RNAi-Mediated Gene Silencing in a Gonad Organ Culture to Study Sex Determination Mechanisms in Sea Turtle



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Method
2.1. Egg Incubation
2.2. Design of Turtle Specific Sox9 siRNAs
2.3. Gonad Culture and Transfection
2.4. Immunofluorescence
2.5. Quantitative PCR
3. Results and Discussion
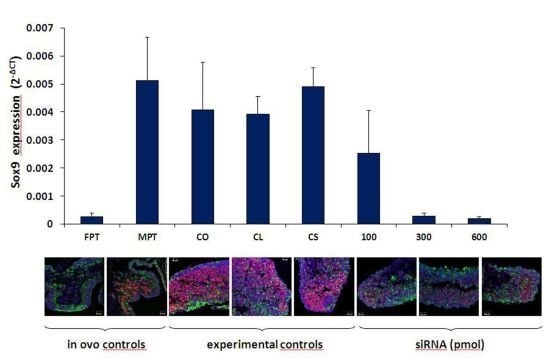
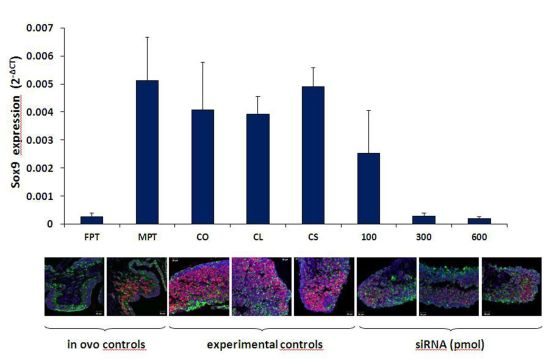
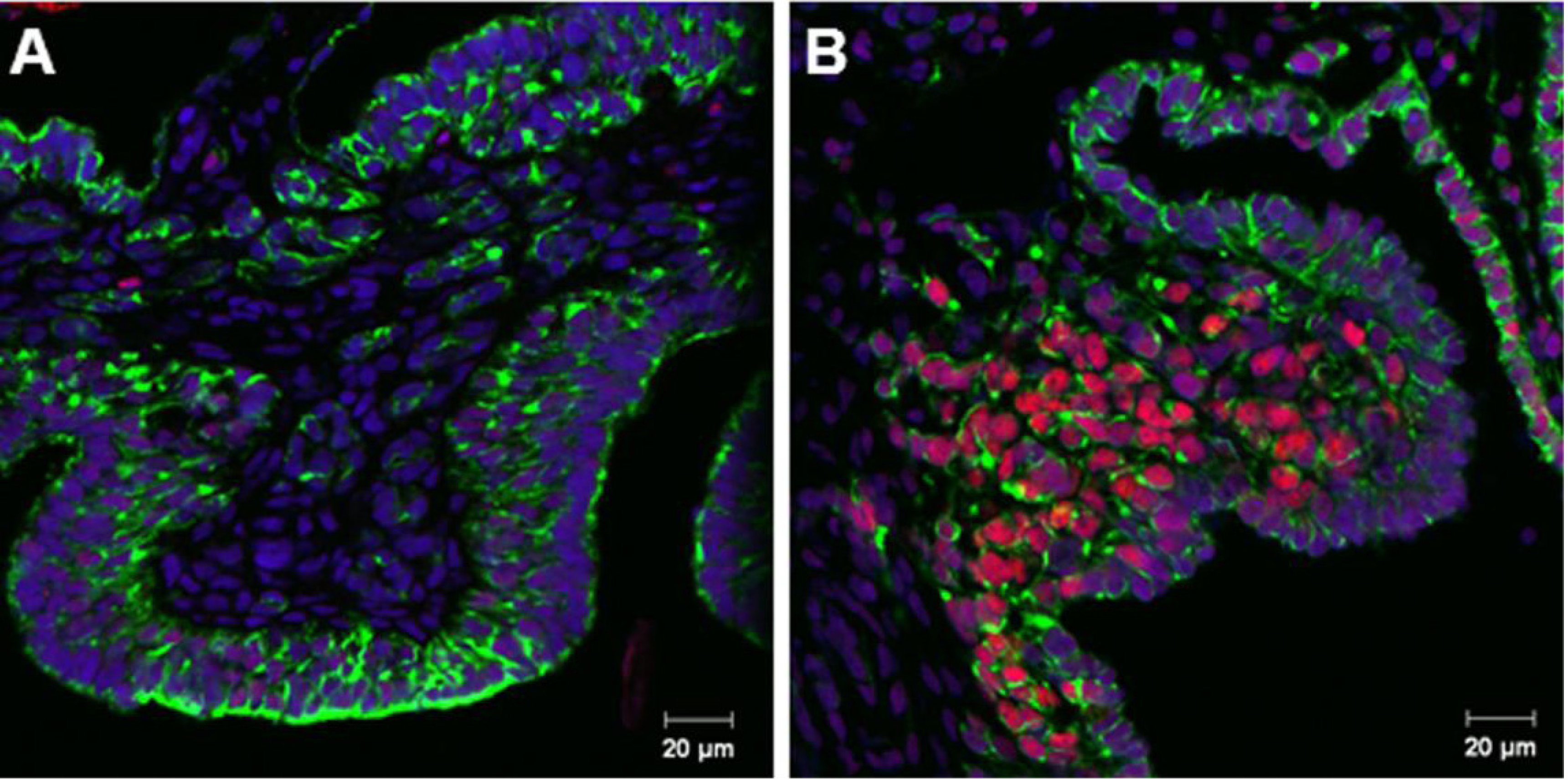
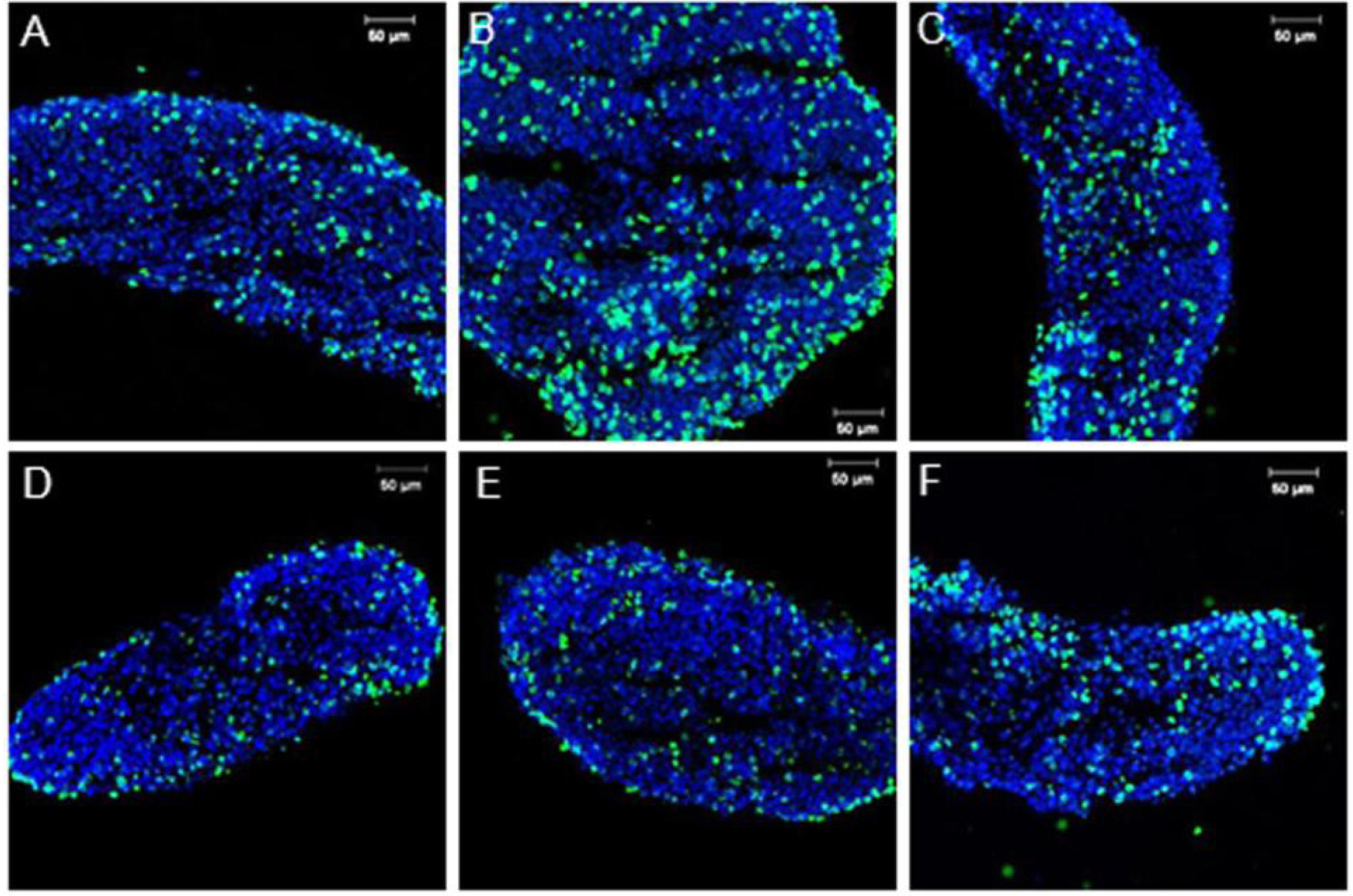
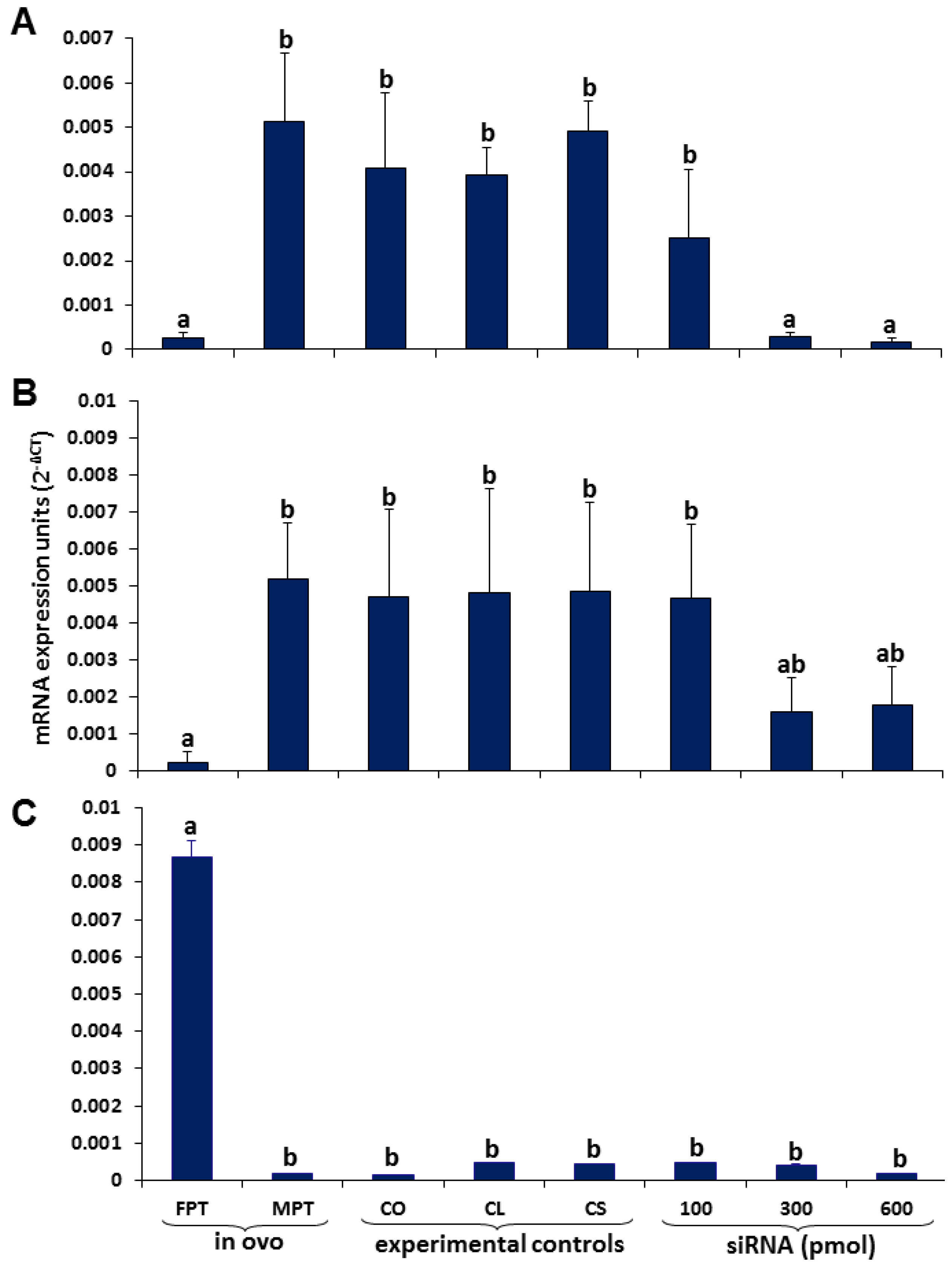
3.1. Sox9 Protein and mRNA Expression
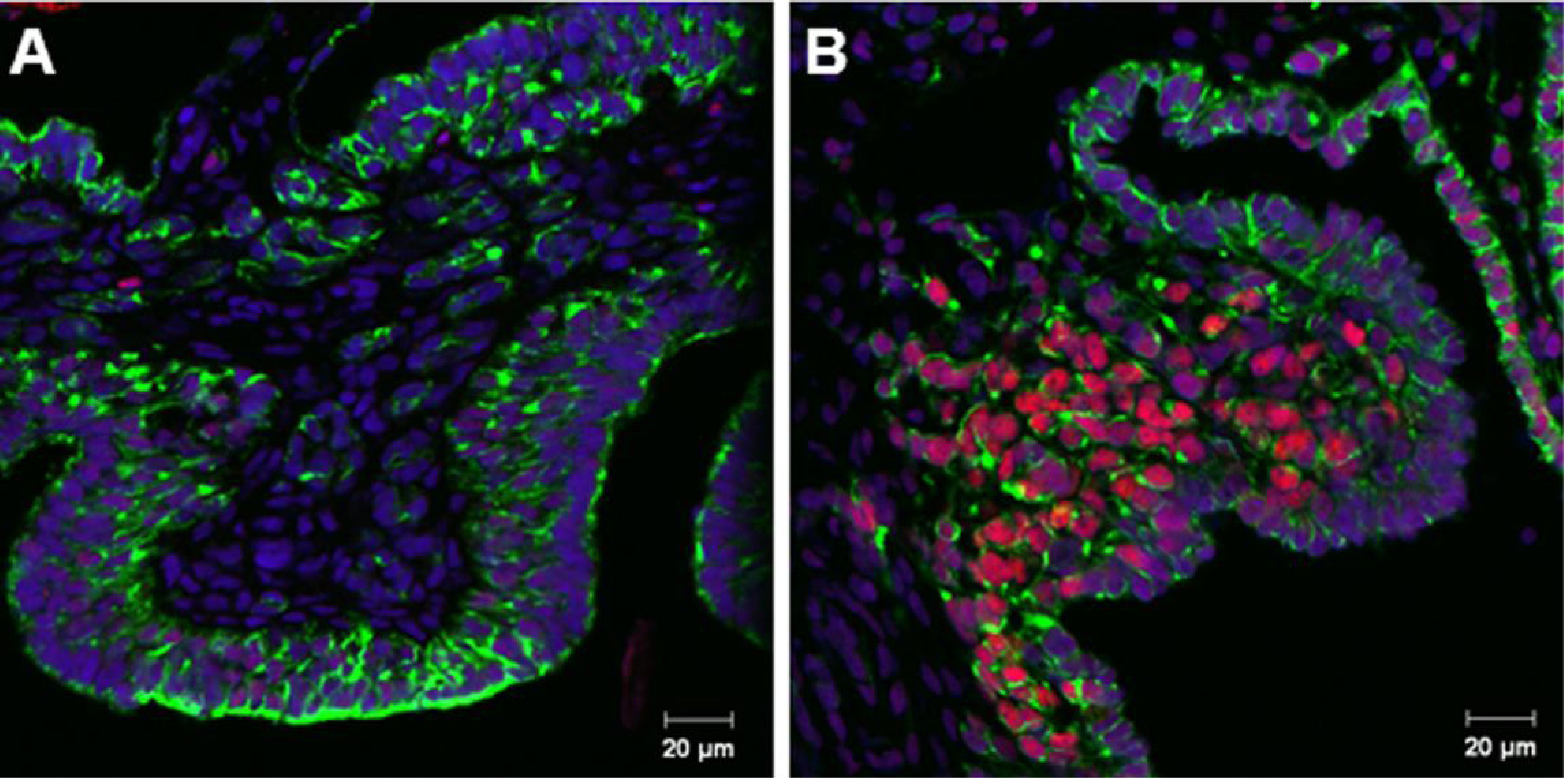
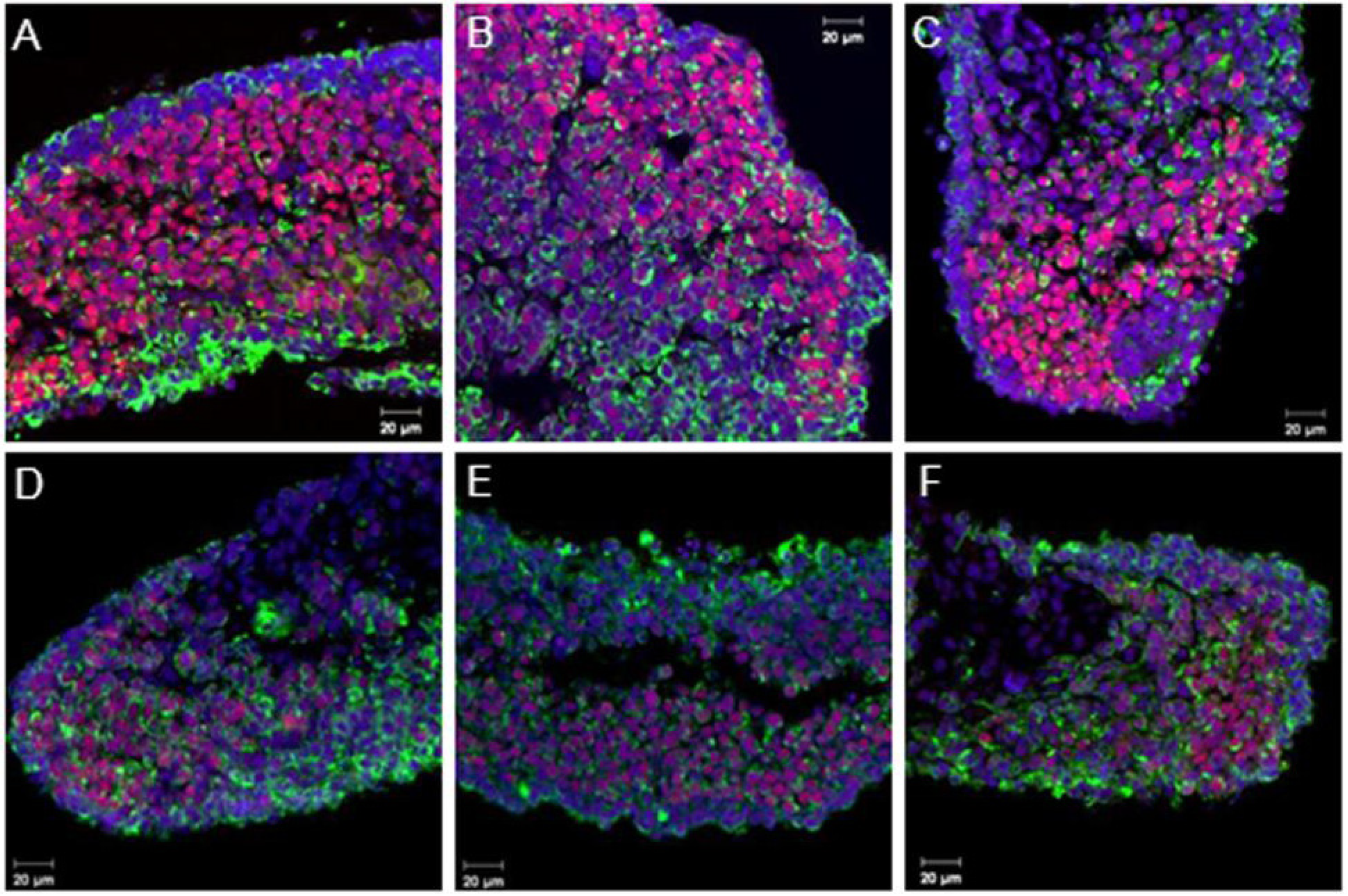
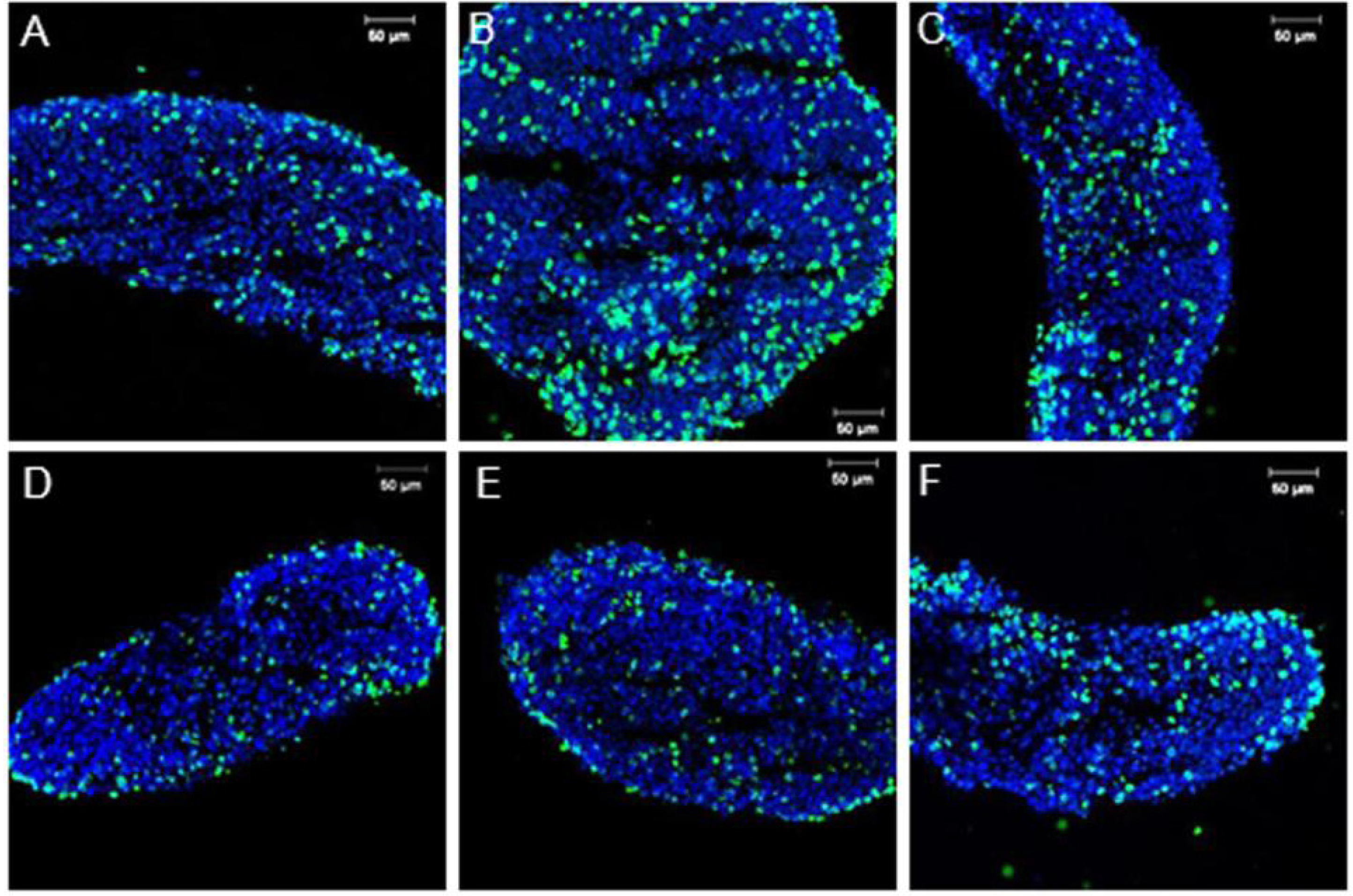
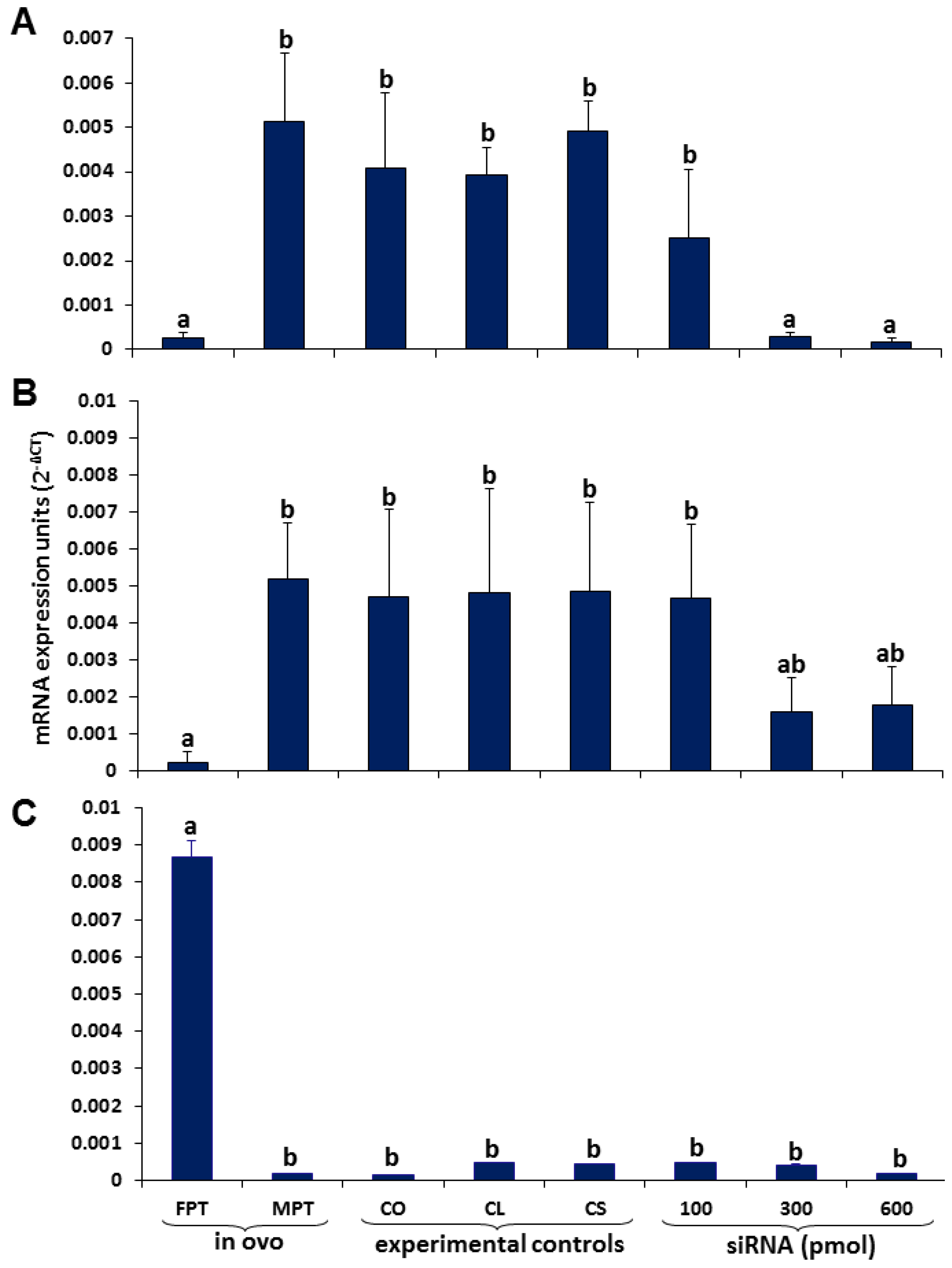
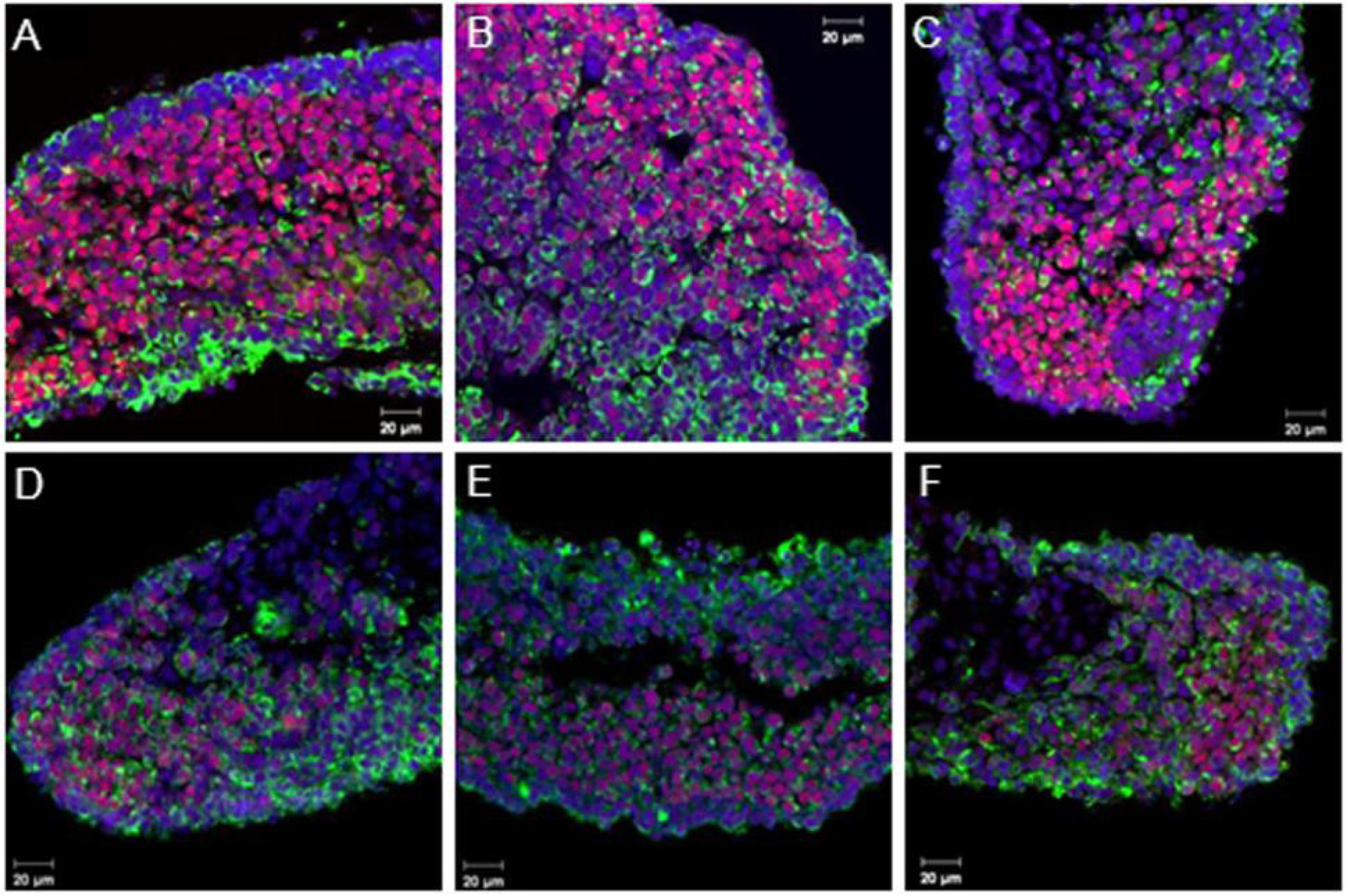
3.2. Coordinated Expression of Sox9 and Amh
3.3. Aromatase Expression
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Wibbels, T. Critical approaches to sex determination in Turtles. In The Biology of Sea Turtles; Lutz, P.L., Musik, J.A., Wineken, J., Eds.; CRC Press Inc: Boca Ratón, FL, USA, 2003; Volume 2, pp. 103–134. [Google Scholar]
- Pieau, C.; Dorizzi, M. Oestrogens and temperatura-dependent sex determination in reptiles: All is in the gonads. J. Endocrinol. 2004, 181, 367–377. [Google Scholar] [CrossRef]
- Koopman, P.; Gubbay, J.; Vivian, N.; Goodfellow, P.; Lovell-Badge, R. Male development of chromosomally female mice transgenic for Sry. Nature 1991, 351, 117–121. [Google Scholar] [CrossRef]
- Shoemaker, C.; Crews, D. Analyzing the coordinated gene network underlying temperature-dependent sex determination in Reptiles. Semin. Cell. Dev. Biol. 2009, 20, 293–303. [Google Scholar] [CrossRef]
- Rhen, T.; Schroeder, A. Molecular mechanisms of sex determination in reptiles. Sex. Dev. 2010, 4, 16–28. [Google Scholar] [CrossRef]
- Chue, J.; Smith, C.A. Sex determination and sexual differentiation in the avian model. FEBS J. 2011, 278, 1027–1034. [Google Scholar] [CrossRef]
- Kent, J.; Wheatley, S.C.; Andrews, J.E.; Sinclair, A.H.; Koopman, P. A male-specific role for SOX9 in vertebrate sex determination. Development 1996, 122, 2813–2822. [Google Scholar]
- Morais da Silva, S.; Hacker, A.; Harley, V.; Goodfellow, P.; Swain, A.; Lovell-Badge, R. Sox9 expression during gonadal development implies a conserved role for gene in testis differentiation in mammals and birds. Nat. Genet. 1996, 14, 62–68. [Google Scholar]
- Western, P.S.; Harry, J.L.; Graves, J.A.M.; Sinclair, A.H. Temperature-dependent sex determination: Upregulation of SOX9 expression after commitment to male development. Dev. Dyn. 1999, 214, 171–177. [Google Scholar] [CrossRef]
- Moreno-Mendoza, N.; Harley, V.R.; Merchant-Larios, H. Differential expression of SOX9 in gonads of the sea turtle Lepidochelys olivacea at male- or female- promoting temperatures. J. Exp. Zool. 1999, 284, 705–710. [Google Scholar] [CrossRef]
- Shoemaker, C.M.; Queen, J.; Crews, D. Response of candidate sex-determinig genes to changes in temperature reveals their involvement in the molecular network undelying temperature-dependent sex determination. Mol. Endocrinol. 2007, 21, 2750–2763. [Google Scholar] [CrossRef]
- Shoemaker, C.; Ramsey, M.; Queen, J.; Crews, D. Expression of Sox9, Mis, and Dmrt1, in the gonad of a species with temperature-dependent sex determination. Dev. Dyn. 2007, 236, 1055–1063. [Google Scholar] [CrossRef]
- Barske, L.A.; Capel, B. Estrogen represses SOX9 during sex determination in the red-eared slider turtle Trachemys scripta. Dev. Biol. 2010, 341, 305–314. [Google Scholar] [CrossRef]
- Harley, V.R.; Clarkson, M.J.; Argentaro, A. The molecular action and regulation of the testis-determining factors, SRY (sex determining region on the Y chromosome) and SOX9 [SRY-related high-mobility group (HMG) box 9]. Endocrinol. Rev. 2003, 24, 466–487. [Google Scholar] [CrossRef]
- Sim, H.; Argentaro, A.; Harley, V.R. Boys, girls and shuttling of SRY and SOX9. Trends Endocrinol. Metab. 2008, 19, 213–222. [Google Scholar] [CrossRef]
- Sekido, R.; Lovell-Badge, R. Sex determination involves synergistic action of SRY and SF1 on a specific Sox9 enhancer. Nature 2008, 453, 930–934. [Google Scholar] [CrossRef]
- Vidal, V.P.; Chaboissier, M.C.; de Rooji, D.G.; Schedl, A. Sox9 induces testis development in XX transgenic mice. Nat. Genet. 2001, 28, 216–217. [Google Scholar]
- Torres-Maldonado, L.C.; Moreno-Mendoza, N.; Landa, A.; Merchant-Larios, H. Timing of SOX9 downregulation and female sex determination in gonads of the sea turtle Lepidochelys olivacea. J. Exp. Zool. 2001, 290, 498–503. [Google Scholar] [CrossRef]
- Torres-Maldonado, L.C.; Landa-Piedra, A.; Moreno-Mendoza, N.; Marmolejo-Valencia, A.; Meza-Martínez, A.; Merchant-Larios, H. Expression profiles of Dax1, Dmrt1, and Sox9 during temperature sex determination in gonads of the sea turtle Lepidochelys olivacea. Gen. Comp. Endocrinol. 2002, 129, 20–26. [Google Scholar] [CrossRef]
- Moreno-Mendoza, N.; Harley, V.R.; Merchant-Larios, H. Temperature regulates SOX9 expression in cultured gonads of Lepidochelys olivacea, a species with temperature sex determination. Dev. Biol. 2001, 229, 319–326. [Google Scholar] [CrossRef]
- Shoemaker-Daly, C.M.; Jackson, K.; Yatsu, R.; Matsumoto, Y.; Crews, D. Genetic network underlying temperature-dependent sex determination is endogenously regulated by temperature in isolated cultured Trachemys scripta gonads. Dev. Dyn. 2010, 239, 1061–1075. [Google Scholar] [CrossRef]
- De Santa Barbara, P.; Bonneaud, N.; Boizet, B. Direct interaction of SRY-related protein SOX9 and Steroidogenic factor 1 regulates transcription of the human anti-Mullerian hormone gene. Mol. Cell Biol. 1998, 18, 6653–6718. [Google Scholar]
- Behringer, R.R.; Finegold, M.J.; Cate, R.L. Müllerian-inhibiting substance, function during mammalian sexual development. Cell 1994, 79, 415–425. [Google Scholar] [CrossRef]
- Merchant-Larios, H.; Díaz-Hernández, V.; Marmolejo-Valencia, A. Gonadal morphogenesis and gene expression in reptiles with temperature-dependent sex determination. Sex. Dev. 2010, 4, 50–61. [Google Scholar] [CrossRef]
- Oreal, E.; Pieau, C.; Mattei, M.G.; Josso, N.; Picard, J.Y.; Eusébe, D.C.; Magre, S. Early expression of Amh in chicken embryonic gonads precedes testicular Sox9 expression. Dev. Dyn. 1998, 212, 522–532. [Google Scholar] [CrossRef]
- Western, P.S.; Sinclair, A.H. Sex, genes and heat: Triggers of diversity. J. Exp. Zool. 2001, 290, 624–631. [Google Scholar] [CrossRef]
- Sifuentes-Romero, I.; Milton, L.S.; García-Gasca, A. Post-trasncriptional gene silencing by RNA interferance in non-mammalian vertebrate systems: Where do we stand? Mut. Res. 2011, 728, 158–171. [Google Scholar] [CrossRef]
- Davies, J.A.; Ladomery, M.; Hohenstein, P.; Michael, L.; Shafe, A.; Spraggon, L.; Hastie, N. Development of an siRNA-based method for repressing specific genes in renal organ culture and its use to show that the Wt1 tumour suppressor is required for nephron differentiation. Hum. Mol. Genet. 2004, 13, 235–246. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, P.; Guo, X.; Zhou, Z.; Sha, J. Hnrnpk, a protein differentially expressed in immature rat ovarian development, is required for normal primordial follicle assembly and development. Endocrinology 2011, 152, 1024–1035. [Google Scholar] [CrossRef]
- Nayak, G.; Prentice, H.M.; Milton, S.L. Role of neuroglobin in regulating reactive oxygen species in the brain of the anoxic-tolerant turtle Trachemys scripta. J. Neurochem. 2009, 110, 603–612. [Google Scholar] [CrossRef]
- Miller, D. Embryology of marine Turtles. In Biology of the Reptilia; Gans, C., Billet, F., Maderson, P.F.A., Eds.; Wiley: New York, NY, USA, 1985; pp. 270–328. [Google Scholar]
- Díaz-Hernández, V.; Marmolejo-Valencia, A.; Harfush, M.; Merchant-Larios, H. Formation of the genital ridges is preceded by a domain of ectopic Sox9-expressing cells in Lepidochelys olivacea. Dev. Biol. 2012, 361, 156–166. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−2ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lance, V.A. Is regulation of Aromatase expression in reptiles the key to understanding temperature-dependent sex determination? J. Exp. Zool. 2009, 311, 314–322. [Google Scholar] [CrossRef]
- Josso, N.; Belville, C.; di Clemente, N.; Picard, J.Y. AMH and AMH receptor defects in persistent Müllerian duct syndrome. Hum. Reprod. Update 2005, 11, 351–356. [Google Scholar] [CrossRef]
- Desvages, G.; Pieau, C. Aromatase activity in gonads of turtle embryos as a function of the incubation temperature of eggs. J. Steroid Biochem. Mol. Biol. 1992, 41, 851–853. [Google Scholar] [CrossRef]
- Desvages, G.; Girondot, M.; Pieau, C. Sensitive stages for the effects of temperature on gonadal aromatase activity in embryos of marine turtle Dermochelys coriacea. Gen. Comp. Endocrinol. 1993, 92, 54–61. [Google Scholar] [CrossRef]
- Jeyasuria, P.; Roosenburg, W.M.; Place, A.R. Role of P-450 Aromatase in sex determination of the diamondback terrapin, Malaclemys terrapin. J. Exp. Zool. 1994, 270, 95–111. [Google Scholar] [CrossRef]
- Rhen, T.; Metzger, K.; Schroeder, A.; Woodward, R. Expression of putative sex-determining genes during the thermosensitive period of gonad development in the snapping turtle, Chelydra serpentina. Sex Dev. 2007, 1, 255–270. [Google Scholar] [CrossRef]
- Ramsey, M.; Crews, D. Adrenal-kidney-gonad complex measurements may not predict gonad-specific changes in gene expression patterns during temperature-dependent sex determination in the red-eared slider turtle (Trachemys scripta elegans). J. Exp. Zool. A Ecol. Gen. Phys. 2007, 307, 463–470. [Google Scholar] [CrossRef]
- Valenzuela, N.; Neuwald, J.L.; Literman, R. Transcriptional evolution underlying vertebrate sexual development. Dev. Dyn. 2012, 242, 307–326. [Google Scholar] [CrossRef]
- Ryan, J.; Ludbrook, L.; Wilhelm, D.; Sinclair, A.; Koopman, P.; Bernard, P.; Harley, V.R. Analysis of gene function in cultured embryonic mouse gonads using nucleofection. Sex Dev. 2011, 5, 7–15. [Google Scholar] [CrossRef]
- Veitia, R.A. FoxL2 versus Sox9: A lifelong “battle of the sexes”. BioEssays 2010, 32, 375–380. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sifuentes-Romero, I.; Merchant-Larios, H.; Milton, S.L.; Moreno-Mendoza, N.; Díaz-Hernández, V.; García-Gasca, A. RNAi-Mediated Gene Silencing in a Gonad Organ Culture to Study Sex Determination Mechanisms in Sea Turtle. Genes 2013, 4, 293-305. https://doi.org/10.3390/genes4020293
Sifuentes-Romero I, Merchant-Larios H, Milton SL, Moreno-Mendoza N, Díaz-Hernández V, García-Gasca A. RNAi-Mediated Gene Silencing in a Gonad Organ Culture to Study Sex Determination Mechanisms in Sea Turtle. Genes. 2013; 4(2):293-305. https://doi.org/10.3390/genes4020293
Chicago/Turabian StyleSifuentes-Romero, Itzel, Horacio Merchant-Larios, Sarah L. Milton, Norma Moreno-Mendoza, Verónica Díaz-Hernández, and Alejandra García-Gasca. 2013. "RNAi-Mediated Gene Silencing in a Gonad Organ Culture to Study Sex Determination Mechanisms in Sea Turtle" Genes 4, no. 2: 293-305. https://doi.org/10.3390/genes4020293