Inositol Polyphosphate Multikinase (IPMK), a Gene Coding for a Potential Moonlighting Protein, Contributes to Human Female Longevity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Population Sample
2.2. SNP Selection and Genotyping
2.3. Quality Control
2.4. Functional Parameters
2.4.1. Disability
2.4.2. Physical Performance
2.4.3. Cognitive Functioning
2.5. Statistical Methods
3. Results
3.1. Association with Longevity
3.1.1. Single SNP Analysis
3.1.2. Haplotype-Based Analysis
3.2. Association with Survival
3.3. Association with Functional Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ukraintseva, S.; Yashin, A.; Arbeev, K.; Kulminski, A.; Akushevich, I.; Wu, D.; Joshi, G.; Land, K.C.; Stallard, E. Puzzling role of genetic risk factors in human longevity: “Risk alleles” as pro-longevity variants. Biogerontology 2016, 17, 109–127. [Google Scholar] [CrossRef]
- Mooijaart, S.P.; van Heemst, D.; Noordam, R.; Rozing, M.P.; Wijsman, C.A.; de Craen, A.J.; Westendorp, R.G.; Beekman, M.; Slagboom, P.E. Polymorphisms associated with type 2 diabetes in familial longevity: The Leiden Longevity Study. Aging 2011, 3, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Sebastiani, P.; Riva, A.; Montano, M.; Pham, P.; Torkamani, A.; Scherba, E.; Benson, G.; Milton, J.N.; Baldwin, C.T.; Andersen, S.; et al. Whole genome sequences of a male and female supercentenarian, ages greater than 114 years. Front. Genet. 2011, 2, 90. [Google Scholar] [CrossRef]
- Lio, D.; Scola, L.; Crivello, A.; Colonna-Romano, G.; Candore, G.; Bonafè, M.; Cavallone, L.; Franceschi, C.; Caruso, C. Gender-specific association between -1082 IL-10 promoter polymorphism and longevity. Genes Immun. 2002, 3, 30–33. [Google Scholar] [CrossRef]
- Kulminski, A.M.; Culminskaya, I.; Arbeev, K.G.; Ukraintseva, S.V.; Arbeeva, L.; Yashin, A.I. Trade-off in the effect of the APOE gene on the ages at onset of cardiovascular disease and cancer across ages, gender, and human generations. Rejuvenation Res. 2013, 16, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.K.; Christensen, K.; Kragstrup, J. Lipid-lowering treatment to the end? A review of observational studies and RCTs on cholesterol and mortality in 80+-year olds. Age Ageing 2010, 39, 674–680. [Google Scholar] [CrossRef] [PubMed]
- van Vliet, P.; Oleksik, A.M.; van Heemst, D.; de Craen, A.J.M.; Westendorp, R.G.J. Dynamics of traditional metabolic risk factors associate with specific causes of death in old age. J. Gerontol. Ser. A 2010, 65, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Nie, C.; Min, J.; Liu, X.; Li, M.; Chen, H.; Xu, H.; Wang, M.; Ni, T.; Li, Y.; et al. Novel loci and pathways significantly associated with longevity. Sci. Rep. 2016, 6, 21243. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, C.J. Moonlighting proteins. Trends Biochem. Sci. 1999, 24, 8–11. [Google Scholar] [CrossRef]
- Jeffery, C.J. Molecular mechanisms for multitasking: recent crystal structures of moonlighting proteins. Curr. Opin. Struct. Biol. 2004, 14, 663–668. [Google Scholar] [CrossRef]
- Bosch, D.; Saiardi, A. Arginine transcriptional response does not require inositol phosphate synthesis. J. Biol. Chem. 2012, 287, 38347–38355. [Google Scholar] [CrossRef] [PubMed]
- Hatch, A.J.; Odom, A.R.; York, J.D. Inositol phosphate multikinase dependent transcriptional control. Adv. Biol. Regul. 2017, 64, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Saiardi, A.; Nagata, E.; Luo, H.R.; Sawa, A.; Luo, X.; Snowman, A.M.; Snyder, S.H. Mammalian inositol polyphosphate multikinase synthesizes inositol 1,4,5-trisphosphate and an inositol pyrophosphate. Proc. Natl. Acad. Sci. USA 2001, 98, 2306–2311. [Google Scholar] [CrossRef] [PubMed]
- Seacrist, C.D.; Blind, R.D. Crystallographic and kinetic analyses of human IPMK reveal disordered domains modulate ATP binding and kinase activity. Sci. Rep. 2018, 8, 16672. [Google Scholar] [CrossRef] [PubMed]
- Resnick, A.C.; Snowman, A.M.; Kang, B.N.; Hurt, K.J.; Snyder, S.H.; Saiardi, A. Inositol polyphosphate multikinase is a nuclear PI3-kinase with transcriptional regulatory activity. Proc. Natl. Acad. Sci. USA 2005, 102, 12783–12788. [Google Scholar] [CrossRef] [PubMed]
- Maag, D.; Maxwell, M.J.; Hardesty, D.A.; Boucher, K.L.; Choudhari, N.; Hanno, A.G.; Ma, J.F.; Snowman, A.S.; Pietropaoli, J.W.; Xu, R.; et al. Inositol polyphosphate multikinase is a physiologic PI3-kinase that activates Akt/PKB. Proc. Natl. Acad. Sci. USA 2011, 108, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, S.F.; Maag, D.; Maxwell, M.J.; Resnick, A.C.; Juluri, K.R.; Chakraborty, A.; Koldobskiy, M.A.; Cha, S.H.; Barrow, R.; et al. Amino acid signaling to mTOR mediated by inositol polyphosphate multikinase. Cell Metab. 2011, 2, 215–221. [Google Scholar] [CrossRef]
- Dailey, M.J.; Kim, S. Inositol polyphosphate multikinase: An emerging player for the central action of AMP-activated protein kinase. Biochem. Biophys. Res. Commun. 2012, 421, 1–3. [Google Scholar] [CrossRef]
- Kim, E.; Beon, J.; Lee, S.; Park, S.J.; Ahn, H.; Kim, M.G.; Park, J.E.; Kim, W.; Yuk, J.M.; Kang, S.J.; et al. Inositol polyphosphate multikinase promotes Toll-like receptor-induced inflammation by stabilizing TRAF6. Sci. Adv. 2017, 3, e1602296. [Google Scholar] [CrossRef]
- Xu, R.; Sen, N.; Paul, B.D.; Snowman, A.M.; Rao, F.; Vandiver, M.S.; Xu, J.; Snyder, S.H. Inositol polyphosphate multikinase is a coactivator of p53-mediated transcription and cell death. Sci. Signal. 2013, 6, ra22. [Google Scholar] [CrossRef]
- Kim, E.; Tyagi, R.; Lee, J.Y.; Park, J.; Kim, Y.R.; Beon, J.; Chen, P.Y.; Cha, J.Y.; Snyder, S.H.; Kim, S. Inositol polyphosphate multikinase is a coactivator for serum response factor-dependent induction of immediate early genes. Proc. Natl. Acad. Sci. USA 2013, 110, 19938–19943. [Google Scholar] [CrossRef] [PubMed]
- Saiardi, A.; Caffrey, J.J.; Snyder, S.H.; Shears, S.B. Inositol polyphosphate multikinase (ArgRIII) determines nuclear mRNA export in Saccharomyces cerevisiae. FEBS Lett. 2000, 468, 28–32. [Google Scholar] [CrossRef]
- Carmody, S.R.; Wente, S.R. mRNA nuclear export at a glance. J. Cell. Sci. 2009, 122, 1933–1937. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Beon, J.; Lee, S.; Park, J.; Kim, S. IPMK: A versatile regulator of nuclear signaling events. Adv. Biol. Regul. 2016, 61, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, J.S.; Wang, Y.; Schork, A.J.; Thompson, W.K.; Karch, C.M.; Cruchaga, C.; McEvoy, L.K.; Witoelar, A.; Chen, C.H.; Holland, D.; et al. Alzheimer’s Disease Neuroimaging Initiative. Association between Genetic Traits for Immune-Mediated Diseases and Alzheimer Disease. JAMA Neurol. 2016, 73, 691–697. [Google Scholar] [CrossRef] [PubMed]
- De Rango, F.; Montesanto, A.; Berardelli, M.; Mazzei, B.; Mari, V.; Lattanzio, F.; Corsonello, A.; Passarino, G. To grow old in southern Italy: a comprehensive description of the old and oldest old in Calabria. Gerontology 2011, 57, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Passarino, G.; Montesanto, A.; De Rango, F.; Garasto, S.; Berardelli, M.; Domma, F.; Mari, V.; Feraco, E.; Franceschi, C.; De Benedictis, G. A cluster analysis to define human aging phenotypes. Biogerontology 2007, 8, 283–290. [Google Scholar] [CrossRef]
- Crocco, P.; Saiardi, A.; Wilson, M.S.; Maletta, R.; Bruni, A.C.; Passarino, G.; Rose, G. Contribution of polymorphic variation of inositol hexakisphosphate kinase 3 (IP6K3) gene promoter to the susceptibility to late onset Alzheimer’s disease. Biochim. Biophys. Acta. 2016, 1862, 1766–1773. [Google Scholar] [CrossRef]
- Katz, S.; Ford, A.B.; Moskowitz, R.W.; Jackson, B.A.; Jaffe, M.W. Studies of illness in the aged. The index of Adl: A standardized measure of biological and psychosocial function. JAMA 1963, 185, 914–919. [Google Scholar] [CrossRef]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. “Mini-mental state”. A practical method for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Plink 1.9 home. Available online: https://www.cog-genomics.org/plink2 (accessed on 8 February 2019).
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed]
- The R Project for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 8 February 2019).
- Austad, S.N.; Fischer, K.E. Sex Differences in Lifespan. Cell Metab. 2016, 23, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Ostan, R.; Monti, D.; Gueresi, P.; Bussolotto, M.; Franceschi, C.; Baggio, G. Gender, aging and longevity in humans: An update of an intriguing/neglected scenario paving the way to a gender-specific medicine. Clin. Sci. (Lond) 2016, 130, 1711–1725. [Google Scholar] [CrossRef] [PubMed]
- Dato, S.; Rose, G.; Crocco, P.; Monti, D.; Garagnani, P.; Franceschi, C.; Passarino, G. The genetics of human longevity: An intricacy of genes, environment, culture and microbiome. Mech. Ageing Dev. 2017, 165, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Garasto, S.; Rose, G.; Derango, F.; Berardelli, M.; Corsonello, A.; Feraco, E.; Mari, V.; Maletta, R.; Bruni, A.; Franceschi, C.; et al. The study of APOA1, APOC3 and APOA4 variability in healthy ageing people reveals another paradox in the oldest old subjects. Ann. Hum. Genet. 2003, 67, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Altomare, K.; Greco, V.; Bellizzi, D.; Berardelli, M.; Dato, S.; De Rango, F.; Garasto, S.; Rose, G.; Feraco, E.; Mari, V.; et al. The allele (A) (-110) in the promoter region of the HSP70-1 gene is unfavorable to longevity in women. Biogerontology 2003, 4, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Deelen, J.; Beekman, M.; Uh, H.W.; Helmer, Q.; Kuningas, M.; Christiansen, L.; Kremer, D.; van der Breggen, R.; Suchiman, H.E.; Lakenberg, N.; et al. Genome-wide association study identifies a single major locus contributing to survival into old age; the APOE locus revisited. Aging Cell 2011, 10, 686–698. [Google Scholar] [CrossRef]
- Zeng, Y.; Nie, C.; Min, J.; Chen, H.; Liu, X.; Ye, R.; Chen, Z.; Bai, C.; Xie, E.; Yin, Z.; et al. Sex Differences in Genetic Associations With Longevity. JAMA Netw. Open 2018, 1, e181670. [Google Scholar] [CrossRef]
- Jeffery, C.J. Moonlighting proteins--an update. Mol Biosyst. 2009, 5, 345–350. [Google Scholar] [CrossRef]
- Houtkooper, R.H.; Williams, R.W.; Auwerx, J. Metabolic networks of longevity. Cell 2010, 142, 9–14. [Google Scholar] [CrossRef]
- Resnick, A.C.; Saiardi, A. Inositol polyphosphate multikinase: Metabolic architect of nuclear inositides. Front. Biosci. 2008, 13, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Saiardi, A.; Resnick, A.C.; Snowman, A.M.; Wendland, B.; Snyder, S.H. Inositol pyrophosphates regulate cell death and telomere length through phosphoinositide 3-kinase-related protein kinases. Proc. Natl. Acad. Sci. USA 2005, 102, 1911–1914. [Google Scholar] [CrossRef] [PubMed]
- Saiardi, A.; Sciambi, C.; McCaffery, J.M.; Wendland, B.; Snyder, S.H. Inositol pyrophosphates regulate endocytic trafficking. Proc. Natl. Acad. Sci. USA 2002, 99, 14206–14211. [Google Scholar] [CrossRef] [PubMed]
- Jadav, R.S.; Chanduri, M.V.; Sengupta, S.; Bhandari, R. Inositol pyrophosphate synthesis by inositol hexakisphosphate kinase 1 is required for homologous recombination repair. J. Biol Chem. 2013, 288, 3312–3321. [Google Scholar] [CrossRef] [PubMed]
- Onnebo, S.M.; Saiardi, A. Inositol pyrophosphates modulate hydrogen peroxide signalling. Biochem. J. 2009, 423, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Szijgyarto, Z.; Garedew, A.; Azevedo, C.; Saiardi, A. Influence of inositol pyrophosphates on cellular energy dynamics. Science 2011, 334, 802–805. [Google Scholar] [CrossRef] [PubMed]
- Wild, R.; Gerasimaite, R.; Jung, J.Y.; Truffault, V.; Pavlovic, I.; Schmidt, A.; Saiardi, A.; Jessen, H.J.; Poirier, Y.; Hothorn, M.; et al. Control of eukaryotic phosphate homeostasis by inositol polyphosphate sensor domains. Science 2016, 352, 986–990. [Google Scholar] [CrossRef]
- Azevedo, C.; Saiardi, A. Eukaryotic Phosphate Homeostasis: The Inositol Pyrophosphate Perspective. Trends Biochem. Sci. 2017, 42, 219–231. [Google Scholar] [CrossRef]
- Illies, C.; Gromada, J.; Fiume, R.; Leibiger, B.; Yu, J.; Juhl, K.; Yang, S.N.; Barma, D.K.; Falck, J.R.; Saiardi, A.; et al. Requirement of inositol pyrophosphates for full exocytotic capacity in pancreatic beta cells. Science 2007, 318, 1299–1302. [Google Scholar] [CrossRef]
- Zhu, Q.; Ghoshal, S.; Rodrigues, A.; Gao, S.; Asterian, A.; Kamenecka, T.M.; Barrow, J.C.; Chakraborty, A. Adipocyte-specific deletion of Ip6k1 reduces diet-induced obesity by enhancing AMPK-mediated thermogenesis. J. Clin. Invest. 2016, 126, 4273–4288. [Google Scholar] [CrossRef]
- Finkel, T. The metabolic regulation of aging. Nat. Med. 2015, 21, 1416–1423. [Google Scholar] [CrossRef]
- Dato, S.; Bellizzi, D.; Rose, G.; Passarino, G. The impact of nutrients on the aging rate: A complex interaction of demographic; environmental and genetic factors. Mech. Ageing Dev. 2016, 154, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Moritoh, Y.; Oka, M.; Yasuhara, Y.; Hozumi, H.; Iwachidow, K.; Fuse, H.; Tozawa, R. Inositol Hexakisphosphate Kinase 3 Regulates Metabolism and Lifespan in Mice. Sci. Rep. 2016, 6, 32072. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.; Chen, Y.; Ahima, R.S.; Kim, S.F. Convergence of IPMK and LKB1-AMPK signaling pathways on metformin action. Mol. Endocrinol. 2014, 28, 1186–1193. [Google Scholar] [CrossRef] [PubMed]
- Stanfel, M.N.; Shamieh, L.S.; Kaeberlein, M.; Kennedy, B.K. The TOR pathway comes of age. Biochim. Biophys. Acta 2009, 1790, 1067–1074. [Google Scholar] [CrossRef] [PubMed]
- Dato, S.; Hoxha, E.; Crocco, P.; Iannone, F.; Passarino, G.; Rose, G. Amino acids and amino acid sensing: Implication for aging and diseases. Biogerontology 2019, 20, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Sei, Y.; Zhao, X.; Forbes, J.; Szymczak, S.; Li, Q.; Trivedi, A.; Voellinger, M.; Joy, G.; Feng, J.; Whatley, M.; et al. A Hereditary Form of Small Intestinal Carcinoid Associated with a Germline Mutation in Inositol Polyphosphate Multikinase. Gastroenterology 2015, 149, 67–78. [Google Scholar] [CrossRef]
- Dovey, C.M.; Diep, J.; Clarke, B.P.; Hale, A.T.; McNamara, D.E.; Guo, H.; Brown, N.W., Jr.; Cao, J.Y.; Grace, C.R.; Gough, P.J.; et al. MLKL Requires the Inositol Phosphate Code to Execute Necroptosis. Mol. Cell. 2018, 70, 936–948.e7. [Google Scholar] [CrossRef]
- Tower, J. Programmed cell death in aging. Ageing Res. Rev. 2015, 23, 90–100. [Google Scholar] [CrossRef]
- Kander, M.C.; Cui, Y.; Liu, Z. Gender difference in oxidative stress: A new look at the mechanisms for cardiovascular diseases. J. Cell Mol. Med. 2017, 21, 1024–1032. [Google Scholar] [CrossRef]
- Gardner, M.; Bann, D.; Wiley, L.; Cooper, R.; Hardy, R.; Nitsch, D.; Martin-Ruiz, C.; Shiels, P.; Sayer, A.A.; Barbieri, M.; et al. Halcyon study team. Gender and telomere length: Systematic review and meta-analysis. Exp. Gerontol. 2014, 51, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.A.; Harrison, D.E.; Astle, C.M.; Fernandez, E.; Flurkey, K.; Han, M.; Javors, M.A.; Li, X.; Nadon, N.L.; Nelson, J.F.; et al. Rapamycin-mediated lifespan increase in mice is dose and sex dependent and metabolically distinct from dietary restriction. Aging Cell 2014, 13, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Roepstorff, C.; Thiele, M.; Hillig, T.; Pilegaard, H.; Richter, E.A.; Wojtaszewski, J.F.; Kiens, B. Higher skeletal muscle alpha2AMPK activation and lower energy charge and fat oxidation in men than in women during submaximal exercise. J. Physiol. 2006, 574, 125–138. [Google Scholar] [CrossRef] [PubMed]



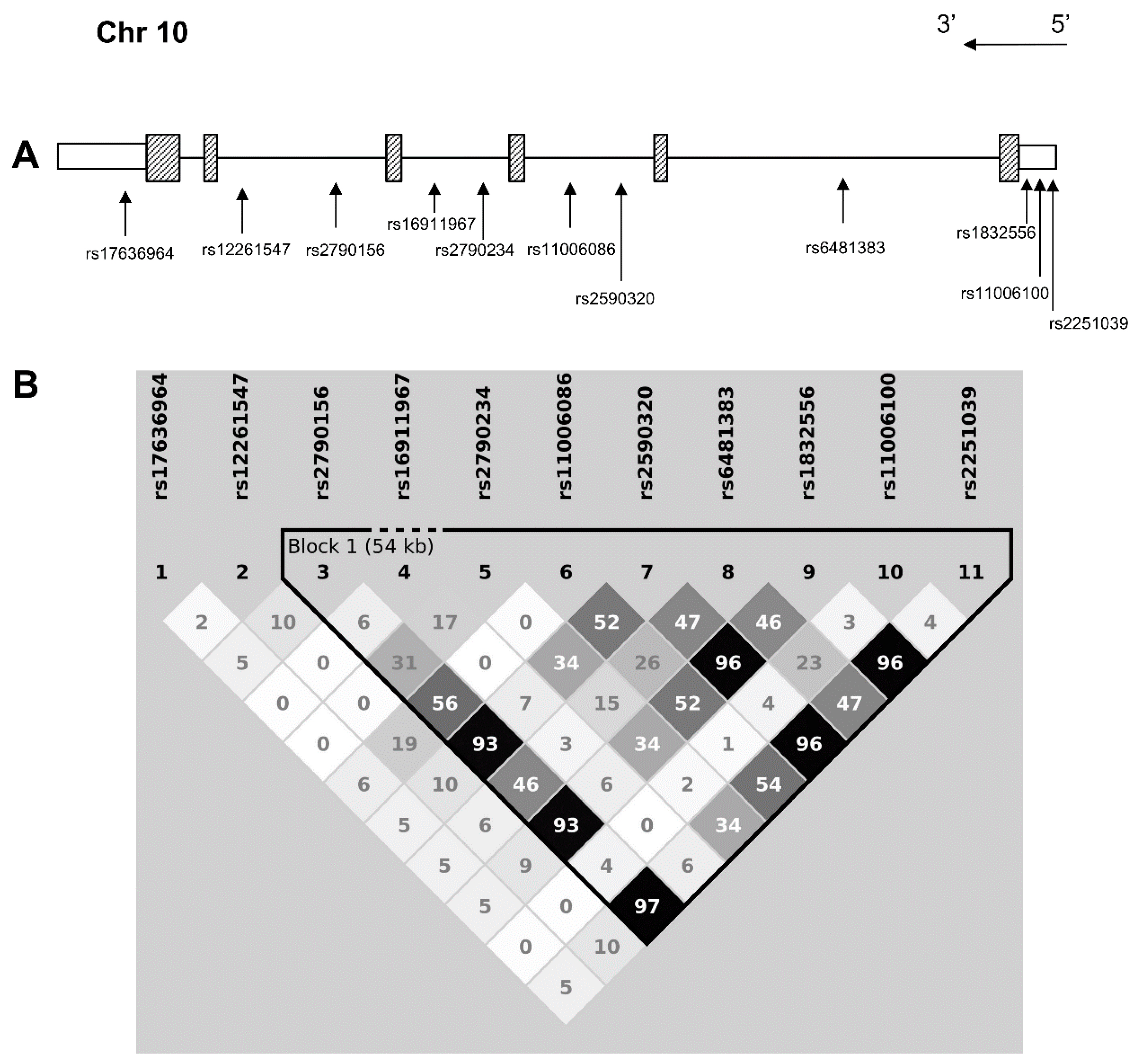{kind=link}
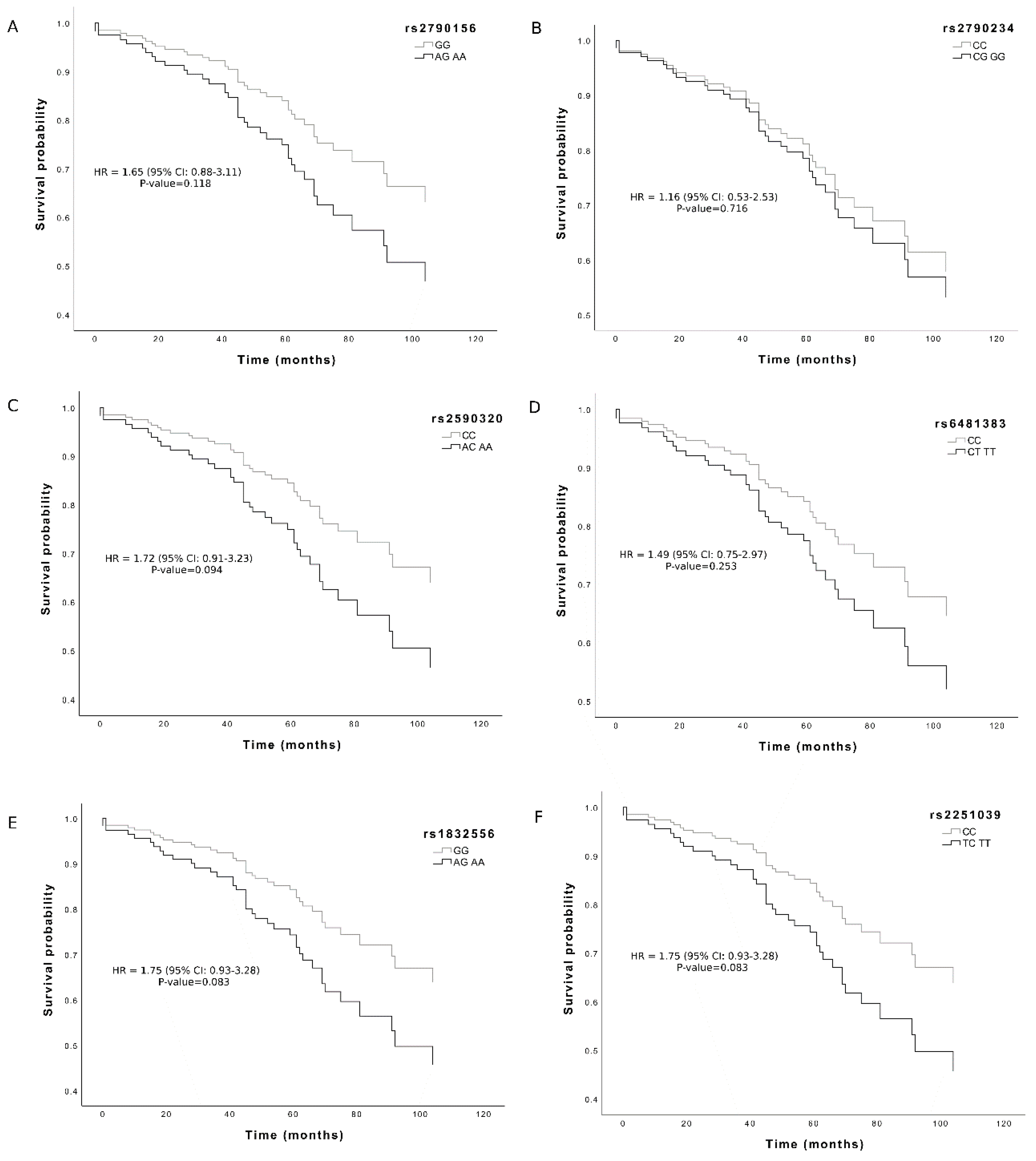{kind=link}
| Elderly Subjects | Long-Lived Subjects | |
|---|---|---|
| N (age) | 309 (74.06 ± 6.95) | 259 (96.92 ± 3.72) |
| Females % | 49.5% | 63.0% |
| Height (cm) | 160.6 (9.7) | 151.4 (9.5) |
| BMI | 26.9 (4.2) | 23.21 (4.1) |
| HG strength [Kg (SD)] | 21.89 (9.9) | 13.04 (6.4) |
| ADL* (% Disabled) | 17% | 69% |
| MMSE | 23.3 (5.4) | 14.0 (6.8) |
| (a) Females | |||
| SNP (Major/Minor Allele) | OR | 95% CI | p-value |
| rs17636964 (G/C) | 1.48 | 0.82–2.662 | 0.185 |
| rs12261547 (G/C) | 1.39 | 0.62–3.11 | 0.415 |
| rs2790156 (G/A) | 0.61 | 0.38–0.98 | 0.042 |
| rs16911967 (G/C) | 0.40 | 0.12–1.33 | 0.136 |
| rs2790234 (C/G) | 0.33 | 0.16–0.67 | 0.002 |
| rs11006086 (T/C) | 0.67 | 0.34–1.32 | 0.255 |
| rs2590320 (C/A) | 0.57 | 0.36–0.91 | 0.019 |
| rs6481383 (C/T) | 0.59 | 0.37–0.94 | 0.026 |
| rs1832556 (G/A) | 0.59 | 0.37–0.94 | 0.028 |
| rs11006100 (T/A) | 0.69 | 0.42–1.14 | 0.154 |
| rs2251039 (C/T) | 0.61 | 0.38–0.97 | 0.038 |
| (b) Males | |||
| SNP | OR | 95% CI | p-value |
| rs17636964 (G/C) | 0.98 | 0.50–1.94 | 0.974 |
| rs12261547 (G/C) | 0.52 | 0.16–1.66 | 0.272 |
| rs2790156 (G/A) | 0.79 | 0.46–1.35 | 0.397 |
| rs16911967 (G/C) | 2.08 | 0.54–7.96 | 0.283 |
| rs2790234 (C/G) | 0.98 | 0.49–1.94 | 0.959 |
| rs11006086 (T/C) | 0.76 | 0.31–1.85 | 0.550 |
| rs2590320 (C/A) | 0.81 | 0.48–1.37 | 0.436 |
| rs6481383 (C/T) | 1.10 | 0.65–1.87 | 0.713 |
| rs1832556 (G/A) | 0.81 | 0.47–1.37 | 0.436 |
| rs11006100 (T/A) | 1.73 | 0.98–3.04 | 0.056 |
| rs2251039 (C/T) | 0.78 | 0.46–1.34 | 0.377 |
| Haplotype | Frequency | Score | p-Value * |
|---|---|---|---|
| A-G-A-T-A-T | 0.067 | −2.897 | 0.002 |
| A-C-A-T-A-T | 0.138 | −0.668 | 0.483 |
| G-C-C-T-G-C | 0.161 | −0.353 | 0.715 |
| G-C-C-C-G-C | 0.616 | 2.155 | 0.024 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Rango, F.; Crocco, P.; Iannone, F.; Saiardi, A.; Passarino, G.; Dato, S.; Rose, G. Inositol Polyphosphate Multikinase (IPMK), a Gene Coding for a Potential Moonlighting Protein, Contributes to Human Female Longevity. Genes 2019, 10, 125. https://doi.org/10.3390/genes10020125
De Rango F, Crocco P, Iannone F, Saiardi A, Passarino G, Dato S, Rose G. Inositol Polyphosphate Multikinase (IPMK), a Gene Coding for a Potential Moonlighting Protein, Contributes to Human Female Longevity. Genes. 2019; 10(2):125. https://doi.org/10.3390/genes10020125
Chicago/Turabian StyleDe Rango, Francesco, Paolina Crocco, Francesca Iannone, Adolfo Saiardi, Giuseppe Passarino, Serena Dato, and Giuseppina Rose. 2019. "Inositol Polyphosphate Multikinase (IPMK), a Gene Coding for a Potential Moonlighting Protein, Contributes to Human Female Longevity" Genes 10, no. 2: 125. https://doi.org/10.3390/genes10020125