Mitochondrial Genome Sequencing Reveals orf463a May Induce Male Sterility in NWB Cytoplasm of Radish

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Mitochondrial DNA Sequencing and Genome Assembly
2.3. Genome Annotation
2.4. Construction of a Phylogenetic Tree, Prediction of Transmembrane Domains, and Sequence Comparisons
2.5. RNA Extraction, RT-PCR, Real-Time PCR, and Amplicon Sequencing
3. Results
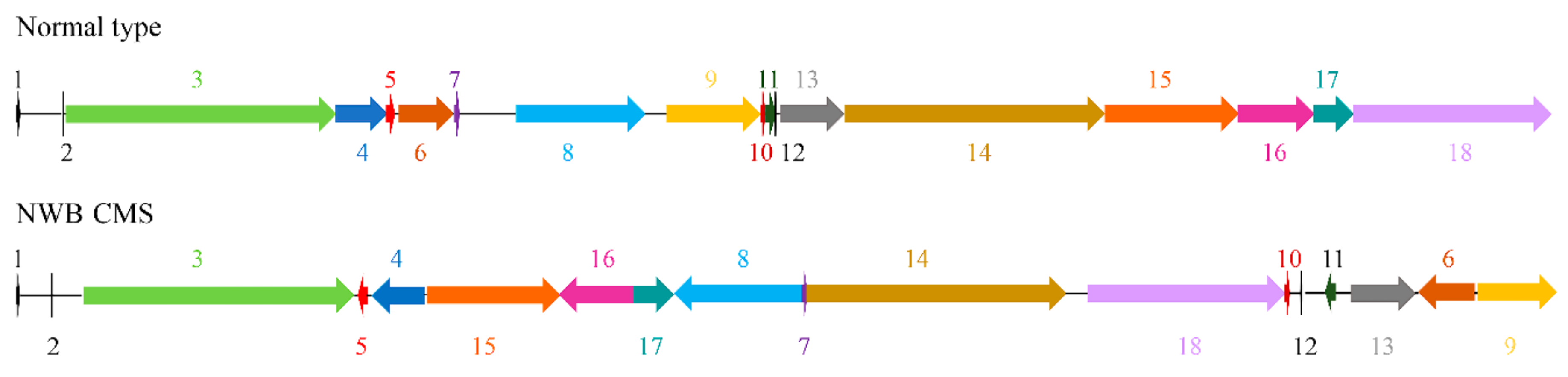
3.1. Mitochondrial Genome Organization of NWB CMS and Normal Radish
3.2. Identification of Sequence Variations in the Genes Encoding Known Proteins between the NWB CMS and Normal Mitochondrial Genomes
3.3. Screening for Specific orf Genes Responsible for the Male Sterility in NWB CMS
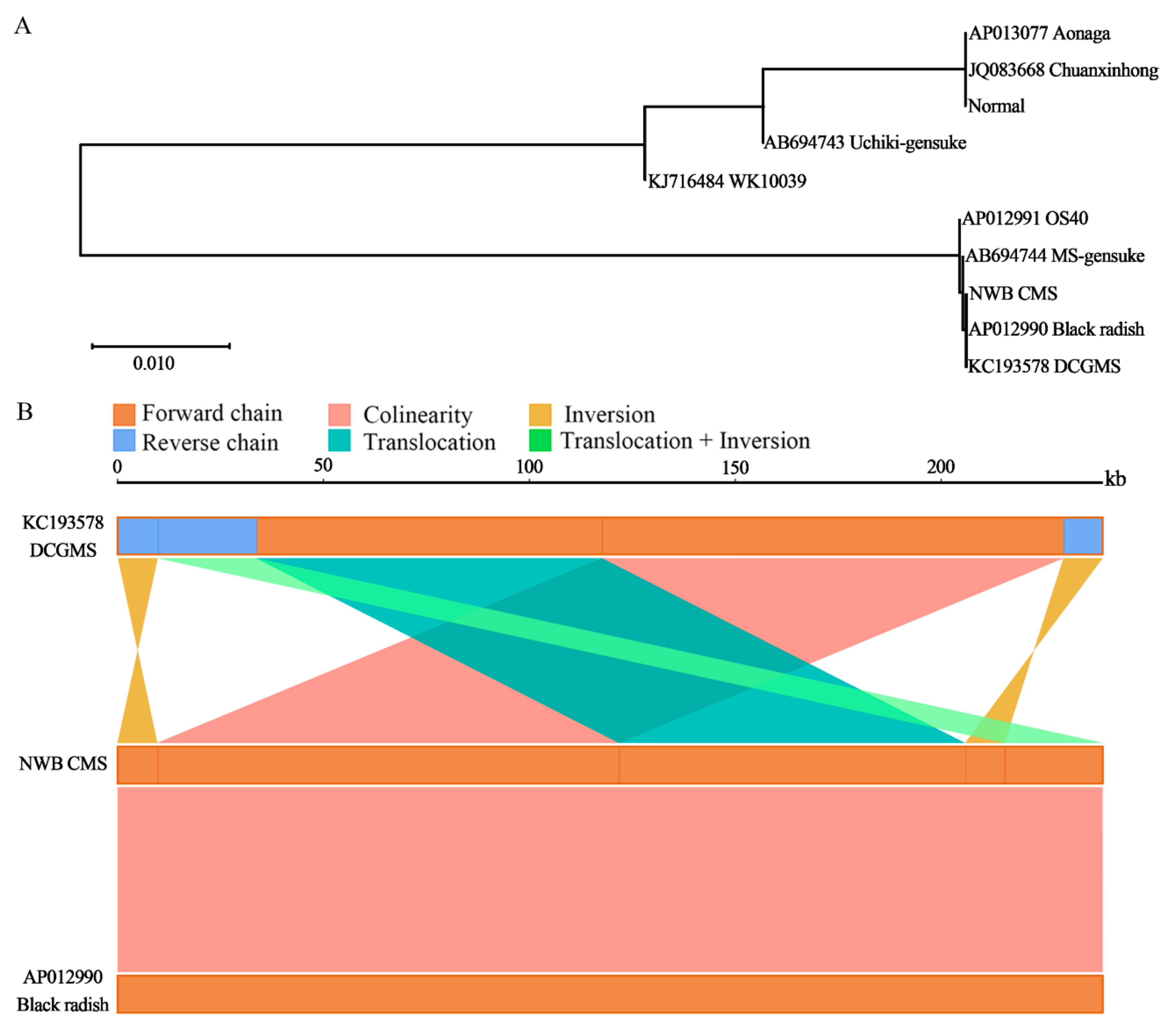
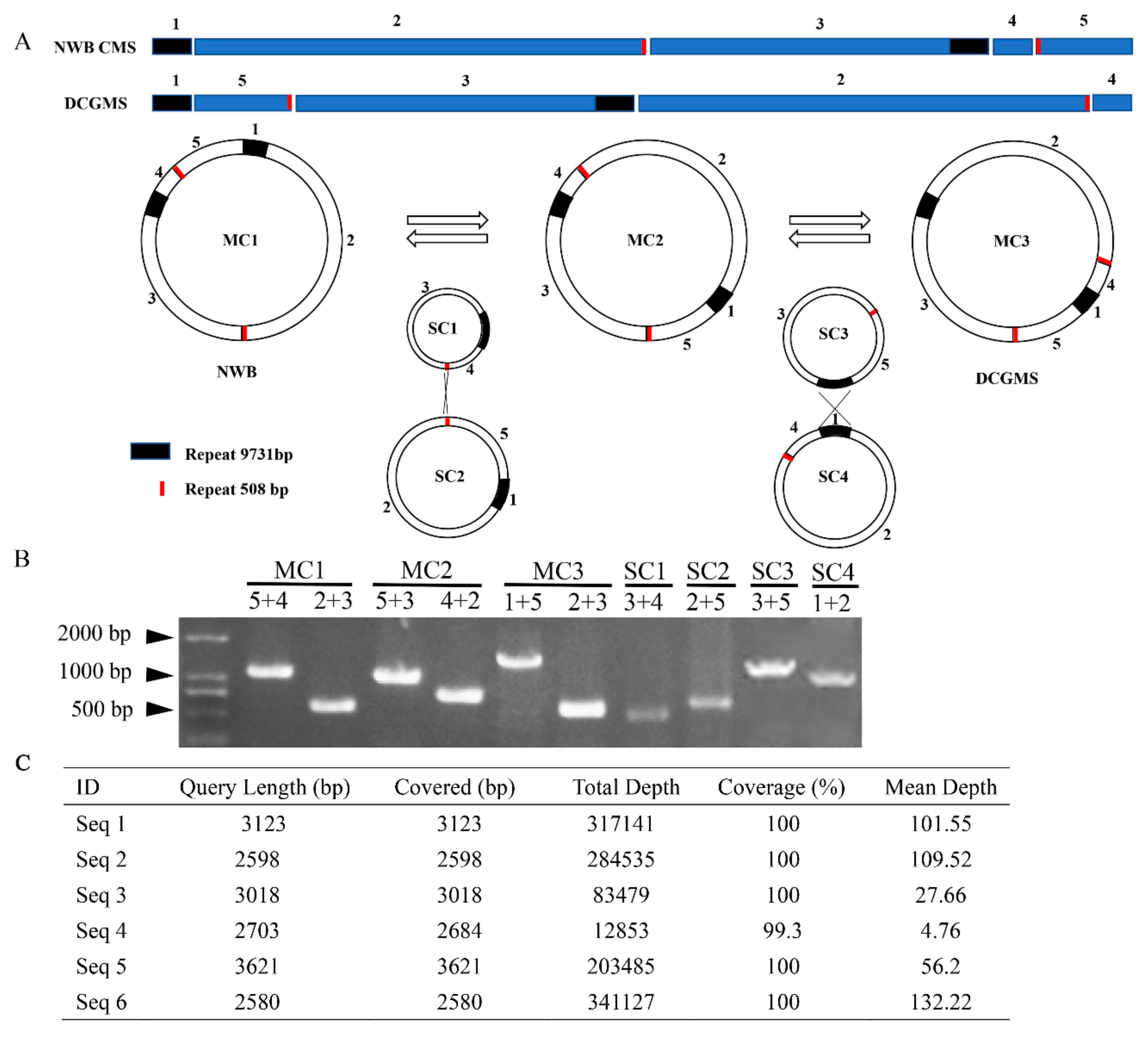
3.4. The NWB CMS and DCGMS Mitogenomes Are Coexisting Isomeric Forms of the Same Mitotype
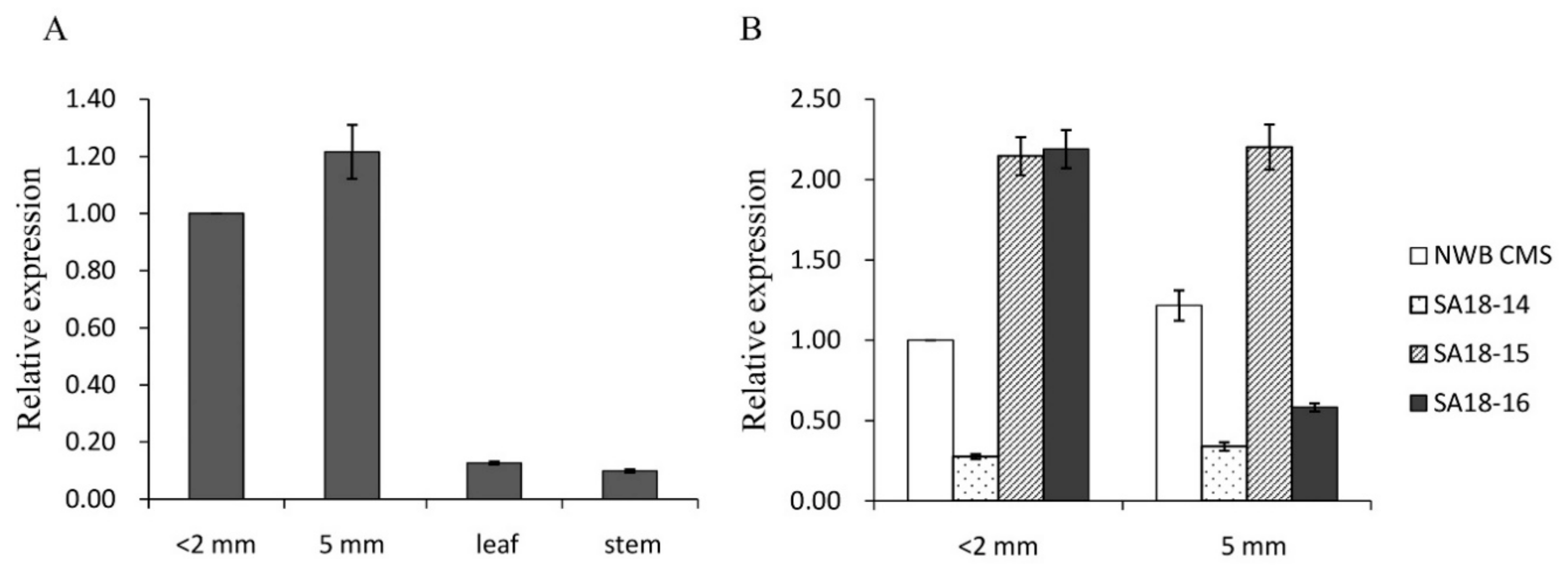
3.5. Expression of orf463a in the Absence or Presence of Rf Genes
3.6. Distribution of orf463a in Various Radish Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CMS | Cytoplasmic male sterility |
| Rf | Restorer of fertility |
| ORF | Open reading frame |
| DCGMS | Dongbu cytoplasmic and genic male sterility |
| NGS | Next-generation sequencing |
| SNP | Single nucleotide polymorphism |
| InDel | Insertion–deletion |
| tRNA | Transfer RNA |
| rRNA | Ribosome RNA |
References
- Curtis, I.S. Genetic engineering of radish: Current achievements and future goals. Plant Cell Rep. 2011, 30, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.R.; Bentolila, S. Interactions of mitochondrial and nuclear genes that affect male gametophyte development. Plant Cell 2004, 16 (Suppl. S1), S154–S169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, D.; Xu, H.; Liu, Z.; Guo, J.; Li, H.; Chen, L.; Fang, C.; Zhang, Q.; Bai, M.; Yao, N.; et al. A detrimental mitochondrial-nuclear interaction causes cytoplasmic male sterility in rice. Nat. Genet. 2013, 45, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Davila, J.; Arrieta-Montiel, M.; Wamboldt, Y.; Cao, J.; Hagmann, J.; Shedge, V.; Xu, Y.Z.; Weigel, D.; Mackenzie, S.A. Double-strand break repair processes drive evolution of the mitochondrial genome in Arabidopsis. BMC Biol. 2011, 9, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, M.; Kazama, T.; Murata, H.; Motomura, K.; Toriyama, K. Whole mitochondrial genome sequencing and transcriptional analysis to uncover an RT102-type cytoplasmic male sterility-associated candidate gene derived from Oryza rufipogon. Plant Cell Physiol. 2013, 54, 1560–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarria, R.; Lyznik, A.; Vallejos, C.E.; Mackenzie, S.A. A cytoplasmic male sterility-associated mitochondrial peptide in common bean is post-translationally regulated. Plant Cell 1998, 10, 1217–1228. [Google Scholar]
- Wise, R.P.; Bronson, C.R.; Schnable, P.S.; Horner, H.T. The genetics, pathology, and molecular biology of T-cytoplasm male sterility in maize. Adv. Agron. 1999, 65, 79–130. [Google Scholar]
- Koizuka, N.; Imai, R.; Fujimoto, H.; Hayakawa, T.; Kimura, Y.; Kohno-Murase, J.; Sakai, T.; Kawasaki, S.; Imamura, J. Genetic characterization of a pentatricopeptide repeat protein gene, orf687, that restores fertility in the cytoplasmic male-sterile Kosena radish. Plant J. 2003, 34, 407–415. [Google Scholar] [CrossRef]
- Kazama, T.; Nakamura, T.; Watanabe, M.; Sugita, M.; Toriyama, K. Suppression mechanism of mitochondrial ORF79 accumulation by Rf1 protein in BT-type cytoplasmic male sterile rice. Plant J. 2008, 55, 619–628. [Google Scholar] [CrossRef]
- Yamamoto, M.P.; Shinada, H.; Onodera, Y.; Komaki, C.; Mikami, T.; Kubo, T. A male sterility-associated mitochondrial protein in wildbeets causes pollen disruption in transgenic plants. Plant J. 2008, 54, 1027–1036. [Google Scholar] [CrossRef]
- Hanson, M.R. Plant mitochondrial mutations and male sterility. Annu. Rev. Genet. 1991, 25, 461–486. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, Y. Male Sterility and Fertility Restoration in Crops. Annu. Rev. Plant Biol. 2014, 65, 579–606. [Google Scholar] [CrossRef] [PubMed]
- Ogura, H. Studies on the new male-sterility in Japanese radish, with special reference to the utilization of this sterility towards the practical raising of hybrid seeds. Mem. Fac. Agric. Kagoshima Univ. 1968, 6, 39–78. [Google Scholar]
- Krishnasamy, S.; Makaroff, C.A. Organ-specific reduction in the abundance of a mitochondrial protein accompanies fertility restoration in cytoplasmic male-sterile radish. Plant Mol. Biol. 1994, 26, 935–946. [Google Scholar] [CrossRef] [PubMed]
- Bonhomme, S.; Budar, F.; Férault, M.; Pelletier, G. A 2.5 kb NcoI fragment of Ogura radish mitochondrial DNA is correlated with cytoplasmic male-sterility in Brassica cybrids. Curr. Genet. 1991, 19, 121–127. [Google Scholar] [CrossRef]
- Bonhomme, S.; Budar, F.; Lancelin, D.; Small, I.; Defrance, M.C.; Pelletier, G. Sequence and transcript analysis of the Nco2.5 Ogura-specific fragment correlated with cytoplasmic male sterility in Brassica cybrids. Mol. Gen. Genet. 1992, 235, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, H.; Terachi, T. Cytoplasmic male sterility and mitochondrial genome variations in radish. In The Radish Genome; Nishi, T., Kitashiba, H., Eds.; Springer Press: Cham, Switzerland, 2017; pp. 151–163. [Google Scholar]
- Nahm, S.H.; Lee, H.J.; Lee, S.W.; Joo, G.Y.; Harn, C.H.; Yang, S.G.; Min, B.W. Development of a molecular marker specific to a novel CMS line in radish (Raphanus sativus L.). Theor. Appl. Genet. 2005, 111, 1191–1200. [Google Scholar] [CrossRef]
- Lee, Y.; Park, S.; Lim, C.; Kim, H.; Lim, H.; Ahn, Y.; Sung, S.; Yoon, M.; Kim, S. Discovery of a novel cytoplasmic male-sterility and its restorer lines in radish (Raphanus sativus L.). Theor. Appl. Genet. 2008, 117, 905–913. [Google Scholar] [CrossRef]
- Park, J.Y.; Lee, Y.P.; Lee, J.; Choi, B.S.; Kim, S.; Yang, T.J. Complete mitochondrial genome sequence and identification of a candidate gene responsible for cytoplasmic male sterility in radish (Raphanus sativus L.) containing DCGMS cytoplasm. Theor. Appl. Genet. 2013, 126, 1763–1774. [Google Scholar] [CrossRef]
- Yamagishi, H.; Tanaka, Y.; Shiiba, S.; Hashimoto, A.; Fukunaga, A. Mitochondrial orf463 causing male sterility in radish is possessed by cultivars belonging to the ‘Niger’ group. Euphytica 2019, 215, 109. [Google Scholar] [CrossRef]
- Wang, Q.B.; Zhang, L.; Zheng, P. Genetic diversity and evolutionary relationship analyses within and among Raphanus species using EST-SSR markers. Mol. Breed. 2015, 35, 62. [Google Scholar] [CrossRef]
- Chen, J.; Guan, R.; Chang, S.; Du, T.; Zhang, H.; Xing, H. Substoichiometrically different mitotypes coexist in mitochondrial genomes of Brassica napus L. PLoS ONE 2011, 6, e17662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borgstrom, E.; Lundin, S.; Lundeberg, J. Large scale library generation for high throughput sequencing. PLoS ONE 2011, 6, e19119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dmitry, A.; Anton, K.; Jeffrey, S.M.; Pavel, A.P. HYBRIDSPADES: An algorithm for hybrid assembly of short and long reads. Bioinformatics 2016, 32, 7. [Google Scholar]
- Haas, B.J.; Salzberg, S.L.; Zhu, W.; Pertea, M.; Allen, J.E.; Orvis, J.; White, O.; Buell, C.R.; Wortman, J.R. Automated eukaryotic gene structure annotation using EvidenceModeler and the Program to Assemble Spliced Alignments. Genome Biol. 2008, 9, R7. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucl. Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Staerfeldt, H.H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucl. Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Wang, Q.B.; Zheng, P.J. Identification and Classification of different male sterile cytoplasms in radish. Acta Agric. Boreali Sin. 2014, 29, 125–129. [Google Scholar]
- Xu, Y.Y.; Zhu, X.W.; Gong, Y.Q.; Xu, L.; Wang, Y.; Liu, L.W. Evaluation of reference genes for gene expression studies in radish (Raphanus sativus L.) using quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2012, 424, 398–403. [Google Scholar] [CrossRef]
- Tanaka, Y.; Tsuda, M.; Yasumoto, K.; Yamagishi, H.; Terachi, T. A complete mitochondrial genome sequence of Ogura-type male-sterile cytoplasm and its comparative analysis with that of normal cytoplasm in radish (Raphanus sativus L.). BMC Genom. 2012, 13, 352. [Google Scholar] [CrossRef] [Green Version]
- Gabay-Laughnan, S.; Newton, K.J. Plant mitochondrial mutations. In Genomics of Chloroplasts and Mitochondria; Bock, R., Knoop, V., Eds.; Springer: Berlin, Germany, 2012; pp. 175–200. [Google Scholar]
- Chang, S.; Chen, J.; Wang, Y.; Gu, B.; He, J.; Chu, P.; Guan, R. The mitochondrial genome of Raphanus sativus and gene evolution of Cruciferous mitochondrial types. J. Genet. Genom. 2013, 40, 117–126. [Google Scholar] [CrossRef]
- Shen, D.; Sun, H.; Huang, M.; Zheng, Y.; Qiu, Y.; Li, X.; Fei, Z. Comprehensive analysis of expressed sequence tags from cultivated and wild radish (Raphanus spp.). BMC Genom. 2013, 14, 721. [Google Scholar] [CrossRef] [Green Version]
- Fu, T.D. Relationship between the origin and evolution of rapeseeds and the development of cytoplasmic male sterile “three lines”. Chin. J. Oil Crop Sci. 1989, 1, 7–10. [Google Scholar]
- Kim, S.; Lee, Y.; Lim, H.; Ahn, Y.; Sung, S. Identification of highly variable chloroplast sequences and development of cpDNA-based molecular markers that distinguish four cytoplasm types in radish (Raphanus sativus L.). Theor. Appl. Genet. 2009, 119, 189–198. [Google Scholar] [CrossRef]
- Kim, K.; Lee, Y.P.; Lim, H.; Han, T.; Sung, S.K.; Kim, S. Identification of Rfd1, a novel restorer-of-fertility locus for cytoplasmic male-sterility caused by DCGMS cytoplasm and development of simple PCR markers linked to the Rfd1 locus in radish (Raphanus sativus L.). Euphytica 2010, 175, 79–90. [Google Scholar] [CrossRef]
- Kazama, T.; Okuno, M.; Watari, Y.; Yanase, S.; Koizuka, C.; Tsuruta, Y.; Sugaya, H.; Toyoda, A.; Itoh, T.; Tsutsumi, N.; et al. Curing cytoplasmic male sterility via Talen-mediated mitochondrial genome editing. Nat. Plants 2019, 5, 722–730. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Position from Start Codon (on CDS) | Nucleotide Variation (Normal-NWB CMS) | Amino Acid Change (Normal-NWB CMS) |
|---|---|---|---|
| coxI | 117 | T-G | Synonymous |
| 207 | T-C | Synonymous | |
| rps3 | 551 | G-A | G-E |
| rps4 | 1052 | T-C | L-P |
| rps12 | 9 | A-G | Synonymous |
| rpl2 | 204 | G-A | Synonymous |
| 94 | A-C | N-H | |
| 93 | A-T | K-N | |
| 92 | A-T | K-I | |
| 91 | A-T | K-I | |
| 90 | G-T | Synonymous | |
| matR | 210 | T-C | Synonymous |
| 488 | C-T | T-I | |
| nad7 | 685 | G-A | V-I |
| Name | Length | Copy | Start–End | Junctions |
|---|---|---|---|---|
| Large repeat | 9731 | Copy 1 | 2–9732 | 1-2-Unique region |
| Copy 2 | 196,094–205,824 | 10-12-11-13 | ||
| R1 | 508 | Copy 1 | 121,189–121,696 | 8-7-14 |
| Copy 2 | 215,328–215,835 | Unique region-6 | ||
| R2 | 307 | Copy 1 | 51,926–52,232 | 3-Unique region |
| Copy 2 | 80,333–80,639 | 15 | ||
| R3 | 232 | Copy 1 | 164–395 | 1 |
| Copy 2 | 112,845–11,3076 | 8 | ||
| Copy 3 | 196,256–196,487 | 10 | ||
| R4 | 152 | Copy 1 | 121,312–121,463 | 7 |
| Copy 2 | 162,014–162,165 | Unique region | ||
| R5 | 147 | Copy 1 | 83,785–83,931 | 15-16 |
| Copy 2 | 95,246–95,392 | 16-17 | ||
| R6 | 141 | Copy 1 | 101,517–101,657 | 17-8 |
| Copy 2 | 165,607–165,747 | Unique region-18 | ||
| R7 | 98 | Copy 1 | 50,042–50,139 | 3 |
| Copy 2 | 121,275–121,372 | 8-7 | ||
| Copy 3 | 215652–215749 | Unique region | ||
| R8 | 87 | Copy 1 | 60,857–60,943 | 4 |
| Copy 2 | 117,566–117,652 | 8 | ||
| R9 | 81 | Copy 1 | 86–166 | 1 |
| Copy 2 | 125–205 | 1 | ||
| Copy 3 | 196,178–196,258 | 18-10 | ||
| Copy 4 | 196,217–196,297 | 18-10 |
| Features of the ORF | orf ID |
|---|---|
| In the unique region | orf84h, orf84g, orf82g, orf77f, orf306a, orf72f, orf71i, orf273a, orf124a, orf91a, orf463a, orf86b |
| In the non-unique region | orf148a, orf79e, orf85f, orf85e |
| Chimeric structure | orf306a, orf72f, orf71i, orf273a, orf91a |
| Transmembrane domain | orf84h, orf306a, orf273a, orf463a |
| Species | Total Accessions | orf463a |
|---|---|---|
| R. sativus var. hortensis Becker (East Asian big long radish) | 52 | 1 |
| R. sativus var. niger Kerner (Black Spanish radish) | 6 | 6 |
| R. sativus var. caudatus Hooker and Anderson (tail-podded radish or rat-tail radish) | 2 | 0 |
| R. sativus var. oleiferus Metzg (oil radish) | 1 | 0 |
| R. sativus var. sativus L. (European small radish) | 22 | 0 |
| R. sativus var. raphanistroides Makino (East Asian wild radish) | 3 | 0 |
| R. raphanistrum | 2 | 1 |
| R. maritimus | 4 | 1 |
| R. landra | 1 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Wang, Q.; Hao, W.; Li, J.; Qi, M.; Zhang, L. Mitochondrial Genome Sequencing Reveals orf463a May Induce Male Sterility in NWB Cytoplasm of Radish. Genes 2020, 11, 74. https://doi.org/10.3390/genes11010074
Wang Y, Wang Q, Hao W, Li J, Qi M, Zhang L. Mitochondrial Genome Sequencing Reveals orf463a May Induce Male Sterility in NWB Cytoplasm of Radish. Genes. 2020; 11(1):74. https://doi.org/10.3390/genes11010074
Chicago/Turabian StyleWang, Yanping, Qingbiao Wang, Wei Hao, Jianxia Li, Meixia Qi, and Li Zhang. 2020. "Mitochondrial Genome Sequencing Reveals orf463a May Induce Male Sterility in NWB Cytoplasm of Radish" Genes 11, no. 1: 74. https://doi.org/10.3390/genes11010074
APA StyleWang, Y., Wang, Q., Hao, W., Li, J., Qi, M., & Zhang, L. (2020). Mitochondrial Genome Sequencing Reveals orf463a May Induce Male Sterility in NWB Cytoplasm of Radish. Genes, 11(1), 74. https://doi.org/10.3390/genes11010074