Gene Expressions for Signal Transduction under Acidic Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Medium for Their Maintenance
2.2. Cell Culture under Different pH Conditions
2.3. DNA Microarray
2.4. Real-Time Quantitative Polymerase Chain Reaction (PCR)

{kind=link}
| Gene name | Sequence | |
|---|---|---|
| 18S rRNA | F; | TAGAGTGTTCAAAGCAGGCCC |
| R; | CCAACAAATAGAACCGCGGT | |
| IL-32 | F; | TCAAAGAGGGCTACCTGGAG |
| R; | TTTCAAGTAGAGGAGTGAGCTCTG | |
| ATP6V0D2 | F; | GACCCAGCAAGACTATATCAACC |
| R; | TGGAGATGAATTTTCAGGTCTTC | |
| TNFRSF9 | F; | AAACGGGGCAGAAAGAAACT |
| R; | CTTCTGGAAATCGGCAGCTA | |
| AREG | F; | GGGAGTGAGATTTCCCCTGT |
| R; | AGCCAGGTATTTGTGGTTCG | |
| DMGDH | F; | GAGCTCACGGCTGGATCTAC |
| R; | CCACCACCTGACCAGTTTCT | |
| ERBB3 | F; | TGCAGTGGATTCGAGAAGTG |
| R; | GGCAAACTTCCCATCGTAGA | |
2.5. Other Reagents
2.6. Statistical Analysis
3. Results
3.1. Highly Expressed Genes under Acidic Conditions in Mesothelioma Cells
| Gene | 2 h | 5 h | 24 h |
|---|---|---|---|
| number of genes | 260 | 175 | 379 |
| receptors | 29 | 22 | 35 |
| signal proteins 1 | 25 | 21 | 32 |
| cytokines 2 | 5 | 10 | 17 |
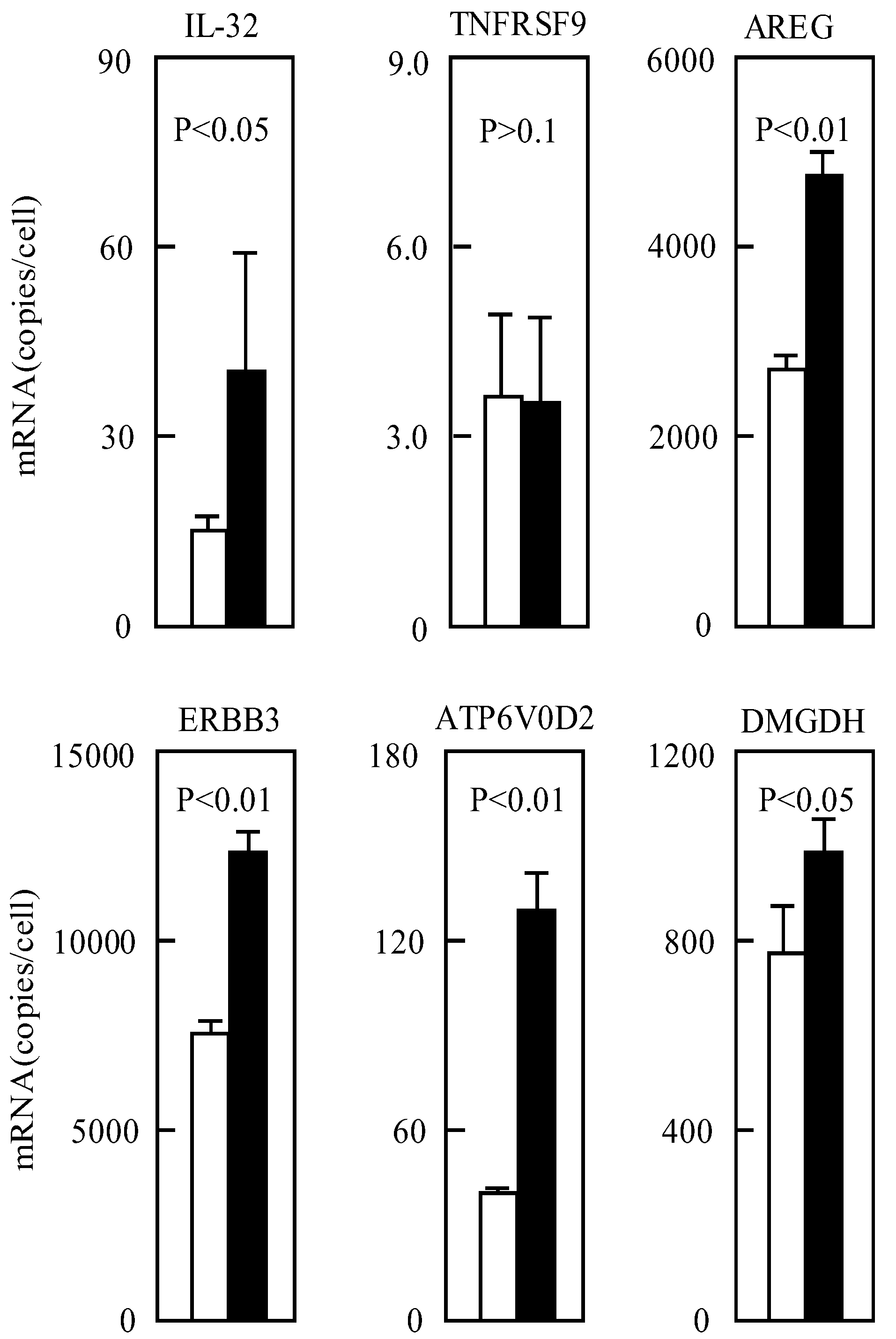
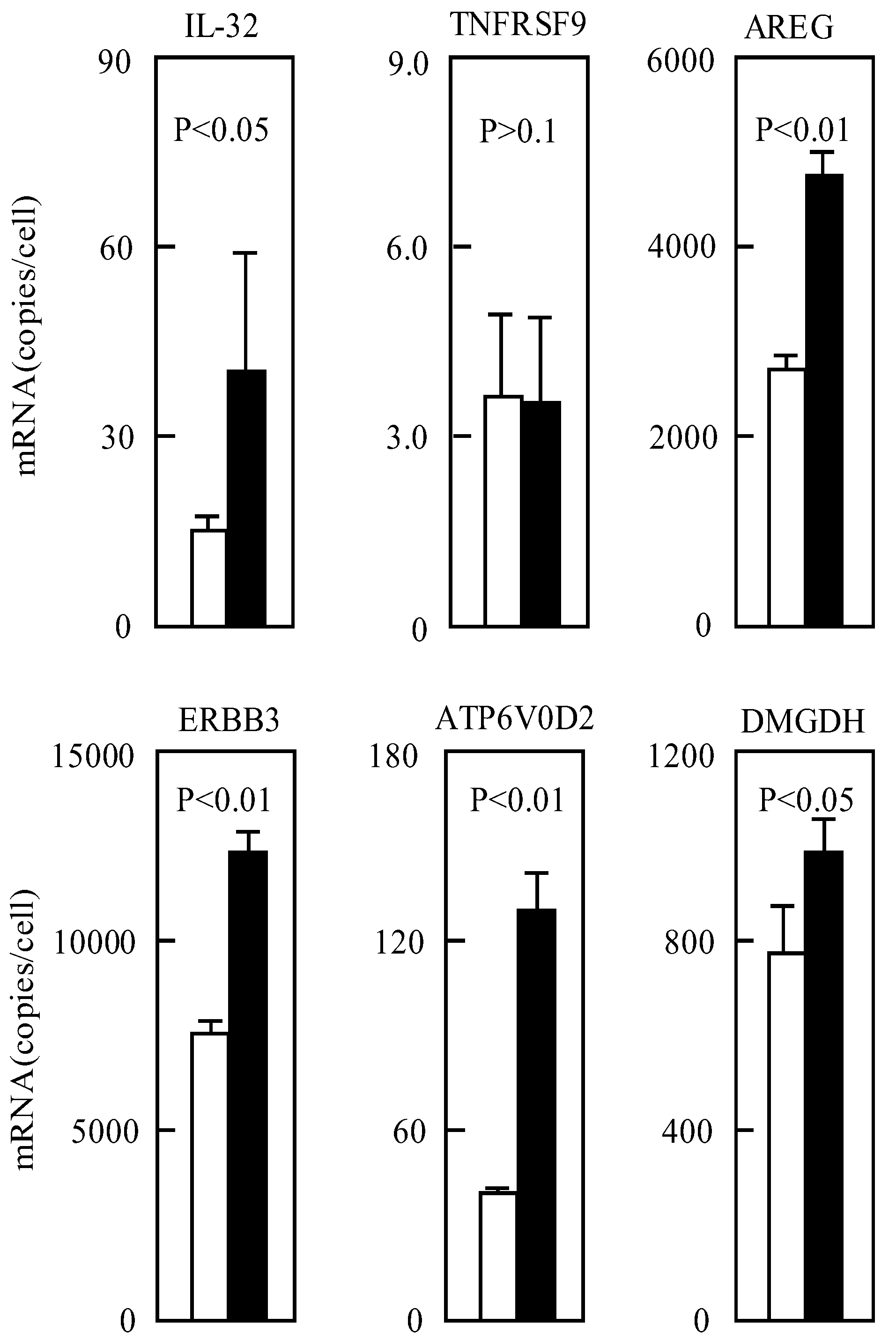
3.2. Gene Expression after Culture for a Short Period at pH 6.7
| Group | Expression level * | Number of genes | |||
|---|---|---|---|---|---|
| 2 h | 5 h | 24 h | Total | Signal ** | |
| A | >2 | >2 | >2 | 15 | 7 |
| B | >2 | <=2 | >2 | 22 | 3 |
| C | <=2 | >2 | >2 | 37 | 8 |
| D | <=2 | <=2 | >2 | 305 | 66 |
| E | >2 | >2 | <=2 | 32 | 6 |
| F | <=2 | >2 | <=2 | 91 | 32 |
| G | >2 | <=2 | <=2 | 191 | 43 |
| total | 693 | 165 | |||
| Gene | Ratio * | Accession number | Description |
|---|---|---|---|
| Group A | |||
| RSPO3 | 7.346 | NM_032784 | R-spondin 3 homolog (Xenopus laevis) |
| IL32 | 3.711 | NM_001012631 | interleukin 32 |
| TAS2R39 | 3.035 | NM_176881 | taste receptor, type 2, member 39 |
| SLAMF8 | 2.751 | NM_020125 | SLAM family member 8 |
| TRAF1 | 2.644 | NM_005658 | TNF receptor-associated factor 1 |
| IL8 | 2.519 | NM_000584 | interleukin 8 |
| RAB33A | 2.356 | NM_004794 | RAB33A, member RAS oncogene family |
| Group B | |||
| LOC553158 | 4.306 | NM_181334 | PRR5-ARHGAP8 fusion |
| PPP1R3E | 3.702 | XM_927029 | protein phosphatase 1, regulatory (inhibitor) subunit 3E |
| BDKRB2 | 2.168 | NM_000623 | bradykinin receptor B2 |
| Group C | |||
| TNFRSF9 | 5.464 | NM_001561 | tumor necrosis factor receptor superfamily, member 9 |
| FGF7 | 3.219 | NM_002009 | fibroblast growth factor 7 (keratinocyte growth factor) |
| ZNF226 | 2.926 | NM_015919 | zinc finger protein 226 |
| MGC17330 | 2.755 | NM_052880 | HGFL gene |
| IL1RAP | 2.551 | NM_134470 | interleukin 1 receptor accessory protein |
| NFKBIZ | 2.159 | NM_001005474 | nuclear factor of κ light polypeptide gene enhancer in B-cells inhibitor, ζ |
| OLR1 | 2.054 | NM_002543 | oxidized low density lipoprotein (lectin-like) receptor 1 |
| TRIB3 | 2.031 | NM_021158 | tribbles homolog 3 (Drosophila) |
| Group D | |||
| ERBB3 | 5.997 | NM_001982 | v-erb-b2 erythroblastic leukemia viral oncogene homolog 3 (avian) |
| AREG | 5.650 | NM_001657 | amphiregulin (schwannoma-derived growth factor) |
| LOC653193 | 4.485 | XM_926448 | similar to Amphiregulin precursor (AR) (Colorectum cell-derived growth factor) (CRDGF) |
| RARRES1 | 3.882 | NM_002888 | retinoic acid receptor responder (tazarotene induced) 1 |
| RRAD | 3.827 | NM_004165 | Ras-related associated with diabetes |
| CRELD1 | 3.707 | NM_001031717 | cysteine-rich with EGF-like domains 1 |
| ARHGAP8 | 3.547 | NM_001017526 | Rho GTPase activating protein 8 |
| GPR78 | 3.302 | NM_080819 | G protein-coupled receptor 78 |
| GDF15 | 3.112 | NM_004864 | growth differentiation factor 15 |
| PTP4A3 | 3.037 | NM_007079 | protein tyrosine phosphatase type IVA, member 3 |
| IL16 | 2.926 | NM_004513 | interleukin 16 (lymphocyte chemoattractant factor) |
| PAQR6 | 2.919 | NM_198406 | progestin and adipoQ receptor family member VI |
| OR52N4 | 2.915 | NM_001005175 | olfactory receptor, family 52, subfamily N, member 4 |
| OR56B1 | 2.913 | NM_001005180 | olfactory receptor, family 56, subfamily B, member 1 |
| PTPRQ | 2.834 | XM_926134 | protein tyrosine phosphatase, receptor type, Q |
| LOC439957 | 2.784 | XM_495805 | similar to Ig κ chain V-I region Walker precursor |
| TNFSF9 | 2.744 | NM_003811 | tumor necrosis factor (ligand) superfamily, member 9 |
| TNFSF7 | 2.714 | NM_001252 | tumor necrosis factor (ligand) superfamily, member 7 |
| GPR87 | 2.641 | NM_023915 | G protein-coupled receptor 87 |
| Group D | |||
| GTF2IRD2B | 2.609 | NM_001003795 | general transcription factor 21 repeat domain containing 2β |
| RGS7 | 2.573 | NM_002924 | regulator of G-protein signalling 7 |
| FOLR3 | 2.506 | NM_000804 | folate receptor 3 (γ) |
| RELB | 2.471 | NM_006509 | v-rel reticuloendotheliosis viral oncogene homolog B, nuclear factor of κ light polypeptide gene enhancer in B-cells 3 (avian) |
| TAS2R40 | 2.459 | NM_176882 | taste receptor, type 2, member 40 |
| CCL3L3 | 2.418 | NM_001001437 | chemokine (C-C motif) ligand 3-like 3 |
| GPR144 | 2.391 | NM_182611 | G protein-coupled receptor 144 |
| RND1 | 2.389 | NM_014470 | Rho family GTPase 1 |
| CD6 | 2.381 | NM_006725 | CD6 molecule |
| ZNF165 | 2.368 | NM_003447 | zinc finger protein 165 |
| ICHTHYIN | 2.353 | XM_371777 | ichthyin protein |
| PKD1L1 | 2.334 | NM_138295 | polycystic kidney disease 1 like 1 |
| NPHP1 | 2.318 | NM_207181 | nephronophthisis 1 (juvenile) |
| PTK6 | 2.312 | NM_005975 | PTK6 protein tyrosine kinase 6 |
| IL15RA | 2.282 | NM_002189 | interleukin 15 receptor, α |
| POU6F1 | 2.271 | NM_002702 | POU domain, class 6, transcription factor 1 |
| TNFRSF10C | 2.268 | NM_003841 | tumor necrosis factor receptor superfamily, member 10c, decoy without an intracellular domain |
| IL15 | 2.248 | NM_172175 | interleukin 15 |
| P2RY12 | 2.233 | NM_176876 | purinergic receptor P2Y, G-protein coupled, 12 |
| MST1 | 2.186 | NM_020998 | macrophage stimulating 1 (hepatocyte growth factor-like) |
| KDR | 2.184 | NM_002253 | kinase insert domain receptor (a type III receptor tyrosine kinase) |
| GPR68 | 2.174 | NM_003485 | G protein-coupled receptor 68 |
| GPR44 | 2.170 | NM_004778 | G protein-coupled receptor 44 |
| RAI17 | 2.162 | NM_020338 | retinoic acid induced 17 |
| OR10V1 | 2.156 | NM_001005324 | olfactory receptor, family 10, subfamily V, member 1 |
| ASB1 | 2.148 | NM_016114 | ankyrin repeat and SOCS box-containing 1 |
| CMTM1 | 2.146 | NM_181293 | CKLF-like MARVEL transmembrane domain containing 1 |
| PHF7 | 2.141 | NM_173341 | PHD finger protein 7 |
| GPRC5D | 2.114 | NM_018654 | G protein-coupled receptor, family C, group 5, member D |
| TP53INP2 | 2.108 | NM_021202 | tumor protein p53 inducible nuclear protein 2 |
| ARHGAP15 | 2.082 | NM_018460 | Rho GTPase activating protein 15 |
| GEFT | 2.066 | NM_182947 | RAC/CDC42 exchange factor |
| PIM1 | 2.062 | NM_002648 | pim-1 oncogene |
| TNFRSF25 | 2.055 | NM_148973 | tumor necrosis factor receptor superfamily, member 25 |
| GPR157 | 2.049 | NM_024980 | G protein-coupled receptor 157 |
| NR2E3 | 2.045 | NM_014249 | nuclear receptor subfamily 2, group E, member 3 |
| LOC619207 | 2.042 | XM_927510 | scavenger receptor protein family member |
| WISP3 | 2.033 | NM_003880 | WNT1 inducible signaling pathway protein 3 |
| P2RX4 | 2.030 | NM_002560 | purinergic receptor P2X, ligand-gated ion channel, 4 |
| RASD2 | 2.029 | NM_014310 | RASD family, member 2 |
| FGF2 | 2.028 | NM_002006 | fibroblast growth factor 2 (basic) |
| RGR | 2.011 | NM_001012720 | retinal G protein coupled receptor |
| Group D | |||
| NRXN2 | 2.011 | NM_015080 | neurexin 2 |
| EDG4 | 2.009 | NM_004720 | endothelial differentiation, lysophosphatidic acid G-protein-coupled receptor, 4 |
| KGFLP1 | 2.006 | NM_174950 | keratinocyte growth factor-like protein 1 |
| PTPRH | 2.005 | NM_002842 | protein tyrosine phosphatase, receptor type, H |
| OR52A5 | 2.001 | NM_001005160 | olfactory receptor, family 52, subfamily A, member 5 |
| Group E | |||
| KLF9 | 3.169 | NM_001206 | Kruppel-like factor 9 |
| CLASP2 | 2.029 | NM_015097 | cytoplasmic linker associated protein 2 |
| E2F5 | 2.267 | NM_001951 | E2F transcription factor 5, p130-binding |
| ZNF474 | 2.203 | NM_207317 | zinc finger protein 474 |
| GPR37 | 2.411 | NM_005302 | G protein-coupled receptor 37 (endothelin receptor type B-like) |
| PAX5 | 2.097 | NM_016734 | paired box gene 5 (B-cell lineage specific activator) |
| Group F | |||
| OR4D6 | 2.809 | NM_001004708 | olfactory receptor, family 4, subfamily D, member 6 |
| OR5B12 | 2.783 | NM_001004733 | olfactory receptor, family 5, subfamily B, member 12 |
| VENTX | 2.515 | NM_014468 | VENT homeobox homolog (Xenopus laevis) |
| UNC5B | 2.438 | NM_170744 | unc-5 homolog B (C. elegans) |
| OR1J4 | 2.416 | NM_001004452 | olfactory receptor, family 1, subfamily J, member 4 |
| NR5A1 | 2.338 | NM_004959 | nuclear receptor subfamily 5, group A, member 1 |
| SESN2 | 2.331 | NM_031459 | sestrin 2 |
| CCL25 | 2.308 | NM_148888 | chemokine (C-C motif) ligand 25 |
| IL21R | 2.301 | NM_021798 | interleukin 21 receptor |
| ATF3 | 2.220 | NM_001030287 | activating transcription factor 3 |
| TLR1 | 2.212 | NM_003263 | toll-like receptor 1 |
| C1QTNF7 | 2.202 | NM_031911 | C1q and tumor necrosis factor related protein 7 |
| TBX19 | 2.186 | NM_005149 | T-box 19 |
| MXD1 | 2.163 | NM_002357 | MAX dimerization protein 1 |
| GTPBP2 | 2.148 | NM_019096 | GTP binding protein 2 |
| NR4A2 | 2.117 | NM_006186 | nuclear receptor subfamily 4, group A, member 2 |
| PHLDA1 | 2.116 | NM_007350 | pleckstrin homology-like domain, family A, member 1 |
| LCP1 | 2.111 | NM_002298 | lymphocyte cytosolic protein 1 (L-plastin) |
| FGFBP1 | 2.111 | NM_005130 | fibroblast growth factor binding protein 1 |
| OR2B11 | 2.078 | NM_001004492 | olfactory receptor, family 2, subfamily B, member 11 |
| OR56A3 | 2.073 | NM_001003443 | olfactory receptor, family 56, subfamily A, member 3 |
| GH2 | 2.071 | NM_002059 | growth hormone 2 |
| PTHLH | 2.061 | NM_002820 | parathyroid hormone-like hormone |
| THBD | 2.059 | NM_000361 | thrombomodulin |
| HGF | 2.055 | NM_000601 | hepatocyte growth factor (hepapoietin A; scatter factor) |
| ARTN | 2.051 | NM_003976 | artemin |
| EPHA8 | 2.040 | NM_020526 | EPH receptor A8 |
| CD200R1 | 2.023 | NM_138939 | CD200 receptor 1 |
| FZD7 | 2.017 | NM_003507 | frizzled homolog 7 (Drosophila) |
| S100A12 | 2.012 | NM_005621 | S100 calcium binding protein A12 (calgranulin C) |
| Group F | |||
| SPIC | 2.008 | NM_152323 | Spi-C transcription factor (Spi-1/PU.1 related) |
| VSIG4 | 2.006 | NM_007268 | V-set and immunoglobulin domain containing 4 |
| Group G | |||
| TCF21 | 3.518 | NM_198392 | transcription factor 21 |
| DOK6 | 2.827 | NM_152721 | docking protein 6 |
| FZD3 | 2.815 | NM_017412 | frizzled homolog 3 (Drosophila) |
| CD86 | 2.686 | NM_006889 | CD86 molecule |
| OR2L13 | 2.677 | NM_175911 | olfactory receptor, family 2, subfamily L, member 13 |
| IL18RAP | 2.557 | NM_003853 | interleukin 18 receptor accessory protein |
| TRPA1 | 2.427 | NM_007332 | transient receptor potential cation channel, subfamily A, member 1 |
| RTP3 | 2.417 | NM_031440 | receptor transporter protein 3 |
| GRIA2 | 2.353 | NM_000826 | glutamate receptor, ionotropic, AMPA 2 |
| OR51E1 | 2.351 | NM_152430 | olfactory receptor, family 51, subfamily E, member 1 |
| GRM2 | 2.343 | NM_000839 | glutamate receptor, metabotropic 2 |
| FCRLM1 | 2.335 | NM_032738 | Fc receptor-like and mucin-like 1 |
| NR4A3 | 2.332 | NM_173199 | nuclear receptor subfamily 4, group A, member 3 |
| SPI1 | 2.288 | NM_003120 | spleen focus forming virus (SFFV) proviral integration oncogene spi1 |
| SMAD6 | 2.282 | NM_005585 | SMAD, mothers against DPP homolog 6 (Drosophila) |
| LOC642400 | 2.281 | XM_925921 | similar to tripartite motif protein 17 |
| CAMTA1 | 2.274 | NM_015215 | calmodulin binding transcription activator 1 |
| GDF6 | 2.260 | NM_001001557 | growth differentiation factor 6 |
| OR51B2 | 2.258 | NM_033180 | olfactory receptor, family 51, subfamily B, member 2 |
| OR7D4 | 2.238 | NM_001005191 | olfactory receptor, family 7, subfamily D, member 4 |
| ECGF1 | 2.227 | NM_001953 | endothelial cell growth factor 1 (platelet-derived) |
| LAIR1 | 2.218 | NM_002287 | leukocyte-associated immunoglobulin-like receptor 1 |
| NELL1 | 2.209 | NM_006157 | NEL-like 1 (chicken) |
| OR8J1 | 2.188 | NM_001005205 | olfactory receptor, family 8, subfamily J, member 1 |
| GRM3 | 2.178 | NM_000840 | glutamate receptor, metabotropic 3 |
| PRKCQ | 2.170 | NM_006257 | protein kinase C, θ |
| PPP1R3F | 2.164 | NM_033215 | protein phosphatase 1, regulatory (inhibitor) subunit 3F |
| LOC642338 | 2.150 | XM_925874 | similar to vomeronasal 1 receptor, C3 |
| CPNE5 | 2.148 | NM_020939 | copine V |
| EPHB6 | 2.134 | NM_004445 | EPH receptor B6 |
| OR51M1 | 2.119 | NM_001004756 | olfactory receptor, family 51, subfamily M, member 1 |
| PTPRC | 2.105 | NM_002838 | protein tyrosine phosphatase, receptor type, C |
| EBF3 | 2.100 | NM_001005463 | early B-cell factor 3 |
| BMPR1B | 2.091 | NM_001203 | bone morphogenetic protein receptor, type IB |
| HRH4 | 2.084 | NM_021624 | histamine receptor H4 |
| SHE | 2.082 | NM_001010846 | Src homology 2 domain containing E |
| T2R55 | 2.073 | NM_181429 | taste receptor T2R55 |
| SBK1 | 2.067 | NM_001024401 | SH3-binding domain kinase 1 |
| RASGRP2 | 2.048 | NM_005825 | RAS guanyl releasing protein 2 (calcium and DAG-regulated) |
| CD96 | 2.047 | NM_005816 | CD96 molecule |
| GRIN2B | 2.028 | NM_000834 | glutamate receptor, ionotropic, N-methyl D-aspartate 2B |
| Group G | |||
| YAF2 | 2.018 | NM_001012424 | YY1 associated factor 2 |
| LCP2 | 2.001 | NM_005565 | lymphocyte cytosolic protein 2 (SH2 domain containing leukocyte protein of 76 kDa) |
3.3. Genes Whose Expression Was Repressed at Acidic pH
| Gene | 2 h | 5 h | 24 h |
|---|---|---|---|
| number of genes | 385 | 141 | 412 |
| receptors | 32 | 14 | 35 |
| signal proteins 1 | 31 | 14 | 76 |
| cytokines 2 | 8 | 0 | 7 |
| Group | Expression level * | Number of genes | |||
|---|---|---|---|---|---|
| 2 h | 5 h | 24 h | Total | Signal ** | |
| A | <0.5 | <0.5 | <0.5 | 8 | 4 |
| B | <0.5 | >=0.5 | <0.5 | 9 | 3 |
| C | >=0.5 | <0.5 | <0.5 | 32 | 4 |
| D | >=0.5 | >=0.5 | <0.5 | 363 | 107 |
| E | <0.5 | <0.5 | >=0.5 | 25 | 8 |
| F | >=0.5 | <0.5 | >=0.5 | 76 | 12 |
| G | <0.5 | >=0.5 | >=0.5 | 343 | 56 |
| total | 856 | 194 | |||
| Gene | Ratio * | Accession number | Description |
|---|---|---|---|
| Group A | |||
| IL11 | 0.158 | NM_000641 | interleukin 11 |
| CCRL2 | 0.324 | NM_003965 | chemokine (C-C motif) receptor-like 2 |
| CD300LG | 0.444 | NM_145273 | CD300 molecule-like family member g |
| ATOH1 | 0.459 | NM_005172 | atonal homolog 1 (Drosophila) |
| Group B | |||
| RASGEF1C | 0.486 | NM_001031799 | RasGEF domain family, member 1C |
| LGR5 | 0.491 | NM_003667 | leucine-rich repeat-containing G protein-coupled receptor 5 |
| HSH2D | 0.493 | NM_032855 | hematopoietic SH2 domain containing |
| Group C | |||
| TLR4 | 0.099 | NM_138554 | toll-like receptor 4 |
| TSSK2 | 0.421 | NM_053006 | testis-specific serine kinase 2 |
| ADRB2 | 0.475 | NM_000024 | adrenergic, β-2-, receptor, surface |
| FLRT2 | 0.400 | NM_013231 | fibronectin leucine rich transmembrane protein 2 |
| Group D | |||
| E2F2 | 0.150 | NM_004091 | E2F transcription factor 2 |
| ADRA2A | 0.193 | NM_000681 | adrenergic, α-2A-, receptor |
| APLN | 0.243 | NM_017413 | apelin, AGTRL1 ligand |
| REEP1 | 0.244 | NM_022912 | receptor accessory protein 1 |
| ARHGAP26 | 0.252 | NM_015071 | Rho GTPase activating protein 26 |
| UHRF1 | 0.259 | NM_013282 | ubiquitin-like, containing PHD and RING finger domains, 1 |
| ZNF367 | 0.261 | NM_153695 | zinc finger protein 367 |
| POU5F1 | 0.267 | NM_203289 | POU domain, class 5, transcription factor 1 |
| RGS4 | 0.269 | NM_005613 | regulator of G-protein signalling 4 |
| RHOJ | 0.269 | NM_020663 | ras homolog gene family, member J |
| MCF2 | 0.270 | NM_005369 | MCF.2 cell line derived transforming sequence |
| CHRNA5 | 0.276 | NM_000745 | cholinergic receptor, nicotinic, α 5 |
| GPR115 | 0.285 | NM_153838 | G protein-coupled receptor 115 |
| SORCS3 | 0.286 | NM_014978 | sortilin-related VPS10 domain containing receptor 3 |
| RBM14 | 0.287 | NM_006328 | RNA binding motif protein 14 |
| PDE4B | 0.298 | NM_001037339 | phosphodiesterase 4B, cAMP-specific (phosphodiesterase E4 dunce homolog, Drosophila) |
| PIK3CG | 0.310 | NM_002649 | phosphoinositide-3-kinase, catalytic, γ polypeptide |
| RGPD2 | 0.311 | NM_001024457 | RANBP2-like and GRIP domain containing 2 |
| TP53RK | 0.314 | NM_033550 | TP53 regulating kinase |
| MAP2K6 | 0.320 | NM_002758 | mitogen-activated protein kinase kinase 6 |
| TP73 | 0.330 | NM_005427 | tumor protein p73 |
| GPR63 | 0.338 | NM_030784 | G protein-coupled receptor 63 |
| FST | 0.340 | NM_006350 | follistatin |
| MPP4 | 0.347 | NM_033066 | membrane protein, palmitoylated 4 (MAGUK p55 subfamily member 4) |
| PDE4D | 0.350 | NM_006203 | phosphodiesterase 4D, cAMP-specific (phosphodiesterase E3 dunce homolog, Drosophila) |
| ANXA10 | 0.355 | NM_007193 | annexin A10 |
| Group D | |||
| RBL1 | 0.355 | NM_002895 | retinoblastoma-like 1 (p107) |
| KIT | 0.360 | NM_000222 | v-kit Hardy-Zuckerman 4 feline sarcoma viral oncogene homolog |
| PBX1 | 0.368 | NM_002585 | pre-B-cell leukemia transcription factor 1 |
| MTUS1 | 0.371 | NM_001001924 | mitochondrial tumor suppressor 1 |
| RORB | 0.386 | NM_006914 | RAR-related orphan receptor B |
| LHX6 | 0.389 | NM_014368 | LIM homeobox 6 |
| PAQR4 | 0.392 | NM_152341 | progestin and adipoQ receptor family member IV |
| ABRA | 0.394 | NM_139166 | actin-binding Rho activating protein |
| GDAP1 | 0.396 | NM_018972 | ganglioside-induced differentiation-associated protein 1 |
| C1QTNF2 | 0.399 | NM_031908 | C1q and tumor necrosis factor related protein 2 |
| CMTM1 | 0.400 | NM_181289 | CKLF-like MARVEL transmembrane domain containing 1 |
| MLR1 | 0.404 | NM_153686 | transcription factor MLR1 |
| TSPAN8 | 0.405 | NM_004616 | tetraspanin 8 |
| SH2D4B | 0.406 | NM_207372 | SH2 domain containing 4B |
| E2F1 | 0.406 | NM_005225 | E2F transcription factor 1 |
| VANGL1 | 0.411 | NM_138959 | vang-like 1 (van gogh, Drosophila) |
| DUSP6 | 0.415 | NM_001946 | dual specificity phosphatase 6 |
| FZD3 | 0.416 | NM_017412 | frizzled homolog 3 (Drosophila) |
| PPARGC1A | 0.417 | NM_013261 | peroxisome proliferative activated receptor, γ, coactivator 1, α |
| HOXB7 | 0.419 | NM_004502 | homeobox B7 |
| PTGER2 | 0.420 | NM_000956 | prostaglandin E receptor 2 (subtype EP2), 53 kDa |
| NGEF | 0.421 | NM_019850 | neuronal guanine nucleotide exchange factor |
| FGF18 | 0.421 | NM_033649 | fibroblast growth factor 18 |
| LOC653528 | 0.425 | XM_927910 | similar to Teratocarcinoma-derived growth factor 2 (Epidermal growth factor-like cripto protein CR3) (Cripto-3 growth factor) |
| OR4N2 | 0.426 | NM_001004723 | olfactory receptor, family 4, subfamily N, member 2 |
| NKX6-2 | 0.429 | NM_177400 | NK6 transcription factor related, locus 2 (Drosophila) |
| NFKBIL2 | 0.431 | NM_013432 | nuclear factor of κ light polypeptide gene enhancer in B-cells inhibitor-like 2 |
| PTPN22 | 0.431 | NM_012411 | protein tyrosine phosphatase, non-receptor type 22 (lymphoid) |
| LOC392269 | 0.432 | XM_928112 | similar to Transcription factor SOX-2 |
| MAL2 | 0.432 | NM_052886 | mal, T-cell differentiation protein 2 |
| SELPLG | 0.434 | NM_003006 | selectin P ligand |
| GPR177 | 0.434 | NM_001002292 | G protein-coupled receptor 177 |
| NCOA5 | 0.437 | NM_020967 | nuclear receptor coactivator 5 |
| RIF1 | 0.437 | NM_018151 | RAP1 interacting factor homolog (yeast) |
| GPR3 | 0.439 | NM_005281 | G protein-coupled receptor 3 |
| CDC14A | 0.439 | NM_003672 | CDC14 cell division cycle 14 homolog A (S. cerevisiae) |
| RP3-509I19.5 | 0.444 | XM_294019 | similar to ECT2 protein (Epithelial cell transforming sequence 2 oncogene) |
| ADORA1 | 0.444 | NM_000674 | adenosine A1 receptor |
| PTCH | 0.446 | NM_000264 | patched homolog (Drosophila) |
| TCF21 | 0.446 | NM_003206 | transcription factor 21 |
| SPRY4 | 0.448 | NM_030964 | sprouty homolog 4 (Drosophila) |
| CBX2 | 0.450 | NM_005189 | chromobox homolog 2 (Pc class homolog, Drosophila) |
| OR6C74 | 0.451 | NM_001005490 | olfactory receptor, family 6, subfamily C, member 74 |
| Group D | |||
| CXCL14 | 0.452 | NM_004887 | chemokine (C-X-C motif) ligand 14 |
| CUBN | 0.453 | NM_001081 | cubilin (intrinsic factor-cobalamin receptor) |
| NRG2 | 0.457 | NM_013985 | neuregulin 2 |
| SGIP1 | 0.457 | NM_032291 | SH3-domain GRB2-like (endophilin) interacting protein 1 |
| GNGT2 | 0.457 | NM_031498 | guanine nucleotide binding protein (G protein), γ transducing activity polypeptide 2 |
| EBF | 0.458 | NM_024007 | early B-cell factor |
| ACVR1C | 0.458 | NM_145259 | activin A receptor, type IC |
| PHTF2 | 0.458 | NM_020432 | putative homeodomain transcription factor 2 |
| RASSF1 | 0.460 | NM_007182 | Ras association (RalGDS/AF-6) domain family 1 |
| GPR109A | 0.462 | NM_177551 | G protein-coupled receptor 109A |
| TSHR | 0.463 | NM_000369 | thyroid stimulating hormone receptor |
| SIM2 | 0.468 | NM_009586 | single-minded homolog 2 (Drosophila) |
| GABRA6 | 0.469 | NM_000811 | γ-aminobutyric acid (GABA) A receptor, alpha 6 |
| LAT2 | 0.469 | NM_032464 | linker for activation of T cells family, member 2 |
| PHKG1 | 0.472 | NM_006213 | phosphorylase kinase, γ 1 (muscle) |
| RGPD4 | 0.473 | XM_496581 | RANBP2-like and GRIP domain containing 4 |
| NKD1 | 0.474 | NM_033119 | naked cuticle homolog 1 (Drosophila) |
| ZNF588 | 0.475 | NM_001013746 | zinc finger protein 588 |
| SH3TC2 | 0.476 | NM_024577 | SH3 domain and tetratricopeptide repeats 2 |
| FZD1 | 0.478 | NM_003505 | frizzled homolog 1 (Drosophila) |
| PKMYT1 | 0.478 | NM_004203 | protein kinase, membrane associated tyrosine/threonine 1 |
| DUSP4 | 0.480 | NM_001394 | dual specificity phosphatase 4 |
| WDR4 | 0.480 | NM_018669 | WD repeat domain 4 |
| WDR76 | 0.481 | NM_024908 | WD repeat domain 76 |
| WDHD1 | 0.483 | NM_001008396 | WD repeat and HMG-box DNA binding protein 1 |
| HOXA7 | 0.485 | NM_006896 | homeobox A7 |
| WDR69 | 0.486 | NM_178821 | WD repeat domain 69 |
| TFAP2C | 0.487 | NM_003222 | transcription factor AP-2 γ (activating enhancer binding protein 2 γ) |
| CDGAP | 0.488 | NM_020754 | Cdc42 GTPase-activating protein |
| RPIB9 | 0.491 | NM_138290 | Rap2-binding protein 9 |
| IFNAR1 | 0.493 | NM_000629 | interferon (α, β and ω) receptor 1 |
| POU3F2 | 0.493 | NM_005604 | POU domain, class 3, transcription factor 2 |
| LOC402279 | 0.497 | XM_377945 | similar to glutamate receptor, metabotropic 8 |
| EYA4 | 0.498 | NM_004100 | eyes absent homolog 4 (Drosophila) |
| ISL1 | 0.498 | NM_002202 | ISL1 transcription factor, LIM/homeodomain, (islet-1) |
| SIRPD | 0.499 | NM_178460 | signal-regulatory protein δ |
| NEDD9 | 0.499 | NM_182966 | neural precursor cell expressed, developmentally down-regulated 9 |
| TLR3 | 0.500 # | NM_003265 | toll-like receptor 3 |
| Group E | |||
| RAPSN | 0.354 | NM_005055 | receptor-associated protein of the synapse, 43 kDa |
| GRAP | 0.363 | NM_006613 | GRB2-related adaptor protein |
| CD48 | 0.410 | NM_001778 | CD48 molecule |
| LOC642966 | 0.428 | XM_926351 | similar to olfactory receptor 139 |
| SALL1 | 0.437 | NM_002968 | sal-like 1 (Drosophila) |
| Group E | |||
| GLIS1 | 0.438 | NM_147193 | GLIS family zinc finger 1 |
| FOLR1 | 0.471 | NM_016725 | folate receptor 1 (adult) |
| NRG4 | 0.482 | NM_138573 | neuregulin 4 |
| Group F | |||
| TACR1 | 0.408 | NM_015727 | tachykinin receptor 1 |
| NHLH1 | 0.414 | NM_005598 | nescient helix loop helix 1 |
| NF2 | 0.420 | NM_181825 | neurofibromin 2 (bilateral acoustic neuroma) |
| LOC440607 | 0.427 | NM_001004340 | Fc-γ receptor I B2 |
| MAF | 0.443 | NM_001031804 | v-maf musculoaponeurotic fibrosarcoma oncogene homolog (avian) |
| ZNF160 | 0.449 | NM_033288 | zinc finger protein 160 |
| DUSP2 | 0.454 | NM_004418 | dual specificity phosphatase 2 |
| SOCS1 | 0.456 | NM_003745 | suppressor of cytokine signaling 1 |
| CHRNA3 | 0.471 | NM_000743 | cholinergic receptor, nicotinic, α 3 |
| CRLF2 | 0.481 | NM_022148 | cytokine receptor-like factor 2 |
| MRAP | 0.487 | NM_178817 | melanocortin 2 receptor accessory protein |
| RPIP8 | 0.491 | NM_006695 | RaP2 interacting protein 8 |
| Group G | |||
| CLEC4G | 0.198 | NM_198492 | C-type lectin superfamily 4, member G |
| FSTL4 | 0.246 | NM_015082 | follistatin-like 4 |
| RAB6C | 0.282 | NM_032144 | RAB6C, member RAS oncogene family |
| CSF3 | 0.295 | NM_172220 | colony stimulating factor 3 (granulocyte) |
| OR2T34 | 0.301 | NM_001001821 | olfactory receptor, family 2, subfamily T, member 34 |
| RRP22 | 0.304 | NM_001007279 | RAS-related on chromosome 22 |
| UTF1 | 0.305 | NM_003577 | undifferentiated embryonic cell transcription factor 1 |
| CHRND | 0.306 | NM_000751 | cholinergic receptor, nicotinic, δ |
| GPR6 | 0.316 | NM_005284 | G protein-coupled receptor 6 |
| ANGPTL6 | 0.324 | NM_031917 | angiopoietin-like 6 |
| OR2M7 | 0.346 | NM_001004691 | olfactory receptor, family 2, subfamily M, member 7 |
| OR10P1 | 0.356 | NM_206899 | olfactory receptor, family 10, subfamily P, member 1 |
| FOXD3 | 0.364 | NM_012183 | forkhead box D3 |
| ZAP70 | 0.377 | NM_207519 | ζ-chain (TCR) associated protein kinase 70 kDa |
| PTGER3 | 0.392 | NM_000957 | prostaglandin E receptor 3 (subtype EP3) |
| CDX4 | 0.395 | NM_005193 | caudal type homeobox transcription factor 4 |
| TBX21 | 0.405 | NM_013351 | T-box 21 |
| TAS2R13 | 0.408 | NM_023920 | taste receptor, type 2, member 13 |
| IL17RE | 0.414 | NM_153482 | interleukin 17 receptor E |
| PRDM9 | 0.415 | NM_020227 | PR domain containing 9 |
| CXCL12 | 0.422 | NM_199168 | chemokine (C-X-C motif) ligand 12 (stromal cell-derived factor 1) |
| LILRA4 | 0.430 | NM_012276 | leukocyte immunoglobulin-like receptor, subfamily A (with TM domain), member 4 |
| LOC642506 | 0.433 | XM_926003 | similar to double homeobox 4c |
| NEUROD6 | 0.435 | NM_022728 | neurogenic differentiation 6 |
| KLF14 | 0.438 | NM_138693 | Kruppel-like factor 14 |
| TFAP2E | 0.439 | NM_178548 | transcription factor AP-2 ε (activating enhancer binding protein 2 ε) |
| Group G | |||
| CCL1 | 0.439 | NM_002981 | chemokine (C-C motif) ligand 1 |
| VAV3 | 0.439 | NM_006113 | vav 3 oncogene |
| IRS3L | 0.444 | XM_498229 | insulin receptor substrate 3-like |
| GPR81 | 0.445 | NM_032554 | G protein-coupled receptor 81 |
| GPR32 | 0.445 | NM_001506 | G protein-coupled receptor 32 |
| GDF7 | 0.446 | NM_182828 | growth differentiation factor 7 |
| WDR42C | 0.447 | XM_293354 | WD repeat domain 42C |
| LOC619207 | 0.454 | XM_927516 | scavenger receptor protein family member |
| FOLR1 | 0.457 | NM_016724 | folate receptor 1 (adult) |
| ADRA1D | 0.457 | NM_000678 | adrenergic, α-1D-, receptor |
| IL12RB2 | 0.459 | NM_001559 | interleukin 12 receptor, β 2 |
| GRIN1 | 0.460 | NM_007327 | glutamate receptor, ionotropic, N-methyl D-aspartate 1 |
| SHC2 | 0.461 | XM_375550 | SHC (Src homology 2 domain containing) transforming protein 2 |
| RAXL1 | 0.464 | NM_032753 | retina and anterior neural fold homeobox like 1 |
| CAMK2B | 0.472 | NM_172084 | calcium/calmodulin-dependent protein kinase (CaM kinase) II β |
| CCL15 | 0.473 | NM_004167 | chemokine (C-C motif) ligand 15 |
| FSHR | 0.474 | NM_000145 | follicle stimulating hormone receptor |
| WDR40B | 0.478 | NM_178470 | WD repeat domain 40B |
| MAFB | 0.482 | NM_005461 | v-maf musculoaponeurotic fibrosarcoma oncogene homolog B (avian) |
| TPRX1 | 0.484 | NM_198479 | tetra-peptide repeat homeobox 1 |
| FLT1 | 0.487 | NM_002019 | fms-related tyrosine kinase 1 (vascular endothelial growth factor/vascular permeability factor receptor) |
| OLIG2 | 0.488 | NM_005806 | oligodendrocyte lineage transcription factor 2 |
| TBXA2R | 0.490 | NM_001060 | thromboxane A2 receptor |
| SSTR5 | 0.490 | NM_001053 | somatostatin receptor 5 |
| MYOG | 0.491 | NM_002479 | myogenin (myogenic factor 4) |
| OR2AG1 | 0.492 | NM_001004489 | olfactory receptor, family 2, subfamily AG, member 1 |
| FOXD4L1 | 0.495 | NM_012184 | forkhead box D4-like 1 |
| PSPN | 0.495 | NM_004158 | persephin |
| PJCG6 | 0.496 | NM_001040066 | similar to olfactory receptor 873 |
| TSHR | 0.499 | NM_001018036 | thyroid stimulating hormone receptor |
3.4. The Gene Expressions in Various Cells
4. Discussion
5. Conclusions
Acknowledgements
References
- Vaupel, P.; Kallinowski, F.; Okunieff, P. Blood flow, oxygen and nutrient supply, and metabolic microenvironment of human tumors: A review. Cancer Res. 1989, 49, 6449–6465. [Google Scholar]
- Helmlinger, G.; Yuan, F.; Dellian, M.; Jain, R.K. Interstitial pH and pO2 gradients in solid tumors in vivo: high-resolution measurements reveal a lack of correlation. Nat. Med. 1997, 3, 177–182. [Google Scholar] [CrossRef]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar]
- Simmen, H.P.; Blaser, J. Analysis of pH and pO2 in abscesses, peritoneal fluid, and drainage fluid in the presence or absence of bacterial infection during and after abdominal surgery. Am. J. Surg. 1993, 166, 24–27. [Google Scholar] [CrossRef]
- Goldie, I.; Nachemson, A. Synovial pH in rheumatoid knee-joints. I. The effect of synovectomy. Acta Orthop. Scand. 1969, 40, 634–641. [Google Scholar] [CrossRef]
- Ward, T.T.; Steigbigel, R.T. Acidosis of synovial fluid correlates with synovial fluid leukocytosis. Am. J. Med. 1978, 64, 933–936. [Google Scholar] [CrossRef]
- Geborek, P.; Saxne, T.; Pettersson, H.; Wollheim, F.A. Synovial fluid acidosis correlates with radiological joint destruction in rheumatoid arthritis knee joints. J. Rheumatol. 1989, 16, 468–472. [Google Scholar]
- Andersson, S.E.; Lexmüller, K.; Johansson, A.; Ekström, G.M. Tissue and intracellular pH in normal periarticular soft tissue and during different phases of antigen induced arthritis in the rat. J. Rheumatol. 1999, 26, 2018–2024. [Google Scholar]
- Ohyama, T.; Igarashi, K.; Kobayashi, H. Physiological role of the chaA gene in sodium and calcium circulations at a high pH in Escherichia coli. J. Bacteriol. 1994, 176, 4311–4315. [Google Scholar]
- Trchounian, A.; Kobayashi, H. Kup is the major K+ uptake system in Escherichiacoli upon hyper-osmotic stress at a low pH. FEBS Lett. 1999, 447, 144–148. [Google Scholar] [CrossRef]
- Fukamachi, T.; Saito, H.; Kakegawa, T.; Kobayashi, H. Different proteins are phosphorylated under acidic environments in Jurkat cells. Immunol. Lett. 2002, 82, 155–158. [Google Scholar] [CrossRef]
- Hirata, S.; Fukamachi, T.; Sakano, H.; Tarora, A.; Saito, H.; Kobayashi, H. Extracellular acidic environments induce phosphorylation of ZAP-70 in Jurkat T cells. Immunol. Lett. 2008, 115, 105–109. [Google Scholar] [CrossRef]
- Lao, Q.; Fukamachi, T.; Saito, H.; Kuge, O.; Nishijima, M.; Kobayashi, H. Requirement of an IκB-β COOH terminal region protein for acidic-adaptation in CHO cells. J. Cell Physiol. 2006, 207, 238–243. [Google Scholar] [CrossRef]
- Fukamachi, T.; Lao, Q.; Okamura, S.; Saito, H.; Kobayashi, H. CTIB (C-Terminus protein of IκB-β): a novel factor required for acidic adaptation. Adv. Exp. Med. Biol. 2006, 584, 219–228. [Google Scholar]
- Wang, X.; Hatatani, K.; Sun, Y.; Fukamachi, T.; Saito, H.; Kobayashi, H. TCR signaling via ZAP-70 induced by CD3 stimulation is more active under acidic conditions. J. Cell Sci. Ther. 2012, S16, 1. [Google Scholar]
- Souza, R.F.; Shewmake, K.; Pearson, S.; Sarosi, G.A., Jr.; Feagins, L.A.; Ramirez, R.D.; Terada, L.S.; Spechler, S.J. Acid increases proliferation via ERK and p38 MAPK-mediated increases in cyclooxygenase-2 in Barrett’s adenocarcinoma cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G743–G748. [Google Scholar] [CrossRef]
- Kato, Y.; Lambert, C.A.; Colige, A.C.; Mineur, P.; Noël, A.; Frankenne, F.; Foidart, J.M.; Baba, M.; Hata, R.; Miyazaki, K.; et al. Acidic extracellular pH induces matrix metalloproteinase-9 expression in mouse metastatic melanoma cells through the phospholipase D-mitogen-activated protein kinase signaling. J. Biol. Chem. 2005, 280, 10938–10944. [Google Scholar] [CrossRef]
- Ihnatko, R.; Kubes, M.; Takacova, M.; Sedlakova, O.; Sedlak, J.; Pastorek, J.; Kopacek, J.; Pastorekova, S. Extracellular acidosis elevates carbonic anhydrase IX in human glioblastoma cells via transcriptional modulation that does not depend on hypoxia. Int. J. Oncol. 2006, 29, 1025–1033. [Google Scholar]
- Xu, L.; Fukumura, D.; Jain, R.K. Acidic extracellular pH induces vascular endothelial growth factor (VEGF) in human glioblastoma cells via ERK1/2 MAPK signaling pathway: Mechanism of low pH-induced VEGF. J. Biol. Chem. 2002, 277, 11368–11374. [Google Scholar]
- Elias, A.P.; Dias, S. Microenvironment changes (in pH) affect VEGF alternative splicing. Cancer Microenviron. 2008, 1, 131–139. [Google Scholar] [CrossRef]
- Hjelmeland, A.B.; Wu, Q.; Heddleston, J.M.; Choudhary, G.S.; MacSwords, J.; Lathia, J.D.; McLendon, R.; Lindner, D.; Sloan, A.; Rich, J.N. Acidic stress promotes a glioma stem cell phenotype. Cell Death Differ. 2010, 18, 829–840. [Google Scholar]
- Tang, X.; Lucas, J.E.; Chen, J.L.; LaMonte, G.; Wu, J.; Wang, M.C.; Koumenis, C.; Chi, J.T. Functional interaction between responses to lactic acidosis and hypoxia regulates genomic transcriptional outputs. Cancer Res. 2012, 72, 491–502. [Google Scholar]
- Irizarry, R.A.; Bolstad, B.M.; Collin, F.; Cope, L.M.; Hobbs, B.; Speed, T.P. Summaries of Affymetrix GeneChip probe level data. Nucleic Acids Res. 2003, 31, e15. [Google Scholar]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics 2003, 4, 249–264. [Google Scholar] [CrossRef]
- Darnel, J.; Lodish, H.; Baltimore, D. Molecular Cell Biology; Scientific American Books Inc.: New York, NY, USA, 1986. [Google Scholar]
- Connor, K.M.; Hempel, N.; Nelson, K.K.; Dabiri, G.; Gamarra, A.; Belarmino, J.; van de Water, L.; Mian, B.M.; Melendez, J.A. Manganese superoxide dismutase enhances the invasive and migratory activity of tumor cells. Cancer Res. 2007, 67, 10260–10267. [Google Scholar] [CrossRef]
- Lao, Q.; Kuge, O.; Fukamachi, T.; Kakegawa, T.; Saito, H.; Nishijima, M.; Kobayashi, H. An IκB-β COOH terminal region protein is essential for the proliferation of CHO cells under acidic stress. J. Cell Physiol. 2005, 203, 186–192. [Google Scholar] [CrossRef]
- Dahl, C.A.; Schall, R.P.; He, H.L.; Cairns, J.S. Identification of a novel gene expressed in activated natural killer cells and T cells. J. Immunol. 1992, 148, 597–603. [Google Scholar]
- Kim, S.H.; Han, S.Y.; Azam, T.; Yoon, D.Y.; Dinarello, C.A. Interleukin-32: A cytokine and inducer of TNFα. Immunity 2005, 22, 131–142. [Google Scholar]
- Mun, S.H.; Kim, J.W.; Nah, S.S.; Ko, N.Y.; Lee, J.H.; Kim, J.D.; Kim, D.K.; Kim, H.S.; Choi, J.D.; Kim, S.H.; et al. Tumor necrosis factor α-induced interleukin-32 is positively regulated via the Syk/protein kinase Cδ/JNK pathway in rheumatoid synovial fibroblasts. Arthritis Rheum. 2009, 60, 678–685. [Google Scholar] [CrossRef]
- Joosten, L.A.; Netea, M.G.; Kim, S.H. IL-32, a proinflammatory cytokine in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2006, 103, 3298–3303. [Google Scholar] [CrossRef]
- Nishida, A.; Andoh, A.; Inatomi, O.; Fujiyama, Y. Interleukin-32 expression in the pancreas. J. Biol. Chem. 2009, 284, 17868–17876. [Google Scholar]
- Castella-Escola, J.; Ojcius, D.M.; LeBoulch, P.; Joulin, V.; Blouquit, Y.; Garel, M.C.; Valentin, C.; Rosa, R.; Climent-Romeo, F.; Cohen-Solal, M. Isolation and characterization of the gene encoding the muscle-specific isozyme of human phosphoglycerate mutase. Gene 1990, 91, 225–232. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fukamachi, T.; Ikeda, S.; Wang, X.; Saito, H.; Tagawa, M.; Kobayashi, H. Gene Expressions for Signal Transduction under Acidic Conditions. Genes 2013, 4, 65-85. https://doi.org/10.3390/genes4010065
Fukamachi T, Ikeda S, Wang X, Saito H, Tagawa M, Kobayashi H. Gene Expressions for Signal Transduction under Acidic Conditions. Genes. 2013; 4(1):65-85. https://doi.org/10.3390/genes4010065
Chicago/Turabian StyleFukamachi, Toshihiko, Syunsuke Ikeda, Xin Wang, Hiromi Saito, Masatoshi Tagawa, and Hiroshi Kobayashi. 2013. "Gene Expressions for Signal Transduction under Acidic Conditions" Genes 4, no. 1: 65-85. https://doi.org/10.3390/genes4010065