The Ufm1 Cascade

Abstract
:1. Introduction

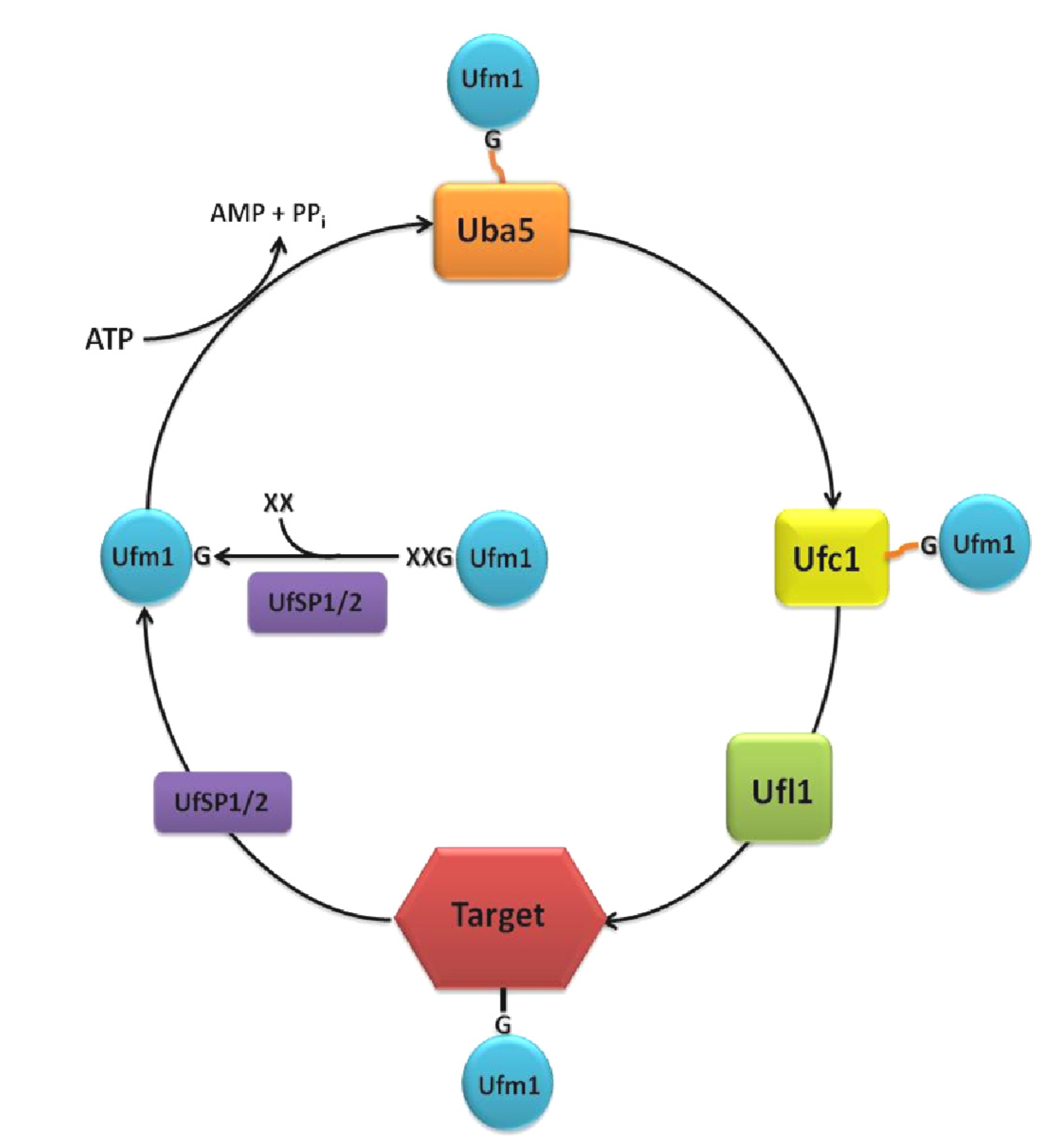{kind=link}
{kind=link}
| Proposed Names | Aliases |
|---|---|
| Ufm1 (ubiquitin-fold modifier 1) | C13orf20 |
| Uba5 (ubiquitin-like modifier activating enzyme 5) | UBE1DC1 |
| Ufc1 (ubiquitin-fold modifier conjugating enzyme 1) | HSPC155 |
| Ufl1 (Ufm1-specific ligase 1) | KIAA0776, NLBP, Maxer, RCAD |
| UfSP1 (Ufm1-specific protease 1) | Inactive Ufm1-specific protease 1 |
| UfSP2 (Ufm1-specific protease 2) | C4orf20 |
| Ufbp1 (Ufm1-binding protein 1 containing a PCI domain) | C20orf116, Dashurin, DDRGK domain containing 1 |
| Cdk5rap3 (CDK5 regulatory subunit-associated protein 3) | LZAP, protein HSF-27, IC53, C53, ARF-binding protein |
2. The Ufm1 Cascade

3. The Role of Ufm1 in ER Homeostasis
4. Ufm1 and Cell Differentiation
5. Ufm1 and Cell Cycle Control
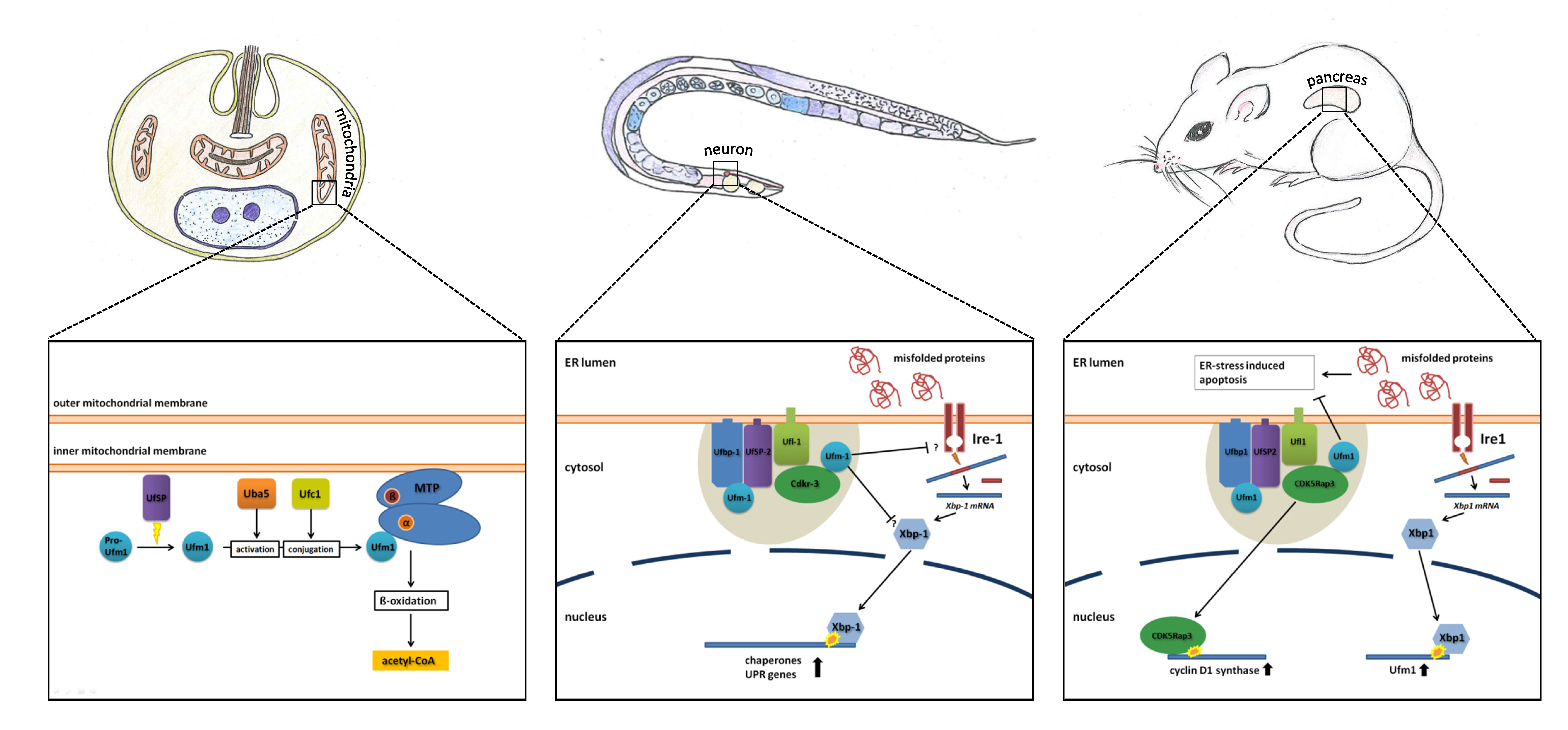
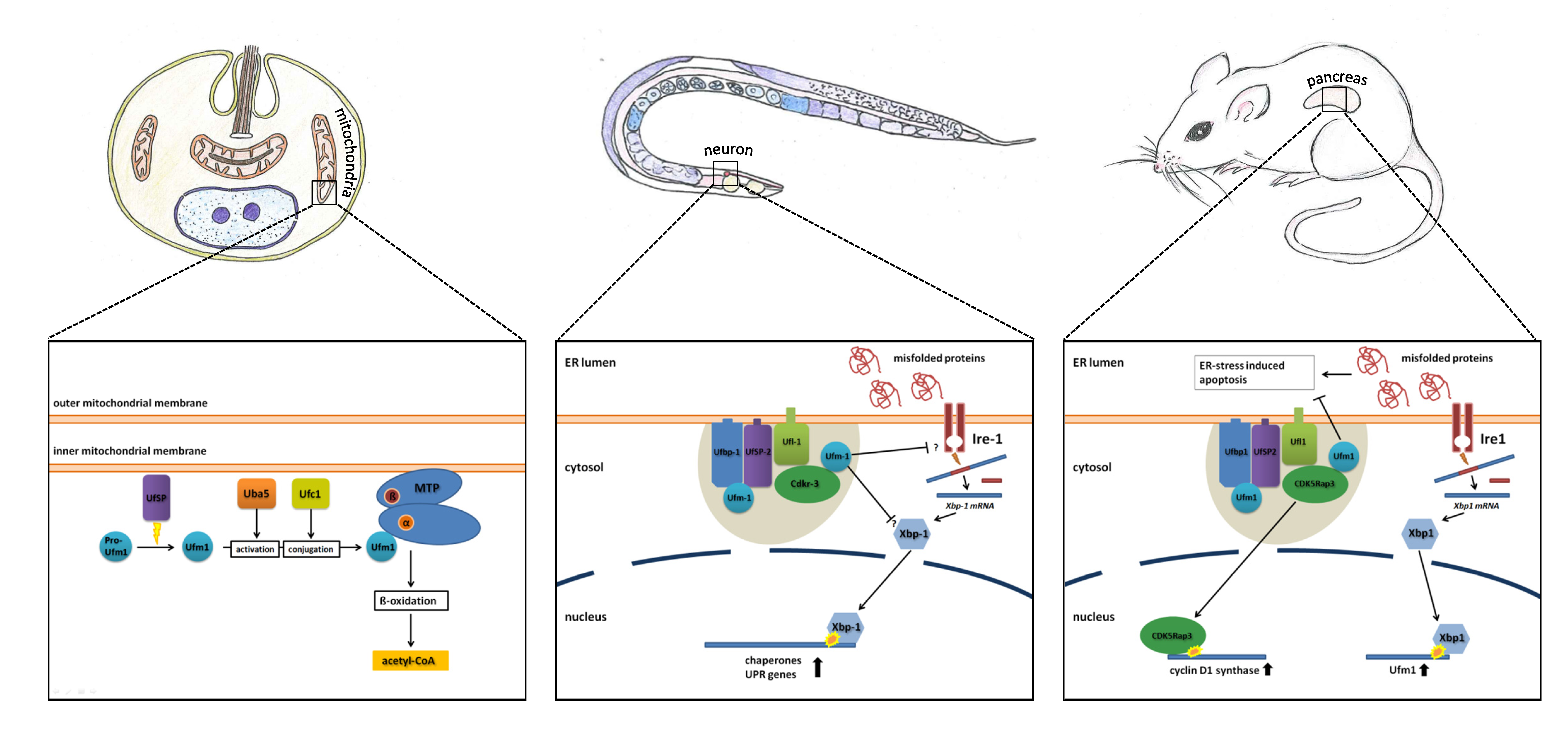
6. Ufm1 in Different Model Organisms
7. Diseases Associated with the Ufm1 Cascade
8. Conclusions
Abbreviations
| APG | Autophagy proteins |
| BiP | Binding immunoglobulin protein |
| CDK5rap3 | CDK5 regulatory subunit-associated protein 3 |
| CSN | COP9 signalosome |
| DUBs | Deubiquitinating enzymes |
| EIF3 | Eukaryotic translation initiation factor 3 |
| ER | Endoplasmic reticulum |
| FCCH | First catalytic cysteine half-domain |
| HECT | Homologous to the E6-AP carboxy terminus |
| I-KB | Inhibitor of κB |
| IRE1 | Inositol-requiring protein 1 |
| MTP | Mitochondrial trifunctional protein |
| NF-κB | Nuclear factor-κB |
| PCI | Proteasome, COP9, Initiation factor-3 |
| RING | Really interesting new gene |
| SCCH | Second catalytic cysteine half-domain |
| SNPs | Single nucleotide polymorphisms |
| SUMO | Small ubiquitin-like modifier |
| UBA | Ubiquitin-associated domain |
| Uba5 | Ubiquitin-like modifier activating enzyme 5 |
| UBL | Ubiquitin-like proteins |
| Ufbp | UFM1-binding protein 1 containing a PCI domain |
| Ufc1 | Ubiquitin-fold modifier conjugating enzyme 1 |
| Ufl1 | Ufm1-specific ligase 1 |
| Ufm1 | Ubiquitin-fold modifier 1 |
| UfSP | Ufm-specific peptidase |
| UPR | Unfolded protein response |
| XBP1 | X-box binding protein 1 |
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hochstrasser, M. Origin and function of ubiquitin-like proteins. Nature 2009, 458, 422–429. [Google Scholar] [CrossRef]
- Cort, J.R.; Chiang, Y.; Zheng, D.; Montelione, G.T.; Kennedy, M.A. Nmr structure of conserved eukaryotic protein zk652.3 from c. Elegans: A ubiquitin-like fold. Proteins 2002, 48, 733–736. [Google Scholar]
- Komatsu, M.; Chiba, T.; Tatsumi, K.; Iemura, S.; Tanida, I.; Okazaki, N.; Ueno, T.; Kominami, E.; Natsume, T.; Tanaka, K. A novel protein-conjugating system for ufm1, a ubiquitin-fold modifier. EMBO J. 2004, 23, 1977–1986. [Google Scholar] [CrossRef]
- Kang, S.H.; Kim, G.R.; Seong, M.; Baek, S.H.; Seol, J.H.; Bang, O.S.; Ovaa, H.; Tatsumi, K.; Komatsu, M.; Tanaka, K.; et al. Two novel ubiquitin-fold modifier 1 (ufm1)-specific proteases, ufsp1 and ufsp2. J. Biol. Chem. 2007, 282, 5256–5262. [Google Scholar] [CrossRef]
- Bacik, J.P.; Walker, J.R.; Ali, M.; Schimmer, A.D.; Dhe-Paganon, S. Crystal structure of the human ubiquitin-activating enzyme 5 (uba5) bound to atp: Mechanistic insights into a minimalistic e1 enzyme. J. Biol. Chem. 2010, 285, 20273–20280. [Google Scholar] [CrossRef]
- Zheng, M.; Gu, X.; Zheng, D.; Yang, Z.; Li, F.; Zhao, J.; Xie, Y.; Ji, C.; Mao, Y. Ube1dc1, an ubiquitin-activating enzyme, activates two different ubiquitin-like proteins. J. Cell. Biochem. 2008, 104, 2324–2334. [Google Scholar] [CrossRef]
- Ichimura, Y.; Kirisako, T.; Takao, T.; Satomi, Y.; Shimonishi, Y.; Ishihara, N.; Mizushima, N.; Tanida, I.; Kominami, E.; Ohsumi, M.; et al. A ubiquitin-like system mediates protein lipidation. Nature 2000, 408, 488–492. [Google Scholar] [CrossRef]
- Tatsumi, K.; Sou, Y.S.; Tada, N.; Nakamura, E.; Iemura, S.; Natsume, T.; Kang, S.H.; Chung, C.H.; Kasahara, M.; Kominami, E.; et al. A novel type of e3 ligase for the ufm1 conjugation system. J. Biol. Chem. 2010, 285, 5417–5427. [Google Scholar] [CrossRef]
- Mizushima, T.; Tatsumi, K.; Ozaki, Y.; Kawakami, T.; Suzuki, A.; Ogasahara, K.; Komatsu, M.; Kominami, E.; Tanaka, K.; Yamane, T. Crystal structure of ufc1, the ufm1-conjugating enzyme. Biochem. Biophys. Res. Commun. 2007, 362, 1079–1084. [Google Scholar] [CrossRef]
- Liu, G.; Aramini, J.; Atreya, H.S.; Eletsky, A.; Xiao, R.; Acton, T.; Ma, L.; Montelione, G.T.; Szyperski, T. Gft nmr based resonance assignment for the 21 kda human protein ufc1. J. Biomol. NMR 2005, 32, 261. [Google Scholar]
- Liu, G.; Forouhar, F.; Eletsky, A.; Atreya, H.S.; Aramini, J.M.; Xiao, R.; Huang, Y.J.; Abashidze, M.; Seetharaman, J.; Liu, J.; et al. Nmr and x-ray structures of human e2-like ubiquitin-fold modifier conjugating enzyme 1 (ufc1) reveal structural and functional conservation in the metazoan ufm1-uba5-ufc1 ubiquination pathway. J. Struct. Funct. Genomics 2009, 10, 127–136. [Google Scholar] [CrossRef]
- Rotin, D.; Kumar, S. Physiological functions of the hect family of ubiquitin ligases. Nat. Rev. Mol. Cell Biol. 2009, 10, 398–409. [Google Scholar] [CrossRef]
- Neziri, D.; Ilhan, A.; Maj, M.; Majdic, O.; Baumgartner-Parzer, S.; Cohen, G.; Base, W.; Wagner, L. Cloning and molecular characterization of dashurin encoded by c20orf116, a pci-domain containing protein. Biochim. Biophys. Acta 2010, 1800, 430–438. [Google Scholar] [CrossRef]
- Pick, E.; Hofmann, K.; Glickman, M.H. Pci complexes: Beyond the proteasome, csn, and eif3 troika. Mol. Cell 2009, 35, 260–264. [Google Scholar] [CrossRef]
- Ha, B.H.; Jeon, Y.J.; Shin, S.C.; Tatsumi, K.; Komatsu, M.; Tanaka, K.; Watson, C.M.; Wallis, G.; Chung, C.H.; Kim, E.E. Structure of ubiquitin-fold modifier 1-specific protease ufsp2. J. Biol. Chem. 2011, 286, 10248–10257. [Google Scholar] [CrossRef]
- Wu, J.; Lei, G.; Mei, M.; Tang, Y.; Li, H. A novel c53/lzap-interacting protein regulates stability of c53/lzap and ddrgk domain-containing protein 1 (ddrgk1) and modulates nf-kappab signaling. J. Biol. Chem. 2010, 285, 15126–15136. [Google Scholar] [CrossRef]
- Mak, G.W.; Lai, W.L.; Zhou, Y.; Li, M.; Ng, I.O.; Ching, Y.P. Cdk5rap3 is a novel repressor of p14arf in hepatocellular carcinoma cells. PLoS One 2012, 7, e42210. [Google Scholar]
- Gusarova, G.A.; Wang, I.C.; Major, M.L.; Kalinichenko, V.V.; Ackerson, T.; Petrovic, V.; Costa, R.H. A cell-penetrating arf peptide inhibitor of foxm1 in mouse hepatocellular carcinoma treatment. J. Clin. Investig. 2007, 117, 99–111. [Google Scholar] [CrossRef]
- Xiao, G.; Fu, J. Nf-kappab and cancer: A paradigm of yin-yang. Am. J. Cancer Res. 2011, 1, 192–221. [Google Scholar]
- Whiteside, S.T.; Israel, A. I kappa b proteins: Structure, function and regulation. Semin. Cancer Biol. 1997, 8, 75–82. [Google Scholar] [CrossRef]
- Chen, C.; Itakura, E.; Weber, K.P.; Hegde, R.S.; de Bono, M. An er complex of odr-4 and odr-8/ufm1 specific protease 2 promotes gpcr maturation by a ufm1-independent mechanism. PLoS Genet. 2014, 10, e1004082. [Google Scholar] [CrossRef]
- Azfer, A.; Niu, J.; Rogers, L.M.; Adamski, F.M.; Kolattukudy, P.E. Activation of endoplasmic reticulum stress response during the development of ischemic heart disease. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H1411–H1420. [Google Scholar] [CrossRef]
- Lemaire, K.; Moura, R.F.; Granvik, M.; Igoillo-Esteve, M.; Hohmeier, H.E.; Hendrickx, N.; Newgard, C.B.; Waelkens, E.; Cnop, M.; Schuit, F. Ubiquitin fold modifier 1 (ufm1) and its target ufbp1 protect pancreatic beta cells from er stress-induced apoptosis. PLoS One 2011, 6, e18517. [Google Scholar] [CrossRef]
- Hertel, P.; Daniel, J.; Stegehake, D.; Vaupel, H.; Kailayangiri, S.; Gruel, C.; Woltersdorf, C.; Liebau, E. The ubiquitin-fold modifier 1 (ufm1) cascade of Caenorhabditis elegans. J. Biol. Chem. 2013, 288, 10661–10671. [Google Scholar] [CrossRef]
- Kwon, J.; Cho, H.J.; Han, S.H.; No, J.G.; Kwon, J.Y.; Kim, H. A novel lzap-binding protein, nlbp, inhibits cell invasion. J. Biol. Chem. 2010, 285, 12232–12240. [Google Scholar]
- Gannavaram, S.; Connelly, P.S.; Daniels, M.P.; Duncan, R.; Salotra, P.; Nakhasi, H.L. Deletion of mitochondrial associated ubiquitin fold modifier protein ufm1 in Leishmania donovani results in loss of beta-oxidation of fatty acids and blocks cell division in the amastigote stage. Mol. Microbiol. 2012, 86, 187–198. [Google Scholar] [CrossRef]
- Shiwaku, H.; Yoshimura, N.; Tamura, T.; Sone, M.; Ogishima, S.; Watase, K.; Tagawa, K.; Okazawa, H. Suppression of the novel er protein maxer by mutant ataxin-1 in bergman glia contributes to non-cell-autonomous toxicity. EMBO J. 2010, 29, 2446–2460. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Wu, J.; Lei, G.; Li, H. Transcriptional regulation of the ufm1 conjugation system in response to disturbance of the endoplasmic reticulum homeostasis and inhibition of vesicle trafficking. PLoS One 2012, 7, e48587. [Google Scholar] [CrossRef]
- Mori, K. Signalling pathways in the unfolded protein response: Development from yeast to mammals. J. Biochem. 2009, 146, 743–750. [Google Scholar] [CrossRef]
- Dudek, M. Investigations of the Ufm1 Pathway and Its Association with a familial Form of Hip Dysplasia. Ph.D. Thesis, University of Manchester, Manchester, UK, 25 September 2013. [Google Scholar]
- Lee, Y.J.; Johnson, K.R.; Hallenbeck, J.M. Global protein conjugation by ubiquitin-like-modifiers during ischemic stress is regulated by micrornas and confers robust tolerance to ischemia. PLoS One 2012, 7, e47787. [Google Scholar]
- Tatsumi, K.; Yamamoto-Mukai, H.; Shimizu, R.; Waguri, S.; Sou, Y.S.; Sakamoto, A.; Taya, C.; Shitara, H.; Hara, T.; Chung, C.H.; et al. The ufm1-activating enzyme uba5 is indispensable for erythroid differentiation in mice. Nat. Commun. 2011, 2, 181. [Google Scholar] [CrossRef]
- Kim, C.H.; Nam, H.S.; Lee, E.H.; Han, S.H.; Cho, H.J.; Chung, H.J.; Lee, N.S.; Choi, S.J.; Kim, H.; Ryu, J.S.; et al. Overexpression of a novel regulator of p120 catenin, nlbp, promotes lung adenocarcinoma proliferation. Cell Cycle 2013, 12, 2443–2453. [Google Scholar] [CrossRef]
- Xi, P.; Ding, D.; Zhou, J.; Wang, M.; Cong, Y.S. Ddrgk1 regulates nf-kappab activity by modulating ikappabalpha stability. PLoS One 2013, 8, e64231. [Google Scholar]
- Gannavaram, S.; Debrabant, A. Programmed cell death in leishmania: Biochemical evidence and role in parasite infectivity. Front. Cell. Infect. Microbiol. 2012, 2, 95. [Google Scholar]
- Gannavaram, S.; Sharma, P.; Duncan, R.C.; Salotra, P.; Nakhasi, H.L. Mitochondrial associated ubiquitin fold modifier-1 mediated protein conjugation in Leishmania donovani. PLoS One 2011, 6, e16156. [Google Scholar]
- Gannavaram, S.; Davey, S.; Lakhal-Naouar, I.; Duncan, R.; Nakhasi, H.L. Deletion of ubiquitin fold modifier protein ufm1 processing peptidase ufsp in L. donovani abolishes ufm1 processing and alters pathogenesis. PLoS Negl. Trop. Dis. 2014, 8, e2707. [Google Scholar] [CrossRef]
- Maran, S.; Lee, Y.Y.; Xu, S.; Rajab, N.S.; Hasan, N.; Syed Abdul Aziz, S.H.; Majid, N.A.; Zilfalil, B.A. Gastric precancerous lesions are associated with gene variants in Helicobacter pylori-susceptible ethnic malays. World J. Gastroenterol. 2013, 19, 3615–3622. [Google Scholar] [CrossRef]
- Rubio, M.D.; Wood, K.; Haroutunian, V.; Meador-Woodruff, J.H. Dysfunction of the ubiquitin proteasome and ubiquitin-like systems in schizophrenia. Neuropsychopharmacology 2013, 38, 1910–1920. [Google Scholar] [CrossRef]
- Back, S.H.; Kaufman, R.J. Endoplasmic reticulum stress and type 2 diabetes. Annu. Rev. Biochem. 2012, 81, 767–793. [Google Scholar] [CrossRef]
- Mitsuda, T.; Omi, T.; Tanimukai, H.; Sakagami, Y.; Tagami, S.; Okochi, M.; Kudo, T.; Takeda, M. Sigma-1rs are upregulated via perk/eif2alpha/atf4 pathway and execute protective function in ER stress. Biochem. Biophys. Res. Commun. 2011, 415, 519–525. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Daniel, J.; Liebau, E. The Ufm1 Cascade. Cells 2014, 3, 627-638. https://doi.org/10.3390/cells3020627
Daniel J, Liebau E. The Ufm1 Cascade. Cells. 2014; 3(2):627-638. https://doi.org/10.3390/cells3020627
Chicago/Turabian StyleDaniel, Jens, and Eva Liebau. 2014. "The Ufm1 Cascade" Cells 3, no. 2: 627-638. https://doi.org/10.3390/cells3020627
APA StyleDaniel, J., & Liebau, E. (2014). The Ufm1 Cascade. Cells, 3(2), 627-638. https://doi.org/10.3390/cells3020627