N2O Emission and Mineral N Release in a Tropical Acrisol Incorporated with Mixed Cowpea and Maize Residues

Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil
2.2. Plant Residues

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical characteristics | Cowpea | Maize |
|---|---|---|
| Total extractable polyphenol (%) | 1.3 | 1.14 |
| Acid detergent lignin (%) | 7.2 | 7.4 |
| Total nitrogen (%) | 3.4 | 0.92 |
| Organic carbon | 39.6 | 42.1 |
| C:N ratio | 11.7 | 45.6 |
2.3. Experimental Setup
2.3.1. Incubation
2.3.2. Gas Sampling and Analysis
2.3.3. Soil Sampling and Analysis
2.3.4. Water Extractable Carbon
2.3.5. Calculations
2.4. Statistical Analysis
3. Results
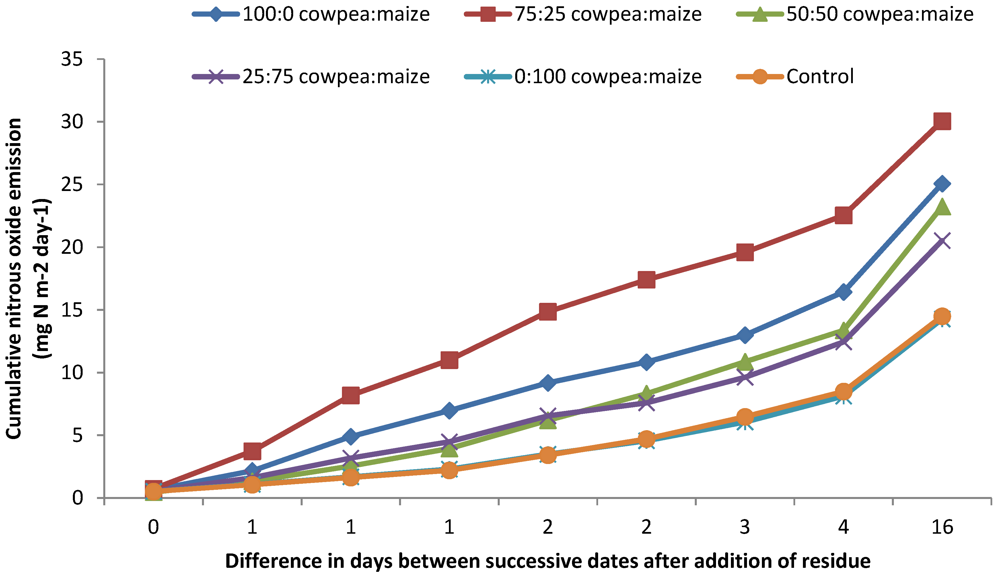
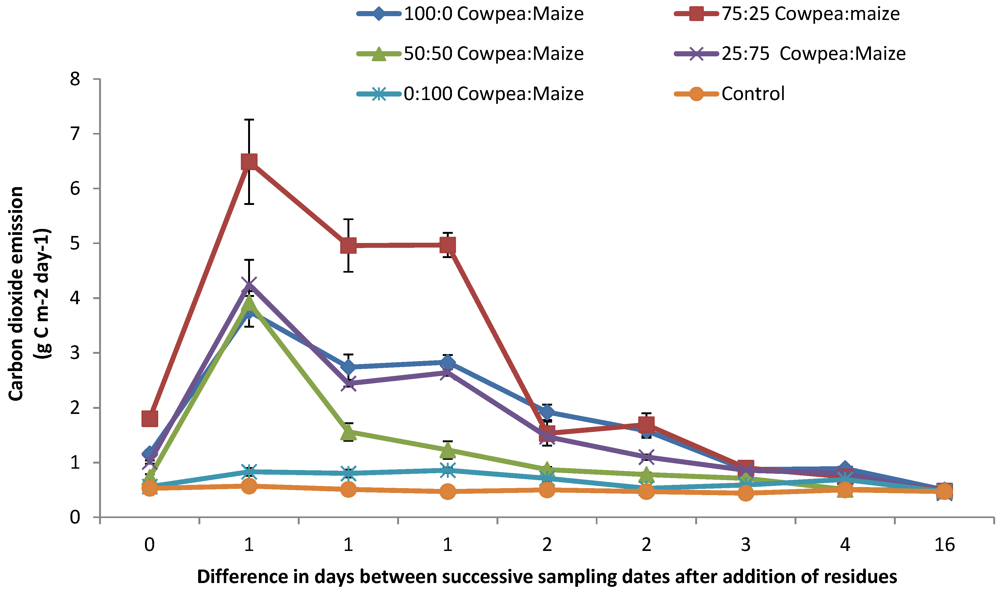
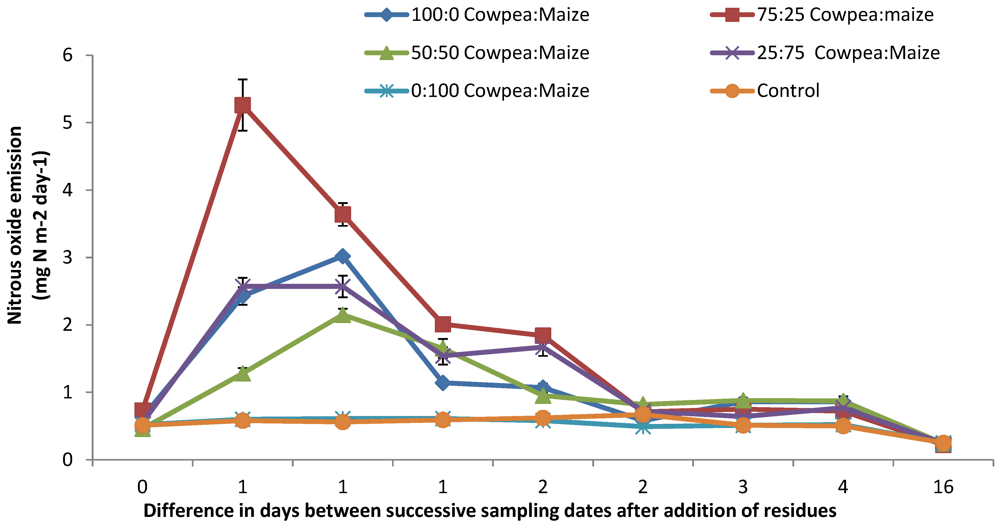
3.1. N2O and CO2 Emissions

| Treatment | N input from residues (kg N ha−1) | Emission factor (%) |
|---|---|---|
| 100:0 cowpea:maize | 136.0 | 5.4 |
| 75:25 cowpea:maize | 111.2 | 12.1 |
| 50:50 cowpea:maize | 86.4 | 9.1 |
| 25:75 cowpea:maize | 61.6 | 11.1 |
| 0:100 cowpea:maize | 36.8 | 0.16 |

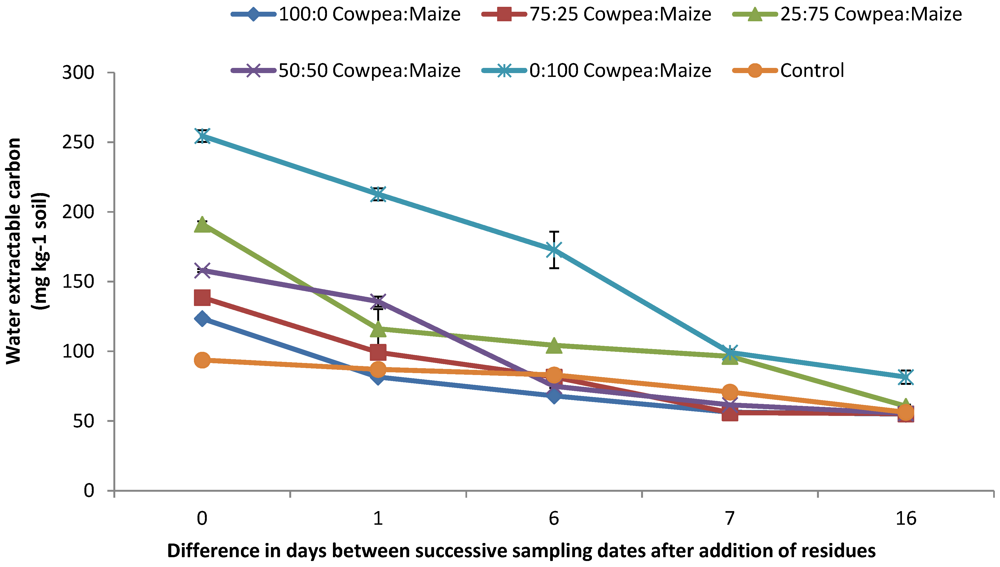
3.2. Water Extractable Carbon

| Treatment | WEC and N2O | CO2 and N2O |
|---|---|---|
| 100:0 Cowpea:maize | r = 0.76 * | r = 0.64, P < 0.05 |
| 75:25 cowpea:maize | r = 0.71 * | r = 0.83 * |
| 50:50 cowpea:maize | r = 0.94 ** | r = 0.20 |
| 25:75 cowpea:maize | r = 0.97 ** | r = 0.78 * |
| 0:100 cowpea:maize | r = 0.56 | r = 0.68 |
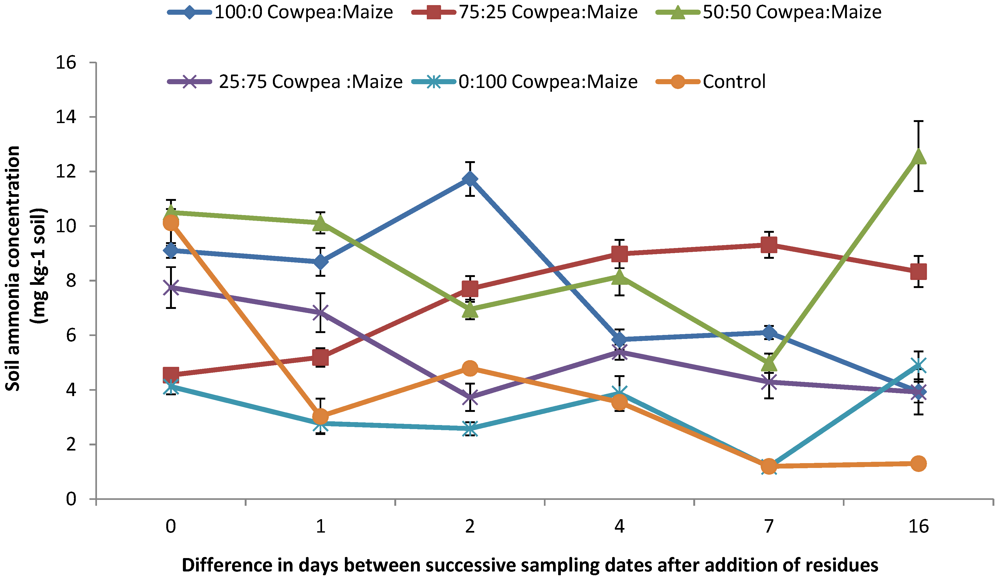
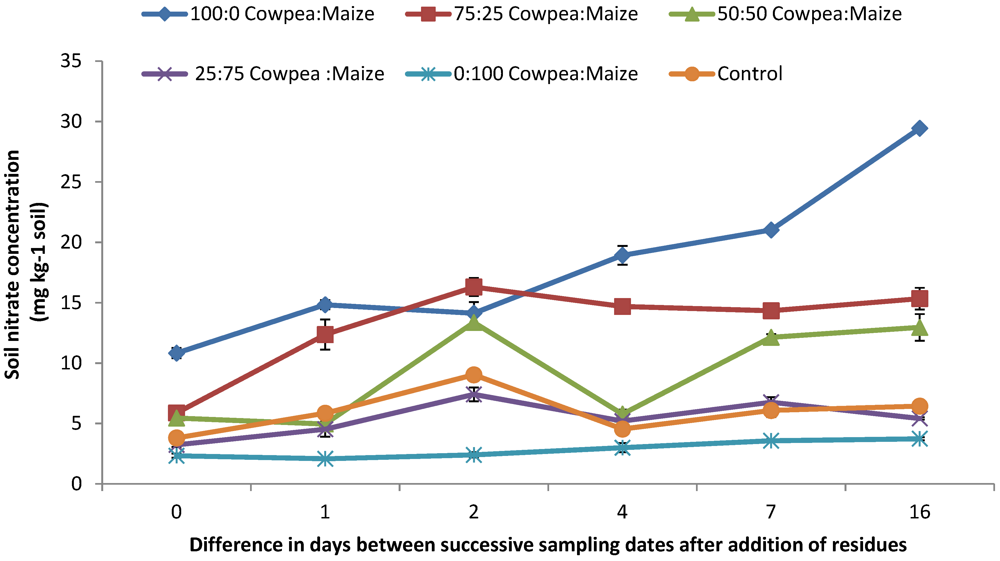
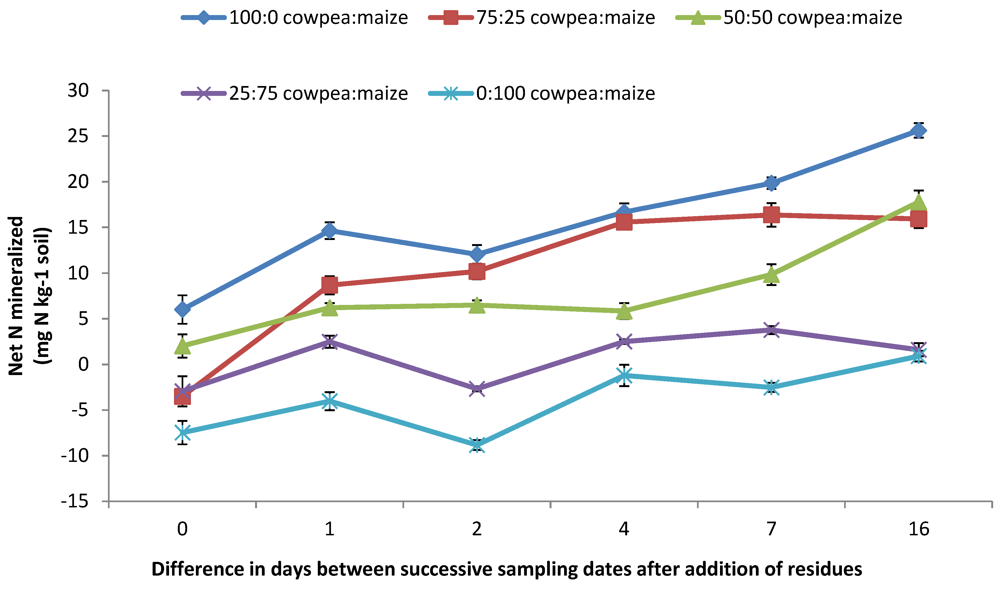
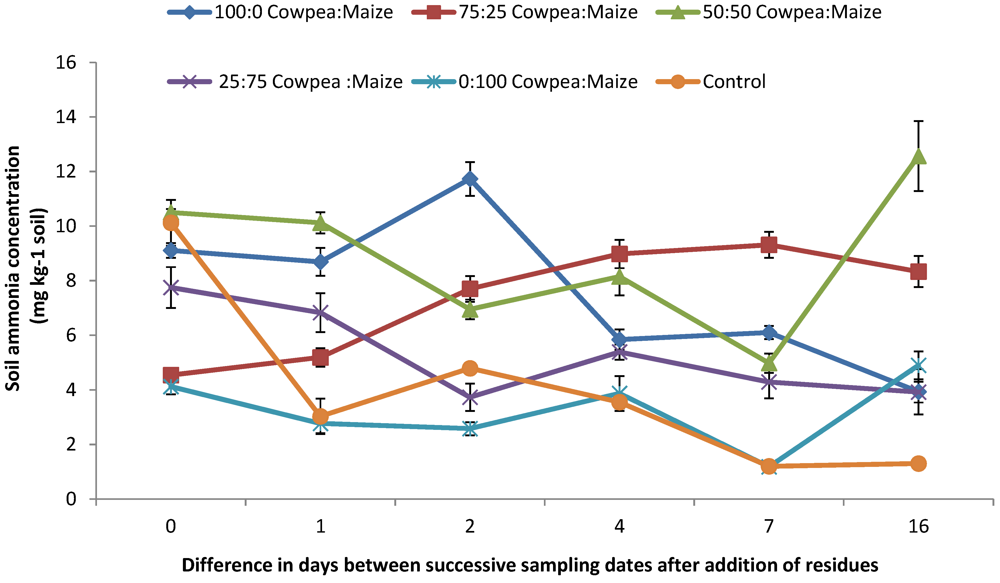
3.3. Soil Available N ( NH4+-N and NO3−-N) and Net N Mineralized


| Treatment | NH4+ + NO3− (mg kg−1 soil) | Net N mineralized |
|---|---|---|
| 100:0 cowpea:maize | 34.5 ± 1.0a | 26.9 ± 1.2a |
| 75:25 cowpea:maize | 25.6 ± 0.6b | 16.0 ± 0.8b |
| 50:50 cowpea:maize | 24.1 ± 1.8b | 16.6 ± 1.8b |
| 25:75 cowpea:maize | 9.32 ± 0.32c | 1.8 ± 0.7c |
| 0:100 cowpea:maize | 7.98 ± 1.2c | 0.47 ± 0.7c |
| Control | 7.5 ± 1.2c | - |
4. Discussion
4.1. N2O and CO2 Emissions
4.2. Mineral N (NO3− and NH4+) Concentrations and N2O Emission
4.3. Implications for Residue Management in Acrisols
5. Conclusions
Acknowledgments
References
- Crews, T.; Peoples, M.B. Legume versus fertilizer source of nitrogen: Ecological trade-offs and human needs. Agric. Ecosyst. Environ. 2004, 102, 279–297. [Google Scholar] [CrossRef]
- IPCC, Technical Summary. In Climate Change 2007: Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S.; Qin, D.; Manning, M.; Chen, Z.; Marquis, M.; Averyt, K.B.; Tignor, M.; Miller, H.L. (Eds.) Cambridge University Press: Cambridge, UK, 2007; pp. 19–91.
- Bouwman, A.F. Soils and the Greenhouse Effect; John Wiley & Sons: Chichester, UK, 1990; pp. 61–627. [Google Scholar]
- Mosier, A.R.; Duxbury, J.M.; Freney, J.R.; Heinemeyer, O.; Minami, K. Assessing and mitigating N2O emissions from agricultural soils. Clim. Chang. 1998, 40, 7–38. [Google Scholar] [CrossRef]
- Abdalla, M.; Jones, M.; Ambus, P.; Williams, M. Emissions of nitrous oxide from Irish arable soils: effects of tillage and reduced N input. Nutr. Cycl. Agroecosyst. 2010, 86, 53–65. [Google Scholar] [CrossRef]
- Freney, J.R. Emission of nitrous oxide from soils used for agriculture. Nutr. Cycl. Agroecosyst. 1997, 49, 1–6. [Google Scholar] [CrossRef]
- Ghana National Inventory of Greenhouse Gases (2000). National Communication to the UNFCCC, p.3. Available online: http://unfccc.int/resource/docs/natc/ghanc1.pdf (accessed on 28 February 2012).
- Agboola, A.A.; Fayemi, A.A. Preliminary trials on the intercropping of maize with different tropical legumes in Western Nigeria. J. Agric. Sci. 1971, 77, 219–225. [Google Scholar] [CrossRef]
- Katsaruware, R.D.; Manyanhaire, I.O. Maize-cowpea intercropping and weed suppression in leaf stripped and detasseled maize in Zimbabwe. EJEAFChe 2009, 8, 1218–1226. [Google Scholar]
- Mariga, I.K. Effect of cowpea planting date and density on the performance of maize-cowpea intercrop. Zimb. J. Agric. Res. 1990, 28, 125–131. [Google Scholar]
- Olufajo, O.O.; Singh, B.B. Advances in Cowpea Cropping Systems Research. In Proceedings of the World Cowpea Conference 111, International Institute of Tropical Agriculture (IITA), Ibadan, Nigeria, 4–8 September, 2000.
- Adeniyan, O.N.; Ayoola, O.T.; Ogunleti, D.O. Evaluation of cowpea cultivars under maize and maize-cassava based intercropping systems. Afr. J. Plant Sci. 2011, 5, 570–574. [Google Scholar]
- Massawe, C.R.; Kaswende, J.S.; Mbwaga, A.M.; Hella, J.P. On-farm Verification of Maize/Cowpea Intercropping on the Control of Striga under Subsistence Farming. In Proceeding of 7th Eastern and Southern Africa Regional Maize Conference, 11–15 February 2001; pp. 165–167.
- Weier, K.I.; Doran, J.W.; Power, J.F.; Walters, D.T. Denitrification and the dinitrogen to nitrous oxide ratio as affected by soil water, available carbon and nitrate. Soil Sci. Soc. Am. J. 1993, 57, 66–72. [Google Scholar]
- Baggs, E.M.; Millar, N.; Ndufa, J.K.; Cadisch, G. Effect of Residue Quality on N2O Emissions from Tropical Soils. In Sustainable Management of Soil Organic Matter; Rees, R.M., Ball, B.C., Campbell, C.D., Watson, C.A., Eds.; CAB International: Edinburgh, UK, 2001; pp. 120–125. [Google Scholar]
- Millar, N.; Baggs, E.M. The chemical composition or quality of agroforestry residues influence N2O emissions after their addition to soils. Soil Biol. Biochem. 2004, 36, 935–943. [Google Scholar] [CrossRef]
- Millar, N.; Ndufa, J.K.; Cadisch, G.; Baggs, E.M. Nitrous oxide emissions following incorporation of improved-fallow residues in the humid tropics. Glob. Biogeochem. Cy. 2004. [Google Scholar] [CrossRef]
- Handayanto, E.; Cadisch, G.; Giller, K.E. Regulating N Mineralisation from Plant Residues by Manipulating Quality. In Driven by Nature: Plant Litter Quality and Decomposition; Cadisch, G., Giller, K.E., Eds.; CAB International: Wallingford, UK, 1997; pp. 175–185. [Google Scholar]
- Peoples, M.B.; Boyer, E.W.; Goulding, K.W.T.; Heffer, P.; Ochoh, V.A.; Vanlauwe, B.; Wood, S.; Yagi, K.; van Cleemput, O. Pathways of Nitrogen Loss and their Impacts on Human Health and the Environment. In Agriculture and the Nitrogen Cycle; Mosier, A.R., Syers, K.J., Freney, J.R., Eds.; The Scientific committee on Problems of the Environment (SCOPE) Island Press: Washington, DC. USA, 2004; pp. 53–69. [Google Scholar]
- Baggs, E.M.; Rees, R.M.; Smith, K.A.; Vinten, A.J.A. Nitrous oxide emission from soils after incorporation of crop residues. Soil Use Manag. 2000, 16, 82–87. [Google Scholar]
- Vityakon, P.; Meepech, S.; Cadisch, G.; Toomsan, B. Soil organic matter and nitrogen transformation mediated by plant residues of different qualities in sandy acid upland and paddy soils. Neth. J. Agric. Sci. 2000, 48, 75–90. [Google Scholar]
- Schwendener, C.M.; Lehman, J.; de Camargo, P.B.; Luizao, R.C.C.; Fernandez, E.C.M. Immobilisation and remineralisation of N following addition of wheat straw into soil: determination of gross transformation rates by 15N-ammonium isotope dilution technique. Soil Biol. Biochem. 2005, 37, 425–432. [Google Scholar] [CrossRef]
- Kaewpradit, W.; Toomsan, B.; Vityakon, P.; Limpinuntana, V.; Saenjan, P.; Jogloy, S.; Patanothai, A.; Cadisch, G. Regulating mineral N emission by mixing groundnut residues with rice straw under field conditions. Eur. J. Soil Sci. 2008, 59, 640–652. [Google Scholar] [CrossRef]
- Myers, R.K.J.; Palm, C.; Cueva, E.; Gunatileke, I.U.N.; Brossard, M. The Synchronisation of Nutrient Mineralisation and Plant Nutrient Demand. In The Biological Management of Tropical Soil Fertility; Woomer, P.L., Swift, M.J., Eds.; Wiley: Chichester, UK, 1994; pp. 81–116. [Google Scholar]
- Robertson, G.P. Nitrification and Denitrification in Humid Tropical Systems. In Mineral Nutrients in Tropical Forest and Savanna Ecosystems; Proctor, J., Ed.; British Ecological Society Special Publication, Blackwell Science: Malden, MA, USA, 1989; Volume 9, pp. 5–69. [Google Scholar]
- Eagle, A.J.; Bird, J.A.; Horwath, W.R.; Linquist, B.A.; Brouder, S.M.; Hill, J.E. Rice yield and nitrogen utilisation efficiency under alternative straw management practices. Agron. J. 2000, 92, 1096–1103. [Google Scholar] [CrossRef]
- Anderson, J.M.; Ingram, J.S.I. Tropical Soil Biology and Fertility: A Handbook of Methods, 2nd ed; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Parton, W.J.; Mosier, A.R.; Ojima, D.S.; Valentine, D.W.; Schimel, D.S.; Weier, K.; Kulmala, A.E. Generalised model for N2 and N2O production from nitrification and denitrification. Glob. Biogeochem. Cy. 1996, 10, 401–412. [Google Scholar] [CrossRef]
- Davidson, E.A.; Hart, S.C.; Shanks, C.A.; Firestone, M.K. Measuring gross nitrogen mineralization, immobilization and nitrification by 15N isotopic pool dilution in intact soil cores. J. Soil Sci. 1991, 42, 335–349. [Google Scholar]
- Huang, Y.; Zou, J.W.; Zheng, X.H. Nitrous oxide emissions as influenced by amendments of plant residues with different C:N ratios. Soil Biol. Biochem. 2004, 36, 973–981. [Google Scholar] [CrossRef]
- Gentile, R.; Vanlauwe, B.; Chivenge, P.; Six, J. Interactive effects from combining fertilizer and organic residue inputs on nitrogen transformations. Soil Biol. Biochem. 2008, 40, 2375–2384. [Google Scholar] [CrossRef]
- Khalil, M.I.; Rosenanin, A.B.; van Cleemput, O.; Fauziah, C.I.; Shamsuddin, J. Nitrous oxide emissions from an ultisol of the humid tropics under maize-groundnut rotation. J. Environ. Qual. 2002, 31, 1071–1078. [Google Scholar] [CrossRef]
- Garcia-Ruiz, R.; Baggs, E.M. N2O emissions from soil following combined application of fertilizer-N and ground weed residues. Plant Soil 2007, 299, 263–274. [Google Scholar] [CrossRef]
- Mole, S.; Waterman, P.G. Tannic acid and proteolytic enzymes: Enzyme inhibition or substrate deprivation? Phytochemistry 1986, 26, 99–102. [Google Scholar]
- Palm, C.A.; Gachengo, C.N.; Delve, R.J.; Cadisch, G.; Giller, K.E. Organic inputs for soil fertility management in tropical agro-ecosystems: application of an organic resource database. Agr. Ecosyst. Environ. 2001, 83, 27–42. [Google Scholar] [CrossRef]
- Constantinides, M.; Fownes, J.H. Nitrogen mineralization from leaves and litter of tropical plants: relationship to nitrogen, lignin and soluble polyphenol concentrations. Soil Biol. Biochem. 1994, 26, 49–55. [Google Scholar] [CrossRef]
- Palm, C.A.; Rowland, A.P. A Minimum Dataset for Characterization of Plant Quality for Decomposition. In Driven by Nature: Plant Litter Quality and Decomposition; Cadisch, G., Giller, K.E., Eds.; CAB International: Wallingford, UK, 1997; pp. 379–392. [Google Scholar]
- Azam, F.; Simmons, F.W.; Mulvaney, R.L. Mineralisation of N from plant residues and its interaction with native soil N. Soil Biol. Biochem. 1993, 25, 1787–1792. [Google Scholar] [CrossRef]
- Bird, J.A.; Horwath, W.R.; Eagle, J.A.; van Kessel, C. Immobilization of fertilizer nitrogen in rice: effect of straw management practices. Soil Sci. Soc. Am. J. 2001, 65, 143–1152. [Google Scholar]
- Heal, O.W.; Anderson, J.M.; Swift, M.J. Plant Litter Quality and Decomposition: An Historical Overview. In Driven by Nature: Plant Litter Quality and Decomposition; Cadisch, G., Giller, K.E., Eds.; CAB International: Wallingford, UK, 1997; pp. 3–30. [Google Scholar]
- Tiedje, J.M.; Sextone, A.J.; Parkin, T.B.; Revbech, N.P.; Shelton, D.R. Anaerobic processes in soils. Plant Soil 1984, 76, 117–212. [Google Scholar]
- Sarkodie-Addo, J.; Lee, H.C.; Baggs, E.M. Nitrous oxide emissions after application of inorganic fertilizer and incorporation of green manure residues. Soil Use Manag. 2003, 19, 331–339. [Google Scholar] [CrossRef]
- Gartner, T.B.; Cardon, Z.B. Decomposition dynamics in mixed species leaf litter. Oikos 2004, 104, 230–246. [Google Scholar] [CrossRef]
- Quemada, M.; Cabrera, M.L. Carbon and nitrogen mineralised from leaves and stems of four cover crops. Soil Sci. Soc. Am. J. 1995, 59, 471–477. [Google Scholar] [CrossRef]
- Mafongoya, P.L.; Giller, K.E.; Palm, C.A. Decomposition and nitrogen release patterns of tree prunnings and litter. Agroforest. Syst. 1998, 38, 77–97. [Google Scholar]
- Chapman, K.; Whittaker, J.B.; Heal, O.W. Metabolic and faunal activity in litters of tree mixtures compared with pure stands. Agric. Ecosyst. Environ. 1988, 24, 33–40. [Google Scholar] [CrossRef]
- Wardle, D.; Lavelle, P. Linkages between soil biota, plant litter quality and decomposition. In Driven by Nature: Plant Litter Quality and Decomposition; Cadisch, G., Giller, K.E., Eds.; CAB International: Wallingford, UK, 1997; pp. 107–124. [Google Scholar]
- Firestone, M.K.; Davidson, E.A. Microbiological Basis of NO and N2O Production and Consumption in soil. In Exchange of Trace Gases between Terrestrial Ecosystems and the Atmosphere; Andreae, M.O., Schimel, D.S., Eds.; John Wiley and Sons: Chichester, UK, 1989; pp. 7–21. [Google Scholar]
- Baggs, E.M. A review of stable isotope techniques for N2O source partitioning in soils: recent progress, remaining challenges and future considerations. Rapid Commun. Mass. Spectrom. 2008, 22, 1664–1672. [Google Scholar] [CrossRef]
- Pansu, M.; Thuriès, L.; Larré-Larroy, M.C.; Bottner, P. Predicting N transformations from organic inputs in soil in relation to incubation time and biochemical composition. Soil Biol. Biochem. 2003, 35, 353–363. [Google Scholar] [CrossRef]
- Zech, W.; Senesi, N.; Guggenberger, G.; Kaiser, K.; Lehman, J.; Miano, T.M.; Miltner, A.; Schroth, O. Factors controlling humification in the tropics. Geoderma 1997, 79, 117–161. [Google Scholar] [CrossRef]
- Skopp, J.; Jawson, M.D.; Doran, J.W. Steady-state aerobic microbial activity as a function of soil water content. Soil Sci. Soc. Am. J. 1990, 54, 1619–1625. [Google Scholar] [CrossRef]
- Vanlauwe, B.; Sanginga, N.; Merkcx, R. Decomposition of four Leucaena and semma prunnings in alley cropping systems under sub-humid tropical conditions: the process and its modifiers. Soil Biol. Biochem. 1997, 29, 131–137. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Frimpong, K.A.; Yawson, D.O.; Agyarko, K.; Baggs, E.M. N2O Emission and Mineral N Release in a Tropical Acrisol Incorporated with Mixed Cowpea and Maize Residues. Agronomy 2012, 2, 167-186. https://doi.org/10.3390/agronomy2030167
Frimpong KA, Yawson DO, Agyarko K, Baggs EM. N2O Emission and Mineral N Release in a Tropical Acrisol Incorporated with Mixed Cowpea and Maize Residues. Agronomy. 2012; 2(3):167-186. https://doi.org/10.3390/agronomy2030167
Chicago/Turabian StyleFrimpong, Kwame A., David O. Yawson, Kofi Agyarko, and Elizabeth M. Baggs. 2012. "N2O Emission and Mineral N Release in a Tropical Acrisol Incorporated with Mixed Cowpea and Maize Residues" Agronomy 2, no. 3: 167-186. https://doi.org/10.3390/agronomy2030167