Functional Assessment of Pharmacological Telomerase Activators in Human T Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
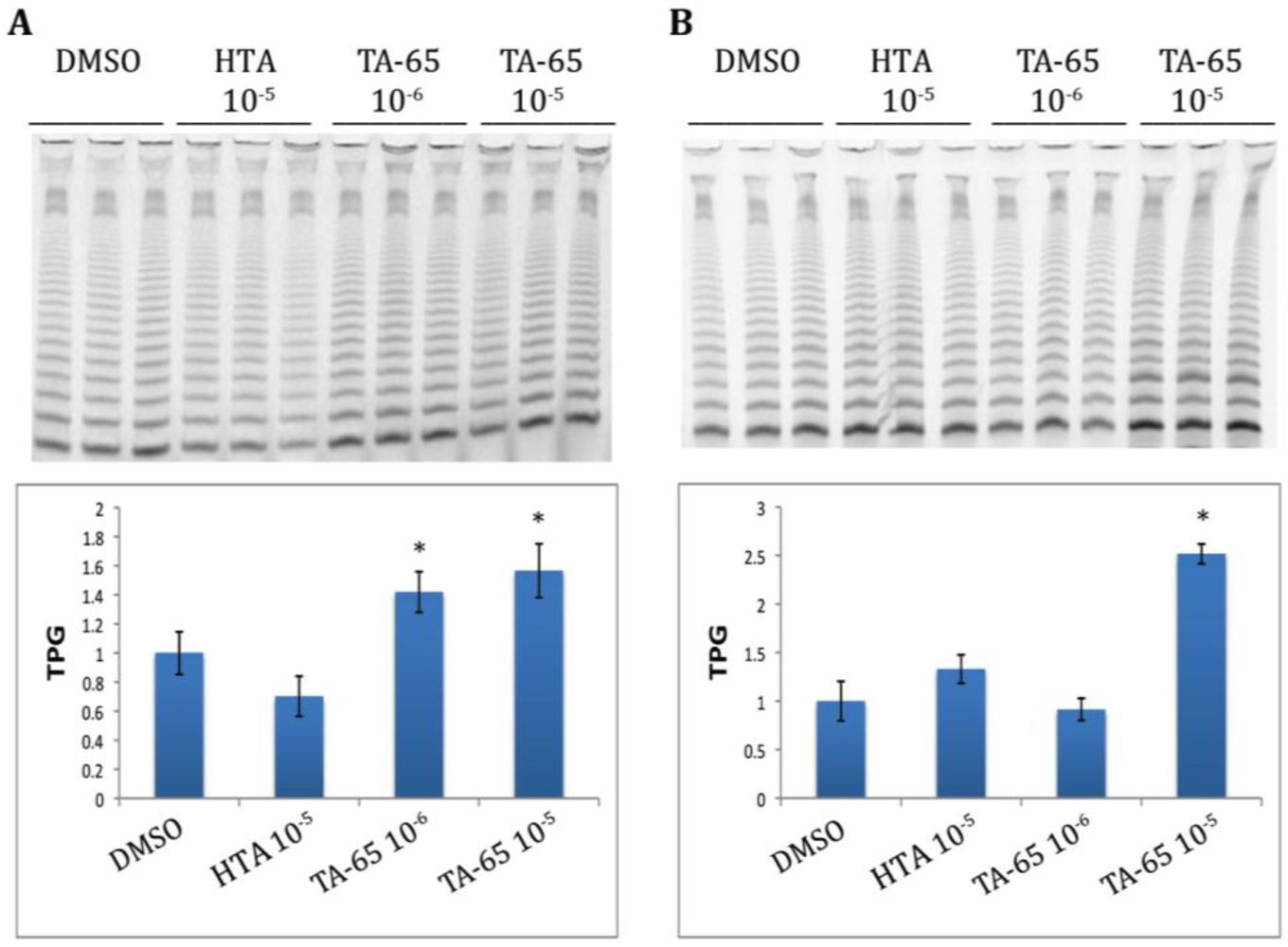
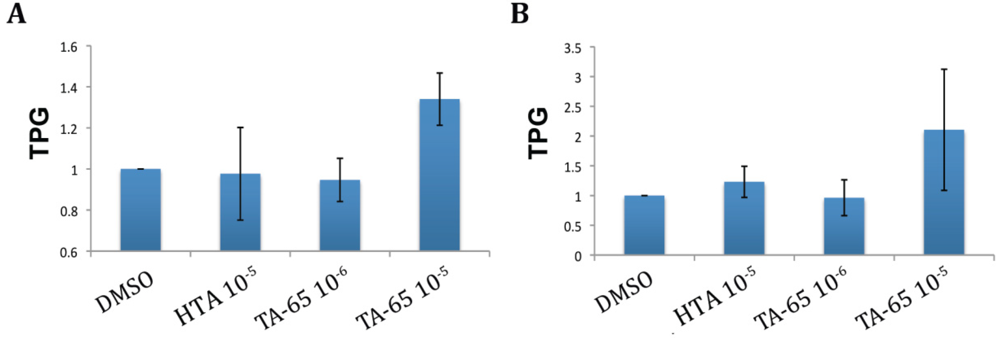
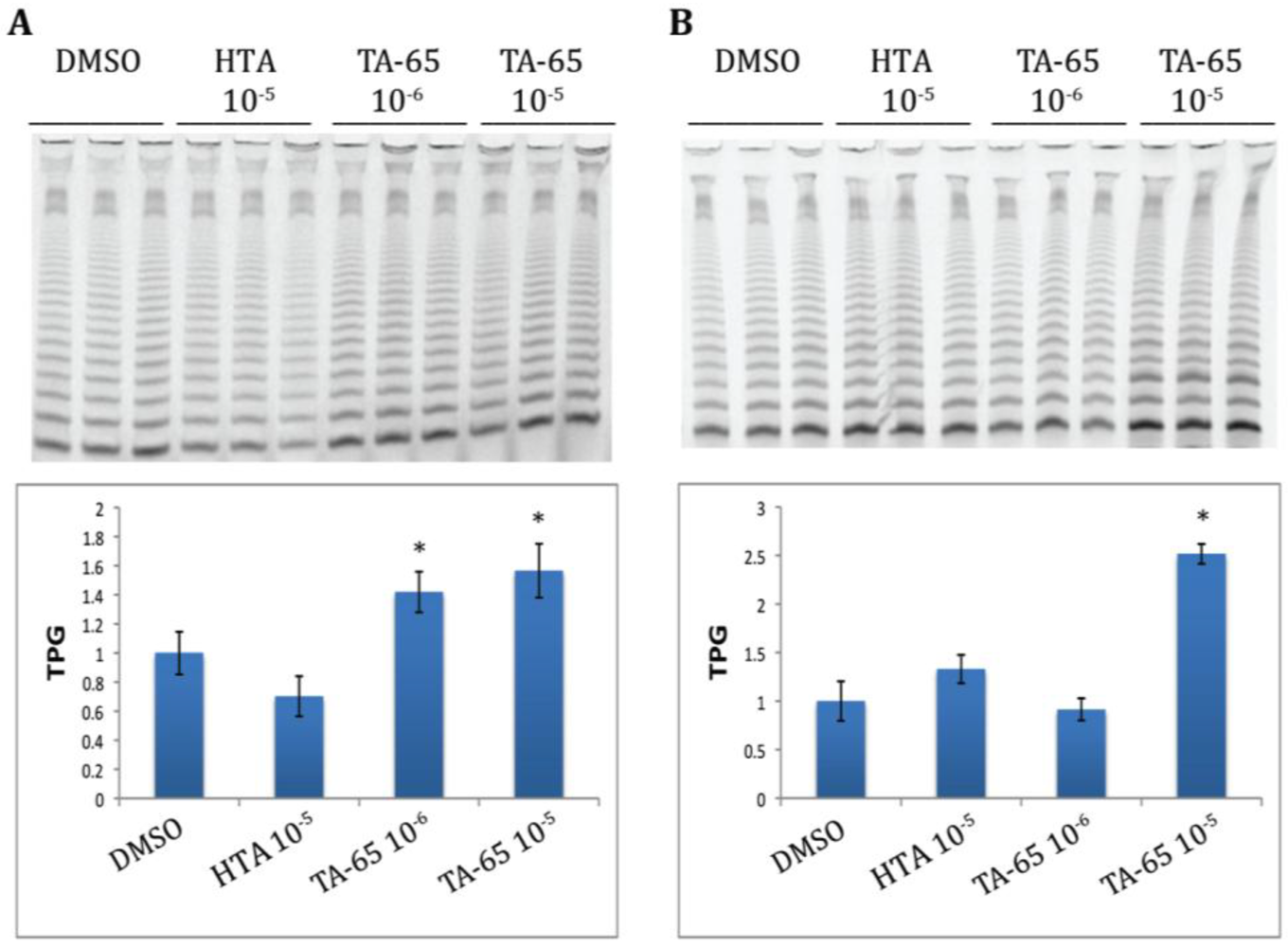
2.1. TA-65 but Not HTA Increased Telomerase Activity in All Donors’ T Cells during Primary and Secondary Stimulations
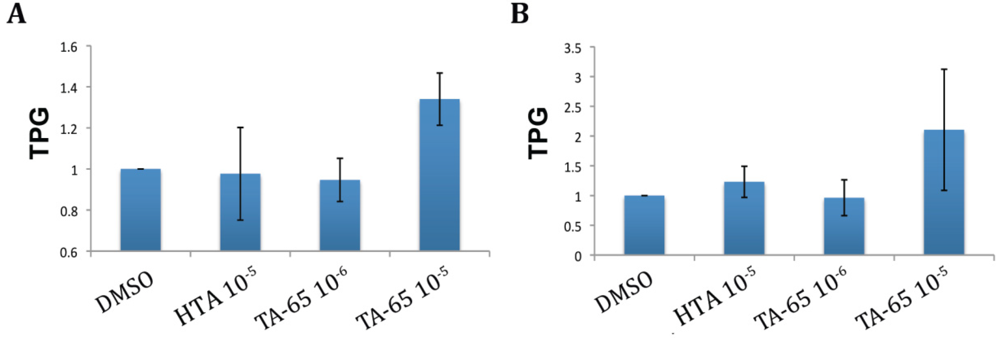
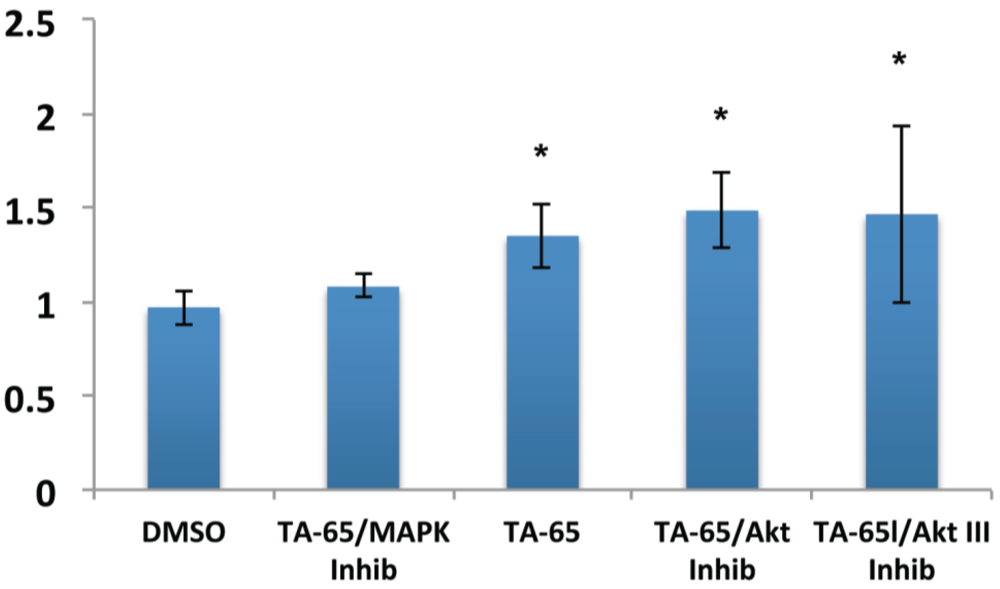
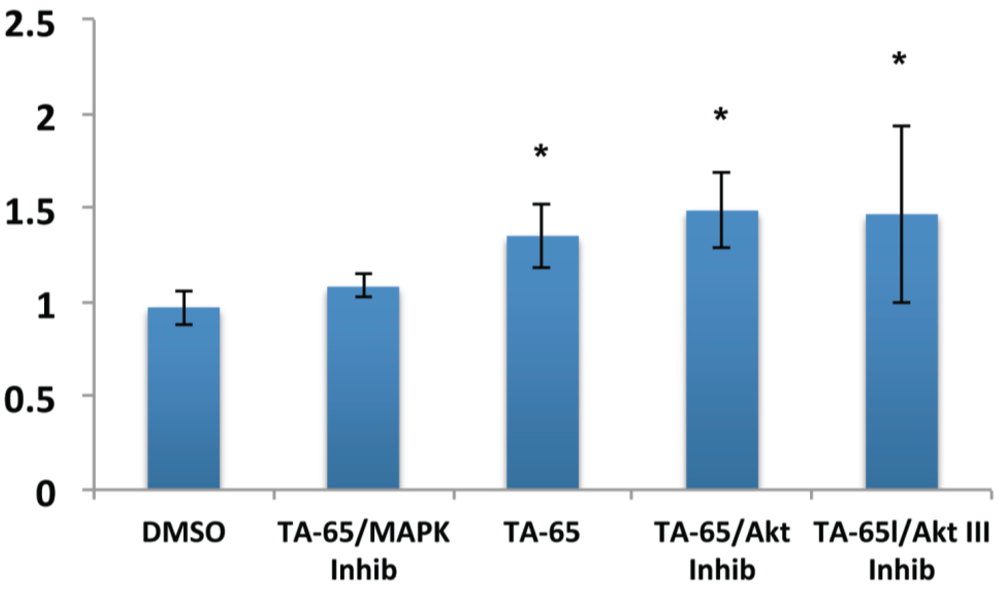
2.2. MAPK Specific Inhibitor Blocks TA-65 Induced Telomerase Activity
2.3. Telomerase Activator TA-65 but not HTA Increases T Cells Proliferation

3. Discussion
4. Experimental Section
4.1. Formulation and Treatments of TA-65 and HTA
4.2. Cell Treatment and Stimulation
4.3. Cell Signaling Inhibitors
4.4. Telomerase Activity Measurements
5. Conclusions
Acknowledgments
Conflict of Interest
References
- de Lange, T. Shelterin: The protein complex that shapes and safeguards human telomeres. Gene Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef]
- Artandi, S.E.; DePinho, R.A. Telomeres and telomerase in cancer. Carcinogenesis 2010, 31, 9–18. [Google Scholar] [CrossRef]
- Counter, C.M.; Avilion, A.A.; LeFeuvre, C.E.; Stewart, N.G.; Greider, C.W.; Harley, C.B.; Bacchetti, S. Telomere shortening associated with chromosome instability is arrested in inmortal cells which express telomerase activity. EMBO J. 1992, 11, 1921–1929. [Google Scholar]
- Price, C. M. Centromeres and telomeres. Curr Opin Cell Biol 1992, 4, 379–84. [Google Scholar] [CrossRef]
- Monteiro, J.; Batliwalla, F.; Ostrer, H.; Gregersen, P.K. Shortened telomeres in clonally expanded CD28-CD8+ T cells imply a replicative history that is distinct from their CD28+CD8+ counterparts. J. Immunol. 1996, 156, 3587–3590. [Google Scholar]
- Roth, A.; Yssel, H.; Pene, J.; Chavez, E.A.; Schertzer, M.; Lansdorp, P.M.; Spits, H.; Luiten, R.M. Telomerase levels control the lifespan of human T lymphocytes. Blood 2003, 102, 849–857. [Google Scholar] [CrossRef]
- Weng, N.P.; Levine, B.L.; June, C.H.; Hodes, R.J. Human naive and memory T lymphocytes differ in telomeric length and replicative potential. Proc. Natl. Acad. Sci. USA 1995, 92, 11091–11094. [Google Scholar] [CrossRef]
- Valenzuela, H.F.; Effros, R.B. Divergent telomerase and CD28 expression patterns in human CD4 and CD8 T cells following repeated encounters with the same antigenic stimulus. Clin. Immunol. 2002, 105, 117–125. [Google Scholar] [CrossRef]
- Hiyama, K.; Hirai, Y.; Kyoizumi, S.; Akiyama, M.; Hiyama, E.; Piatyszek, M.A.; Shay, J.W.; Ishioka, S.; Yamakido, M. Activation of telomerase in human lymphocytes and hematopoietic progenitor cells. J. Immunol. 1995, 155, 3711–3715. [Google Scholar]
- Jaskelioff, M.; Muller, F.L.; Paik, J.H.; Thomas, E.; Jiang, S.; Adams, A.C.; Sahin, E.; Kost-Alimova, M.; Protopopov, A.; Cadinanos, J.; et al. Telomerase reactivation reverses tissue degeneration in aged telomerase-deficient mice. Nature 2011, 469, 102–106. [Google Scholar]
- de Jesus, B.B.; Schneeberger, K.; Vera, E.; Tejera, A.; Harley, C.B.; Blasco, M.A. The telomerase activator TA-65 elongates short telomeres and increases health span of adult/old mice without increasing cancer incidence. Aging Cell 2011, 10, 604–621. [Google Scholar] [CrossRef]
- Dagarag, M.; Evazyan, T.; Rao, N.; Effros, R.B. Genetic manipulation of telomerase in HIV-specific CD8+ T cells: enhanced antiviral functions accompany the increased proliferative potential and telomere length stabilization. J. Immunol. 2004, 173, 6303–6311. [Google Scholar]
- Fauce, S.R.; Jamieson, B.D.; Chin, A.C.; Mitsuyasu, R.T.; Parish, S.T.; Ng, H.L.; Kitchen, C.M.; Yang, O.O.; Harley, C.B.; Effros, R.B. Telomerase-based pharmacologic enhancement of antiviral function of human CD8+ T lymphocytes. J. Immunol. 2008, 181, 7400–7406. [Google Scholar]
- Harley, C.B.; Liu, W.; Blasco, M.; Vera, E.; Andrews, W.H.; Briggs, L.A.; Raffaele, J.M. A natural product telomerase activator as part of a health maintenance program. Rejuvenation Res. 2011, 14, 45–56. [Google Scholar]
- Harley, C.B.; Futcher, A.B.; Greider, C.W. Telomeres shorten during ageing of human fibroblasts. Nature 1990, 345, 458–460. [Google Scholar] [CrossRef]
- Allsopp, R.C.; Vaziri, H.; Patterson, C.; Goldstein, S.; Younglai, E.V.; Futcher, A.B.; Greider, C.W.; Harley, C.B. Telomere length predicts replicative capacity of human fibroblasts. Proc. Natl. Acad. Sci. USA 1992, 89, 10114–10118. [Google Scholar]
- Bodnar, A.G.; Ouellette, M.; Frolkis, M.; Holt, S.E.; Chiu, C.P.; Morin, G.B.; Harley, C.B.; Shay, J.W.; Lichtsteiner, S.; Wright, W.E. Extension of life-span by introduction of telomerase into normal human cells. Science 1998, 279, 349–352. [Google Scholar] [CrossRef]
- Parish, S.T.; Wu, J.E.; Effros, R.B. Sustained CD28 expression delays multiple features of replicative senescence in human CD8 T lymphocytes. J. Clin. Immunol. 2010, 30, 798–805. [Google Scholar] [CrossRef]
- Hooijberg, E.; Ruizendaal, J.J.; Snijders, P.J.; Kueter, E.W.; Walboomers, J.M.; Spits, H. Immortalization of human CD8+ T cell clones by ectopic expression of telomerase reverse transcriptase. J. Immunol. 2000, 165, 4239–4245. [Google Scholar]
- Migliaccio, M.; Amacker, M.; Just, T.; Reichenbach, P.; Valmori, D.; Cerottini, J.C.; Romero, P.; Nabholz, M. Ectopic human telomerase catalytic subunit expression maintains telomere length but is not sufficient for CD8+ T lymphocyte immortalization. J. Immunol. 2000, 165, 4978–84. [Google Scholar]
- Denzler, K.L.; Waters, R.; Jacobs, B.L.; Rochon, Y.; Langland, J.O. Regulation of inflammatory gene expression in PBMCs by immunostimulatory botanicals. PLoS One 2010, 5, e12561. [Google Scholar]
- Auyeung, K.K.; Mok, N.L.; Wong, C.M.; Cho, C.H.; Ko, J.K. Astragalus saponins modulate mTOR and ERK signaling to promote apoptosis through the extrinsic pathway in HT-29 colon cancer cells. Int. J. Mol. Med. 2010, 26, 341–9. [Google Scholar]
- Collins, R.A. A ten-year audit of traditional Chinese medicine and other natural product research published in the Chinese Medical Journal (2000–2009). Chin. Med. J. (Engl) 2011, 124, 1401–1408. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Molgora, B.; Bateman, R.; Sweeney, G.; Finger, D.; Dimler, T.; Effros, R.B.; Valenzuela, H.F. Functional Assessment of Pharmacological Telomerase Activators in Human T Cells. Cells 2013, 2, 57-66. https://doi.org/10.3390/cells2010057
Molgora B, Bateman R, Sweeney G, Finger D, Dimler T, Effros RB, Valenzuela HF. Functional Assessment of Pharmacological Telomerase Activators in Human T Cells. Cells. 2013; 2(1):57-66. https://doi.org/10.3390/cells2010057
Chicago/Turabian StyleMolgora, Brenda, Riley Bateman, Greg Sweeney, Danielle Finger, Taylor Dimler, Rita B. Effros, and Hector F. Valenzuela. 2013. "Functional Assessment of Pharmacological Telomerase Activators in Human T Cells" Cells 2, no. 1: 57-66. https://doi.org/10.3390/cells2010057
APA StyleMolgora, B., Bateman, R., Sweeney, G., Finger, D., Dimler, T., Effros, R. B., & Valenzuela, H. F. (2013). Functional Assessment of Pharmacological Telomerase Activators in Human T Cells. Cells, 2(1), 57-66. https://doi.org/10.3390/cells2010057