
Impact of Hood Steaming on Tuber Vitality of Yellow Nutsedge (Cyperus esculentus)
by
 , ,
, ,
Jeroen Feys
1,*,
Sander De Ryck
1,
Clara Sciffer
2 ,
,
Dirk Reheul
1,
Joos Latré
3,
Danny Callens
4 and
Benny De Cauwer
1 1
Weed Science Unit, Department of Plants and Crops, Faculty of Bioscience Engineering, Ghent University, 9000 Gent, Belgium
2
Flanders Research Institute for Agriculture, Fisheries and Food (ILVO), 9820 Merelbeke, Belgium
3
Biosciences and Food Sciences Department, Faculty of Science and Technology, University College Ghent, Research Station HoGent-UGent, 9820 Bottelare, Belgium
4
Research and Advice Centre for Agriculture and Horticulture (INAGRO VZW), 8800 Rumbeke-Beitem, Belgium
*
Author to whom correspondence should be addressed.
Agronomy 2024, 14(5), 918; https://doi.org/10.3390/agronomy14050918
Submission received: 19 March 2024
/
Revised: 23 April 2024
/
Accepted: 24 April 2024
/
Published: 26 April 2024
(This article belongs to the Section Weed Science and Weed Management)
Abstract
:Cyperus esculentus is a hard-to-control, destructive perennial weed propagating and spreading through rhizomes and tubers. Currently, a combination of mechanical, cultural, and chemical measures sustained over time is required for satisfactory control of arable crops. Hood steaming is a promising thermal technique for rapid depletion of the soil tuber bank. The effect of hood steaming on C. esculentus tuber vitality was investigated using a dose–response experiment on two localities in Belgium. Steaming was performed for five durations (2, 4, 8, 16, and 32 min). Moreover, the effects of tuber burial depth (5, 15, and 25 cm), genetic C. esculentus clone (three different clones), and pedohydrological parameters (sandy and sandy, loamy soil) on hood steaming efficacy were determined. Additionally, the heat sensitivity of genetically diverse C. esculentus tubers was investigated in a laboratory incubator experiment in which incubation temperature and duration varied. To control tubers at depths of 5, 15, and 25 cm, treatment durations of 8, 16, and 32 min were required, respectively. In general, clones producing large tubers showed lower heat sensitivity. As pedohydrological parameters may affect heat transfer into the soil, they may affect steaming efficacy as well. To obtain complete control of C. esculentus tubers, soil temperature should be 50 °C or more for at least 42 min. Hood steaming is a highly suitable alternative technique for a rapid and strong depletion of the soil tuber bank in small well-delineated C. esculentus patches.
1. Introduction
Cyperus esculentus L. (yellow nutsedge) is a perennial weed that is very prolific and hard to control in many crops. It is considered one of the most troublesome weeds in the world [1] and can lead to huge losses in arable crops (e.g., 60% in sugar beets [Beta vulgaris L.] and 40% in potatoes [Solanum tuberosum L.]) [2]. High infestation levels (80–100% of the field covered with C. esculentus shoots) led to yield losses of 86%, 90%, and 93% in leek [Allium porrum L.], onions [Allium cepa L.], and Brussels sprouts [Brassica oleracea var. gemmifera (de Candolle) Zenker], respectively [3]. In maize (Zea mays L.), every 100 shoots per square meter is expected to cause a yield loss of 8% [4].
Cyperus esculentus is a cosmopolitan weed but prefers tropical and subtropical regions [5]. Reproduction occurs mainly via tubers and rhizomes. In Western Europe, mother tubers start to germinate during April and initiate a process of intensive shoot formation. De Cauwer et al. [6] planted C. esculentus mother tubers in a 10 L pot (1 tuber per pot). At the end of the growing season, 29 to 91 shoots were counted. Formation of daughter tubers starts in May or June and continues till autumn. In regions with a temperate climate, one mother tuber can generate 700 or more daughter tubers during one growing season [7]. Tubers can survive low temperatures (−10 °C on the soil surface), and the most persistent tubers may remain germinative for 10 years [8]. Cyperus esculentus also produces viable seeds, but few to no seedlings are found in fields under outdoor conditions [9]. Seedlings would probably lack the vigor to survive under field conditions [10,11].
In Belgium, over 50,000 hectares of cropland are infected with C. esculentus (J Feys pers. comm.). Successful control of C. esculentus tuber banks in cropped areas requires years of intensive control. Because of the insufficient efficacy of single control measures against C. esculentus, integrated weed management systems combining cultural, mechanical, and chemical control measures are required [7,12,13]. Moreover, curative management systems providing effective control in the short term are lacking.
So far, thermal techniques to reduce the C. esculentus tuber bank, such as electrocution, soil steaming, and soil solarization are poorly implemented in practice. Nevertheless, C. esculentus tubers are highly sensitive to heat [14]. With exposure to hot air for 16 h, tubers are killed at an air temperature of 50 °C. At temperatures of 55 °C and 60 °C, tubers are killed after an exposure of 8 h and 2 h, respectively. Soil steaming is considered the most promising thermal method, as one single treatment may potentially lead to 100% control of the tuber bank. This is clearly not the case for electrocution, where multiple treatments and a season-long black fallow are required to obtain a complete depletion of the tuber bank [15]. Soil solarization is another thermal technique, but its efficacy decreases rapidly with increasing soil depth. For example, at the depths of 2.5 and 15 cm, seed germination of Portulaca oleracea L. (common purslane) after solarization was 23.3 and 55.9%, respectively [16]. As a result, soil solarization is deemed ineffective in reducing the soil tuber bank of C. esculentus, as C. esculentus tubers may occur up to a depth of 45 cm [10,17].
There are several types of soil steaming. Sheet steaming is an application technique in which steam is released under a heat-resistant steam foil that covers the soil surface. Hood steaming is very similar to sheet steaming, but in this method steam is released under metal hoods that do not stay in place after steaming. Other steaming techniques directly inject the steam into the soil by using a grid of long perforated metal spikes that are pushed into the soil and connected with a steam source [18]. During hood steaming, multiple zones are created in the soil (Figure 1). The upper zone (the topsoil) becomes filled by overheated steam and eventually no longer contains liquid water. The second ‘two-phase’ zone contains both liquid water and water vapor. In this zone, both the processes of evaporation and condensation occur. The deeper in this zone, the higher the water saturation, until a condensation front is formed. The condensation front, which moves downwards during the hood steaming process, forms the border with the next, third zone where the soil is completely water-saturated. Irrespective of the soil texture, the condensation front in relatively dry soils (moisture content about 40% of field capacity) is located near the surface. In moist soils (moisture content about 80% of field capacity), the condensation front is located between 5 and 16 cm. In the zone beyond the condensation front (the third zone), heat can only be transferred by conduction. In the fourth, deepest soil zone, soil temperature is not affected by steaming [18,19].
The parameter ‘soil heat flux’ describes the amount of thermal energy that moves through an area of soil during a unit of time [20]. During the steaming process, the heat flux through the soil is dependent on the heat exchange between the solid, liquid, and gas phases of the soil. The heat flux may be affected by several pedohydrological parameters. In the experiment of Melander and Kristensen [21], higher (on average 6–7 °C) temperatures were needed to obtain a similar control rate of seedlings when steaming was performed on a coarse soil containing 27.2% large (>6 mm) aggregates compared to a fine soil containing 16.1% large (>6 mm) aggregates. Gay et al. [18] performed hood steaming on sandy and sandy loamy soils differing in moisture content. Heat transfer to deeper soil layers was faster in soil with a moisture content of about 80% of field capacity compared to soil with a moisture content of about 40% of field capacity.
The effect of some soil steaming systems on C. esculentus tuber vitality has been assessed by different authors. In Switzerland, three steaming methods were compared [22], namely in situ hood steaming (hood with 30 cm long steam injector spikes that were pushed into the soil, steaming duration 15 min), in situ sheet steaming (without steam injection, steaming duration 6 to 8 h), and ex situ steaming using a steam station (capacity of 6 to 10 square meters of infested soil per hour [23]) in which excavated C. esculentus-infested soil was steamed. The steam station was placed on the farm, and the infected soil needed to be excavated and transported to the farm. They concluded that steaming is highly effective in reducing shoot emergence. No new C. esculentus shoots emerged after hood steaming and ex situ steaming with the steam station. However, the absence of shoot formations does not automatically mean that tubers were dead, as steaming might have induced tuber dormancy. For the sheet steaming method, the researchers reported a shoot emergence reduction of 95% relative to an untreated control. Most likely, sheet steaming was less suitable for heating deeper soil layers (>25 cm) that may still contain tubers. These tubers could possibly even be activated by the heat [22]. Also, the effect of steaming with spikes has been investigated [24]. Spikes were inserted at full length (20 cm) into the soil and were connected to a steam generator. The treated beds were covered with a whitened plastic film to minimize solarization effects. Steaming reduced C. esculentus tuber vitality by 96% relative to untreated control tubers (vitality of 31.9%). Tuber burial depth or burial location (in the central zone or near plot edges) did not affect the efficacy of either method. Despite its effectiveness, in situ soil steaming remains poorly implemented. This may be due to the high application costs. For example, the fuel consumption ranges between 3000 and 5000 L ha−1 [25]. Moreover, 70 to 100 h are needed to treat 1 hectare. Additionally, steaming leads to a large reduction in (or even elimination of) soil living organisms. A steaming treatment may lead to a flush in carbon mineralization and a shift in the genetic structure of the soil bacterial community. However, these strong impacts are quickly reversible, which enables farmers to sow a crop approximately 1 week after treatment [26]. Nevertheless, steaming could be particularly useful for the control of small C. esculentus patches on croplands and for use in cropped or non-cropped areas where chemical methods are not allowed (e.g., near water bodies or in nature conservation areas). To increase the cost-effectiveness and robustness of steaming, additional knowledge is required about the critical energy dose needed to kill tubers and the impact of biological and pedohydrological factors on the efficacy of soil steaming against C. esculentus. In the literature, there are no data available about the heat sensitivity of different C. esculentus clones. Indeed, there is a lot of genetic diversity within this species, as shown by De Ryck et al. [27]. Moreover, the potential effects of the parameters of soil texture, soil moisture content, and tuber burial depth will be assessed. Therefore, the following hypotheses were formulated: (H1) the efficacy of hood steaming depends on the genetic C. esculentus clone, (H2) the efficacy of hood steaming depends on the tuber burial depth, and (H3) the efficacy of hood steaming depends on pedohydrological parameters, namely soil texture and soil moisture content.
To test these hypotheses, a field experiment was set up in which the effects of the tuber burial depth and the clonal variation on the efficacy of steaming were investigated on sandy loam soil and sandy soil. To assess potential year effects, the experiment was performed in two consecutive years, namely in 2020 and 2021. The experiment was a dose–response experiment to determine the optimal duration of steaming. During the steaming treatments, soil temperature was monitored to quantify the heat sensitivity of the different C. esculentus clones. The heat sensitivity of genetically diverse C. esculentus tubers was also investigated in a laboratory incubator experiment, with the genetic clone being the only variable factor. In this experiment, tubers were thermally treated by exposing them to various incubation temperatures and durations.
2. Materials and Methods
2.1. Experiments
2.1.1. Field Experiment
Experimental Sites
In the spring of 2020 and 2021, the efficacy of steaming was evaluated by performing two dose–response field experiments at Melle (3°49′ E, 50°58′ N) and Sint-Niklaas (4°11′ E, 51°11′ N), Belgium. The distance between the two locations is approximately 35 km. Location Melle has a sandy loam soil (8.6% clay, 51.6% loam, and 39.9% sand) with an organic carbon content of 1.1% and a pHKCl of 5.65. Location Sint-Niklaas has a sandy soil (6.2% clay, 10.3% loam, and 83.3% sand) with an organic carbon content of 1.14% and a pHKCl of 5.2. The gravimetric soil moisture contents at different depths (corresponding to the tuber burial depths; see below) at both locations are given in Table 1.
Steaming Equipment (Figure 2)
In our experiments, hood steaming was performed with a Polyvap 2000 CE machine (Simox, Contamine-sur-Arve, France). The machine consists of a steam generator working at low pressure (30,000–40,000 Pa) and a water tank of 300 L. The steam generator produces overheated steam of 120 °C. Then, the steam generated by the boiler is conveyed through a flexible hose to an aluminum hooded steam application unit with a surface of 1 m2 and a height of 15 cm. Prior to steaming, the hood walls are pushed 3 cm deep into the soil to force the steam released under the hood to move into the soil and to avoid steam losses to the air. The machine is also equipped with an alternator (Sincro ER2CAT, Soga Energy Team, Montecchio Maggiore, Italy), producing single-phase electric power of 230 V and 50 Hz. The whole machine is constructed on a trailer which can be easily pulled with a tractor or automobile [28,29]. Fuel consumption (diesel with a heating value of 43.8 MJ kg−1 and density of 0.855 kg L−1 [30]) during steaming was 4.6 L h−1. The water consumption was 70 L h−1.
Figure 2.
(Left): The steam generator, water tank, and flexible hose connected between the steam generator and the hooded application unit. (Right): The aluminum hooded application unit.
Figure 2.
(Left): The steam generator, water tank, and flexible hose connected between the steam generator and the hooded application unit. (Right): The aluminum hooded application unit.

Experimental Set-Up
On each experimental site, a dose–response experiment was carried out in which tubers of three C. esculentus clones (Ardooie, Bree, and Meulebeke) were buried at three different soil depths (5, 15, and 25 cm) and exposed to six different steaming durations. This dose–response field experiment was run twice, namely in 2020 and 2021. Each combination of clone, burial depth, and steaming duration was laid down in four replicates. An experimental unit consisted of a buried nylon bag with 25 C. esculentus tubers of a particular clone. Each nylon bag measured 30 cm × 10 cm and was handmade using a gas- and water-permeable woven Suzuki fruit fly net (Duranet, Oostende, Belgium) with a mesh size of 0.97 mm × 0.83 mm.
- Genetic clone
The genetic distinctness of the chosen clones, namely Ardooie, Bree, and Meulebeke, was confirmed by De Ryck et al. [27]. Clones were named after the place in Belgium where they were originally sampled. Only medium-sized tubers with a weight falling between 80% and 120% of the clone-specific mean tuber weight were used. Tubers of the Ardooie, Bree, and Meulebeke clones had mean fresh tuber weights of 217.1, 261.0, and 456.4 mg, respectively. The tubers were produced and harvested the year before the experimental set-up (thus, in 2019 and 2020). After the harvest, they were stored at 5 °C until used. Prior to packaging in the nylon bags, tubers were imbibed in tap water for 12 h. To avoid creating preferential pathways for gas or fluid flow inside the bags (e.g., via macropores), tubers were intermingled with 500 mL of original crumbled soil.
- Burial depth
Prior to steaming, the soil was tilled to a depth of 30 cm using a deep tine cultivator with a knife roller. Deep tillage crumbles and aerates the soil thus fostering steam penetration into the soil. After soil tillage, 6 plots (1 m2, 1 plot for each steam duration) were demarcated on the field. Then, three gullies were dug in each plot. The gullies had depths of 5, 15, and 25 cm (checked by a ruler) and a width of 22 cm. Then, the nylon bags filled with C. esculentus tubers were horizontally laid down in the gullies (Figure 3).
In each plot, 36 nylon bags (3 gullies × 3 clones × 4 repeats) were buried. After laying the nylon bags in the gullies, the gullies were filled up again with the original soil. Thereafter, the steam treatment was performed on each of the 6 plots. The nylon bags were exhumed 1 h after the end of the treatment.
- Steaming duration
Six different steaming durations were applied (1, 2, 4, 8, 16, and 32 min at location Melle, and 2, 4, 8, 16, 32, and 64 min at location Sint-Niklaas). The steam durations chosen in this experiment were based on preliminary work aimed at determining the steam duration range that allows fitting dose–response curves and calculating the minimum steam duration required for 90% control of C. esculentus tubers at different depths. Moreover, for each genetic clone, an untreated control was included. These tubers were stored in the fridge at 5 °C until vitality assessment.
2.1.2. Laboratory Experiment
Incubator
The experiment was performed by using a general-purpose incubator (model 1535) produced by Sheldon Manufacturing Inc.® (Cornelius, OR, USA). The incubator contains 5 heating elements which are spatially distributed to guarantee a uniform temperature in the incubator. The inner height, width, and depth of the incubator are 51 cm, 61 cm, and 61 cm, respectively. Temperatures can be set with a precision of 0.1 °C and a maximum of 65 °C [31]. The experiment took place in 2021, from July 13 to July 15, in Ghent, Belgium.
Experimental Set-Up
The experiment was a dose–response experiment in which tubers of 3 particular C. esculentus clones (Ardooie, Bree, and Oostkamp) were heated at 4 particular temperatures (50, 55, 60, and 65 °C) for 6 durations of exposure. For each combination of genetic clone, temperature, and exposure duration, 3 repeats were foreseen. An experimental unit consisted of a petri dish (LLG-Petri dish PS, diameter 90 mm, LLG, Meckenheim, Germany) with 15 C. esculentus tubers of a particular C. esculentus clone. The petri dishes were prepared 24 h before the treatment. Preparation was performed as described by Webster [14]. A total of 15 tubers of a particular clone were placed between two filter papers (LLG-Rundfilter, diameter 90 mm, LLG, Meckenheim, Germany). Then, 5 mL of water was added to each petri dish to simulate the hydrated conditions in the soil. Thereafter, petri dishes were sealed airtight with tape (Petri Seal® type K427.1, Carl Roth GmbH+Co. KG, Karlsruhe, Germany). After preparation, the petri dishes were placed in a climate chamber (17.3 °C) until the treatment.
- Genetic clone
The clones Ardooie and Bree were the same clones as the ones tested in the field experiment. Clone Oostkamp replaced clone Meulebeke, but both clones produce large tubers and belong to the same genetic cluster, as shown by De Ryck et al. [27]. The clones Ardooie, Bree, and Oostkamp had mean fresh tuber weights of 217.1, 261.0, and 500.0 mg, respectively. Only medium-sized tubers with weight falling between 80 and 120% of the clone-specific mean weight were selected. The tubers were produced and harvested in 2020 and were kept at 5 °C until used.
- Exposure duration and temperature
During the hot air treatment, the petri dishes with tubers were subjected to 50, 55, 60, and 65 °C for 6 temperature-specific exposure durations (Table 2).
2.2. Measurements
2.2.1. Soil Temperature
In the field experiment, continuous measurements of soil temperatures at the three burial depths of 5, 15, and 25 cm were performed from the start of the steaming duration until one hour after the end of the steaming duration. For this purpose, 3 sensors (Onset 12-bit temperature smart sensors, Onset, Bourne, MA, USA) connected to a data logger (Data logger micro station, Forestry Suppliers, Jackson, MS, USA) were used. Sensors were carefully buried near the nylon bags. These sensors monitored the temperature every second and had a measuring range between −40 °C and 100 °C.
2.2.2. Tuber Sprouting Assessment
In both experiments, tuber sprouting was assessed after the treatments. For this purpose, the tubers were planted 2 cm deep in 150-cell trays. The substrate in the trays was a 1:1 mixture of potting soil and steamed sandy loam soil. The sandy loam soil contained 10.0% clay, 46.7% silt, and 43.3% sand. The organic matter content was 2.6%, and the pHKCl was 5.5. After planting, the trays were placed under a rain shelter greenhouse with sprinkler irrigation. Depending on daily water evapotranspiration, the irrigation varied between 2.5 and 3.8 mm day−1.
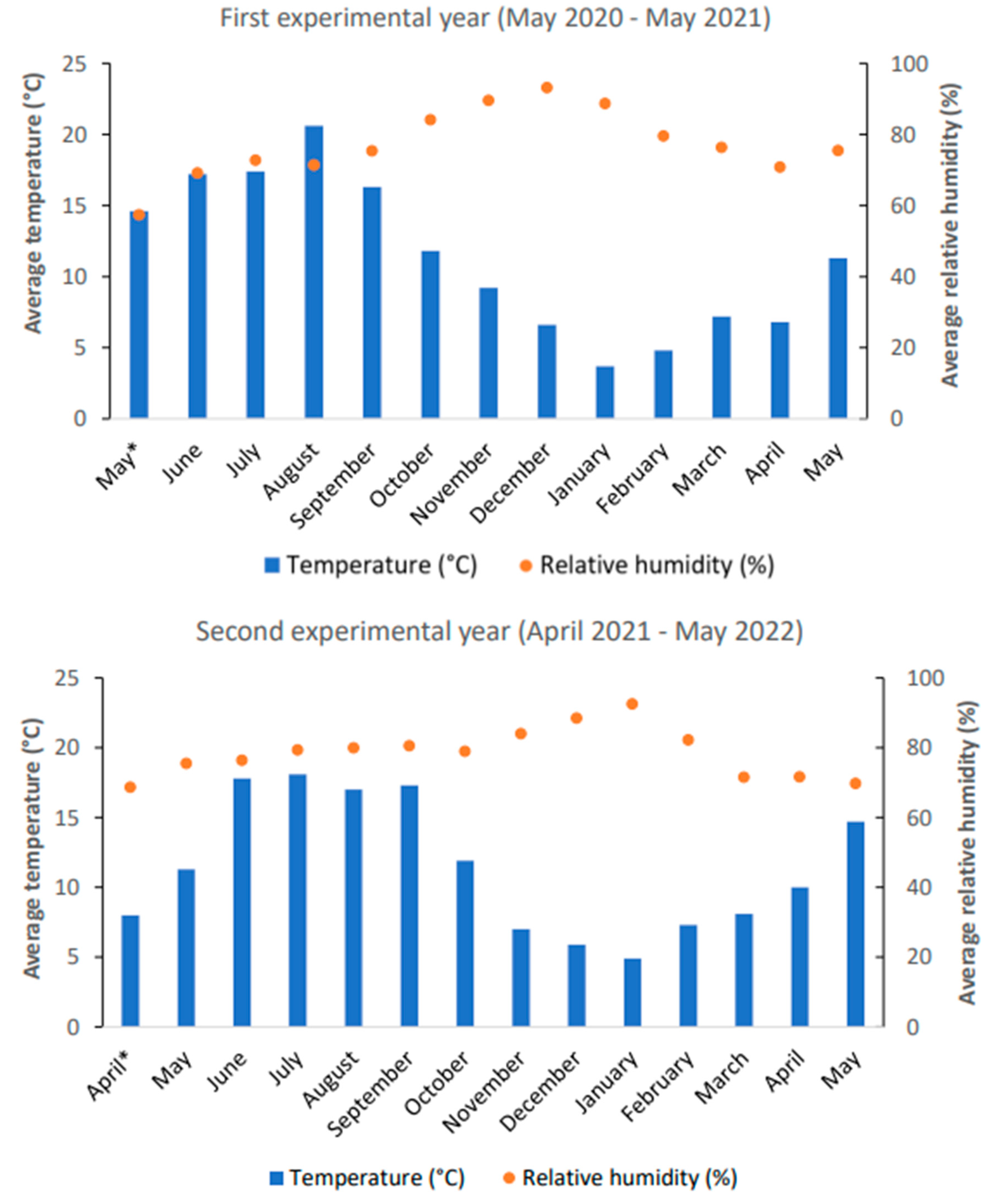
During the screening period (12 months), all cells were monitored weekly for sprouting tubers. Sprouted tubers were counted and removed from the trays. At the end of the screening period, a vitality percentage could be calculated for each experimental unit. The vitality percentage was calculated by dividing the number of sprouted tubers by the total number of tubers (25 for the field experiment, 15 for the laboratory experiment) and multiplying the result obtained by 100. The climatic conditions in the greenhouse during the 12-month screening periods in 2020–2021 and 2021–2022 are given in Figure 4. During the winter months, the trays were kept free of frost.
2.3. Statistical Data Analysis
All data were analyzed in RStudio, version 4.1.3 [32]. All data were analyzed using parametric or non-parametric tests run at the 5% significance level. The obtained data from our dose–response experiments were analyzed with the drc package [33]. The variable tuber vitality was the dependent variable. The factors of steaming duration (field experiment) and incubation duration (laboratory experiment) were the independent variables. The assumptions of normality and homoscedasticity were checked with a Q-Q plot and a Levene test, respectively. Thereafter, the construction of dose–response curves was performed according to Streibig et al. [34]. Based on these curves, ED90 values and selectivity indices were determined using the delta method [35]. The ED90 value is the required dose to obtain a 90% reduction in tuber vitality (compared to the untreated control). The higher this value, the lower the efficacy of the applied treatments. The selectivity index is a parameter that can be used to compare two dose–response curves. It is the ratio between the ED90 for one dose–response curve and the ED90 for another dose–response curve. In the field experiment, this index was used to determine whether the factors of genetic clone, burial depth, and location affect the efficiency of hood steaming. In the laboratory experiment, this factor was used to determine whether the factor genetic clone affects the efficiency of the hot air treatment.
3. Results
3.1. Field Experiment
3.1.1. Monitoring of Soil Temperature
At both experimental sites and in both experimental years, the temperature was measured at the burial depths of 5, 15, and 25 cm during and until one hour after each treatment. Moreover, during each treatment, it was calculated for how long the temperature exceeded 50, 65, 80, and 95 °C. These results are given in Figure 5 and Table 3.
At both locations and in both years, 2 min of steaming led to a limited heating of the upper soil layer (5 cm deep). The maximum observed temperatures were 23.4 and 35.7 °C at Melle (2020) and Sint-Niklaas (2020), respectively. At the deeper soil layers (15 and 25 cm deep), almost no heating was observed.
When steaming was performed for 4 min, the heating effect at 5 cm depth was restricted at Melle, where the temperature did not exceed 50 °C in both experimental years. At Sint-Niklaas, however, the temperature in the upper soil layer quickly increased, especially in 2020. In 2020, the temperature exceeded the thresholds of 50 °C and 65 °C for 2260 and 780 s, respectively. At the deeper soil layers, the heating effect was still restricted or absent at both locations and in both experimental years.
A treatment of 8 min led to a strong heating of the upper soil layer. For example, at Sint-Niklaas, the temperature exceeded 95 °C for 523 and 267 s in 2020 and 2021, respectively. Remarkably, at Melle, the threshold of 95 °C was exceeded only in 2020 (for 409 s). At the depths of 15 and 25 cm, the temperature did not exceed 50 °C at both locations and in both experimental years.
A treatment of 16 min led to very high temperatures in the upper soil layer. At both locations and in both experimental years, the temperature was higher than 95 °C for at least 855 s. At 15 cm depth, the heating effect was much stronger at Melle. For example, the temperature at Melle (in 2020) exceeded 95 °C for 451 s, while at Sint-Niklaas, the maximum observed temperature was only 52.4 °C. At 25 cm depth, the heating effect was restricted at both locations and in both experimental years.
The longest steaming duration (32 min) led to strong heating effects at depths of 5 and 15 cm. At a depth of 5 cm, the exceedance of the 95 °C threshold varied between 2220 and 2698 s, depending on the location and the experimental year. In 2020, at a depth of 15 cm, the threshold of 95 °C was exceeded for 1692 and 1418 s at Melle and Sint-Niklaas, respectively. In 2021, the heating effect at 15 cm depth was even more pronounced. In the deepest soil layer (25 cm), a differential heating effect between the years was observed. For example, in 2020, the temperature exceeded 95 °C for 47 s at Melle. In 2022, the temperature exceeded 95 °C for 670 s at Melle.
3.1.2. Tuber Vitality Measures
Effect of Location
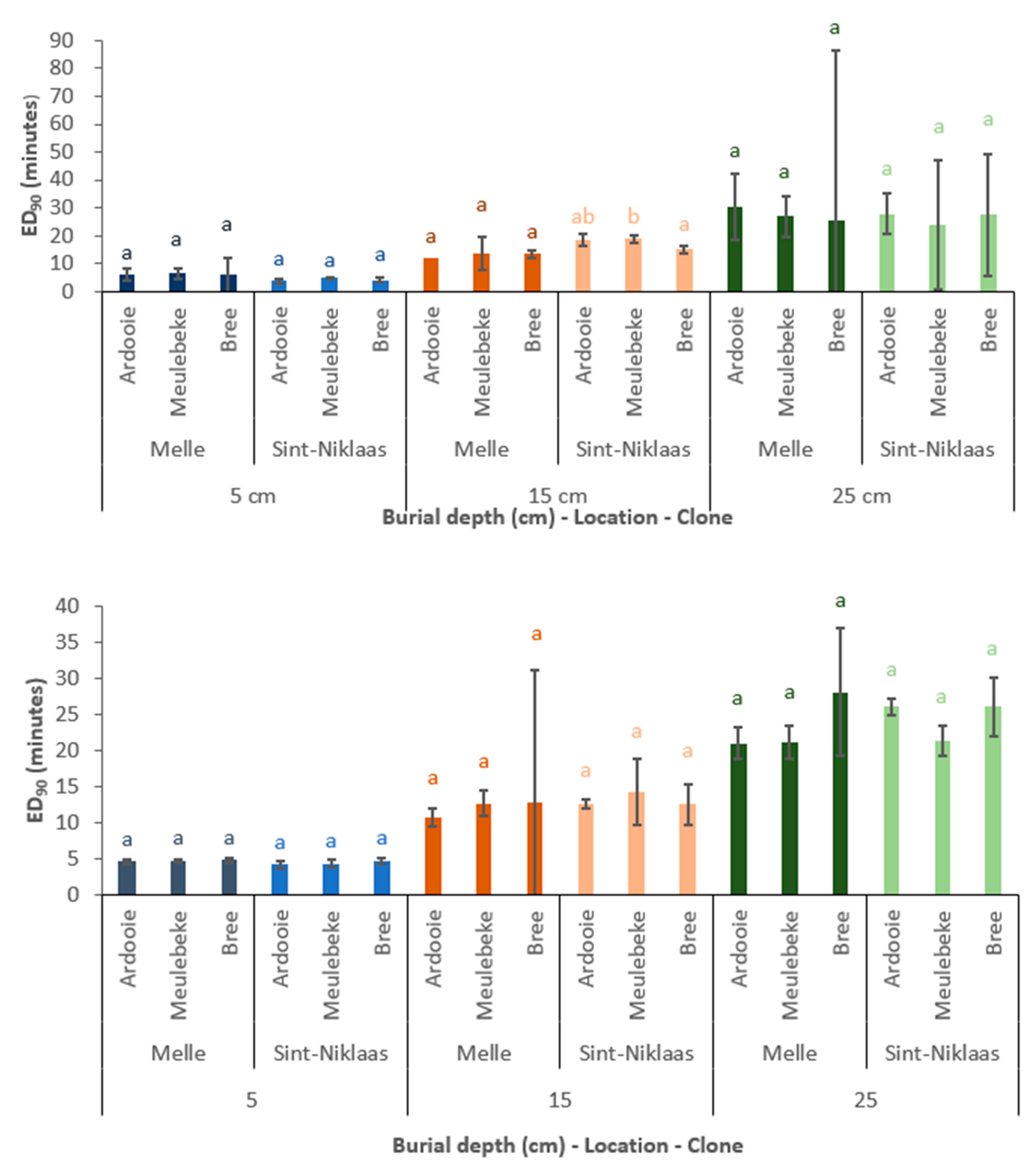
In both experimental years, for each unique combination of clone, burial depth, and location, an ED90 value was determined. In Figure 6, the ED90 values are compared among locations.
Figure 6 suggests that the factor location (linked with the factors of soil texture and soil moisture content) affects the ED90 value to a very limited extent. In the second experimental year (2021), no significant differences in ED90 values were observed among locations. This was also observed in the first experimental year (2020) at the burial depths of 15 and 25 cm. At a burial depth of 5 cm, the ED90 value is higher (1.63 to 1.89 min) in sandy, loamy soil (Melle) compared to sandy soil (Sint-Niklaas). However, this difference was statistically significant only for the Meulebeke clone.
Effect of Burial Depth
The factor of burial depth clearly affects the ED90 value (Figure 7). At a burial depth of 15 cm compared to 5 cm, the increase in ED90 value varied between 5.68 min and 13.73 min. For 10 of the 12 unique combinations of experimental year, clone, and location, this increase in ED90 value was statistically significant. When the burial depth further increased from 15 to 25 cm, the increase in ED90 value varied between 5.92 and 15.21 min. However, only in the second experimental year (2021) were these differences statistically significant. The absence of statistical differences in 2020 might be explained by the high standard errors of the ED90 values at 25 cm depth. In the first experimental year (2020), the ED90 values averaged over clones and locations were 5.4, 14.9, and 25.6 min at burial depths of 5, 15, and 25 cm, respectively. In the second experimental year (2021), these ED90 values were 4.8, 13.3, and 25.8 min, respectively.
Effect of Genetic Clone
As illustrated by Figure 8, there is a limited effect of the factor clone on the determined ED90 values. Within 11 of the 12 combinations of burial depth, location, and experimental year, the clone did not affect the ED90 value. However, in 2020, at location Sint-Niklaas and a burial depth of 15 cm, the ED90 value of clone Meulebeke was significantly higher than the ED90 value of clone Bree (18.79 and 15.01 min, respectively).
3.2. Laboratory Experiment
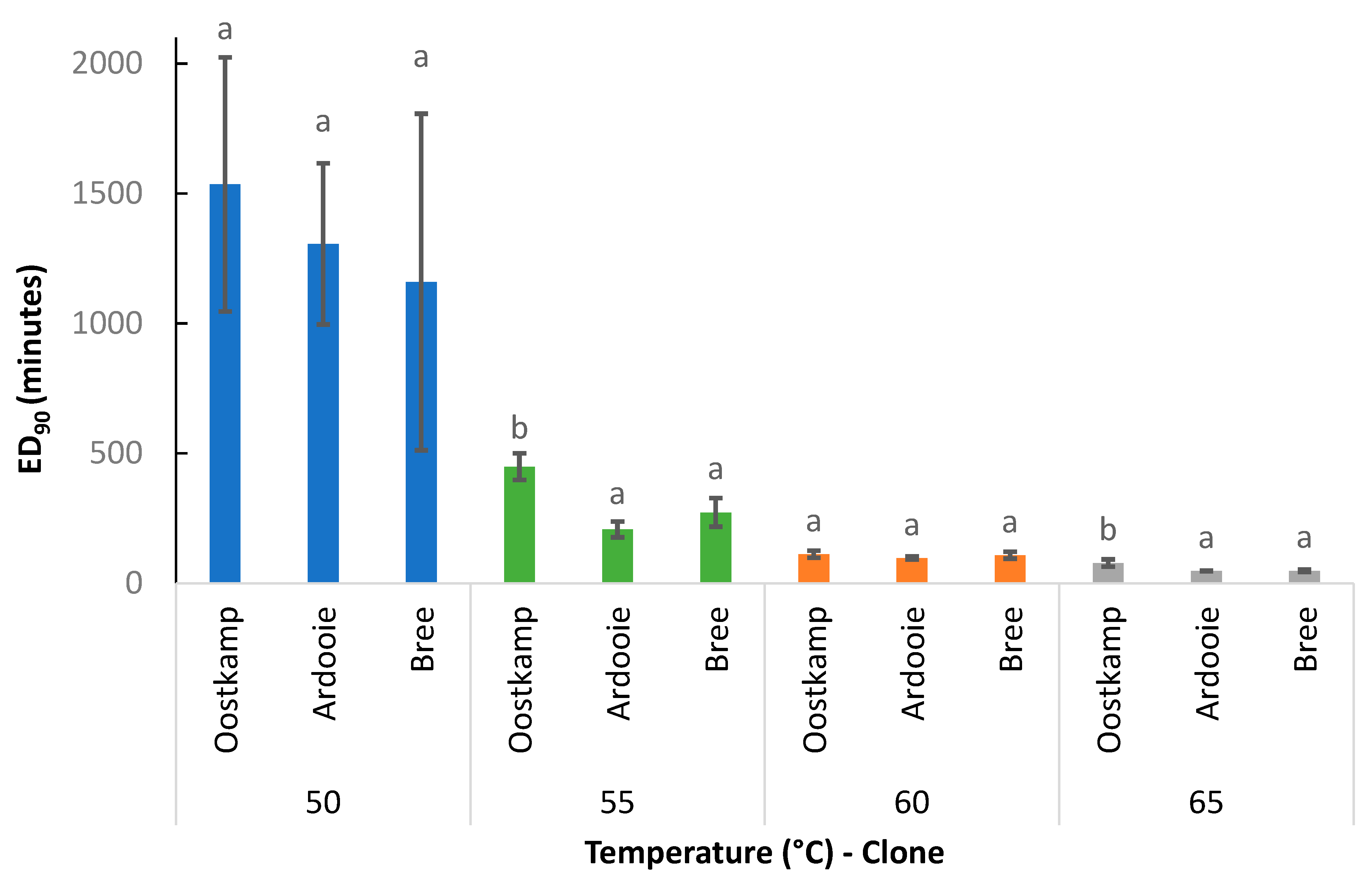
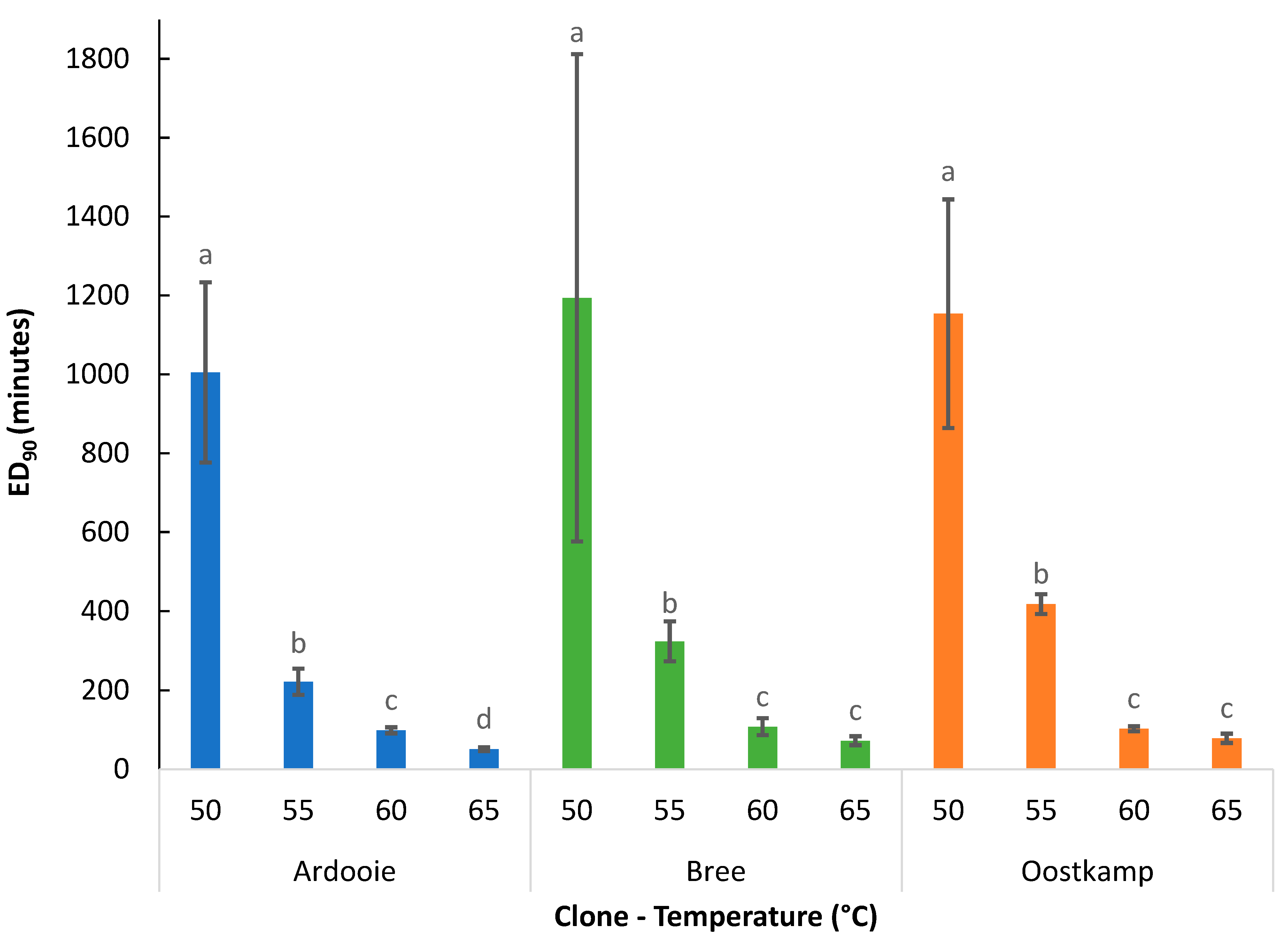
3.2.1. Effect of Temperature
In Figure 9, the effect of the applied temperature on the ED90 value is illustrated. For all investigated clones, the ED90 value was affected by exposure temperature. The ED90 value significantly decreased with increasing exposure temperature within the temperature range from 50 to 60 °C, irrespective of clone. A further decrease in ED90 value was observed when exposure temperature increased from 60 °C to 65 °C, but this decrease was only significant for clone Ardooie. Averaged over the clones, exposure durations of 1118, 321, 103, and 67 min at 50, 55, 60, and 65 °C, respectively, were required for a 90% reduction in tuber vitality.
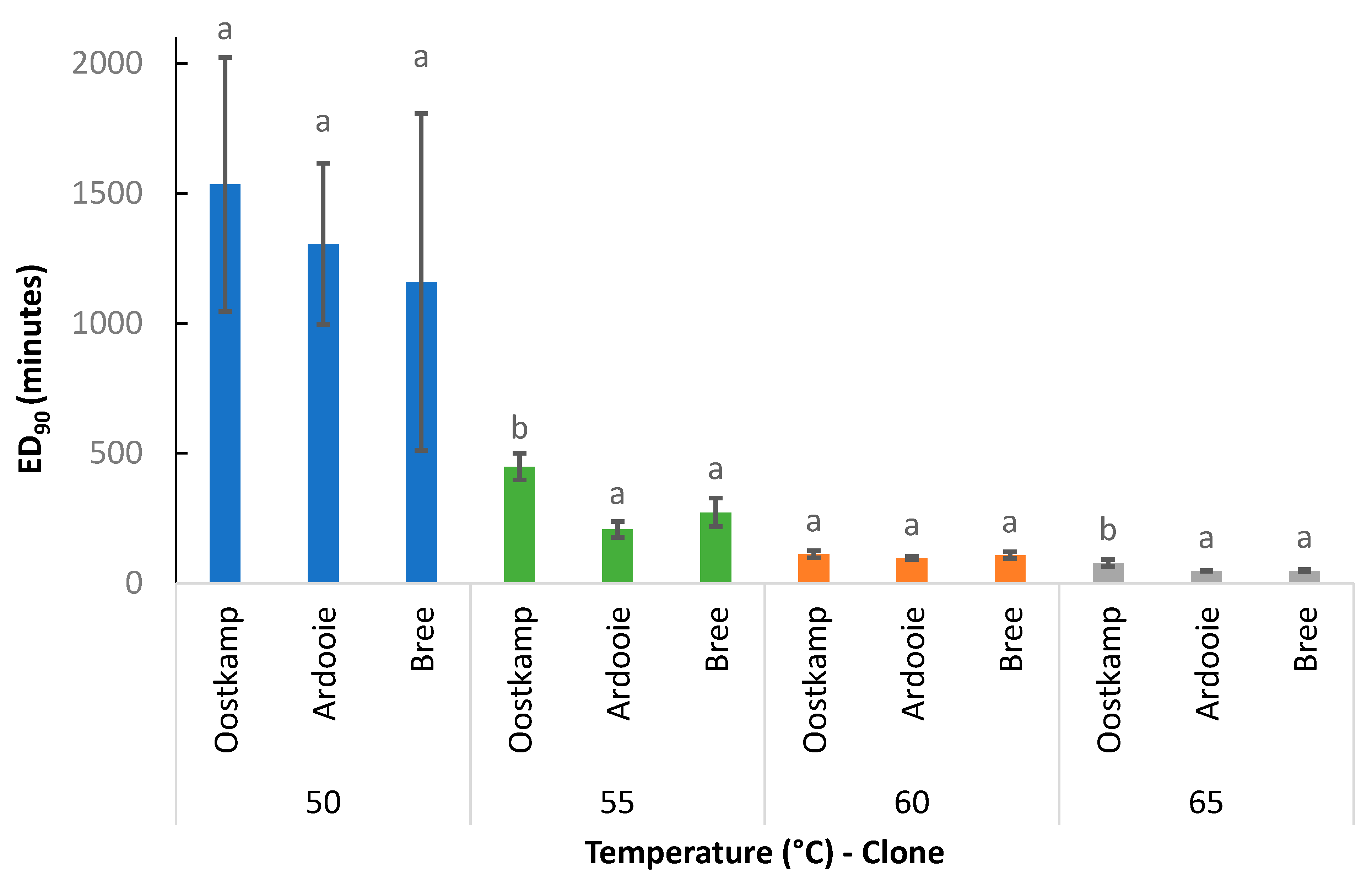
3.2.2. Effect of Genetic Clone
In Figure 10, the effect of the genetic C. esculentus clone on the ED90 value is illustrated. ED90 values were consistently higher for clone Oostkamp producing large tubers (mean fresh tuber weight of 500.0 mg) than for clones Ardooie and Bree, producing relatively small tubers (mean fresh tuber weights of 217.1 and 261.0 mg, respectively), irrespective of exposure temperature. However, the clone effect was only statistically significant at 55 and 65 °C. Averaged over the applied temperatures, the ED90 values for clones Oostkamp, Ardooie, and Bree were 543, 415, and 398 min, respectively.
4. Discussion
Hypothesis 1 stated that the efficiency of the hood steaming method depends on the genetic C. esculentus clone. This hypothesis is partly supported. In the field experiment, the genetic clone did affect the ED90 value to a limited extent. For example, at Sint-Niklaas in 2020, tubers of clone Bree (light tubers) were significantly more susceptible to hood steaming than tubers of clone Meulebeke (heavy tubers) (Figure 8). In the laboratory experiment, the effect of the genetic clone was confirmed at exposure temperatures of 55 and 65 °C, at which clones forming light tubers were most heat sensitive. In all cases with significant clone effects, the clone with the heaviest tubers was the most heat tolerant (clone Meulebeke in the field experiment, clone Oostkamp in the lab experiment). A number of explanations may account for these clone effects. Firstly, clones may differ in the degree to which axillary bud meristems, formed at the distal end of the tuber [36,37], are physically enclosed by protective scale-like sheaths (cladophylls). Axillary buds covered by numerous big phylloclades are better protected against heat damage. Secondly, clones may also differ in those chemical constituents that have a large impact on the thermal conductivity of outer tissue layers that protect bud meristems, in particular water and oils. Indeed, Matthiesen and Stoller [38] found that tuber lipid concentration differs among C. esculentus clones. Thirdly, clones differ in number of nodes per tuber. Large tubers have more nodes and hence, bear more axillary buds, thus increasing the resprouting ability of thermally treated tubers.
According to hypothesis 2, the efficiency of the hood steaming method is affected by tuber burial depth. This hypothesis is clearly supported. For example, in 2021 (the second experimental year), the ED90 value at 15 cm burial depth was significantly higher than the ED90 value at 5 cm depth within all combinations of clone and location (Figure 7). Averaged over clones and locations, the increase in ED90 value was 8.5 min. Furthermore, all ED90 values at 25 cm depth were statistically higher than the ED90 values at 15 cm depth. Averaged over clones and locations, the ED90 value was lengthened by 12.5 min. Also, in 2020 (the first experimental year), increasing ED90 values were observed with increasing burial depths. However, the differences in ED90 values at different burial depths were not always statistically significant in 2020. Nevertheless, our results underpin the assumption that hood steaming might be less energy-efficient for tubers at greater burial depths (>25 cm), as suggested by Keller et al. [22]. For example, in 2021 at Melle, soil temperature at 5 cm depth increased from 19.6 °C to 99.9 °C within 7.13 min of steaming. At 25 cm depth, soil temperature increased from 19.6 °C to 99.7 °C within 27.05 min. The deeper soil layers warm up more slowly due to the build-up of a condensation front, which strongly reduces steam penetration into the soil. In a condensation front, heat transfer predominantly occurs through conduction [18,39]. In naturally infested soils, tubers may occur at greater depths, up to a depth of 45 cm. Generally, up to 10% of the tubers are located below 25 cm [10,17]. Stoller and Sweet [10] observed in a field experiment that these deeply positioned tubers (up to 45 cm) can produce emerging shoots. This implies that very long steaming durations (probably between 30 and 60 min) are needed to control the whole tuber bank. Hence, alternative steaming techniques using direct steam injection into the soil might be an interesting option to improve efficacy and/or energy efficiency.
Hypothesis 3 stated that the efficiency of the hood steaming method is dependent on pedohydrological parameters, namely soil texture and soil moisture content. This hypothesis is only partly supported. In 2020, ED90 values for tubers buried at 5 cm depth tended to be lower (difference ranging from 1.63 to 1.89 min) at Sint-Niklaas than at Melle. However, this difference was statistically significant only for clone Meulebeke. At the depths of 15 and 25 cm, no differences in ED90 values were observed among locations. In the second experimental year (2021), no differences in ED90 values were observed among locations at any burial depth. So, in our experiments, the factor location affected the ED90 value only to a limited extent. The weak effect of location on steam efficacy is surprising, given the impact of soil texture and soil moisture content on heat capacity and thermal conductivity [40]. The specific heat capacity of a soil gives an indication of how much energy (Joules) is needed to heat 1 kg of that soil by 1 °C. The higher the specific heat capacity of a soil, the more slowly it heats, and the higher the ED90 value. The effect of soil moisture content on soil heat capacity is very clear. For example, the gravimetric heat capacities of dry and saturated sandy soils are approximately 800 and 1632 J kg−1 K−1, respectively [41]. At Sint-Niklaas, the upper soil layer (5 cm) heated up faster in 2020 than in 2021. After 2 min of steaming, a maximum temperature of 35.7 °C was observed in 2020, compared to 24.1 °C in 2021. This might be linked to the lower gravimetric soil moisture content in the upper soil layer in 2020 (2020: 8.56%, 2021: 10.14%).
Also, soil texture affects the soil heat capacity. Abu-Hamdeh [40] observed that clay soils generally have a higher gravimetric heat capacity than sandy soils at a particular soil moisture content and soil density. This is probably a result of the soil’s specific mineralogy and the differences in sand, silt, and clay fractions. Clay and silt have a heat capacity of approximately 1350 J kg−1 K−1, while sand has a heat capacity of approximately 1180 J kg−1 K−1 [42]. Quartz, the most common constituent of sand, has a relatively low specific heat capacity (740 J kg−1 K−1 at 20 °C) [42,43]. Based on soil texture and soil moisture content, soil heat capacity is expected to be higher at location Melle (heaviest soil and highest soil water content) than at location Sint-Niklaas. Hence, the soil at Melle is expected to heat up more slowly than the soil at Sint-Niklaas, especially near the soil surface. The time evolution of soil temperature obtained after soil steaming in 2020 confirms this expectation. For example, after 2 min of hood steaming, the upper soil layer was heated to 23.4 °C at Melle and 35.6 °C at Sint-Niklaas. So, the upper soil layer at Sint-Niklaas takes less energy to raise its temperature than the upper layer at Melle. As a result, the ED90 values tend to be lower at Sint-Niklaas. However, this tendency towards lower ED90 values in the upper soil layer was not observed in 2021. This might be explained by the varying soil moisture content between the experimental years. The difference in soil moisture content in the upper soil layer of both locations was 7.56 percentage points in 2020 and only 2.85 percentage points in 2021. As a result, the soil heat capacities of both topsoils were more comparable in 2021.
At the deeper burial depths (15 and 25 cm), the soil at Melle seemed to warm up faster than at Sint-Niklaas, as suggested by Figure 5. As described by Gay et al. [18], the condensation front is located at a depth between 5 and 16 cm in moist soils. In drier soils, the condensation front occurs at an even more superficial level. Beyond the condensation front, heat transfer is possible only via conduction. Thus, to declare differences in heating rate among locations, the thermal conductivity of the soils should be compared. This can be explained by the differential thermal conductivity of the soils at both locations. The parameter thermal conductivity (expressed as W m−1 K−1) describes the rate at which heat is transferred by conduction through a unit cross-section area of a certain material [44]. The thermal conductivity of soil is dependent on pedohydrological parameters. The finer the soil texture, the lower the thermal conductivity. This might be linked with soil mineralogy. Quartz and clay minerals have thermal conductivities of 8.8 and 3.0 W m−1 °C−1, respectively [43]. As a result, fine clay soils (high content of clay minerals) tend to have a lower thermal conductivity than coarse sandy soils (high content of quartz). However, according to Malek et al. [45], soil moisture content is the most important parameter influencing soil thermal conductivity. The higher the soil moisture content, the higher the thermal conductivity, which is linked with the relatively high thermal conductivity of water (0.598 W m−1 K−1) [46]. This might be an explanation for the faster heating of the deeper soil layers at Melle. Moreover, vertical steam penetration may also be better in the soil at Melle as a result of the high number of air-filled macrospores in a soil largely consisting of large soil aggregates. On the other hand, the soil at Melle is characterized by a higher heat capacity, which might lead to a slower heating relative to the soil at Sint-Niklaas. Apparently, as the soil was heating faster at Melle, this effect was limited. The faster heating at Melle compared to Sint-Niklaas corresponds to the general assumption that sandy soils take more time to heat than sandy loam soils due to their dense structure [47,48]. This was also observed by Melander and Kristensen [21,49], who steamed a sandy and sandy, loamy soil in a laboratory rig.
Despite the faster heating of the deeper soil layers at Melle, no statistical differences in ED90 values were observed among locations. This can be explained by the shape of the dose–response curves. Within a short amount of time, tuber vitalities decline from approximately 100% to 0% at both locations. For example, tubers buried at 15 cm depth had a vitality of approximately 100% after a steaming duration of 8 min. After a steaming duration of 16 min, tuber vitality has already declined to 0%. As a result, standard errors of calculated ED90 values for tubers buried at 25 cm were too high to detect significant differences in ED90 values among locations. Another reason for the lack of a location effect might be the differential soil aggregate size distribution of both locations. Coarse aggregates (soil at Melle) may facilitate fast heat penetration through their macropore spaces but may slow down the heat transport within aggregates, thus protecting tubers encapsulated in these clods.
As a rule of thumb, steaming durations of 8, 16, and 32 min are needed to obtain 100% control of tubers at depths of 5, 15, and 25 cm, respectively. In this process, the recorded temperatures were not necessarily very high (>65 °C). For example, a steaming duration of 8 min led to a 100% control of superficial tubers at Melle in 2021. During and until one hour after this treatment, the temperature exceeded 50 °C for 2185 s, while the threshold of 65 °C was not exceeded (Table 3). At Sint-Niklaas in 2021, a steaming duration of 16 min gave 100% control of tubers at 15 cm depth. During and until one hour after the treatment, even the threshold of 50 °C was not exceeded. However, it should be stressed that the temperature exceeded 47.5 °C for 1483 s. Moreover, there was no tendency towards a declining temperature at the end of the temperature registration (Figure 4). In 2020 (at Sint-Niklaas), the temperature at 15 cm depth exceeded 50 °C for 1915 s (Table 3), which gave 100% control of tubers. At the end of the temperature monitoring, the temperature was still higher than 50 °C (Figure 4). The threshold of 50 °C was probably still exceeded for a few additional minutes. Based on previous data, it could be stated that a 100% tuber control can be achieved when the temperature exceeds 50 °C for at least 2500 s, corresponding to approximately 42 min. In the lab experiment, much higher durations were required for a complete kill. For example, at 50 °C, the ED90 value was 1118 min averaged over all clones. This more or less corresponds with the findings of Webster [14], who observed that C. esculentus tubers are killed at 50 °C (hot air) with an exposure duration of 16 h (=960 min). The higher exposure durations for 90% control in the lab experiment compared to the field experiment are not surprising. In contrast with the field experiment, where heat is conductively transported to the tubers via the solid, liquid, and gaseous phases, heat transport in the lab experiments only occurs via the gaseous phase. As thermal conductivity is 24 times higher in water than in air (0.598 W m−1 K−1 versus 0.026 W m−1 K−1) [46,50], the heat transfer was much faster in the field experiment.
5. Conclusions
Hood steaming might be an interesting alternative technique to control C. esculentus tubers in small infested patches (a few square meters), especially in zones in which herbicides are not allowed or in organic agriculture. Larger infested patches might be less interesting due to the high energy, time, and water consumption during the process. In our two-year experiments, C. esculentus clones producing small tubers seemed to be more sensitive to hood steaming. The higher heat sensitivity of these clones was clearly confirmed in the laboratory experiment. Soil moisture content affected steam efficiency to a small extent. The higher the soil moisture content, the better the heat conduction in the deeper soil layers. However, differences in soil moisture content did not lead to significantly different ED90 values among locations. The effect of tuber burial depth on steaming efficiency was very clear. As a rule of thumb, 8, 16, and 32 min of hood steaming are required to achieve 100% tuber control at depths of 5, 15, and 25 cm, respectively. The temperature should exceed 50 °C for at least 2500 s to obtain 100% tuber control. To improve steam penetration into the soil, the soil should be tilled before the treatment. Ideally, tillage depth should correspond with the maximum burial depth of the C. esculentus tubers. After the treatment, the treated area should be visually inspected for emerging C. esculentus shoots. Future research should focus on reducing the required energy, time, and water consumption during the process, for example, by using perforated spikes that inject the steam directly into the soil mass. Moreover, the dependency on fossil-based energy sources should be phased out, e.g., by implementing photovoltaic energy systems or other renewable energy technologies. In the future, the importance of the steaming technique might increase as more and more herbicides lose their authorization or herbicide-resistant C. esculentus clones may spread rapidly.
Author Contributions
J.F.: conceptualization, methodology, analysis, writing—original draft, writing—reviewing and editing, and resources. S.D.R.: conceptualization, methodology, analysis, and resources. C.S.: methodology, analysis, and resources. D.R.: writing—reviewing and editing. J.L.: conceptualization, methodology, analysis, and resources. D.C.: conceptualization, methodology, analysis, and resources. B.D.C.: conceptualization, methodology, analysis, writing—reviewing and editing, resources, and supervision. All authors have read and agreed to the published version of the manuscript.
Funding
Both this research and the APC were funded by Flanders Innovation and Entrepreneurship (VLAIO), grant number AIO.LAN.2021.0003.01.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author. The data are not publicly available due to privacy reasons.
Acknowledgments
This study was financially supported by Flanders Innovation and Entrepreneurship (VLAIO). Special thanks to Simox Belgium for providing the steaming equipment. Furthermore, special thanks to the technical staff of Ghent University and the University College of Ghent (HOGENT) for the practical aid during the experiments.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. The World’s Worst Weeds; Kriegar Publishing: Malabar, FL, USA, 1991; ISBN 0894644157. [Google Scholar]
- Bohren, C.; Wirth, J. Souchet comestible (Cyperus esculentus L.): Situation actuelle en Suisse. Rech. Agron. Suisse 2016, 4, 460–467. [Google Scholar]
- Total, R.; Collet, L.; Heyer, J.; Keller, M. Yield losses in vegetable and arable crops caused by yellow nutsedge (Cyperus esculentus) in farmers fields in Switzerland. Julius-Kühn-Archiv 2018, 458, 473–477. [Google Scholar]
- Stoller, E.W.; Wax, L.M.; Slife, F.W. Yellow nutsedge (Cyperus esculentus) competition and control in corn (Zea mays). Weed Sci. 1979, 27, 32–37. [Google Scholar] [CrossRef]
- Pascual, B.; Maroto, J.V.; López-Galarza, S.; Sanbautista, A.; Alagarda, J. Chufa (Cyperus esculentus L. var. sativus Boeck.): An unconventional crop. Studies related to applications and cultivation. Econ. Bot. 2000, 54, 439–448. [Google Scholar] [CrossRef]
- De Cauwer, B.; De Ryck, S.; Claerhout, S.; Biesemans, N.; Reheul, D. Differences in growth and herbicide sensitivity among Cyperus esculentus clones found in Belgian maize fields. Weed Res. 2017, 57, 234–246. [Google Scholar] [CrossRef]
- Vonnez, J. Rapport Final Relatif au Projet OFAG: Stratégies de Lutte Contre le Souchet Comestible. Available online: https://www.souchet-comestible.ch/fileadmin/PDF/rapport_final_souchet/Rapport_final_projet_souchet_comestible.pdf (accessed on 13 July 2023).
- Demeyere, A. Praktijkgids Gewasbescherming. Module IPM Akkerbouw. Departement Landbouw En Visserij. Available online: https://www.vlaanderen.be/publicaties/praktijkgids-gewasbescherming-module-ipm-akkerbouw (accessed on 13 July 2023).
- Lapham, J.; Drennan, D.S.H. The fate of yellow nutsedge (Cyperus esculentus) seed and seedlings in soil. Weed Sci. 1990, 38, 125–128. [Google Scholar] [CrossRef]
- Stoller, E.W.; Sweet, R.D. Biology and life cycle of purple and yellow nutsedges (Cyperus rotundus and C. esculentus). Weed Technol. 1987, 1, 66–73. [Google Scholar] [CrossRef]
- Keller, M.; Eppler, L.; Collet, L.; Wirth, J.; Total, R. Beim Erdmandelgras auf nummer sicher gehen: Auch blütenbildung und abblühen verhindern! Gemüsebau Info 2015, 22, 7–9. [Google Scholar]
- Follak, S.; Belz, R.; Bohren, C.; De Castro, O.; Del Guacchio, E.; Pascual-Seva, N.; Schwarz, M.; Verloove, F.; Essl, F. Biological flora of Central Europe: Cyperus esculentus L. Perspect. Plant Ecol. Evol. Syst. 2016, 23, 33–51. [Google Scholar] [CrossRef]
- De Ryck, S.; Reheul, D.; De Cauwer, B. Impacts of herbicide sequences and vertical tuber distribution on the chemical control of yellow nutsedge (Cyperus esculentus L.). Weed Res. 2021, 61, 454–464. [Google Scholar] [CrossRef]
- Webster, T.M. High temperatures and durations of exposure reduce nutsedge (Cyperus spp.) tuber viability. Weed Sci. 2003, 51, 1010–1015. [Google Scholar] [CrossRef]
- Feys, J.; De Cauwer, B.; Reheul, D.; Sciffer, C.; Clercx, S.; Palmans, S. Impact of electrocution on shoot and tuber vitality of yellow nutsedge (Cyperus esculentus). Agriculture 2023, 13, 696. [Google Scholar] [CrossRef]
- El-Keblawy, A.; Al-Hammadi, F. Soil amendments enhance soil solarisation efficiency in controlling weeds under the environment of the United Arab Emerites. In Proceedings of the 16th Australian Weeds Conference, Cairns Convention Centre, North Queensland, Australia, 18–22 May 2008. [Google Scholar]
- Tumbleson, M.E.; Kommedahl, T. Reproductive potential of Cyperus esculentus by tubers. Weeds 1961, 9, 646–653. [Google Scholar] [CrossRef]
- Gay, P.; Piccarolo, P.; Aimonino, D.R.; Tortia, C. A high efficiency steam soil disinfestation system, part I: Physical background and steam supply optimisation. Biosyst. Eng. 2010, 107, 74–85. [Google Scholar] [CrossRef]
- Gay, P.; Piccarolo, P.; Aimonino, D.R.; Tortia, C. Soil parameters effects on steam disinfestation efficiency. In Agricultural and biosystems engineering for a sustainable world. In Proceedings of the International Conference on Agricultural Engineering, Hersonissos, Crete, Greece, 23–25 June 2008. [Google Scholar]
- Sauer, T.J.; Horton, R. Soil heat flux. In Micrometeorology in Agricultural Systems; Hatfield, J.L., Baker, J.M., Viney, M.K., Eds.; American Society of Agronomy: Madison, WI, USA, 2005; ISBN 089118158X. [Google Scholar]
- Melander, B.; Kristensen, J.K. Soil steaming effects on weed seedling emergence under the influence of soil type, soil moisture, soil structure and heat duration. Ann. Appl. Biol. 2011, 158, 194–203. [Google Scholar] [CrossRef]
- Keller, M.; Collet, L.; Total, R. Using steam to eradicate Cyperus esculentus infestations in vegetable fields in Switzerland. In Proceedings of the Joint EWRS Workshop of the Working Groups Physical and Cultural Weed Control and Crop-Weed Interactions, Nyon, Switzerland, 2–5 April 2017. [Google Scholar]
- Total, R.; Collet, L.; Keller, M. Souchet comestible: Élimination des foyers primaires par traitement à la vapeur. Agrocope Transf. 2016, 137, 1–6. [Google Scholar]
- Samtani, J.B.; Gilbert, C.; Weber, J.B.; Subbarao, K.V.; Goodhue, R.E.; Fennimore, S.A. Effect of steam and solarization treatments on pest control, strawberry yield, and economic returns relative to methyl bromide fumigation. HortScience 2012, 47, 64–70. [Google Scholar] [CrossRef]
- Pinel, M.P.C.; Bond, W.; de Coursey Williams, M.; Johnston, E.; White, J.G. Field Vegetables: Assessment of the Potential for Mobile Soil Steaming Machinery to Control Diseases, Weeds and Mites of Field Salad and Related Crops; Final Report, Horticultural Development Council Project; Horticultural Development Council: East Malling, UK, 1999. [Google Scholar]
- Roux-Michollet, D.; Dudal, Y.; Jocteur-Monrozier, L.; Czarnes, S. Steam treatment of surface soil: How does it affect water-soluble organic matter, C mineralization, and bacterial community composition? Biol. Fert. Soils 2010, 46, 607–616. [Google Scholar] [CrossRef]
- De Ryck, S.; Reheul, D.; De Riek, J.; De Keyser, E.; De Cauwer, B. Genetic and morphological variation of Belgian Cyperus esculentus L. clonal populations and their significance for integrated management. Agronomy 2023, 13, 572. [Google Scholar] [CrossRef]
- Simox. Polyvap—Onkruid Verdelgen Met Respect Voor Milieu en Gezondheid. Available online: https://www.bluesphere.be/images/sites/51/article/117/1/Folder_Stoomgeneratoren.pdf (accessed on 14 August 2023).
- Feredec Bretagne. Guide des Alternatives au Désherbage Chimique dans les Communes. Available online: https://www.sagebaiededouarnenez.org/telechargement/Phytos_non_agricoles/Guide-des-alternatives-au-d%C3%A9sherbage-chimique-dans-les-communes.pdf (accessed on 14 August 2023).
- Jaichandar, S.; Annamalai, K. The status of biodiesel as an alternative fuel for diesel engine—An overview. J. Sustain. Energy Environ. 2011, 2, 71–75. [Google Scholar]
- Artisan Technology Group. General Purpose Incubators. Available online: https://www.artisantg.com/info/VWR_1500_Series_Datasheet.pdf (accessed on 11 December 2023).
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 17 August 2023).
- Ritz, C.; Streibig, J.C. Bioassay analysis using R. J. Stat. Softw. 2005, 12, 1–22. [Google Scholar] [CrossRef]
- Streibig, J.C.; Rudemo, M.; Jensen, J.E. Dose-response curves and statistical models. In Herbicide Bioassays; Streibig, J.C., Kudsk, P., Eds.; CRC Press: Boca Raton, FL, USA, 1993; ISBN 0849366038. [Google Scholar]
- Van Der Vaart, A.W. Asymptotic Statistics; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Thullen, R.J.; Keeley, P.E. Yellow nutsedge sprouting and resprouting potential. Weed Sci. 1975, 23, 333–337. [Google Scholar] [CrossRef]
- Kehler, R. Investigations on Yellow Nutsedge. Master’s Thesis, University of Manitoba, Winnipeg, Canada, 1991. [Google Scholar]
- Matthiesen, R.L.; Stoller, E.W. Tuber composition in yellow nutsedge (Cyperus esculentus (L.)) variants. Weed Res. 1978, 18, 373–377. [Google Scholar] [CrossRef]
- Dabbene, F.; Gay, P.; Tortia, C. Modeling and control of steam soil disinfestation processes. Biosyst. Eng. 2003, 84, 247–256. [Google Scholar] [CrossRef]
- Abu-Hamdeh, N.H. Thermal properties of soils as affected by density and water content. Biosyst. Eng. 2003, 86, 97–102. [Google Scholar] [CrossRef]
- Hamdhan, I.N.; Clarke, B.G. Determination of thermal conductivity of coarse and fine sand soils. In Proceedings of the World Geothermal Congress, Bali, Indonesia, 25–29 April 2010. [Google Scholar]
- Zhu, X.; Gao, Z.; Chen, T.; Wang, W.; Lu, C.; Zhang, Q. Study on the thermophysical properties and influencing factors of regional surface shallow rock and soil in China. Front. Earth Sci. 2022, 10, 864548. [Google Scholar] [CrossRef]
- Waples, D.W.; Waples, J.S. A review and evaluation of specific heat capacities of rocks, minerals, and subsurface fluids. Part 1: Minerals and nonporous rocks. Nat. Resour. Res. 2004, 13, 97–122. [Google Scholar] [CrossRef]
- Ratna, D. Thermal properties of thermosets. In Thermosets: Structure, Properties and Applications; Guo, Q., Ed.; Woodhead Publishing: Sawston, UK, 2012; ISBN 978-0-85709-086-7. [Google Scholar]
- Malek, K.; Malek, K.; Khanmohammadi, F. Response of soil thermal conductivity to various soil properties. Int. Commun. Heat Mass 2021, 127, 105516. [Google Scholar] [CrossRef]
- Huber, M.L.; Perkins, R.A.; Friend, D.G.; Sengers, J.V.; Assael, M.J.; Metaxa, I.N.; Miyagawa, K.; Hellmann, R.; Vogel, E. New international formulation for the thermal conductivity of H2O. J. Phys. Chem. Ref. Data 2012, 41, 033102. [Google Scholar] [CrossRef]
- Runia, W.T. A recent development in steam sterilisation. Acta Hortic. 1983, 152, 195–200. [Google Scholar] [CrossRef]
- Minuto, G.; Gilardi, G.; Kejji, S.; Gullino, M.L.; Garibaldi, A. Effect of physical nature of soil and humidity on stream disinfestation. Acta Hortic. 2005, 698, 257–262. [Google Scholar] [CrossRef]
- Melander, B.; Jørgensen, M.H. Soil steaming to reduce intra-row weed seedling emergence. Weed Res. 2005, 45, 202–211. [Google Scholar] [CrossRef]
- The Engineering Toolbox. Air–Thermal Conductivity vs. Temperature and Pressure. Available online: https://www.engineeringtoolbox.com/air-properties-viscosity-conductivity-heat-capacity-d_1509.html (accessed on 15 April 2024).
Figure 1.
The different soil zones and corresponding temperatures during the hood steaming process. Figure retrieved from Gay et al. [18].
Figure 1.
The different soil zones and corresponding temperatures during the hood steaming process. Figure retrieved from Gay et al. [18].

Figure 3.
(Left): The gullies with depths of 5 cm (left), 15 cm (middle), and 25 cm (right). (Right): detail of the nylon bags which were laid in the gullies.
Figure 3.
(Left): The gullies with depths of 5 cm (left), 15 cm (middle), and 25 cm (right). (Right): detail of the nylon bags which were laid in the gullies.

Figure 4.
The average temperature (°C) and average relative humidity (%) during the screening period in the first (above) and second (below) experimental years. (*): In the first experimental year, the screening period started on 13 May. In the second experimental year, the screening period started on 28 April.
Figure 4.
The average temperature (°C) and average relative humidity (%) during the screening period in the first (above) and second (below) experimental years. (*): In the first experimental year, the screening period started on 13 May. In the second experimental year, the screening period started on 28 April.

Figure 5.
The measured temperatures (°C) at a particular burial depth during and until one hour after steaming for a particular duration. The end of a steaming duration is indicated with a red arrow. The treatments took place in 2020 and 2021 at Melle and Sint-Niklaas.
Figure 5.
The measured temperatures (°C) at a particular burial depth during and until one hour after steaming for a particular duration. The end of a steaming duration is indicated with a red arrow. The treatments took place in 2020 and 2021 at Melle and Sint-Niklaas.
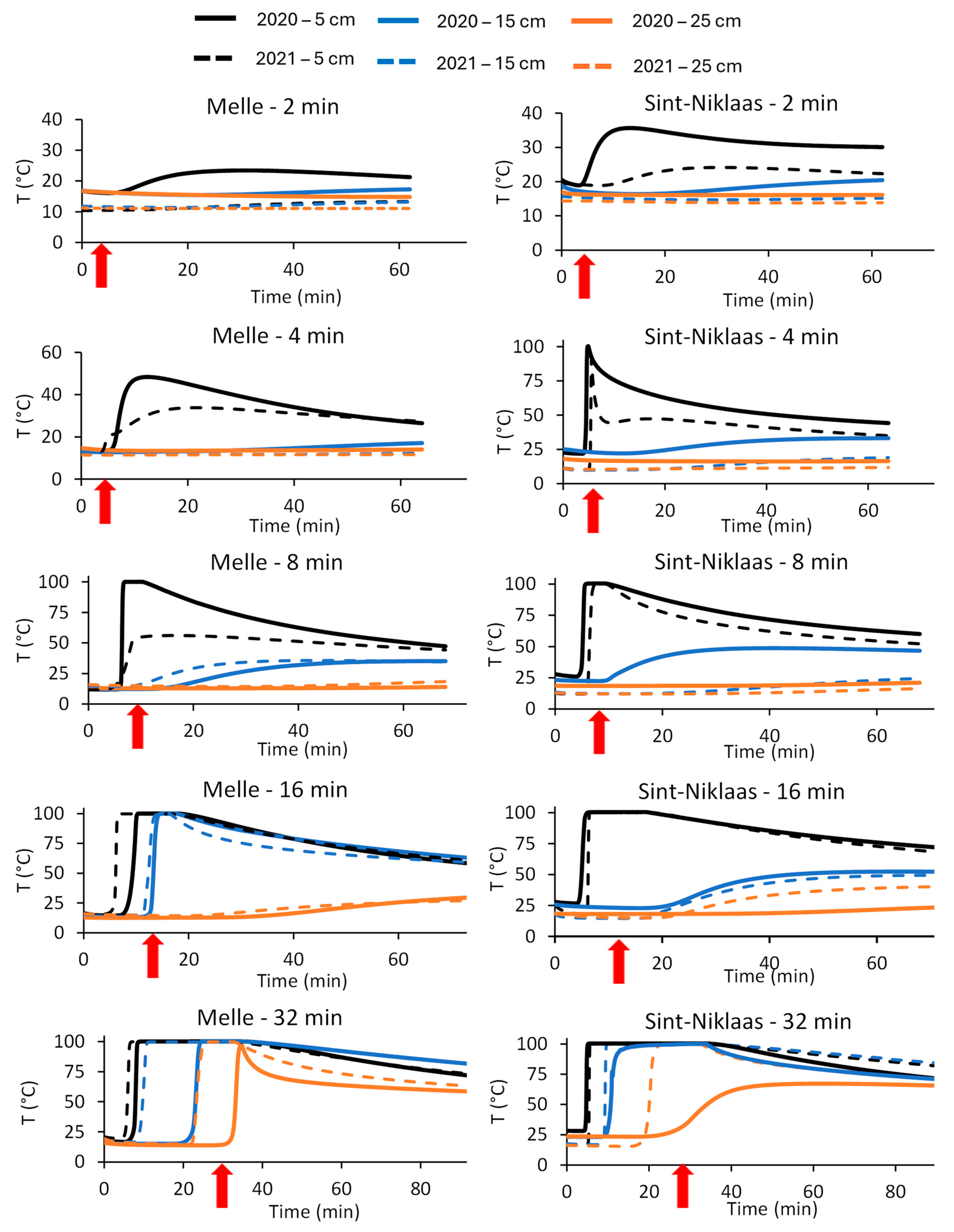
Figure 6.
The estimated ED90 values (mean ± standard error) for each combination of clone, burial depth, and location. Means without a common letter are significantly different (p < 0.05). Comparison within a particular combination of clone and burial depth only. (Top): first experimental year (2020). (Bottom): second experimental year (2021).
Figure 6.
The estimated ED90 values (mean ± standard error) for each combination of clone, burial depth, and location. Means without a common letter are significantly different (p < 0.05). Comparison within a particular combination of clone and burial depth only. (Top): first experimental year (2020). (Bottom): second experimental year (2021).

Figure 7.
The estimated ED90 values (mean ± standard error) for each combination of clone, location, and burial depth (cm). Means without a common letter are significantly different (p < 0.05). Comparison within a particular combination of clone and location only. (Top): first experimental year (2020). (Bottom): second experimental year (2021).
Figure 7.
The estimated ED90 values (mean ± standard error) for each combination of clone, location, and burial depth (cm). Means without a common letter are significantly different (p < 0.05). Comparison within a particular combination of clone and location only. (Top): first experimental year (2020). (Bottom): second experimental year (2021).

Figure 8.
The estimated ED90 values (mean ± standard error) for each combination of burial depth, location, and clone. Means without a common letter are significantly different (p < 0.05). Comparison within a particular combination of burial depth and location only. (Top): first experimental year (2020). (Bottom): second experimental year (2021).
Figure 8.
The estimated ED90 values (mean ± standard error) for each combination of burial depth, location, and clone. Means without a common letter are significantly different (p < 0.05). Comparison within a particular combination of burial depth and location only. (Top): first experimental year (2020). (Bottom): second experimental year (2021).
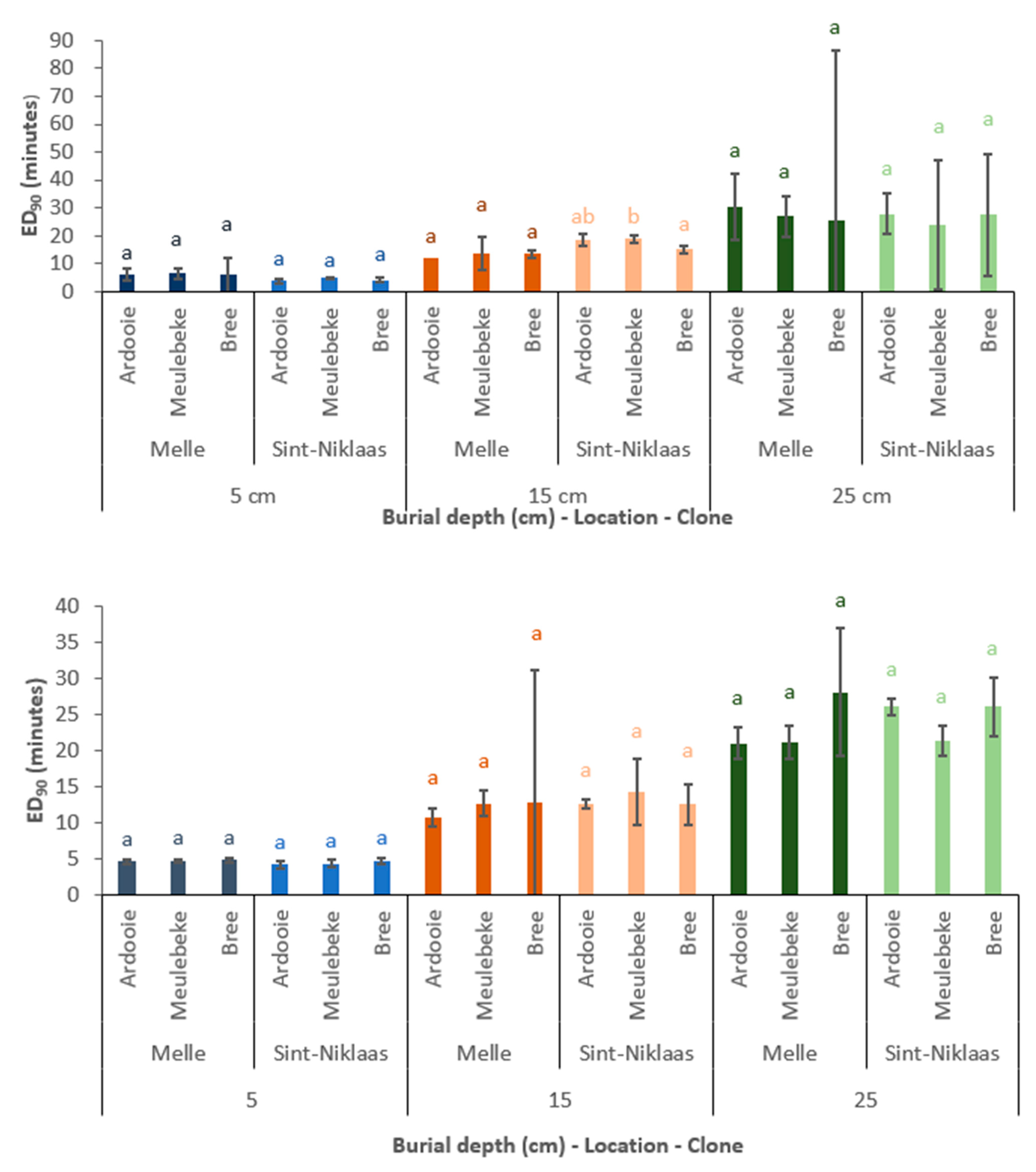
Figure 9.
The ED90 values (mean ± standard error) for all combinations of genetic clones and temperatures (°C). Means without a common letter are statistically different. Comparison within clone only.
Figure 9.
The ED90 values (mean ± standard error) for all combinations of genetic clones and temperatures (°C). Means without a common letter are statistically different. Comparison within clone only.

Figure 10.
The ED90 values (mean ± standard error) for all combinations of temperature (°C) and genetic clone. Means without a common letter are statistically different. Comparison within temperature only.
Figure 10.
The ED90 values (mean ± standard error) for all combinations of temperature (°C) and genetic clone. Means without a common letter are statistically different. Comparison within temperature only.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Gravimetric soil moisture content (%) 1 h prior to hood steaming for all combinations of soil depth, experimental site, and year.
Table 1.
Gravimetric soil moisture content (%) 1 h prior to hood steaming for all combinations of soil depth, experimental site, and year.
| Location | Year | Day | Depth (cm) | Gravimetric Soil Moisture Content (%) |
|---|---|---|---|---|
| Melle | 2020 | 13 May | 5 | 16.12 |
| 15 | 14.95 | |||
| 25 | 18.58 | |||
| 2021 | 28 April | 5 | 12.99 | |
| 15 | 15.35 | |||
| 25 | 17.64 | |||
| Sint-Niklaas | 2020 | 19 May | 5 | 8.56 |
| 15 | 11.12 | |||
| 25 | 12.57 | |||
| 2021 | 12 May | 5 | 10.14 | |
| 15 | 10.65 | |||
| 25 | 11.27 |
Table 2.
Specific exposure durations for each investigated temperature in the laboratory experiment.
Table 2.
Specific exposure durations for each investigated temperature in the laboratory experiment.
| Temperature (°C) | ||||
|---|---|---|---|---|
| 50 | 55 | 60 | 65 | |
| Exposure durations (min) | 0 | 0 | 0 | 0 |
| 25 | 13 | 7 | 7 | |
| 48 | 24 | 12 | 12 | |
| 96 | 48 | 24 | 24 | |
| 192 | 96 | 48 | 48 | |
| 384 | 192 | 96 | 96 | |
| 768 | 384 | 192 | 192 | |
Table 3.
Residence time above a particular temperature threshold, at a particular burial depth, steaming duration, location (Melle or Sint-Niklaas), and experimental year (2020 or 2021).
Table 3.
Residence time above a particular temperature threshold, at a particular burial depth, steaming duration, location (Melle or Sint-Niklaas), and experimental year (2020 or 2021).
| Location | Steaming Duration (min) | Temperature Threshold (°C) | Residence Time (s) above an Indicated Temperature Threshold at a Particular Burial Depth | |||||
|---|---|---|---|---|---|---|---|---|
| 2020 | 2021 | |||||||
| 5 cm | 15 cm | 25 cm | 5 cm | 15 cm | 25 cm | |||
| Melle | 2 | 50 | 0 | 0 | 0 | 0 | 0 | 0 |
| 65 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 80 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 95 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 4 | 50 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 65 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 80 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 95 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 8 | 50 | 3310 | 0 | 0 | 2185 | 0 | 0 | |
| 65 | 1831 | 0 | 0 | 0 | 0 | 0 | ||
| 80 | 984 | 0 | 0 | 0 | 0 | 0 | ||
| 95 | 409 | 0 | 0 | 0 | 0 | 0 | ||
| 16 | 50 | 4001 | 3764 | 0 | 4196 | 3820 | 0 | |
| 65 | 3005 | 3266 | 0 | 3432 | 2313 | 0 | ||
| 80 | 1737 | 1496 | 0 | 1918 | 752 | 0 | ||
| 95 | 855 | 451 | 0 | 954 | 281 | 0 | ||
| 32 | 50 | 5047 | 4136 | 3528 | 5170 | 4939 | 4121 | |
| 65 | 5040 | 4121 | 1314 | 5160 | 4921 | 3627 | ||
| 80 | 3895 | 4106 | 243 | 4083 | 4904 | 1493 | ||
| 95 | 2331 | 1692 | 47 | 2220 | 2511 | 670 | ||
| Sint-Niklaas | 2 | 50 | 0 | 0 | 0 | 0 | 0 | 0 |
| 65 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 80 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 95 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 4 | 50 | 2260 | 0 | 0 | 102 | 0 | 0 | |
| 65 | 780 | 0 | 0 | 40 | 0 | 0 | ||
| 80 | 213 | 0 | 0 | 14 | 0 | 0 | ||
| 95 | 38 | 0 | 0 | 0 | 0 | 0 | ||
| 8 | 50 | 3773 | 0 | 0 | 3696 | 0 | 0 | |
| 65 | 2898 | 0 | 0 | 1695 | 0 | 0 | ||
| 80 | 1369 | 0 | 0 | 700 | 0 | 0 | ||
| 95 | 523 | 0 | 0 | 267 | 0 | 0 | ||
| 16 | 50 | 4261 | 1915 | 0 | 4188 | 0 | 0 | |
| 65 | 4252 | 0 | 0 | 4186 | 0 | 0 | ||
| 80 | 2711 | 0 | 0 | 2386 | 0 | 0 | ||
| 95 | 1169 | 0 | 0 | 1143 | 0 | 0 | ||
| 32 | 50 | 5238 | 4880 | 3478 | 5196 | 4963 | 4316 | |
| 65 | 5235 | 4867 | 2803 | 5194 | 4960 | 4299 | ||
| 80 | 3879 | 2869 | 0 | 5192 | 4956 | 2234 | ||
| 95 | 2381 | 1418 | 0 | 2698 | 2699 | 840 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Feys, J.; De Ryck, S.; Sciffer, C.; Reheul, D.; Latré, J.; Callens, D.; De Cauwer, B. Impact of Hood Steaming on Tuber Vitality of Yellow Nutsedge (Cyperus esculentus). Agronomy 2024, 14, 918. https://doi.org/10.3390/agronomy14050918
AMA Style
Feys J, De Ryck S, Sciffer C, Reheul D, Latré J, Callens D, De Cauwer B. Impact of Hood Steaming on Tuber Vitality of Yellow Nutsedge (Cyperus esculentus). Agronomy. 2024; 14(5):918. https://doi.org/10.3390/agronomy14050918
Chicago/Turabian StyleFeys, Jeroen, Sander De Ryck, Clara Sciffer, Dirk Reheul, Joos Latré, Danny Callens, and Benny De Cauwer. 2024. "Impact of Hood Steaming on Tuber Vitality of Yellow Nutsedge (Cyperus esculentus)" Agronomy 14, no. 5: 918. https://doi.org/10.3390/agronomy14050918
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.