
Extracellular Trapping of Soil Contaminants by Root Border Cells: New Insights into Plant Defense

Abstract
:1. Root Border Cells
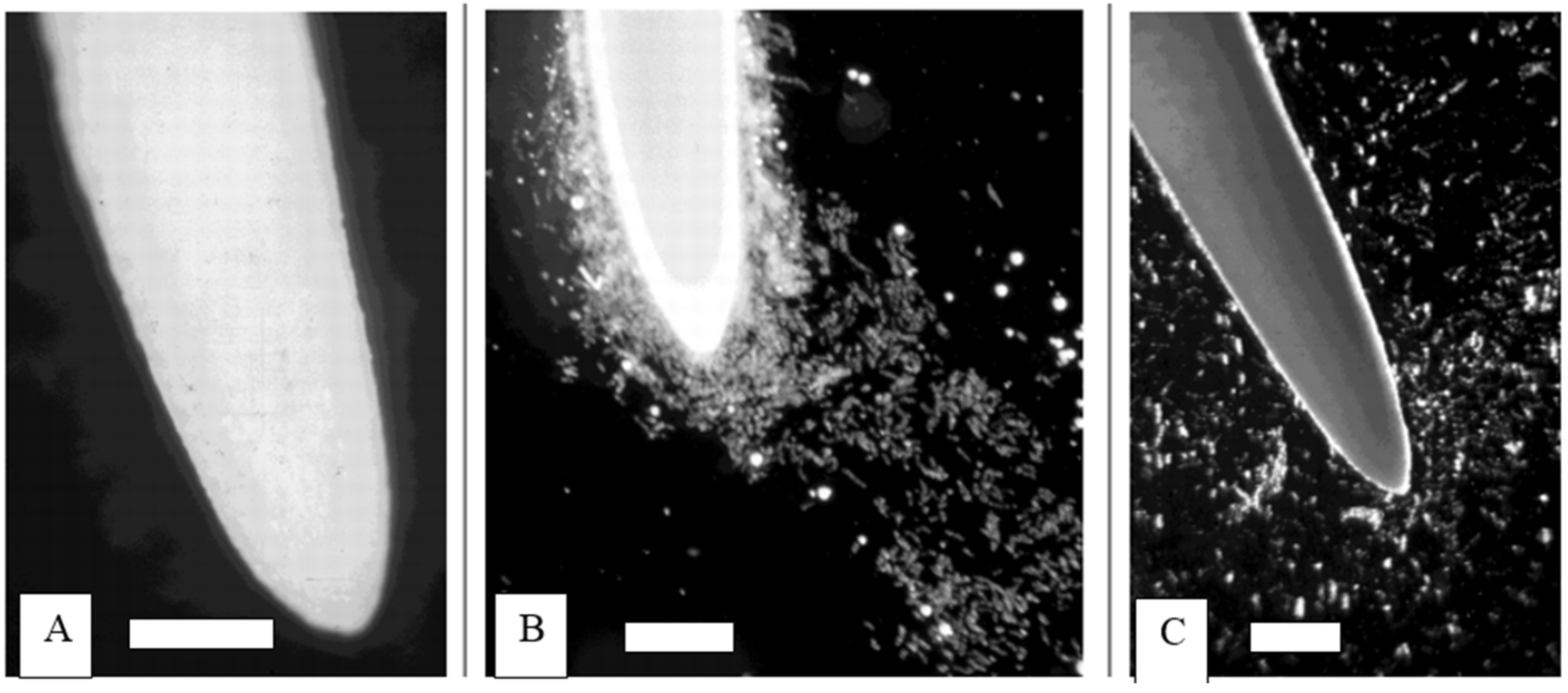
2. Extracellular Traps in Animals and Plants
3. Border Cell Trapping of Aluminum

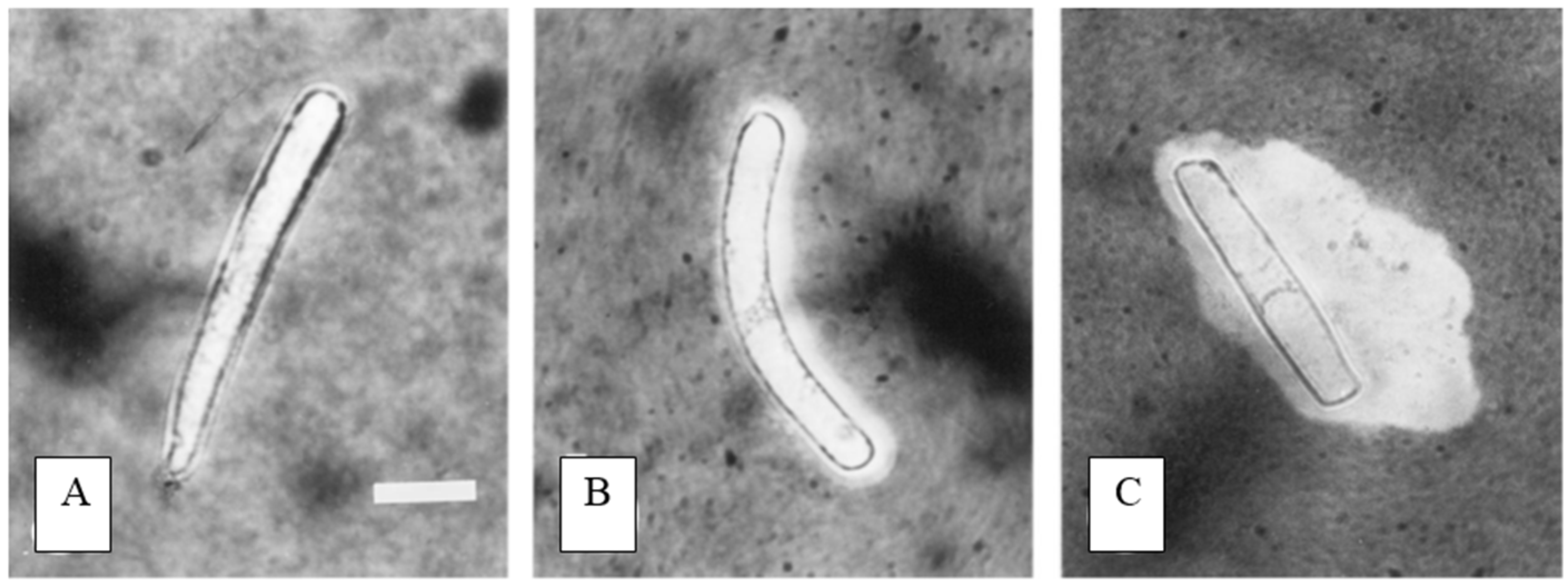
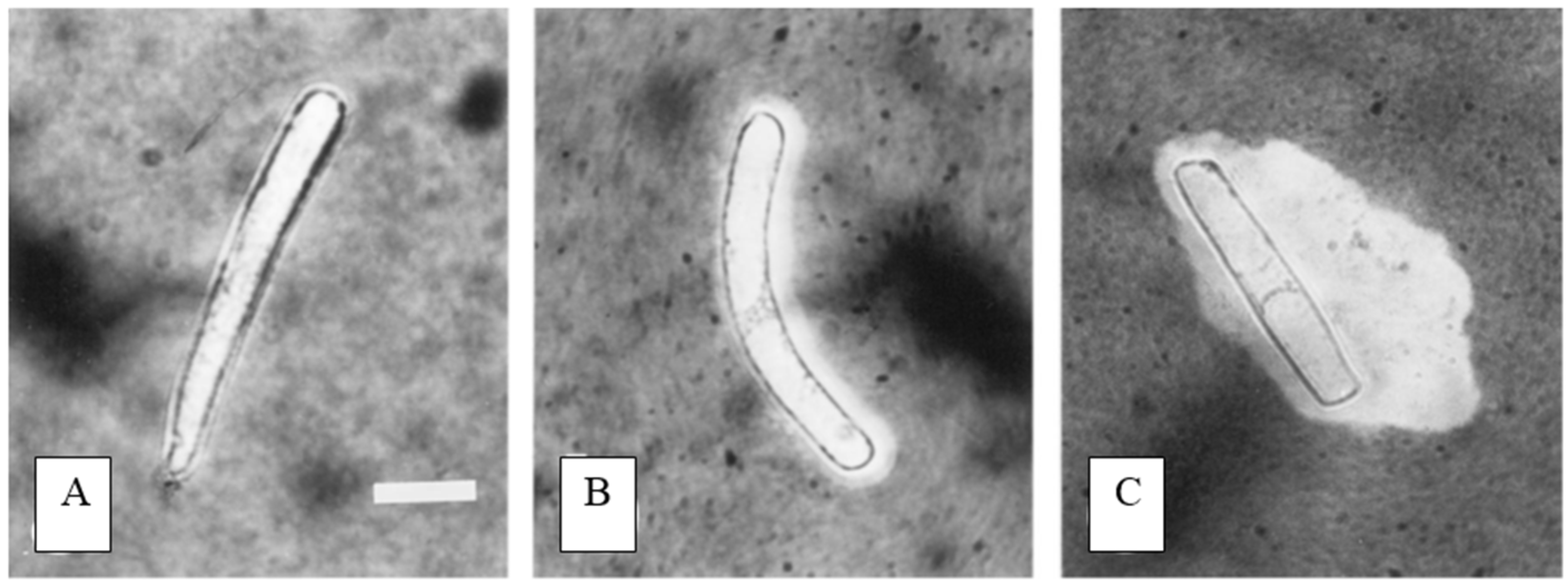
4. Border Cell Trapping of Other Soil Contaminants

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Metal | Plant | Reference |
|---|---|---|---|
| 2001 | aluminum | snapbean | [21] |
| 2003 | aluminum | pea | [22] |
| 2003 | aluminum | wheat | [24] |
| 2003 | copper | Silene | [25] |
| 2003 | cadmium | coyotillo | [26] |
| 2004 | aluminum | barley | [27] |
| 2005 | aluminum | barley | [28] |
| 2006 | aluminum | pea | [29] |
| 2007 | aluminum+boron | pea | [30] |
| 2008 | aluminum | cowpea | [31] |
| 2008 | iron | rice | [32] |
| 2008 | lead, mercury | mung bean | [33] |
| 2009 | aluminum | pea | [34] |
| 2011 | aluminum | rice | [35] |
| 2011 | aluminum | soybean | [36] |
| 2011 | copper, nickel, zinc | cowpea | [37] |
| 2011 | iron | rice | [38] |
| 2012 | iron | rice | [39] |
| 2012 | arsenic | cowpea | [40] |
| 2012 | iron, aluminum | rice | [41] |
| 2012 | aluminum | oats | [42] |
| 2012 | arsenic | fern | [43] |
| 2013 | boron, aluminum | pea | [44] |
| 2013 | aluminum | soybean | [45] |
| 2013 | copper | cotton | [46] |
| 2014 | cadmium | fern | [47] |
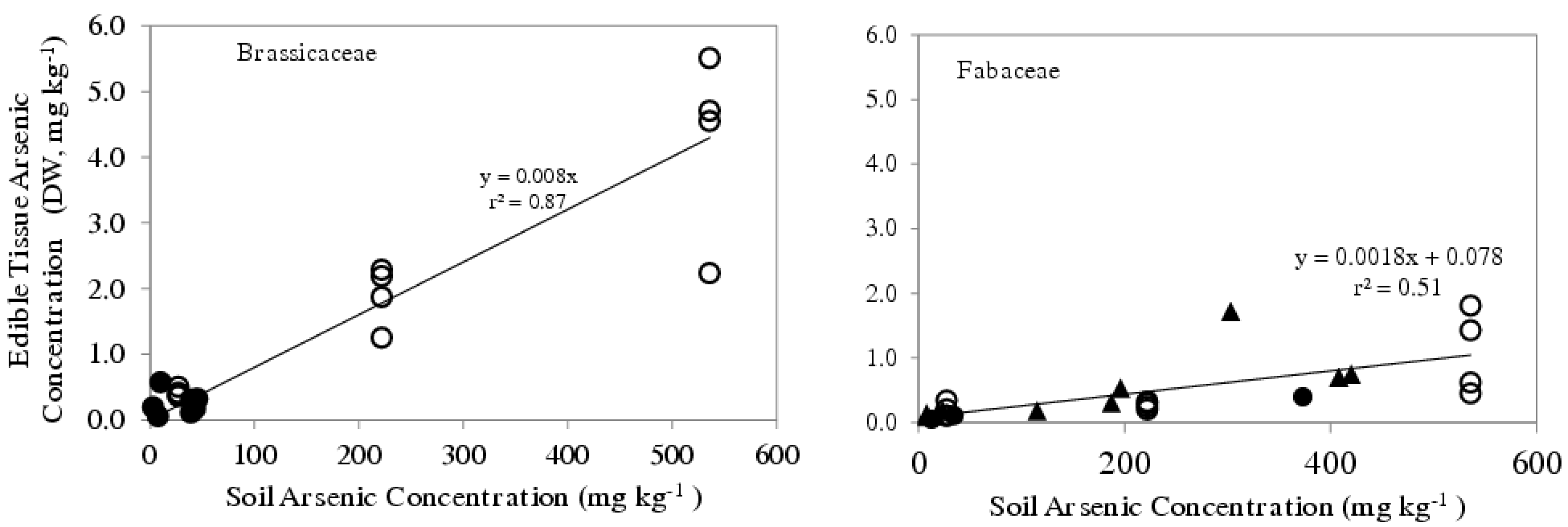
5. Border Cell Number vs. Arsenic Uptake into Edible Plants

6. Rhizofiltration vs. Rhizoprotection
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Knudson, L. Viability of detached root-cap cells. Am. J. Bot. 1919, 6, 309–310. [Google Scholar] [CrossRef]
- Hawes, M.C.; Wheeler, H. Factors affecting victorin-induced cell death: Temperature and plasmolysis. Physiol. Plant Pathol. 1982, 20, 137–144. [Google Scholar] [CrossRef]
- Brigham, L.A.; Woo, H.H.; Hawes, M.C. Differential expression of proteins and mRNAs from border cells and root tips of pea. Plant Physiol. 1995, 109, 457–463. [Google Scholar] [PubMed]
- Hawes, M.C.; Brigham, L.A.; Wen, F.; Woo, H.H.; Zhu, Y. Function of root border cells in plant health: Pioneers in the rhizosphere. Annu. Rev. Phytopathol. 1998, 36, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Watson, B.S.; Bedair, M.F.; Urbanczyk-Wochniak, E.; Huhman, D.V.; Yang, D.S.; Allen, S.N.; Li, W.; Tang, Y.; Sumner, L.W. Integrated metabolomics and transcriptomics reveal enhanced specialized metabolism in Medicago truncatula root border cells. Plant Physiol. 2015, 167, 1699–1716. [Google Scholar] [CrossRef] [PubMed]
- Hawes, M.C.; Curlango-Rivera, G.; Wen, F.; White, G.J.; VanEtten, H.D.; Xiong, Z. Extracellular DNA: The tip of root defenses? Plant Sci. 2011, 180, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrach, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Metzler, K.D.; Goosmann, C.; Lubojemska, A.; Zychlinsky, A.; Papayanopoulos, V. A myeloperoxidase-containing complex regulates neutrophil elastase release and actin dynamics during NETosis. Cell Rep. 2014, 8, 883–896. [Google Scholar] [CrossRef] [PubMed]
- Nasser, W.; Bersa, S.B.; Olsen, R.J.; Dean, M.A.; Rice, K.A.; Long, S.W.; Kristinsson, K.G.; Gottfredsson, M.; Vuopio, J.; Raisanen, K.; et al. Evolutionary pathway to increased virulence and epidemic group A Streptococcus disease derived from 3615 genome sequences. Proc. Natl. Acad. Sci. USA 2014, 111, E1768–E1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchanan, J.T.; Simpson, A.J.; Aziz, R.K.; Liu, G.Y.; Kristian, S.A.; Kotb, M.; Feramisco, J.; Nizet, V. DNase expression allows the pathogen group A Streptococcus to escape killing in neutrophil extracellular traps. Curr. Biol. 2006, 16, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Wen, F.; VanEtten, H.D.; Tsaprailis, G.; Hawes, M.C. Extracellular proteins in Pisum sativum L. root tip and border cell exudates. Plant Physiol. 2007, 143, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Wen, F.; White, G.A.; Xiong, Z.; VanEtten, H.D.; Hawes, M.C. Extracellular DNA is required for root tip resistance to fungal infection. Plant Physiol. 2009, 151, 820–829. [Google Scholar] [CrossRef] [PubMed]
- Tuan, T.; Hawes, M.C.; Allen, C. Extracellular DNases contribute to virulence of Ralstonia solanacearum. Phytopathology 2013, 103, 147–148. [Google Scholar]
- Liz, R.; Simard, J.; Leonardi, L.; Girard, D. Silver nanoparticles rapidly induce atypical human neutrophil cell death by a process involving inflammatory caspases and reactive oxygen species and induce neutrophil extracellular traps release upon cell adhesion. Int. Immunopharmacol. 2015, 28, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Mushtakova, V.M.; Fomina, V.A.; Rogovin, V.V. Toxic effect of heavy metals on human blood neutrophils. Biol. Bulletin 2005, 32, 276–278. [Google Scholar] [CrossRef]
- Niermiec, J.J.; de Samber, B.; Garrevoet, J.; Vergucht, E.; Vekemans, B.; de Rycke, R.; Björn, E.; Sandblad, L.; Wellenreuther, G.; Falkenberg, G.; et al. Trace element landscape of resting and activated human neutrophils on the sub-micrometer level. Metallomics 2015, 7, 996–1010. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.; Delhaize, E. Adaptations to aluminium toxicity. In Plant Stress Physiology; CABI: Wallingford, UK, 2012; pp. 171–193. [Google Scholar]
- Klug, B.; Kirchner, T.W.; Horst, W.J. Differences in aluminum accumulation and resistance between genotypes of the genus Fagopyrum. Agronomy 2015, 5, 418–434. [Google Scholar] [CrossRef]
- Richard, C.; Munyinda, K.; Kinkese, T.; Osiru, D.S. Genotypic variation in seedling tolerance to aluminum toxicity in historical maize inbred lines of Zambia. Agronomy 2015, 5, 200–219. [Google Scholar] [CrossRef]
- Ryan, P.R.; Ditomaso, J.M.; Kochian, L.V. Aluminum toxicity in roots: An investigation of spatial sensitivity and the role of the root cap. J. Exp. Bot. 1993, 44, 437–446. [Google Scholar] [CrossRef]
- Miyasaka, S.; Hawes, M.C. Possible role of root border cells in detection and avoidance of aluminum toxicity. Plant Physiol. 2013, 125, 1978–1987. [Google Scholar] [CrossRef]
- Brigham, L.A.; Miyasaka, S.; Hawes, M.C. Avoidance of aluminum toxicity: Role of root border cells. Plant Nut. Dev. 2001, 92, 452–453. [Google Scholar]
- Dyrssen, D.; Haraldsson, C.; Nyberg, E.; Wedborg, M. Complexation of aluminum with DNA. J. Inorg. Biochem. 1987, 29, 67–75. [Google Scholar] [CrossRef]
- Zhu, M.Y.; Ahn, S.; Matsumoto, H. Inhibition of growth and development of root border cells in wheat by Al. Physiol. Plant. 2003, 117, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Llugany, M.; Lombini, A.; Poschenrieder, C.; Dinelli, E.; Barcelo, J. Different mechanisms account for enhanced copper resistance in Silene armeria ecotypes from mine spoil and serpentine sites. Plant Soil 2003, 251, 55–63. [Google Scholar] [CrossRef]
- Zelko, I.; Lux, A. Effect of cadmium on Karwinskia humboldtiana roots. Biologia 2003, 59, 205–209. [Google Scholar]
- Pan, J.; Ye, D.; Wang, L.; Hua, J.; Zhao, G.; Pan, W.; Han, N.; Zhu, M. Root border cell development is a temperature-insensitive and Al-sensitive process in barley. Plant Cell Physiol. 2004, 45, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Tamas, L.; Budikova, S.; Huttova, J.; Mistrik, I.; Simonovicova, M. Aluminum-induced cell death of barley-root border cells is correlated with peroxidase- and oxalate oxidase-mediated hydrogen peroxide production. Plant Cell Rep. 2005, 24, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Feng, Y.M.; Goldbach, H.E. Mist culture for mass harvesting of root border cells: Aluminum effects. J. Plant Nutr. Soil Sci. 2006, 169, 670–674. [Google Scholar] [CrossRef]
- Yu, M.; Goldbach, H.E. Influence of boron on Al absorption and Ca release of root border cells of pea (Pisum sativum). In Advances in Plant and Animal Boron Nutrition; Springer: Dordrecht, The Netherlands, 2007; pp. 63–68. [Google Scholar]
- Chen, W.; Liu, P.; Xu, G.; Cai, M.; Yu, H.; Chen, M. Effects of Al3+ on the biological characteristics of cowpea root border cells. Acta Physiol. Plant. 2008, 30, 303–308. [Google Scholar] [CrossRef]
- Xing, C.; Zhu, M.; Cai, M.; Liu, P.; Xu, G.; Wu, S. Developmental characteristics and response to iron toxicity of root border cells in rice seedlings. J. Zhejiang Univ. Sci. B. 2008, 9, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Zhu, L.; Liu, X.Y.; Zhang, Y.; Zhao, N. Individual and joint effects of lead and mercury on the viability of root border cells in mung bean (Vigna radiata). In Proceedings of the International Symposium on Environmental Science and Technology, Shanghai, China, 2–5 June 2009; pp. 254–258.
- Yu, M.; Shen, R.; Xiao, H.; Xu, J.; Wang, H.; Wang, H.; Zeng, Q.; Bien, J. Boron alleviates aluminum toxicity in pea (Pisum sativum). Plant Soil 2009, 314, 87–98. [Google Scholar] [CrossRef]
- Cai, M.; Zhang, S.; Xing, C.; Wang, F.; Wang, N.; Zhu, L. Developmental characteristics and aluminum resistance of root border cells in rice seedlings. Plant Sci. 2008, 180, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Wang, F.; Li, R.; Zhang, S.; Wang, W.; Xu, G. Response and tolerance of root border cells to aluminum toxicity in soybean seedlings. J. Inorg. Biochem. 2011, 105, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Kopittke, P.M.; Menzies, N.W.; de Jonge, M.D.; McKenna, B.D.; McKenna, B.A.; Donner, E.; Webb, R.I.; Paterson, D.J.; Howard, D.L.; Ryan, C.G.; et al. In situ distribution and speciation of toxic Cu, Ni and Zn in hydrated roots of cowpea. Plant Physiol. 2011, 156, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zheng, G.H.; Liu, P.; Song, J.M.; Xu, G.D.; Cai, M.Z. Morphological and physiological responses of root tip cells to Fe2+ toxicity in rice. Acta Physiol. Plant. 2011, 33, 683–689. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.P.; Liu, P.; Song, J.M.; Xu, G.D.; Zheng, G.H. Effect of toxic Fe2+ levels on the biological characteristics of rice root border cells. Russ. J. Plant Physiol. 2012, 59, 766–771. [Google Scholar] [CrossRef]
- Kopittke, P.M.; de Jonge, M.D.; Menzies, N.W.; Wang, P.; Donner, E.; McKenna, B.A.; Paterson, D.; Howard, D.L.; Lombi, E. Examination of the distribution of arsenic in hydrated and fresh cowpea roots using two- and three-dimensional techniques. Plant Physiol. 2012, 159, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Zhang, S.; Xing, C.; Wang, F.; Zhu, L.; Wang, N.; Liu, L. Interaction between iron plaque and root border cells ameliorates aluminum toxicity of Oryza sativa differing in aluminum tolerance. Plant Soil. 2012, 353, 155–167. [Google Scholar] [CrossRef]
- Radmer, L.; Tesfaye, M.; Somers, D.A.; Temple, S.J.; Vance, C.P.; Samac, D.A. Aluminum resistance mechanisms in oat (Avena sativa L.). Plant Soil 2012, 351, 121–134. [Google Scholar] [CrossRef]
- Forino, L.M.C.; Castiglione, M.R.; Bartoli, G.; Balestri, M.; Andreuci, A.; Tagliasacchi, A.M. Arsenic-induced morphogenic response in roots of arsenic hyperaccumulator fern Pteris vittata. J. Hazard. Mater. 2012, 235, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yu, M.; Wang, C. Influence of boron and aluminum on production and viability of root border cells of pea (Pisum sativum). Adv. Plant Anim. Boron Nut. 2013, 30, 69–74. [Google Scholar]
- Cai, M.; Wang, N.; Xing, C.; Wang, F.; Wu, K.; Du, X. Immobilization of aluminum with mucilage secreted by root cap and root border cells is related to aluminum resistance in Glycine max L. Environ. Sci. Pollut. Res. 2013, 20, 8924–8933. [Google Scholar] [CrossRef] [PubMed]
- Curlango-Rivera, G.; Huskey, D.A.; Mostafa, A.; Kessler, J.O.; Xiong, Z.; Hawes, M.C. Intraspecies variation in cotton border cell production: Rhizosphere microbiome implications. Am. J. Bot. 2013, 100, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Balestri, M.; Ceccarini, A.; Forino, L.M.C.; Zelko, I.; Martinka, M.; Lux, A.; Ruffini Castiglione, M. Cadmium uptake, localization and stress-induced morphogenic response in the fern Pteris vittata. Planta 2014, 239, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Curlango-Rivera, G.; Duclos, D.V.; Ebolo, J.J.; Hawes, M.C. Transient exposure of root tips to primary and secondary metabolites: Impact on root growth and production of border cells. Plant Soil. 2010, 306, 206–216. [Google Scholar] [CrossRef]
- Hawes, M.C.; Curlango-Rivera, G.; Xiong, Z.; Kessler, J.O. Roles of root border cells in plant defense and regulation of rhizosphere microbial populations by extracellular DNA “trapping”. Plant Soil 2012, 355, 1–16. [Google Scholar] [CrossRef]
- Odell, R.E.; Dumlao, M.R.; Samar, D.; Silk, W.K. Stage-dependent border cell and carbon flow from roots to rhizosphere. Am. J. Bot. 2008, 95, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Shen, R.; Liu, J.; Chen, R.; Xu, M.; Yang, Y.; Xiao, H.; Wang, H.; Wang, H.; Wang, C. The role of root border cells in aluminum resistance of pea (Pisum sativum) grown in mist culture. J. Plant Nutr. Soil Sci. 2009, 172, 528–534. [Google Scholar] [CrossRef]
- Ramirez-Andreotta, M.D.; Brusseau, M.L.; Artiola, J.F.; Maier, R.M. A greenhouse and field-based study to determine the accumulation of arsenic in common homegrown vegetables grown in mining-affected soils. Sci. Total Environ. 2013, 443, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Driouich, A.; Follet-Gueye, M.; Vicre-Gibouin, M.; Hawes, M.C. Root border cells and secretions as critical elements in plant host defense. Curr. Opin. Plant. Biol. 2013, 16, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Hawes, M.C.; Pueppke, S.G. Sloughed peripheral root cap cells: Yield from different species and callus formation from single cells. Am. J. Bot. 1986, 73, 1466–1473. [Google Scholar] [CrossRef]
- Vangronsveld, J.; Herzig, R.; Weyens, N.; Boulet, J.; Adriaensen, K.; Ruttens, A.; Thewys, T.; Vassilev, A.; Meers, E.; Nehnevajova, E.; et al. Phytoremediation of contaminated soils and groundwater: Lessons from the field. Environ. Sci. Pollut. Res. 2009, 16, 765–794. [Google Scholar] [CrossRef] [PubMed]
- Interstate Technology & Regulatory Council (ITRC). Phytotechnology Technical and Regulatory Guidance and Decision Trees; PHYTO-3. Interstate Technology & Regulatory Council, Phytotechnologies Team: Washington, DC, USA, 2009. Available online: http://www.itrcweb.org (accessed on 30 October 2015).
- Dushenkov, V.; Nanda, K.; Motto, H.; Raskin, I. Rhizofiltration: The use of plants to remove heavy metals from aqueous streams. Environ. Sci. Technol. 1995, 30, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Anawar, H.M.; Garcia-Sanchez, A.; Tari Kul Alam, M.; Rahman, M.M. Phytofiltration of water polluted with arsenic and heavy metals. Int. J. Environ. Pollut. 2008, 33, 292–312. [Google Scholar] [CrossRef]
- Arthur, E.L.; Rice, P.J.; Rice, P.J.; Anderson, T.A.; Baladie, S.M.; Henderson, K.D.; Coats, J.R. Phytoremediation—An overview. Crit. Rev. Plant. Sci. 2005, 24, 109–122. [Google Scholar] [CrossRef]
- Cheng, S.P. Heavy metals in plants and phytoremediation. Environ. Sci. Pollut. Res. 2003, 10, 335–340. [Google Scholar] [CrossRef]
- Cooney, C.M. Sunflowers remove radionuclides from water in ongoing phytoremediation field tests. Environ. Sci. Technol. 1996, 30, 194. [Google Scholar] [CrossRef]
- Meagher, R.B.; Heaton, A.C.P. Strategies for the engineered phytoremediation of toxic element pollution: Mercury and arsenic. J. Ind. Microbiol. Biotech. 2004, 32, 502–513. [Google Scholar] [CrossRef] [PubMed]
- Page, V.; Feller, U. Heavy metals in crop plants: Transport and redistribution processes on the whole plant level. Agronomy 2015, 5, 447–463. [Google Scholar] [CrossRef]
- Raskin, I. Phytoremediation of metals: Using plants to remove pollutants from the environment. Curr. Opin. Biotechnol. 1997, 8, 221–226. [Google Scholar] [CrossRef]
- Shah, K.; Nongkynrih, J.M. Metal hyperaccumulation and bioremediation. Biol. Plant. 2007, 51, 618–634. [Google Scholar] [CrossRef]
- Cooper, P.C.; Palmer, L.J.; Chapple, I.L.C. Neutrophil extracellular traps as a new paradigm in innate immunity: Friend or foe? Periodontol. 2000 2013, 63, 165–197. [Google Scholar] [CrossRef] [PubMed]
- Curlango-Rivera, G.; Flores-Lara, Y.; Cho, I.; Huskey, D.A.; Xiong, Z.; Hawes, M.C. Signals controlling extracellular trap formation in plant and animal immune responses. Clin. Microbiol. 2014, 3, 5–7. [Google Scholar]
- Haichar, F.; Santaella, C.; Heulin, T.; Achouak, W. Root exudates mediated interactions belowground. Soil Biol. Biochem. 2015, 77, 69–80. [Google Scholar] [CrossRef]
- Hawes, M.C.; Brigham, L.A. Impact of root border cells on microbial populations in the rhizosphere. Adv. Plant Pathol. 1992, 8, 118–148. [Google Scholar]
- Durand, C.; Vicre-Gibouin, M.; Follet-Gueye, M.L.; Duponchel, L.; Moreau, M.; Lerouge, P.; Driouich, A. The organization pattern of root border-like cells of Arabidopsis is dependent on cell wall homogalacturonan. Plant Physiol. 2009, 150, 1411–1421. [Google Scholar] [CrossRef] [PubMed]
- Driouich, A.; Durnad, C.; Cannesan, M.A.; Percoco, G.; Vicré-Gibouin, M. Border cells versus border-like cells: Are they alike? J. Exp. Bot. 2010, 61, 3827–3831. [Google Scholar] [CrossRef] [PubMed]
- Endo, I.; Tange, T.; Osawa, H. A cell-type-specific defect in border cell formation in the Acacia mangium root cap developing an extraordinary sheath of sloughed-off cells. Ann. Bot. 2011, 108, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Nguema-Ona, E.; Vicre-Gibouin, M.; Cannesan, M.; Driouich, A. Arabinogalactan proteins in root-microbe interactions. Trends Plant Sci. 2013, 18, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Cannesan, M.; Durand, C.; Burel, C.; Gangneux, C.; Lerouge, P.; Ishii, T.; Laval, K.; Follet-Gueye, M.L.; Driouich, A.; Vicré-Gibouin, M. Effect of arabinogalactan proteins from the root caps of pea and Brassica napus on Aphanomyces euteiches zoospore chemotaxis and germination. Plant Physiol. 2012, 159, 1658–1670. [Google Scholar] [CrossRef] [PubMed]
- Cannesan, M.A.; Gangneux, C.; Lanoue, A.; Giron, D.; Laval, K.; Hawes, M.; Driouich, A.; Vicré-Gibouin, M. Association between border cell responses and localized root infection by pathogenic Aphanomyces euteiches. Ann. Bot. 2011, 108, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Henry, H.F.; Burken, J.G.; Maier, R.M.; Newman, L.A.; Rock, S.; Schnoor, J.L.; Suk, W.A. Phytotechnologies—Preventing Exposures, Improving Public Health. Int. J. Phytoremediation 2013, 15, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Colzi, I.; Pignattelli, S.; Glorni, E.; Papini, A.; Connelli, C. Linking root traits to copper exclusion mechanisms in Silene paradoxa L. (Caryophyllaceae). Plant Soil 2015, 390, 1–15. [Google Scholar] [CrossRef]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hawes, M.C.; McLain, J.; Ramirez-Andreotta, M.; Curlango-Rivera, G.; Flores-Lara, Y.; Brigham, L.A. Extracellular Trapping of Soil Contaminants by Root Border Cells: New Insights into Plant Defense. Agronomy 2016, 6, 5. https://doi.org/10.3390/agronomy6010005
Hawes MC, McLain J, Ramirez-Andreotta M, Curlango-Rivera G, Flores-Lara Y, Brigham LA. Extracellular Trapping of Soil Contaminants by Root Border Cells: New Insights into Plant Defense. Agronomy. 2016; 6(1):5. https://doi.org/10.3390/agronomy6010005
Chicago/Turabian StyleHawes, Martha C., Jean McLain, Monica Ramirez-Andreotta, Gilberto Curlango-Rivera, Yolanda Flores-Lara, and Lindy A. Brigham. 2016. "Extracellular Trapping of Soil Contaminants by Root Border Cells: New Insights into Plant Defense" Agronomy 6, no. 1: 5. https://doi.org/10.3390/agronomy6010005
APA StyleHawes, M. C., McLain, J., Ramirez-Andreotta, M., Curlango-Rivera, G., Flores-Lara, Y., & Brigham, L. A. (2016). Extracellular Trapping of Soil Contaminants by Root Border Cells: New Insights into Plant Defense. Agronomy, 6(1), 5. https://doi.org/10.3390/agronomy6010005