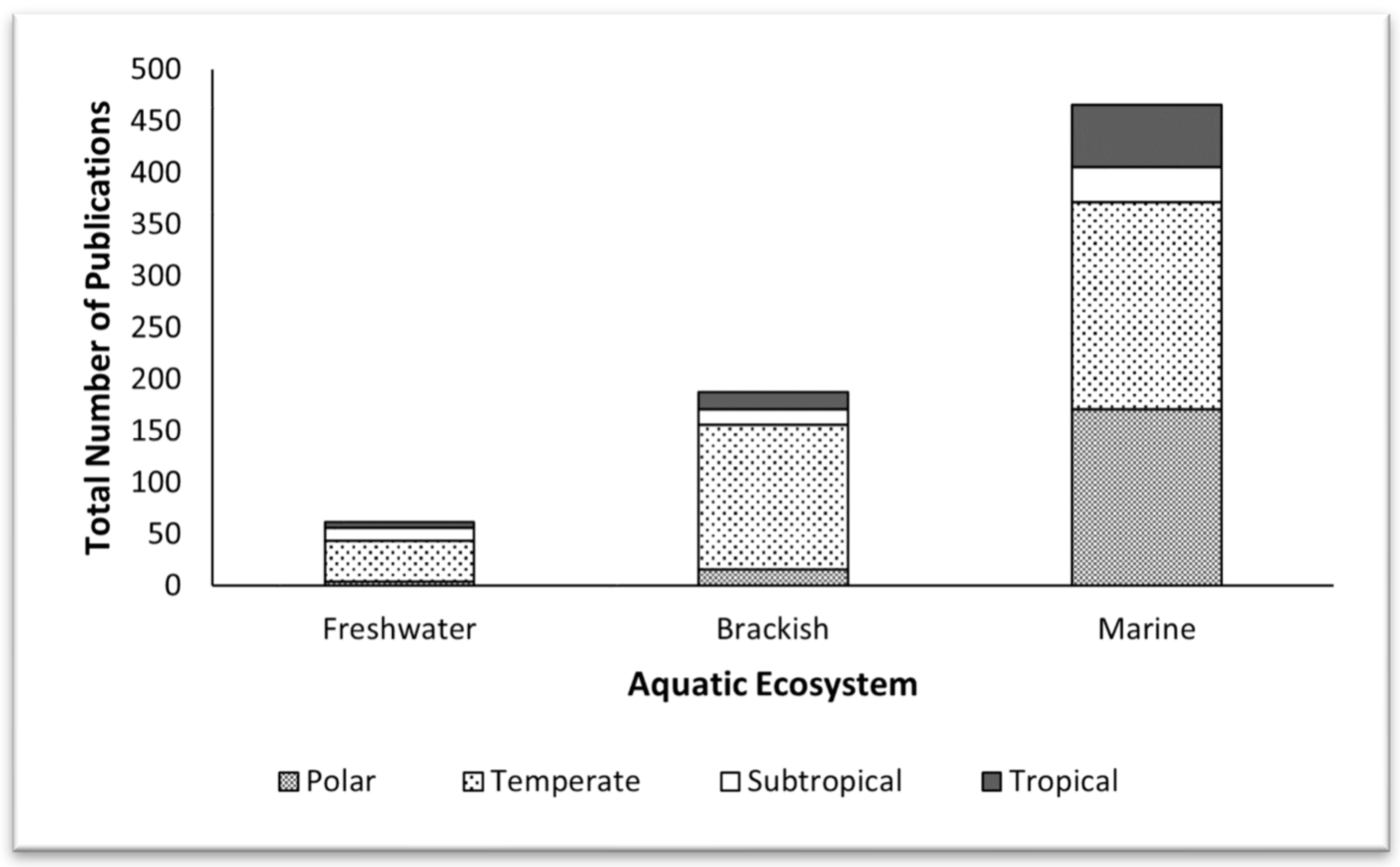
Figure 4.
Posterior probability density estimates for different climate zones and ecosystems.
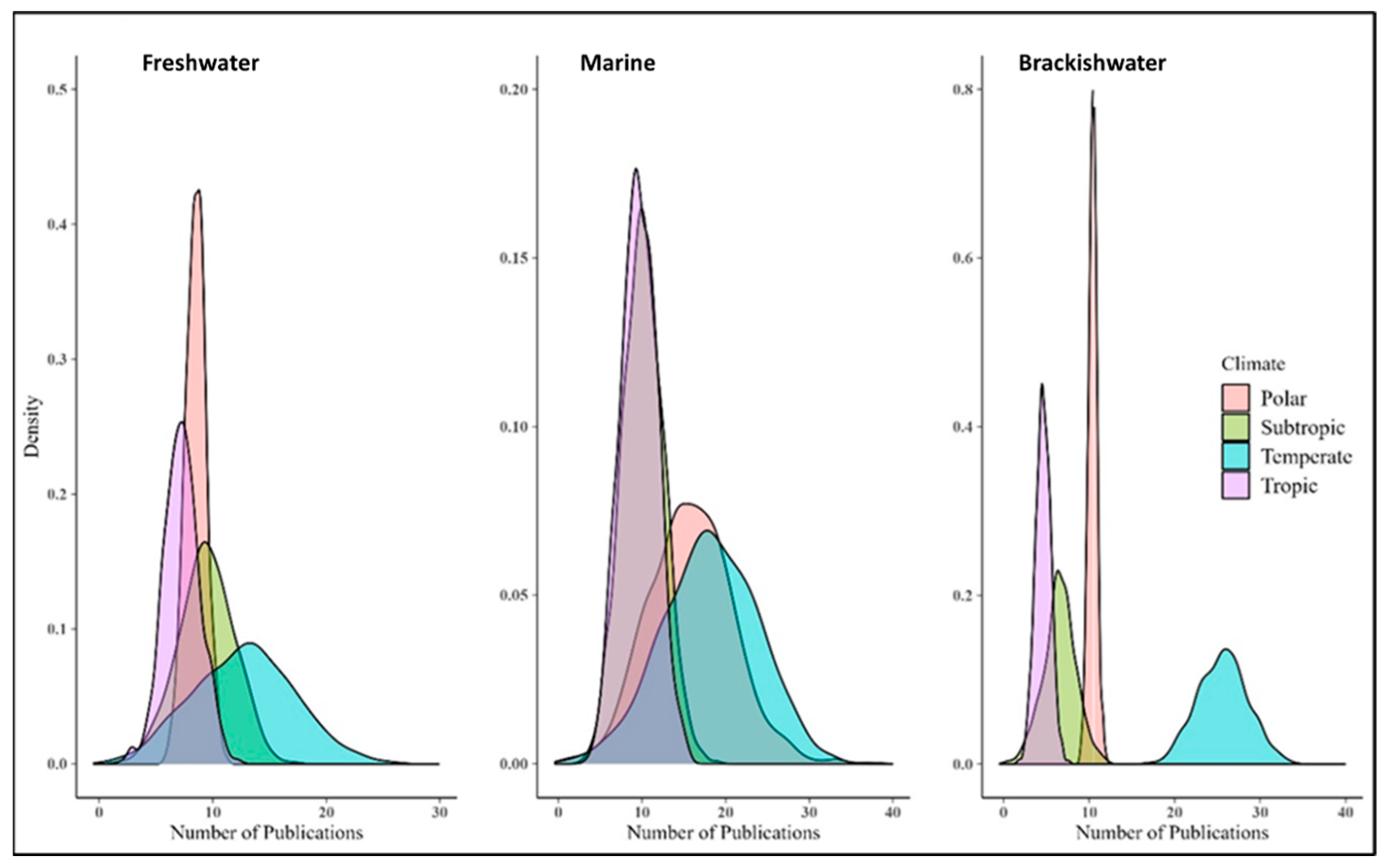
Figure 5.
Total number of scientific publications in tropical ecosystems on different continents (A) and water bodies (B).
Figure 5.
Total number of scientific publications in tropical ecosystems on different continents (A) and water bodies (B).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Adhiambo, R.; Mensah, P.K.; Acheampong, E. Widespread Geographical Disparities in Phytoplankton Ecology Research in the Face of Climate Change: A Review. Water 2023, 15, 4288. https://doi.org/10.3390/w15244288
AMA Style
Adhiambo R, Mensah PK, Acheampong E. Widespread Geographical Disparities in Phytoplankton Ecology Research in the Face of Climate Change: A Review. Water. 2023; 15(24):4288. https://doi.org/10.3390/w15244288
Chicago/Turabian StyleAdhiambo, Rael, Paul Kojo Mensah, and Emmanuel Acheampong. 2023. "Widespread Geographical Disparities in Phytoplankton Ecology Research in the Face of Climate Change: A Review" Water 15, no. 24: 4288. https://doi.org/10.3390/w15244288
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.