1. Introduction
Fruit production across the world relies on inter-specific grafted trees, rather than intra-specific seedlings, which has enabled the expansion of growing areas, including various soil and environmental conditions. Due to comprehensive scion and rootstock breeding, sweet cherry is ranked in sixth place according to total temperate fruit production [
1]. Sweet cherries are cultivated in more than 40 countries worldwide, with the production increases from year to year, due to high consumer demands [
2]. Intensive fruit production is inconceivable without size-controlling rootstocks characterized by high compatibility and productivity [
3]. One rootstock is generally considered as desirable if it raises early bearing, highly productive compact fruit trees suitable for pedestrian orchards. If those planned and selected genetically inherited characteristics are present, rootstock gains prompt popularity among producers and it spreads across the world very rapidly. Marketing and sales pressure of large quantities from nursery production can cause low-adaptive rootstock expansion, regardless of environmental restrictions. In rootstock selection strategies, the focus is on phenotypically (assumed mainly to be genetically inherited) described properties such as good anchorage, good radial and axial hydraulic conductivity, and water use efficiency. Atmospheric impact is often neglected as it is considered that the root system is not so exposed and susceptible to the influence of meteorological conditions, as the scion is [
4]. Previous models developed based on cross-sections of the anatomical parameters of one- and two-year-old roots and rootstock stems of leading cherry rootstocks and selections proved to be reliable for size-controlling capacity prediction within species and interspecific hybrids of the genus
Prunus [
3,
5,
6]. The same autochthonous cherry germplasm has shown annual adjustments towards the seasons. Approaching autumn, the majority of the investigated genotypes increased their vessel numbers and decreased vessel sizes to provide mechanical support, in contrast to spring wood, when the vessel size was greater and was a function of abundant water conductance [
3,
5]. Under dry summer conditions, this is of great importance due to the fact that narrow vessels in the stem are generally more resistant to drought-induced embolism than wide vessels of the same stem [
4,
7,
8,
9]. Climate scenarios for the Balkan region predict an increase in average surface temperatures from 2.2 to 5.1 °C for the period 2080–2100. For the same period, the models predict that precipitation over land might vary between −4% and −27% [
10]. Summer drought in Balkan region is prominent, with climate changes prevailing in the last decade being characterized by unevenly distributed precipitation and high temperatures. Dehydration (long dry periods during summer months) and rehydration (short lasting but heavy rainfalls) cycles during vegetation have become a common phenomenon that limits fruit growing [
3]. Persistent, long dry intervals of up to 2–3 weeks in the period of xylem formation directly influence and leave imprints on its major properties, since division, expansion, and maturation of xylem cells happens over several days to a few weeks [
11,
12]. Short conduit formation span and very variable environmental conditions require high temporal plasticity for the individual plant’s successful water uptake, overall functioning, and survival [
11].
Analyzing the possible autochthonous cherry rootstock candidates’ rapid adaptability of earlywood to overcome the last years’ embolized large vessels by the production of new functional ones, Ljubojević et al. [
3] suggested the potential for cherry cambial and xylem plasticity. Due to proposed xylem plasticity, those genotypes are assumed to be better adapted to drought and embolism caused by freezing/thawing and hydration/dehydration cycles. Those aboveground stem xylem adjustments are generally well known and recognized over the many deciduous and coniferous trees and herbaceous perennial taxa, due to dendro-chronology and herb-chronology investigations [
13,
14,
15,
16], but to the authors’ best knowledge, there has been no such report on root cambial and xylem environmentally driven plasticity [
17], especially in fruit rootstocks. According to Brunner et al. [
17], roots are capable of withstanding drought events due to various morphological and physiological adjustments, but there is still not enough knowledge regarding root anatomical responses to drought, as they are marginally mentioned in reviews and often avoided in research.
Considering results from previous studies on stems, the general aim of this research was to examine whether the cherry root cambium possess the similar water-stress adaptation ability as the scion cambium does. The specific aim of this study was to assess whether there is a shift in root xylem structure due to precipitation fluctuations and temperature increase during the growing season in two contrasting root systems—deep and vertically oriented in P. fruticosa versus shallow in P. cerasus ‘Oblačinska’.
4. Discussion
The root system is generally supported and buffered by the surrounding soil. With such assistance, roots are able to develop large conduits with a lower risk of hydraulic pathway breakage [
4]. However, this soil-mediated protection is diminished if drought events last for a long time and are followed by the heavy rainfalls, especially if the root system is shallow. As the root system is the ‘hidden half’ [
22], it is very difficult to evaluate; even with the modern technologies and approaches, more in-depth analysis across the plant taxa is required. In rootstock breeding, this hidden half is actually the ‘major half’, of the importance for future grafted trees’ performances. The occurrence of the plasticity in stems’ responses directed towards improved water use efficiency during drought events might be of even greater importance in roots, due to their function in water and nutrient uptake. In the presented study, root plasticity varied between species and the levels of structural shifts.
European ground cherry genotypes followed typical semi-ring porous patterns—vessel size decreased as the roots matured, with a very similar number of vessels being formed across the zones. European ground cherry mother trees from which roots were collected received higher amounts of precipitation during May (82.6 mm) and June (60.2 mm), resulting in a more moderate response to the atmospheric conditions compared to Oblačinska sour cherry (
Figure 2).
As for the structural shifts, the first clearly obvious modification was the overall radial growth expressed through the cross-sectional radius and area. It seems that the decrease in root diameter is a trait for increasing the plant acquisition of limited water and productivity under drought [
23]. Annual ring width reduction is also a very common response in branches forming during high temperatures and limited precipitation conditions [
9,
11,
17]. In the present study, monthly precipitation as well as the monthly sum of temperatures in general positively affected radial growth, especially secondary wood—xylem formation. However, the scarce and uneven distribution of precipitations constrained secondary thickening.
Treder and Konopacki [
24] discussed the significance of 2.5 and 5 mm of precipitation thresholds for orchard water balance, emphasizing the complexity of interaction between rainfall and temperature, as well as the amount and intensity of precipitation. It was noted that the 5 mm threshold could be sufficient if the rain is light and continues over a couple of days. In the present study, the number of dry days—PPT
<5mm during May, July, and August were in a statistically significant negative correlation with radial secondary growth—cross-section area, percentage of secondary wood area, and wood/cortex ratio. Investigating xylem plasticity in
Larix decidua and
Picea abies, Bryukhanova and Fonti [
8] described the site climate conditions as dry—summer months with day temperatures from 25 to 35 °C, and little monthly precipitation (45 mm), causing annual xylem adjustments. Other authors also emphasized 40–50 mm of monthly precipitation as a threshold for trees’ successful growth [
25,
26]. Investigating the growth patterns of mangrove trees, Menezes et al. [
27] observed that a lesser tree ring width was closely related to a number of months with less than 50 mm of precipitation.
As for the fruiting trees, the general orchard requirements for successful fruiting and bud formation for the next year are 600–900 mm of precipitation during vegetation [
28]. An ideal distribution of that amount of precipitation should be around 100 mm per month i.e., 25 mm per week. From investigating irrigation requirements for peach orchards, it was concluded that depending on the cultivation system, the monthly needs during June, July, and August varied from 46 to 65 mm, 88–121 mm, and 121–162 mm, respectively [
29]. Considering these observations during the entire growing season, roots from the locality of Dešilovo grew in dry conditions—a lack of precipitation, since monthly precipitation did not exceed 45 mm, whilst roots from the other two localities faced this constraint during July and August. Altogether, with a number of continuous dry days, this has led to such growth stagnation that cross-sections from Dešilovo were one complete wood zone thinner.
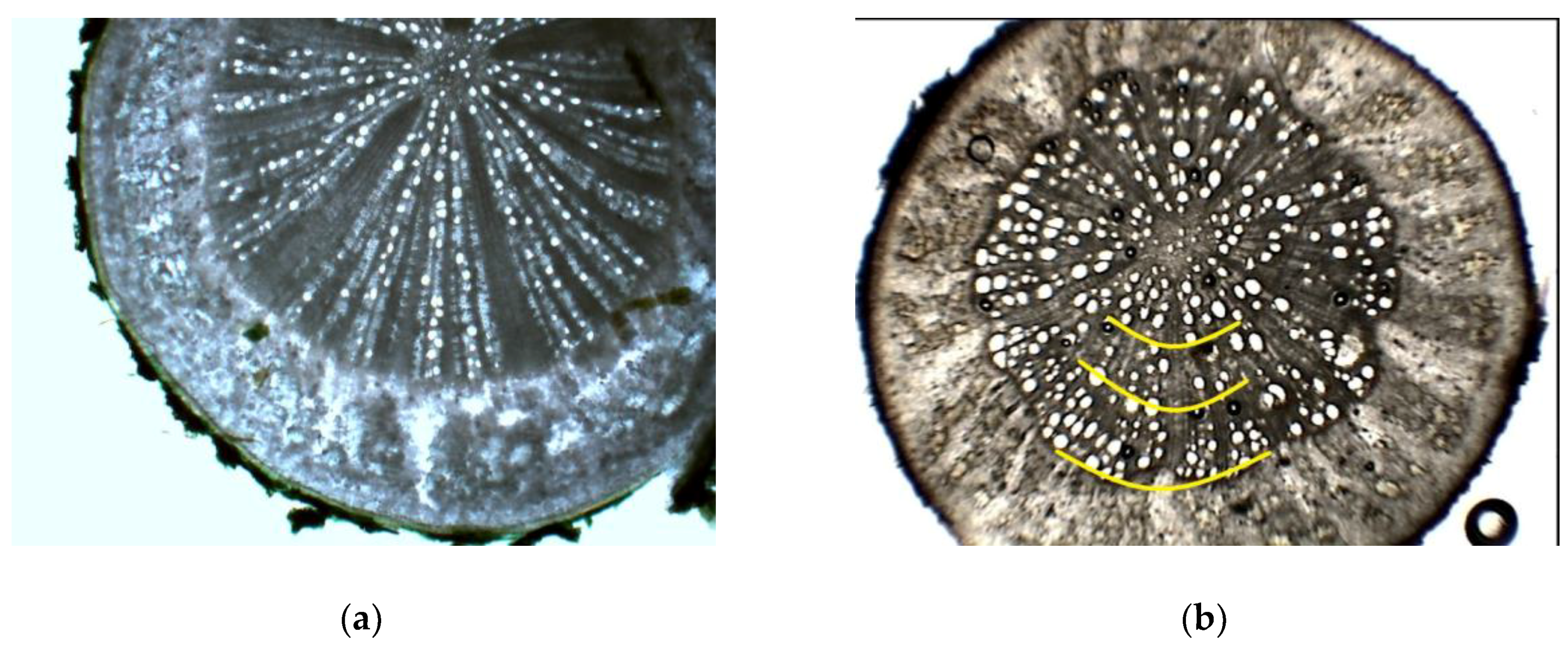
In some root cross-sections, there were traces of very thin middle wood zone (
Figure 2d), while in others it did not appear. These conditionally divided zones for measurement purposes corresponded to false wood rings with shifts in vessel lumen areas from larger to smaller, which was characteristic of semi-ring porous species. The transition from one to another false ring was very noticeable, due to differences in vessel sizes and density, as well as the prominent region of differently colored very dense wood that occurred between rings in most of the investigated genotypes (
Figure 2). Both species did not cease their growth and vessel production prematurely due to drought events late in the summer, as would be expected, but they differed in newly produced xylem conduit sizes. In poplar trees (
Populus nigra L. ×
P. maximowiczii, clone Kamabuchi), Arend and Fromm [
30] also reported a strong seasonally dependent pattern—the development of xylem elements was much more sensitive to drought in early than in late summer. European ground cherry genotypes, characterized by deeper, vertically oriented root systems, produced smaller late summer vessels compared to those from late spring and early summer. On the contrary, roots of the Oblačinska sour cherry shallow root system produced larger late summer vessels compared to those that formed earlier during vegetation. In previous anatomical research conducted on standard cherry rootstocks of different size-controlling capacities, the rootstocks ‘Colt’ and ‘PHL-A’, also characterized by shallow, horizontally oriented root systems [
31], showed the same increase in vessels size as in ‘Oblačinska’, whilst other three did not follow this pattern [
21].
Reviewing the growth dynamics in tropical trees, Worbes [
25] have shown that even regular shoot rings can form two small false rings related to two occasions of precipitation during the dry season, with 50 mm rainfall over one or two days, followed by dry periods of up to three weeks. According to this study, during alternating periods of rainfall and drought, cambial activity started and stopped within short periods, resulting in two small rings, which was the case in the investigated cherry roots. Dry periods with 38 (Fruška gora), 60 (Dešilovo), and 36 (Prokuplje) dry days, resulted in a different number of false growth rings (three or two) imprinted in the analyzed roots. There is an indication of a relation between dry periods–rainy episodes, and the occurrence of intra-annual root xylem rings with the ability for wide conduit production after cambial activity evocation. This seems to be the compensation for reduced radial growth and lesser wood area. It was previously suggested by Ljubojević [
3] that ‘Oblačinska’ sour cherry might have a unique strategy that enables it to supply shoots with a sufficient amount of water and nutrients. Large xylem conduits with a low resistance to water flow and high theoretical hydraulic capacity might compensate for lower water uptake from shallower and lesser-branched root systems. A similar pattern was noted by Singh et al. [
32] on the rice root’s anatomical and histological shifts towards larger xylem vessels under water stress in resistant genotypes. The same strategy was concluded by Von Arx et al. [
13], where upon release from drought stress, long-living perennial herb plants of
Potentilla diversifolia were able to shift the vessel sizes towards larger diameters, enabling them to utilize increased soil moisture.
Overall precipitation and its monthly distribution, as well as the sum of temperatures and monthly average temperatures (except for August), positively correlated with the percentage of the secondary wood area and wood/cortex ratio, whilst it negatively correlated with the percentage of secondary cortex area. On the contrary, due to the obtained correlation matrices, it seems that a number of continuous days with negligible or absent precipitation (PPT<5mm) promoted secondary cortex (phloem) formation. A greater phloem portion under drought stress might be a strategy to amplify the storage capacity for the photoassimilates needed for the growth and development that emerges upon drought release.
The most interesting shift in vessel lumen area due to reduced precipitation in July and August was the production of larger vessels in the investigated ‘Oblačinska’ sour cherry roots as they matured. According to Eilmann et al. [
33], the increased lumen diameter mitigated the reduction in the conducting area caused by drought, and reduced tree-ring widths due to the fourth power relation to the conduit diameter (Hagen Poiseuille’s law according to [
7,
21,
34,
35,
36]). Drought induced larger vessel lumen diameters, and consequently higher specific hydraulic conductivity in ponderosa pine growing at a semi-arid site compared to a moderate mountain site, were previously reported by Maherali and DeLucia [
37]. Very specific statistically significant positive correlations occurred between the average vessel number on complete cross-section and PPT
<5mm in May, June, and July. This suggests that the cherry root maintenance strategy is to mitigate the reduced wood area during drought events, and vice versa; when wood growth is promoted, vessel number was reduced.
Although changing environmental conditions did trigger the shift of large and small vessels, as well as the occurrence of false rings, it seems that their size was mainly genetically controlled. The scarcity of statistically significant correlations implies that there is an inherited mechanism that keeps vessel size under internal control.
5. Conclusions
The assumed impact of atmospheric conditions on cherry root cambial plasticity in this study was fortified by the occurrence of false rings and alterations in conduit sizes in secondary roots collected from in vivo mother plants, and their absence in clonally propagated roots of the same age and genotypes. Under atmospheric impacts, both of the investigated species altered their radial root growth imprinting stops and starts in a cambial activity that resulted in the occurrence of intra-annual false rings. Along with the false rings, European ground cherry followed a usual early–late wood pattern, while Oblačinska sour cherry produced larger vessels as an alternative to smaller cross-sections, and a weaker shallow root system. Although this represents its valuable ability to actively respond to temporal environmental challenges, priority should be narrowed to the Oblačinska sour cherry genotypes with moderate vessel lumen areas, due to the profound vulnerability of large conduits to embolism/cavitation. Changing atmospheric conditions triggered the shifts of large and small vessels throughout the false rings, but their size seems to be mainly genetically controlled. Taking into consideration all the above, genotypes with moderate vessel lumen areas—less than or around 1200 μm2 in the inner zone, as well as no greater than 1500 μm2 in the outer zone—are presumed to be both size-controlling and stable upon the drought events. Thus, further field trials will be focused on the SV2 European ground cherry genotype, and the OV13, OV32, and OV34 Oblačinska sour cherry genotypes.
With the changing environment, rootstock breeding selection goals are to be changed accordingly. The preferred rootstocks should be able to cope with an uneven distribution of precipitation and increased temperatures during the late spring and summer months. For future breeding purposes and for the development of size-controlling prediction models, equal importance should be given to the determination of environmentally related xylem plasticity, as well as genetically controlled conduit size. Investigations of the cherry genepool indicated its ability to optimize its intra-annual anatomical architecture to the specific environment, adjusting both secondary wood and secondary cortex characteristics, which should be further investigated inter-annually.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
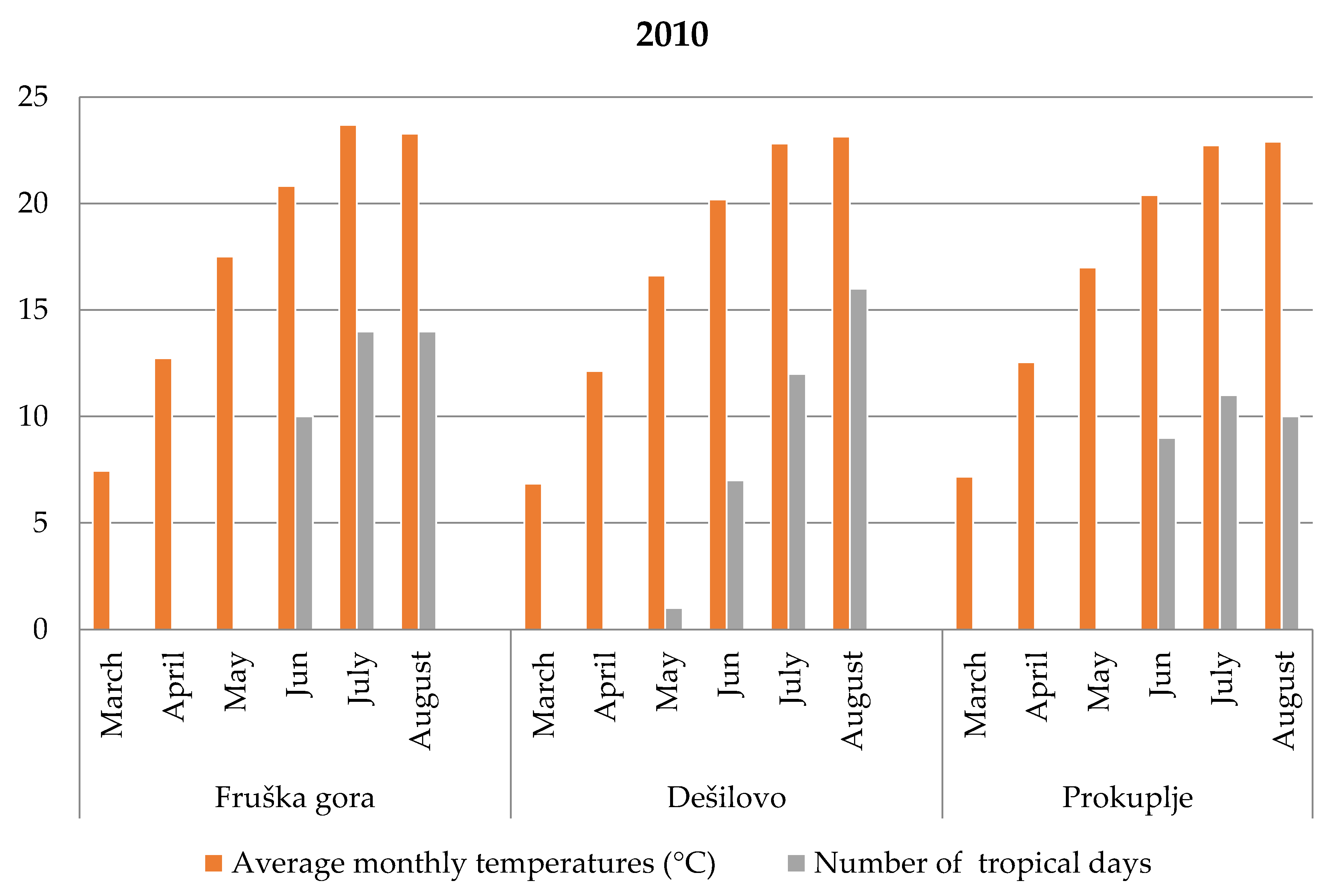{kind=link}