Analysis of the Association Between TERC and TERT Genetic Variation and Leukocyte Telomere Length and Human Lifespan—A Follow-Up Study


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Laboratory Methods
2.3. Statistical Analysis
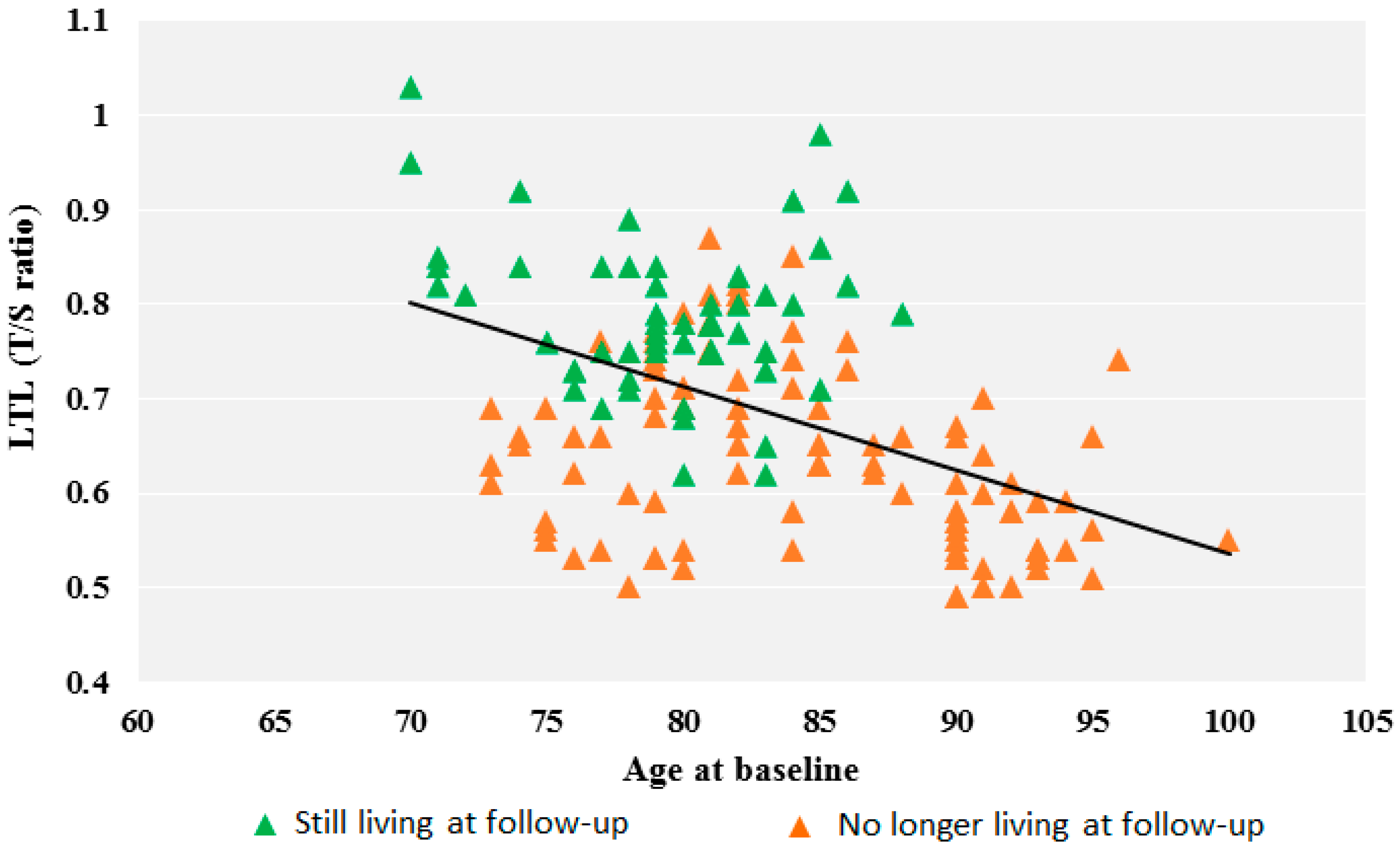
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.Y.; Collins, K. Telomere maintenance and disease. Lancet 2003, 362, 983–988. [Google Scholar] [CrossRef]
- Blackburn, E.H.; Greider, C.W.; Szostak, J.W. Telomeres and telomerase: The path from maize, Tetrahymena and yeast to human cancer and aging. Nat. Med. 2006, 12, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Cawthon, R.M.; Smith, K.R.; O’Brien, E.; Sivatchenko, A.; Kerber, R.A. Association between telomere length in blood and mortality in people aged 60 years or older. Lancet 2003, 361, 393–395. [Google Scholar] [CrossRef]
- Kimura, M.; Hjelmborg, J.V.B.; Gardner, J.P.; Bathum, L.; Brimacombe, M.; Lu, X.; Christiansen, L.; Vaupel, J.W.; Aviv, A.; Christensen, K. Telomere length and mortality: A study of leukocytes in elderly Danish twins. Am. J. Epidemiol. 2008, 167, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Lapham, K.; Kvale, M.N.; Lin, J.; Connell, S.; Croen, L.A.; Dispensa, B.P.; Fang, L.; Hesselson, S.; Hoffmann, T.J.; Iribarren, C.; et al. Automated Assay of Telomere Length Measurement and Informatics for 100,000 Subjects in the Genetic Epidemiology Research on Adult Health and Aging (GERA) Cohort. Genetics 2015, 200, 1061–1072. [Google Scholar] [CrossRef] [PubMed]
- Daniali, L.; Benetos, A.; Susser, E.; Kark, J.D.; Labat, C.; Kimura, M.; Desai, K.; Granick, M.; Aviv, A. Telomeres shorten at equivalent rates in somatic tissues of adults. Nat. Commun. 2013, 4, 1597. [Google Scholar] [CrossRef]
- Barrett, J.H.; Iles, M.M.; Dunning, A.M.; Pooley, K.A. Telomere length and common disease: Study design and analytical challenges. Hum. Genet. 2015, 134, 679–689. [Google Scholar] [CrossRef]
- Sanders, J.L.; Newman, A.B. Telomere length in epidemiology: A biomarker of aging, age-related disease, both, or neither? Epidemiol. Rev. 2013, 35, 112–131. [Google Scholar] [CrossRef]
- Needham, B.L.; Rehkopf, D.; Adler, N.; Gregorich, S.; Lin, J.; Blackburn, E.H.; Epel, E.S. Leukocyte telomere length and mortality in the National Health and Nutrition Examination Survey, 1999–2002. Epidemiology 2015, 26, 528–535. [Google Scholar] [CrossRef]
- Codd, V.; Nelson, C.P.; Albrecht, E.; Mangino, M.; Deelen, J.; Buxton, J.L.; Hottenga, J.J.; Fischer, K.; Esko, T.; Surakka, I.; et al. Identification of seven loci affecting mean telomere length and their association with disease. Nat. Genet. 2013, 45, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Scheller Madrid, A.; Rode, L.; Nordestgaard, B.G.; Bojesen, S.E. Short Telomere Length and Ischemic Heart Disease: Observational and Genetic Studies in 290 022 Individuals. Clin. Chem. 2016, 62, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Atzmon, G.; Cho, M.; Cawthon, R.M.; Budagov, T.; Katz, M.; Yang, X.; Siegel, G.; Bergman, A.; Huffman, D.M.; Schechter, C.B.; et al. Evolution in health and medicine Sackler colloquium: Genetic variation in human telomerase is associated with telomere length in Ashkenazi centenarians. Proc. Natl. Acad. Sci. USA 2010, 107 (Suppl. 1), 1710–1717. [Google Scholar] [CrossRef] [PubMed]
- Codd, V.; Mangino, M.; van der Harst, P.; Braund, P.S.; Kaiser, M.; Beveridge, A.J.; Rafelt, S.; Moore, J.; Nelson, C.; Soranzo, N.; et al. Common variants near TERC are associated with mean telomere length. Nat. Genet. 2010, 42, 197–199. [Google Scholar] [CrossRef] [PubMed]
- Njajou, O.T.; Blackburn, E.H.; Pawlikowska, L.; Mangino, M.; Damcott, C.M.; Kwok, P.-Y.; Spector, T.D.; Newman, A.B.; Harris, T.B.; Cummings, S.R.; et al. A common variant in the telomerase RNA component is associated with short telomere length. PLoS ONE 2010, 5, e13048. [Google Scholar] [CrossRef]
- Shen, Q.; Zhang, Z.; Yu, L.; Cao, L.; Zhou, D.; Kan, M.; Li, B.; Zhang, D.; He, L.; Liu, Y. Common variants near TERC are associated with leukocyte telomere length in the Chinese Han population. Eur. J. Hum. Genet. 2011, 19, 721–723. [Google Scholar] [CrossRef]
- Soerensen, M.; Thinggaard, M.; Nygaard, M.; Dato, S.; Tan, Q.; Hjelmborg, J.; Andersen-Ranberg, K.; Stevnsner, T.; Bohr, V.A.; Kimura, M.; et al. Genetic variation in TERT and TERC and human leukocyte telomere length and longevity: A cross-sectional and longitudinal analysis. Aging Cell 2012, 11, 223–227. [Google Scholar] [CrossRef]
- Concetti, F.; Lucarini, N.; Carpi, F.M.; Di Pietro, F.; Dato, S.; Capitani, M.; Nabissi, M.; Santoni, G.; Mignini, F.; Passarino, G.; et al. The functional VNTR MNS16A of the TERT gene is associated with human longevity in a population of Central Italy. Exp. Gerontol. 2013, 48, 587–592. [Google Scholar] [CrossRef]
- Crocco, P.; Barale, R.; Rose, G.; Rizzato, C.; Santoro, A.; De Rango, F.; Carrai, M.; Fogar, P.; Monti, D.; Biondi, F.; et al. Population-specific association of genes for telomere-associated proteins with longevity in an Italian population. Biogerontology 2015, 16, 353–364. [Google Scholar] [CrossRef]
- Wang, L.; Soria, J.-C.; Chang, Y.-S.; Lee, H.-Y.; Wei, Q.; Mao, L. Association of a functional tandem repeats in the downstream of human telomerase gene and lung cancer. Oncogene 2003, 22, 7123–7129. [Google Scholar] [CrossRef]
- Hofer, P.; Baierl, A.; Feik, E.; Führlinger, G.; Leeb, G.; Mach, K.; Holzmann, K.; Micksche, M.; Gsur, A. MNS16A tandem repeats minisatellite of human telomerase gene: A risk factor for colorectal cancer. Carcinogenesis 2011, 32, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, M.; Amininia, S.; Ebrahimi, M.; Hashemi, S.M.; Taheri, M.; Ghavami, S. Association between hTERT polymorphisms and the risk of breast cancer in a sample of Southeast Iranian population. BMC Res. Notes 2014, 7, 895. [Google Scholar] [CrossRef] [PubMed]
- Scarabino, D.; Broggio, E.; Gambina, G.; Pelliccia, F.; Corbo, R.M. Common variants of human TERT and TERC genes and susceptibility to sporadic Alzheimers disease. Exp. Gerontol. 2017, 88, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Maubaret, C.G.; Salpea, K.D.; Romanoski, C.E.; Folkersen, L.; Cooper, J.A.; Stephanou, C.; Li, K.W.; Palmen, J.; Hamsten, A.; Neil, A.; et al. Association of TERC and OBFC1 haplotypes with mean leukocyte telomere length and risk for coronary heart disease. PLoS ONE 2013, 8, e83122. [Google Scholar] [CrossRef] [PubMed]
- Cresta, M.; Gregorio, G. Il disegno dello studio LONCILE. Riv. Antropol. 2001, 79, 11–18. [Google Scholar]
- Corbo, R.M.; Pinto, A.; Scacchi, R. Gender-specific association between FSHR and PPARG common variants and human longevity. Rejuvenation Res. 2013, 16, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef]
- Cawthon, R.M. Telomere measurement by quantitative PCR. Nucleic Acids Res. 2002, 30, e47. [Google Scholar] [CrossRef]
- Scarabino, D.; Broggio, E.; Gambina, G.; Corbo, R.M. Leukocyte telomere length in mild cognitive impairment and Alzheimer’s disease patients. Exp. Gerontol. 2017, 98, 143–147. [Google Scholar] [CrossRef]
- Terwilliger, J.; Ott, J. Handbook for Human Genetic Linkage; John Hopkins University Press: Baltimore, MD, USA, 1994. [Google Scholar]
- Wang, L.; Wei, Q.; Wang, L.-E.; Aldape, K.D.; Cao, Y.; Okcu, M.F.; Hess, K.R.; El-Zein, R.; Gilbert, M.R.; Woo, S.Y.; et al. Survival prediction in patients with glioblastoma multiforme by human telomerase genetic variation. J. Clin. Oncol. 2006, 24, 1627–1632. [Google Scholar] [CrossRef]
- Jin, G.; Yoo, S.S.; Cho, S.; Jeon, H.-S.; Lee, W.-K.; Kang, H.-G.; Choi, Y.Y.; Choi, J.E.; Cha, S.-I.; Lee, E.B.; et al. Dual roles of a variable number of tandem repeat polymorphism in the TERT gene in lung cancer. Cancer Sci. 2011, 102, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Fu, G.; Wei, J.; Shi, J.; Pan, W.; Ren, Y.; Xiong, X.; Xia, J.; Shen, Y.; Li, H.; et al. The identification of two regulatory ESCC susceptibility genetic variants in the TERT-CLPTM1L loci. Oncotarget 2016, 7, 5495–5506. [Google Scholar] [CrossRef] [PubMed]
- Martin-Ruiz, C.M.; Baird, D.; Roger, L.; Boukamp, P.; Krunic, D.; Cawthon, R.; Dokter, M.M.; van der Harst, P.; Bekaert, S.; de Meyer, T.; et al. Reproducibility of Telomere Length Assessment--An International Collaborative Study. Int. J. Epidemiol. 2015, 44, 1749–1754. [Google Scholar] [CrossRef] [PubMed]
- Müezzinler, A.; Zaineddin, A.K.; Brenner, H. A systematic review of leukocyte telomere length and age in adults. Ageing Res. Rev. 2013, 12, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Mather, K.A.; Jorm, A.F.; Parslow, R.A.; Christensen, H. Is telomere length a biomarker of aging? A review. J. Gerontol. A Biol. Sci. Med. Sci. 2011, 66, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Mons, U.; Müezzinler, A.; Schöttker, B.; Dieffenbach, A.K.; Butterbach, K.; Schick, M.; Peasey, A.; De Vivo, I.; Trichopoulou, A.; Boffetta, P.; et al. Leukocyte Telomere Length and All-Cause, Cardiovascular Disease, and Cancer Mortality: Results from Individual-Participant-Data Meta-Analysis of 2 Large Prospective Cohort Studies. Am. J. Epidemiol. 2017, 185, 1317–1326. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhan, Y.; Pedersen, N.L.; Fang, F.; Hägg, S. Telomere Length and All-Cause Mortality: A Meta-analysis. Ageing Res. Rev. 2018, 48, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Liu, X.-R.; Reynolds, C.A.; Pedersen, N.L.; Hägg, S.; Clements, M.S. Leukocyte Telomere Length and All-Cause Mortality: A Between-Within Twin Study with Time-Dependent Effects Using Generalized Survival Models. Am. J. Epidemiol. 2018, 187, 2186–2191. [Google Scholar] [CrossRef]
- Honig, L.S.; Kang, M.S.; Cheng, R.; Eckfeldt, J.H.; Thyagarajan, B.; Leiendecker-Foster, C.; Province, M.A.; Sanders, J.L.; Perls, T.; Christensen, K.; et al. Heritability of telomere length in a study of long-lived families. Neurobiol. Aging 2015, 36, 2785–2790. [Google Scholar] [CrossRef]
- Bischoff, C.; Petersen, H.C.; Graakjaer, J.; Andersen-Ranberg, K.; Vaupel, J.W.; Bohr, V.A.; Kølvraa, S.; Christensen, K. No association between telomere length and survival among the elderly and oldest old. Epidemiology 2006, 17, 190–194. [Google Scholar] [CrossRef]
- Martin-Ruiz, C.M.; Gussekloo, J.; van Heemst, D.; von Zglinicki, T.; Westendorp, R.G.J. Telomere length in white blood cells is not associated with morbidity or mortality in the oldest old: A population-based study. Aging Cell 2005, 4, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Svensson, J.; Karlsson, M.K.; Ljunggren, Ö.; Tivesten, Å.; Mellström, D.; Movérare-Skrtic, S. Leukocyte telomere length is not associated with mortality in older men. Exp. Gerontol. 2014, 57, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflammaging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Rane, G.; Dai, X.; Shanmugam, M.K.; Arfuso, F.; Samy, R.P.; Lai, M.K.P.; Kappei, D.; Kumar, A.P.; Sethi, G. Ageing and the telomere connection: An intimate relationship with inflammation. Ageing Res. Rev. 2016, 25, 55–69. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gene/Genotype | Not Long-Lived | Long-Lived |
|---|---|---|
| TERC rs12696304 | ||
| G/G | 11 (12.4) | 5 (7.0) |
| G/C | 35 (39.3) | 31 (43.7) |
| C/C | 43 (48.3) | 35 (49.3) |
| TOTAL | 89 | 71 |
| p | 0.52 | |
| TERC rs3772190 | ||
| C/C | 56 (67.5) | 40 (58.8) |
| C/T | 22 (26.5) | 25 (36.8) |
| T/T | 5 (6.0) | 3 (4.4) |
| TOTAL | 83 | 68 |
| p | 0.40 | |
| TERC rs16847897 | ||
| C/C | 7 (8.6) | 8 (11.9) |
| C/G | 36 (44.4) | 34 (50.7) |
| G/G | 38 (46.9) | 25 (37.3) |
| TOTAL | 81 | 67 |
| p | 0.47 | |
| TERT VNTR | ||
| MNS16A | ||
| L/L L/S 1 | 32 (36.0) 42 (47.2) | 13 (18.8) 35 (50.7) |
| S/S 1 TOT | 15 (16.9) 89 | 21 (30.4) 69 |
| p | 0.018 | |
| TERT rs2853691 | ||
| A/A | 41 (45.1) | 48 (67.6) |
| A/G1 | 42 (46.2) | 22 (31.0) |
| G/G1 | 8 (8.8) | 1 (1.4) |
| TOT | 91 | 71 |
| p | 0. 004 | |
| TERT rs33954691 | ||
| C/C | 77 (85.6) | 62 (84.9) |
| C/T1 | 11 (12.2) | 9 (12.3) |
| T/T1 | 2 (2.2) | 2 (2.7) |
| TOTAL | 90 | 73 |
| p | 0.91 | |
| TERT rs2736098 | ||
| C/C | 56 (64.4) | 52 (72.2) |
| C/T1 | 30 (34.5) | 17 (23.6) |
| T/T1 | 1 (1.1) | 3 (4.2) |
| TOTAL | 87 | 72 |
| p | 0. 29 | |
| SNP/Genotypes | Age at Death |
|---|---|
| VNTR MNS16A | |
| L/L | 87.6 ± 6.0 (45) |
| L/S | 88.4 ± 5.4 (77) |
| S/S | 90.8 ± 6.2 (36) |
| p | 0.04 |
| rs2853691 | |
| A/A | 89.9 ± 5.7 (89) |
| A/G | 87.6 ± 5.7 (64) |
| G/G | 85.8 ± 3.7 (9) |
| p | 0.01 |
| VNTR MNS16A/rs2853691 Haplotype | Not Long-Lived | Long-Lived |
|---|---|---|
| L-A | 0.294 | 0.278 |
| L-G | 0.301 | 0.164 |
| S-A 1 | 0.397 | 0.548 |
| S-G 1 | 0.008 | 0.010 |
| p | 0.03 | |
| All Ages | Age 70–79 Years | Age 80–89 Years | ≥ 90 Years | |
|---|---|---|---|---|
| Total sample | 0.69 ± 0.12 (153) | 0.72 ± 0.12 (54) | 0.73 ± 0.10 (63) | 0.57 ± 0.06 (36) |
| No longer living at follow-up | 0.63 ± 0.09 (99) | 0.63 ± 0.08 (26) | 0.69 ± 0.09 (37) | 0.57 ± 0.06 (37) |
| Still living at follow-up | 0.79 ± 0.08 (54) | 0.80 ± 0.08 (28) | 0.77 ± 0.09 (26) | / |
| p1 | <0.0001 | <0.0001 | 0.0007 |
| L/L + G/G or A/G | L/S or S/S + A/A | |
|---|---|---|
| Not long-lived | 0.63 ± 0.08 (16) | 0.64 ± 0.11 (40) |
| Long-lived | 0.72 ± 0.10 (6) | 0.66 ± 0.06 (37) |
| p | 0.05 | 0.57 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scarabino, D.; Peconi, M.; Pelliccia, F.; Corbo, R.M. Analysis of the Association Between TERC and TERT Genetic Variation and Leukocyte Telomere Length and Human Lifespan—A Follow-Up Study. Genes 2019, 10, 82. https://doi.org/10.3390/genes10020082
Scarabino D, Peconi M, Pelliccia F, Corbo RM. Analysis of the Association Between TERC and TERT Genetic Variation and Leukocyte Telomere Length and Human Lifespan—A Follow-Up Study. Genes. 2019; 10(2):82. https://doi.org/10.3390/genes10020082
Chicago/Turabian StyleScarabino, Daniela, Martina Peconi, Franca Pelliccia, and Rosa Maria Corbo. 2019. "Analysis of the Association Between TERC and TERT Genetic Variation and Leukocyte Telomere Length and Human Lifespan—A Follow-Up Study" Genes 10, no. 2: 82. https://doi.org/10.3390/genes10020082