


Effect of Drip Irrigation, N, K, and Zn Coupling on
Abstract
:1. Introduction
2. Materials and Methods
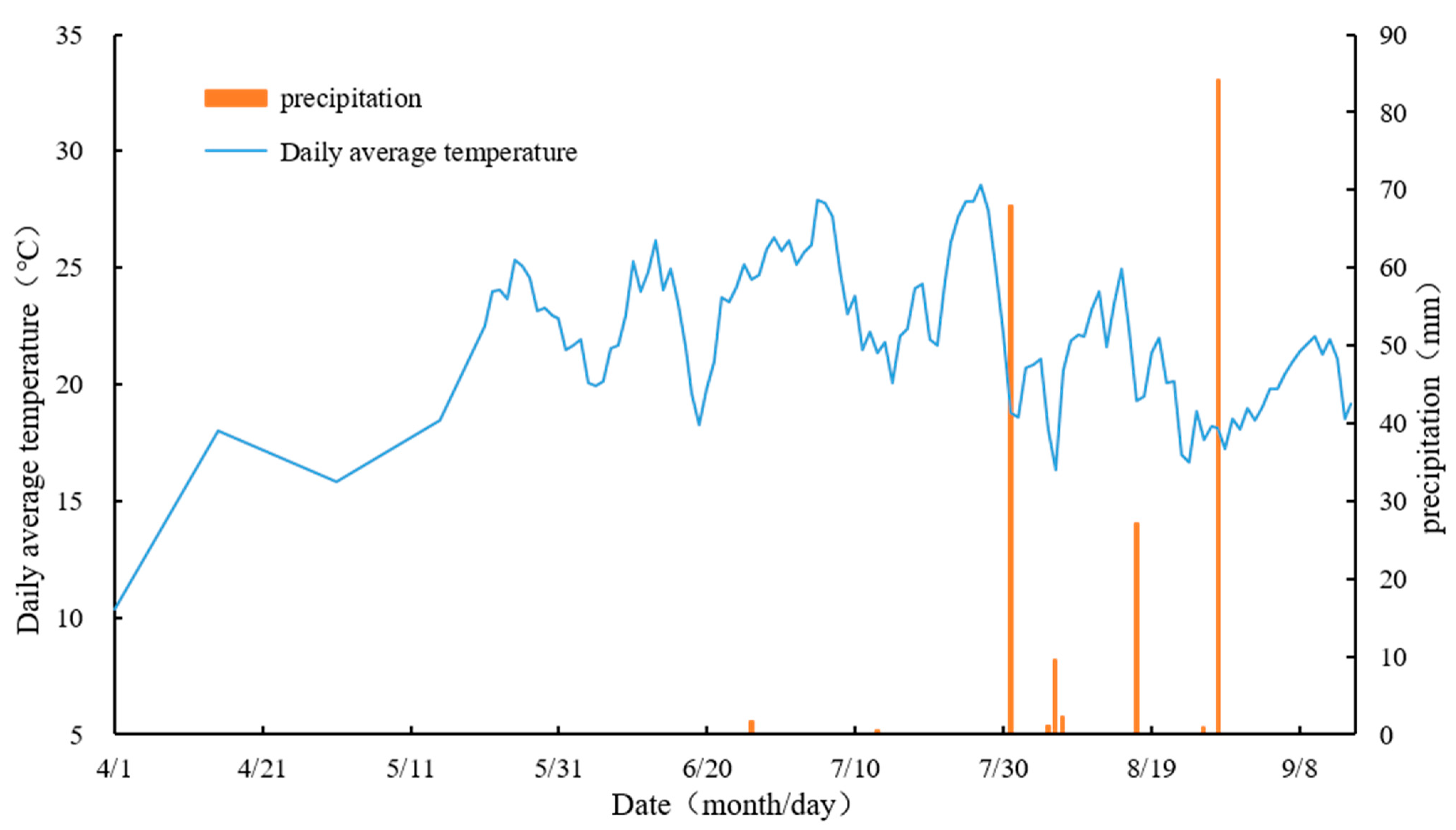
2.1. Overview of the Study Area
2.2. Experimental Materials
2.3. Experimental Design
2.4. Data Acquisition and Analysis
- (1)
- Sample collection
- (2)
- Construction of regression models
- (3)
- Data processing and mapping
3. Results and Analysis
3.1. Analysis of Pn in Apple Leaves
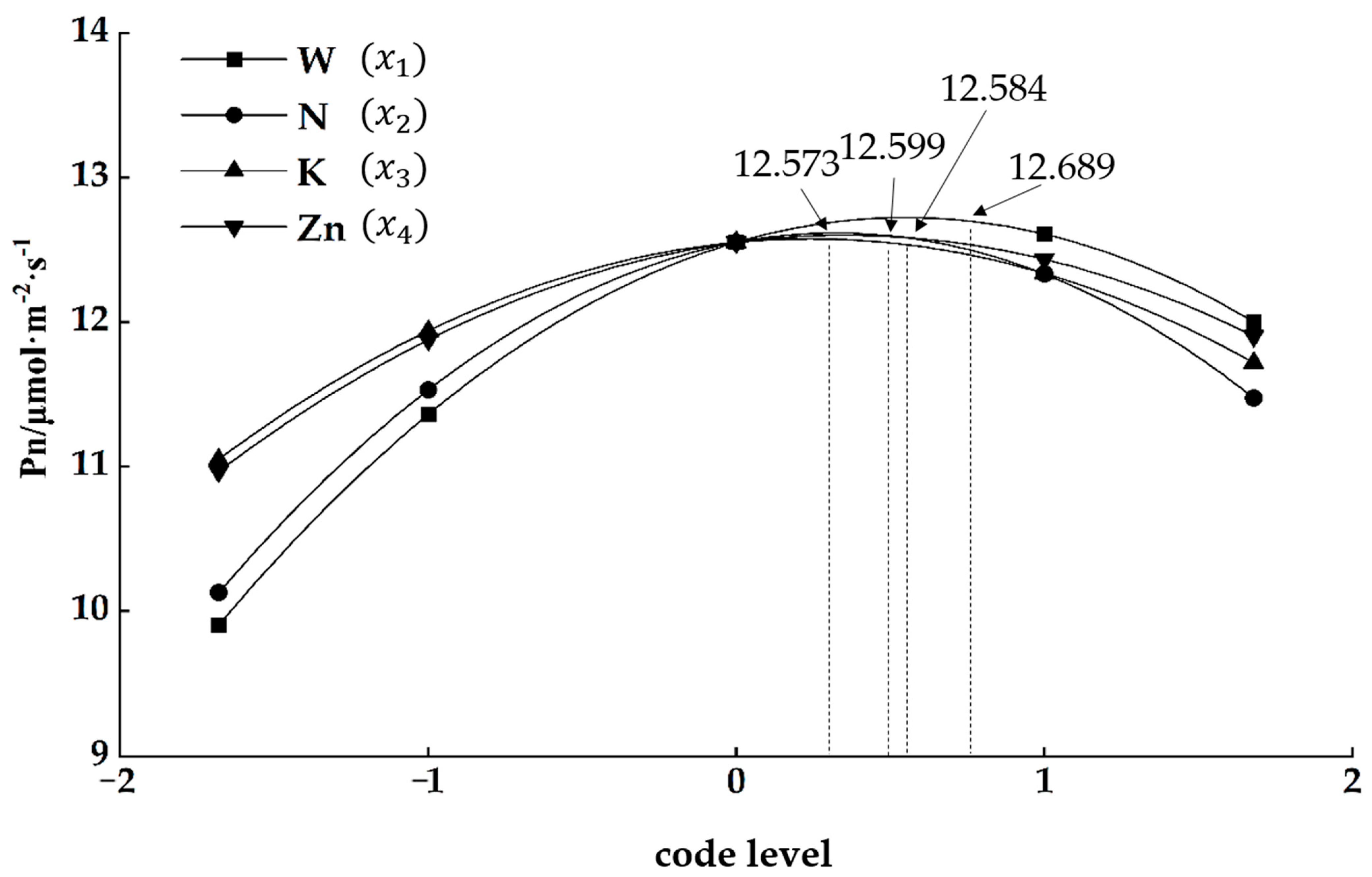
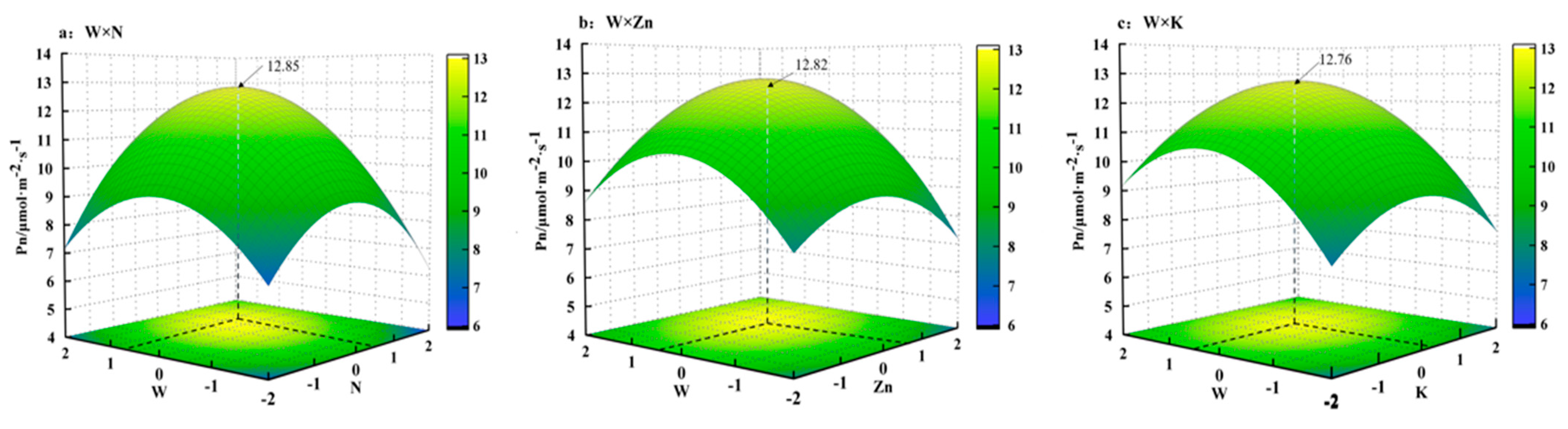
3.2. Regression Analysis
- (1)
- Main effect analysis
- (2)
- Single factor effects
- (3)
- Two-factor interaction effect analysis
- (4)
- Three-factor interaction effect analysis
- (5)
- Optimal combination scheme
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fatichi, S.; Pappas, C.; Ivanov, V.Y. Modeling plant-water interactions: An ecohydrological overview from the cell to the global scale. Wires Water 2016, 3, 327–368. [Google Scholar] [CrossRef]
- Tracy, L.; Silvère, V.C. Speedy stomata, Photosynthesis and plant water use efficiency. New Phytol. 2019, 221, 93–98. [Google Scholar]
- Han, H.; Yang, H.; Ma, X.; Feng, J.; Li, X.; Tissue, D.T. Seasonal maintenance of leaf level carbon balance facilitated by thermal acclimation of leaf respiration but not photosynthesis in three angiosperm species. Environ. Exp. Bot. 2022, 195, 104781. [Google Scholar] [CrossRef]
- Retta, M.; Ho, Q.T.; Yin, X.; Verboven, P.; Berghuijs, H.N.C.; Struik, P.C.; Nicolaï, B.M. A two-dimensional microscale model of gas exchange during photosynthesis in maize (Zea mays L.) leaves. Plant Sci. 2016, 246, 37–51. [Google Scholar] [CrossRef]
- Luo, C.; Wang, R.; Li, C.; Zheng, C.; Dou, X. Photosynthetic characteristics, soil nutrients, and their interspecific competitions in an apple–soybean alley cropping system subjected to different drip fertilizer regimes on the Loess Plateau, China. Agric. Water Manag. 2023, 275, 108001. [Google Scholar] [CrossRef]
- Hosseinzadeh, S.R.; Amiri, H.; Ismaili, A. Evaluation of photosynthesis, physiological, and biochemical responses of chickpea (Cicer arietinum L. cv. Pirouz) under water deficit stress and use of vermicompost fertilizer. J. Integr. Agric. 2018, 17, 2426–2437. [Google Scholar] [CrossRef]
- Chen, Y.H.; Liu, L.; Guo, Q.S.; Zhu, Z.B.; Zhang, L.X. Effects of different water management options and fertilizer supply on photosynthesis, fluorescence parameters and water use efficiency of Prunella vulgaris seedlings. Blological Res. 2016, 49, 1–9. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, H.; Chen, Q.; Ren, J. Coupling effects of depth of film-bottomed tillage and amount of irrigation and nitrogen fertilizer on spring wheat yield. Soil Tillage Res. 2007, 94, 251–261. [Google Scholar] [CrossRef]
- Hyson, D.A. A comprehensive review of apples and apple components and their relationship to human health. Adv. Nutr. 2011, 2, 408–420. [Google Scholar] [CrossRef]
- Wu, Z.; Pan, C. State analysis of apple industry in china. IOP Conf. Ser. Earth Environ. Sci. 2021, 831, 012067. [Google Scholar] [CrossRef]
- Zhou, J.; Zhao, D.; Chen, Y.; Kang, G.; Cheng, C. Analysis of apple producing area changes in China. J. Fruit Sci. 2021, 38, 372–384. [Google Scholar]
- Cao, H.; Wang, H.B.; Li, Y.; Hamani, A.K.M.; Zhang, N.; Wang, X.P.; Gao, Y. Evapotranspiration partition and dual crop coefficients in apple orchard with dwarf stocks and dense planting in arid region, aksu oasis, southern Xinjiang. Agriculture 2021, 11, 1167. [Google Scholar] [CrossRef]
- Ferree, D.C.; Warrington, I.J.; Ebrary, I. Apples: Botany, Production and Uses; CABI Publishing: New York, NY, USA, 2003. [Google Scholar]
- Heping, Z.; Mingyi, Z.; Qi, Z.; Zhifeng, S.; Jinlong, C. Analysis of agricultural irrigation water-using coefficient in Xinjiang arid region. Trans. Chin. Soc. Agric. Eng. 2013, 29, 100–107. [Google Scholar]
- Thevs, N.; Peng, H.; Rozi, A.; Zerbe, S.; Abdusalih, N. Water allocation and water consumption of irrigated agriculture and natural vegetation in the Aksu-Tarim river basin, Xinjiang, China. J. Arid. Environ. 2015, 112, 87–97. [Google Scholar] [CrossRef]
- Fereres, E.; Soriano, M.A. Deficit irrigation for reducing agricultural water use. J. Exp. Bot. 2007, 58, 147–159. [Google Scholar] [CrossRef]
- Wang, H.; Wu, L.; Cheng, M.; Fan, J.; Zhang, F.; Zou, Y.; Chau, H.W.; Gao, Z.; Wang, X. Coupling effects of water and fertilizer on yield, water and fertilizer use efficiency of drip-fertigated cotton in northern Xinjiang, China. Field Crops Res. 2018, 219, 169–179. [Google Scholar] [CrossRef]
- Liu, X.; Li, M.; Guo, P.; Zhang, Z. Optimization of water and fertilizer coupling system based on rice grain quality. Agric. Water Manag. 2019, 221, 34–46. [Google Scholar] [CrossRef]
- Cao, X.S.; Feng, Y.Y.; Li, H.P.; Zheng, H.X.; Wang, J.; Tong, C.F. Effects of subsurface drip irrigation on water consumption and yields of alfalfa under different water and fertilizer conditions. J. Sens. 2021, 2021, 6617437. [Google Scholar] [CrossRef]
- Yang, L.; Lin, Y.; Kong, J.; Yu, Y.; He, Q.; Su, Y.; Li, J.; Qiu, Q. Effects of fertilization and dry-season irrigation on the timber production and carbon storage in subtropical Eucalyptus plantations. Ind. Crops Prod. 2023, 192, 116143. [Google Scholar] [CrossRef]
- Yu, F.; Truong, T.V.; He, Q.; Hua, L.; Su, Y.; Li, J.Y. Dry Season Irrigation Promotes Leaf Growth in Eucalyptus urophylla x E. grandis under Fertilization. Forest 2019, 10, 67. [Google Scholar] [CrossRef]
- Man, J.; Wang, D.; White, P.J.; Yu, Z. The length of micro-sprinkling hoses delivering supplemental irrigation affects photosynthesis and dry matter production of winter wheat. Field Crops Res. 2014, 168, 65–74. [Google Scholar] [CrossRef]
- Chen, X.; Qi, Z.; Gui, D.; Sima, M.W.; Zeng, F.; Li, L.; Li, X.; Feng, S. Responses of cotton photosynthesis and growth to a new irrigation control method under deficit irrigation. Field Crops Res. 2022, 275, 108373. [Google Scholar] [CrossRef]
- Shareef, M.; Gui, D.; Zeng, F.; Waqas, M.; Zhang, B.; Iqbal, H. Water productivity, growth, and physiological assessment of deficit irrigated cotton on hyperarid desert-oases in northwest China. Agric. Water Manag. 2018, 206, 1–10. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, L.; Shen, Q.; Yang, J.; Wu, J. Effects of water stress on photosynthesis, yield, and water use efficiency in winter wheat. Water 2020, 12, 2127. [Google Scholar] [CrossRef]
- Han, K.; Liu, Y.; Zhang, J.; Hao, Y.; Wang, W.; Zhang, L. Effects of water stress on photosynthesis and chlorophyll fluorescence of the sugar beet. J. Agric. Resour. Environ. 2015, 32, 463–470. [Google Scholar]
- Abdalla, M.; Carminati, A.; Cai, G.C.; Javaux, M.; Ahmed, M.A. Stomatal closure of tomato under drought is driven by an increase in soil-root hydraulic resistance. Plant Cell Environ. 2021, 44, 425–431. [Google Scholar] [CrossRef]
- Ma, S.-C.; Zhang, W.-Q.; Duan, A.-W.; Wang, T.-C. Effects of controlling soil moisture regime based on root-sourced signal characteristics on yield formation and water use efficiency of winter wheat. Agric. Water Manag. 2019, 221, 486–492. [Google Scholar] [CrossRef]
- Dos Santos, M.R.; Neves, B.R.; da Silva, B.L.; Donato, S.L.R. Yield, water use efficiency and physiological characteristic of ‘Tommy Atkins’ mango under partial rootzone drying irrigation system. J. Water Resour. Prot. 2015, 7, 1029–1037. [Google Scholar] [CrossRef]
- Wei, Z.; Abdelhakim, L.O.A.; Fang, L.; Peng, X.; Liu, J.; Liu, F. Elevated CO2 effect on the response of stomatal control and water use efficiency in amaranth and maize plants to progressive drought stress. Agric. Water Manag. 2022, 266, 107609. [Google Scholar] [CrossRef]
- Liu, R.-X.; Zhou, Z.-G.; Guo, W.-Q.; Chen, B.-L.; Oosterhuis, D.M. Effects of N fertilization on root development and activity of water-stressed cotton (Gossypium hirsutum L.) plants. Agric. Water Manag. 2008, 95, 1261–1270. [Google Scholar] [CrossRef]
- Heerema, R.J.; VanLeeuwen, D.; Hilaire, R.S.; Gutschick, V.P.; Cook, B. Leaf photosynthesis in nitrogen-starved ‘Western’ pecan is lower on fruiting shoots than non-fruiting shoots during kernel fill. J. Am. Soc. Hortic. Sci. 2014, 139, 267–274. [Google Scholar] [CrossRef]
- Sun, J.; Feng, Z.; Leakey, A.D.B.; Zhu, X.; Bernacchi, C.J.; Ort, D.R. Inconsistency of mesophyll conductance estimate causes the inconsistency for the estimates of maximum rate of Rubisco carboxylation among the linear, rectangular and non-rectangular hyperbola biochemical models of leaf photosynthesis—A case study of CO2 enrichment and leaf aging effects in soybean. Plant Sci. 2014, 226, 49–60. [Google Scholar]
- Mu, X.; Chen, Y. The physiological response of photosynthesis to nitrogen deficiency. Plant Physiol. Biochem. 2021, 158, 76–82. [Google Scholar] [CrossRef]
- Zhu, X.-G.; de Sturler, E.; Long, S.P. Optimizing the distribution of resources between enzymes of carbon metabolism can dramatically increase photosynthetic rate: A numerical simulation using an evolutionary algorithm. Plant Physiol. 2007, 145, 513–526. [Google Scholar] [CrossRef]
- Vos, J.; van der Putten, P.E.L. Effect of nitrogen supply on leaf growth, leaf nitrogen economy and photosynthetic capacity in potato. Field Crops Res. 1998, 59, 63–72. [Google Scholar] [CrossRef]
- Makino, A.; Suzuki, Y.; Ishiyama, K. Enhancing photosynthesis and yield in rice with improved N use efficiency. Plant Sci. 2022, 325, 111475. [Google Scholar] [CrossRef]
- Xu, X.X.; Du, X.; Wang, F.; Sha, J.C.; Chen, Q.; Tian, G.; Zhu, Z.L.; Ge, S.F.; Jiang, Y.M. Effects of potassium levels on plant growth, accumulation and distribution of carbon, and nitrate metabolism in apple dwarf rootstock seedlings. Front. Plant Sci. 2020, 11, 904. [Google Scholar] [CrossRef]
- Ju, F.; Pang, J.; Huo, Y.; Zhu, J.; Yu, K.; Sun, L.; Loka, D.A.; Hu, W.; Zhou, Z.; Wang, S.; et al. Potassium application alleviates the negative effects of salt stress on cotton (Gossypium hirsutum L.) yield by improving the ionic homeostasis, photosynthetic capacity and carbohydrate metabolism of the leaf subtending the cotton boll. Field Crops Res. 2021, 272, 108288. [Google Scholar] [CrossRef]
- Degl’Innocenti, E.; Hafsi, C.; Guidi, L.; Navari-Izzo, F. The effect of salinity on photosynthetic activity in potassium-deficient barley species. J. Plant Physiol. 2009, 166, 1968–1981. [Google Scholar] [CrossRef]
- Kanai, S.; Moghaieb, R.E.; El-Shemy, H.A.; Panigrahi, R.; Mohapatra, P.K.; Ito, J.; Nguyen, N.T.; Saneoka, H.; Fujita, K. Potassium deficiency affects water status and photosynthetic rate of the vegetative sink in green house tomato prior to its effects on source activity. Plant Sci. 2011, 180, 368–374. [Google Scholar] [CrossRef]
- Hiratsuka, S.; Suzuki, M.; Nishimura, H.; Nada, K. Fruit photosynthesis in Satsuma mandarin. Plant Sci. 2015, 241, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Srinivasarao, C.; Shanker, A.K.; Kundu, S.; Reddy, S. Chlorophyll fluorescence induction kinetics and yield responses in rainfed crops with variable potassium nutrition in K deficient semi-arid alfisols. J. Photochem. Photobiol. B Biol. 2016, 160, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Zahoor, R.; Zhao, W.; Dong, H.; Snider, J.L.; Abid, M.; Iqbal, B.; Zhou, Z. Potassium improves photosynthetic tolerance to and recovery from episodic drought stress in functional leaves of cotton (Gossypium hirsutum L.). Plant Physiol. Biochem. 2017, 119, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Sturikova, H.; Krystofova, O.; Huska, D.; Adam, V. Zinc, zinc nanoparticles and plants. J. Hazard. Mater. 2018, 349, 101–110. [Google Scholar] [CrossRef]
- Siddiqui, Z.A.; Parveen, A.; Ahmad, L.; Hashem, A. Effects of graphene oxide and zinc oxide nanoparticles on growth, chlorophyll, carotenoids, proline contents and diseases of carrot. Sci. Hortic. 2019, 249, 374–382. [Google Scholar] [CrossRef]
- Barman, H.; Das, S.K.; Roy, A. Zinc in soil environment for plant health and management strategy. Univers. J. Agric. Res. 2018, 6, 149–154. [Google Scholar] [CrossRef]
- Rossi, L.; Fedenia, L.N.; Sharifan, H.; Ma, X.; Lombardini, L. Effects of foliar application of zinc sulfate and zinc nanoparticles in coffee (Coffea arabica L.) plants. Plant Physiol. Biochem. 2019, 135, 160–166. [Google Scholar] [CrossRef]
- Davarpanah, S.; Tehranifar, A.; Davarynejad, G.; Abadía, J.; Khorasani, R. Effects of foliar applications of zinc and boron nano-fertilizers on pomegranate (Punica granatum cv. Ardestani) fruit yield and quality. Sci. Hortic. 2016, 210, 57–64. [Google Scholar] [CrossRef]
- Fu, C.; Li, M.; Zhang, Y.; Zhang, Y.; Yan, Y.; Wang, Y.A. Morphology, photosynthesis, and internal structure alterations in field apple leaves under hidden and acute zinc deficiency. Sci. Hortic. 2015, 193, 47–54. [Google Scholar] [CrossRef]
- Mahdi, A.; Abdul, S. Effect of plant regulators, zinc nanoparticles and irrigation intervals on leaf content of endogenous hormones and nutrients in sunflower (Helianthus Annuus L.). Plant Arch. 2020, 20, 2720–2725. [Google Scholar]
- Zand-Parsa, S.; Sepaskhah, A.R. Optimal applied water and nitrogen for corn. Agric. Water Manag. 2001, 52, 73–85. [Google Scholar] [CrossRef]
- Rahman, S.M.; Khalil, M.I.; Ahmed, M.F. Yield-water relations and nitrogen utilization by wheat in salt-affected soils of Bangladesh. Agric. Water Manag. 1995, 28, 49–56. [Google Scholar] [CrossRef]
- Yanjun, Y.; Jinhua, Z.; Linjing, C.; Aiqing, J.; Pingyi, G. Effects of fertilizer levels and plant density on chlorophyll contents, its fluorescence and grain yield of setaria italica. Int. J. Agric. Biol. 2018, 20, 737–744. [Google Scholar]
- Liu, Z.; Zhong, H.; Weize, R.; Yang, Z.; Chen, Y.; Liu, Y. Effects of combined application of N, P and K on yield and active ingredient content of rhizome of Paris polyphylla var.chinensis. Chin. Tradit. Herb. Drugs 2019, 50, 6103–6113. [Google Scholar]
- Ren, L.Q. Regression Design and Optimization; Science Press: Beijing, China, 2009. [Google Scholar]
- Du, Q.; Li, J.; Pan, T.; Chang, Y.; Liu, G.; Wang, P.; Zheng, G. The compound effects of water and fertilizer on yield and quality of tomato under drip irrigation. Agric. Res. Arid. Areas 2015, 33, 10–17. [Google Scholar]
- Tuiwong, P.; Lordkaew, S.; Prom-u-thai, C. Improving Grain Zinc Concentration in Wetland and Upland Rice Varieties Grown under Waterlogged and Well-Drained Soils by Applying Zinc Fertilizer. Agronomy 2021, 11, 554. [Google Scholar] [CrossRef]
- Wang, C.; She, H.Z.; Liu, X.B.; Hu, D.; Ruan, R.W. Effects of fertilization on leaf photosynthetic characteristics and grain yield in tartary buckwheat Yunqiao1. Photosynth. Int. J. Photosynth. Res. 2017, 55, 77–84. [Google Scholar] [CrossRef]
- Chhaya; Yadav, B.; Jogawat, A.; Gnanasekaran, P.; Kumari, P.; Lakra, N.; Lal, S.K.; Pawar, J.; Narayan, O.P. An overview of recent advancement in phytohormones-mediated stress management and drought tolerance in crop plants. Plant Gene 2021, 25, 100264. [Google Scholar] [CrossRef]
- Dong, Z.; Zhang, X.; Li, J.; Zhang, C.; Wei, T.; Yang, Z.; Cai, T.; Zhang, P.; Ding, R.; Jia, Z. Photosynthetic characteristics and grain yield of winter wheat (Triticum aestivum L.) in response to fertilizer, precipitation, and soil water storage before sowing under the ridge and furrow system: A path analysis. Agric. For. Meteorol. 2019, 272, 12–19. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, H.S.; Li, Y.F.; Yan, H.J.; Li, J.S. Modeling the effects of plastic film mulching on irrigated maize yield and water use efficiency in sub-humid Northeast China. Int. J. Agric. Biol. Eng. 2017, 10, 69–84. [Google Scholar]
- Liao, Z.; Zeng, H.; Fan, J.; Lai, Z.; Zhang, C.; Zhang, F.; Wang, H.; Cheng, M.; Guo, J.; Li, Z.; et al. Effects of plant density, nitrogen rate and supplemental irrigation on photosynthesis, root growth, seed yield and water-nitrogen use efficiency of soybean under ridge-furrow plastic mulching. Agric. Water Manag. 2022, 268, 107688. [Google Scholar] [CrossRef]
- Zhen, X.; Zhang, Q.; Sanz-Saez, A.; Chen, C.Y.; Dang, P.M.; Batchelor, W.D. Simulating drought tolerance of peanut varieties by maintaining photosynthesis under water deficit. Field Crops Res. 2022, 287, 108650. [Google Scholar] [CrossRef]
- Dos Santos, M.R.; Donato, S.L.R.; Coelho, E.F.; Cotrim, P.R.; De Castro, I.N. Irrigation deficit strategies on physiological and productive parameters of ‘Tommy Atkins’ mango. Rev. Caatinga 2016, 29, 173–182. [Google Scholar] [CrossRef]
- Bhusal, N.; Kim, H.S.; Han, S.-G.; Yoon, T.-M. Photosynthetic traits and plant–water relations of two apple cultivars grown as bi-leader trees under long-term waterlogging conditions. Environ. Exp. Bot. 2020, 176, 104111. [Google Scholar] [CrossRef]
- Chen, H.; Qualls, R.G.; Blank, R.R. Effect of soil flooding on photosynthesis, carbohydrate partitioning and nutrient uptake in the invasive exotic Lepidium latifolium. Aquat. Bot. 2005, 82, 250–268. [Google Scholar] [CrossRef]
- Braun, N.A.; Davis, A.W.; Theg, S.M. The chloroplast tat pathway utilizes the transmembrane electric potential as an energy source. Biophys. J. 2007, 93, 1993–1998. [Google Scholar] [CrossRef]
- Sperling, O.; Karunakaran, R.; Erel, R.; Yasuor, H.; Klipcan, L.; Yermiyahu, U. Excessive nitrogen impairs hydraulics, limits photosynthesis, and alters the metabolic composition of almond trees. Plant Physiol. Biochem. 2019, 143, 265–274. [Google Scholar] [CrossRef]
- Wang, H.; Jin, J.Y. Photosynthetic rate, chlorophyll fluorescence parameters, and lipid peroxidation of maize leaves as affected by zinc deficiency. Photosynth. -Praha 2006, 2, 41–42. [Google Scholar] [CrossRef]
- Tuiwong, P.; Lordkaew, S.; Veeradittakit, J.; Jamjod, S.; Prom-u-thai, C. Efficacy of nitrogen and zinc application at different growth stages on yield, grain zinc, and nitrogen concentration in rice. Agronomy 2022, 12, 2093. [Google Scholar] [CrossRef]
- Niyigaba, E.; Twizerimana, A.; Mugenzi, I.; Ngnadong, W.A.; Ye, Y.P.; Wu, B.M.; Hai, J.B. Winter wheat grain quality, zinc and iron concentration affected by a combined foliar spray of zinc and iron fertilizers. Agronomy 2019, 9, 250. [Google Scholar] [CrossRef]
- Rahman, N.; Schoenau, J. Bioavailability, Speciation, and crop responses to copper, zinc, and boron fertilization in South-Central saskatchewan soil. Agronomy 2022, 12, 1837. [Google Scholar] [CrossRef]
- Wang, H.; Jin, J.; Zhou, W. Effect of zinc application on physiological response of maize to different soil moisture regimes. Plant Nutr. Fertitizer Sci. 2004, 10, 367–373. [Google Scholar]
- Li, J.; Zhou, Y.; Lu, Z.; Cong, R.; Li, X.; Ren, T.; Lu, J. The effects of combined nitrogen and potassium application on photosynthesis and nitrogen allocation in photosynthetic organs of winter oilseed rape (Brassica napus L.) silique wall. J. Plant Nutr. Fertitizer 2022, 28, 869–879. [Google Scholar]
- Zhang, P.; Dong, T.; Jin, H.; Pei, D.; Pervaiz, T.; Ren, Y.; Jia, H.; Fang, J. Analysis of photosynthetic ability and related physiological traits in nodal leaves of grape. Sci. Hortic. 2022, 304, 111251. [Google Scholar] [CrossRef]
- Wang, X.; Cao, H.; Niu, Y.; Shi, X. Influence of water and nitrogen coupling on photosynthesis characteristic of pear-jujube tree. J. Irrig. Drain. 2012, 31, 53–55. [Google Scholar]
- Sun, X.; Chai, Z.; Jiang, P.; Fang, L. Effects on the photosynthetic characteristics and the quality of the apple under the water and nitrogen coupling. Res. Soil Water Conserv. 2010, 17, 271–274. [Google Scholar]
- Li, Z.; Zou, H.; Lai, Z.; Zhang, F.; Fan, J. Optimal drip fertigation regimes improved soil micro-environment, root growth and grain yield of spring maize in arid northwest china. Agronomy 2023, 13, 227. [Google Scholar] [CrossRef]
- Kaur, A.; Singh, K.B.; Gupta, R.K.; Alataway, A.; Dewidar, A.Z.; Mattar, M.A. Interactive effects of nitrogen application and irrigation on water use, growth and tuber yield of potato under subsurface drip irrigation. Agronomy 2023, 13, 11. [Google Scholar] [CrossRef]
- Qu, Z.; Chen, Q.; Feng, H.; Hao, M.; Niu, G.; Liu, Y.; Li, C. Interactive effect of irrigation and blend ratio of controlled release potassium chloride and potassium chloride on greenhouse tomato production in the Yellow River Basin of China. Agric. Water Manag. 2022, 261, 107346. [Google Scholar] [CrossRef]
- Ali, M.M.E.; Petropoulos, S.A.; Selim, D.A.F.H.; Elbagory, M.; Othman, M.M.; Omara, A.E.; Mohamed, M.H. Plant growth, yield and quality of potato crop in relation to potassium fertilization. Agronomy 2021, 11, 675. [Google Scholar] [CrossRef]
- Kuzin, A.; Solovchenko, A. Essential role of potassium in apple and its implications for management of orchard fertilization. Plants 2021, 10, 2624. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Ródenas, R.; Lara, A. The combination of K+ deficiency with other environmental stresses: What is the outcome? Physiol. Plant. 2019, 165, 264–276. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| () | 800.0 | 150.0 | 300.0 | 22.5 |
| (1) | 700.0 | 135.0 | 270.0 | 15.8 |
| (0) | 550.0 | 112.5 | 225.0 | 11.3 |
| (−1) | 400.0 | 90.0 | 180.0 | 6.7 |
| () | 300.0 | 75.0 | 150.0 | 0 |
| 148.6 | 22.3 | 44.6 | 6.7 | |
| Factors | Stage | Factor Levels | ||||
|---|---|---|---|---|---|---|
| −1.682 | −1 | 0 | 1 | 1.682 | ||
|
| AFS | 26.25 | 31.50 | 39.25 | 47.25 | 52.50 |
| YFS | 105.00 | 126.00 | 157.00 | 189.00 | 210.00 | |
| FES | 288.75 | 346.50 | 431.75 | 519.75 | 577.50 | |
| FRS | 105.00 | 126.00 | 157.00 | 189.00 | 210.00 | |
| TGP | 525.00 | 630.00 | 785.00 | 945.00 | 1050.00 | |
|
| AFS | 26.25 | 31.50 | 39.38 | 47.25 | 52.50 |
| YFS | 131.25 | 157.50 | 196.88 | 236.25 | 262.50 | |
| FES | 105.00 | 126.00 | 157.40 | 189.00 | 210.00 | |
| FRS | 0 | 0 | 0 | 0 | 0 | |
| TGP | 262.50 | 315.00 | 393.75 | 472.50 | 525.00 | |
|
| AFS | 0.53 | 0.70 | 0.96 | 1.23 | 1.40 |
| YFS | 1.58 | 2.10 | 2.89 | 3.68 | 4.20 | |
| FES | 6.30 | 8.40 | 11.55 | 14.70 | 16.80 | |
| FRS | 2.10 | 2.80 | 3.85 | 4.90 | 5.60 | |
| TGP | 10.50 | 14.00 | 19.25 | 24.50 | 28.00 | |
|
| AFS | 0 | 7.10 | 17.50 | 27.90 | 35.00 |
| YFS | 0 | 7.10 | 17.50 | 27.90 | 35.00 | |
| FES | 0 | 7.10 | 17.50 | 27.90 | 35.00 | |
| FRS | 0 | 0 | 0 | 0 | 0 | |
| TGP | 0 | 21.30 | 52.45 | 83.70 | 105.00 | |
| No. | Factor Code | Implementation | |||||||
|---|---|---|---|---|---|---|---|---|---|
|
(mm) |
(kg/hm2) |
(kg/hm2) |
(kg/hm2) | ||||||
| T1 | 1 | 1 | 1 | 1 | 700 | 135.0 | 270 | 15.825 | 12.960 a |
| T2 | 1 | 1 | −1 | −1 | 700 | 135.0 | 180 | 6.675 | 11.226 ef |
| T3 | 1 | −1 | 1 | −1 | 700 | 90.0 | 270 | 6.675 | 10.428 hi |
| T4 | 1 | −1 | −1 | 1 | 700 | 90.0 | 180 | 15.825 | 10.797 gh |
| T5 | −1 | 1 | 1 | −1 | 400 | 135.0 | 270 | 6.675 | 10.227 ij |
| T6 | −1 | 1 | −1 | 1 | 400 | 135.0 | 180 | 15.825 | 10.106 ij |
| T7 | −1 | −1 | 1 | 1 | 400 | 90.0 | 270 | 15.825 | 10.062 ij |
| T8 | −1 | −1 | −1 | −1 | 400 | 90.0 | 180 | 6.675 | 9.517 k |
| T9 | −1.682 | 0 | 0 | 0 | 300 | 112.5 | 225 | 11.250 | 9.900 jk |
| T10 | 1.682 | 0 | 0 | 0 | 800 | 112.5 | 225 | 11.250 | 11.692 d |
| T11 | 0 | −1.682 | 0 | 0 | 550 | 75.0 | 225 | 11.250 | 10.119 ij |
| T12 | 0 | 1.682 | 0 | 0 | 550 | 150.0 | 225 | 11.250 | 11.168 efg |
| T13 | 0 | 0 | −1.682 | 0 | 550 | 112.5 | 150 | 11.250 | 11.025 ef |
| T14 | 0 | 0 | 1.682 | 0 | 550 | 112.5 | 300 | 11.250 | 11.431 de |
| T15 | 0 | 0 | 0 | −1.682 | 550 | 112.5 | 225 | 0 | 10.897 fg |
| T16 | 0 | 0 | 0 | 1.682 | 550 | 112.5 | 225 | 22.500 | 11.659 d |
| T17 | 0 | 0 | 0 | 0 | 550 | 112.5 | 225 | 11.250 | 12.733 ab |
| T18 | 0 | 0 | 0 | 0 | 550 | 112.5 | 225 | 750.000 | 12.580 bc |
| T19 | 0 | 0 | 0 | 0 | 550 | 112.5 | 225 | 750.000 | 12.643 abc |
| T20 | 0 | 0 | 0 | 0 | 550 | 112.5 | 225 | 750.000 | 12.752 ab |
| T21 | 0 | 0 | 0 | 0 | 550 | 112.5 | 225 | 750.000 | 12.677 ab |
| T22 | 0 | 0 | 0 | 0 | 550 | 112.5 | 225 | 750.000 | 12.265 c |
| T23 | 0 | 0 | 0 | 0 | 550 | 112.5 | 225 | 750.000 | 12.575 bc |
| Source | Df | SS | MS | F Value | p Value |
|---|---|---|---|---|---|
| 1 | 5.314 | 5.314 | 82.62 | 0.0001 ** | |
| 1 | 2.197 | 2.197 | 34.16 | 0.0001 ** | |
| 1 | 0.540 | 0.540 | 8.39 | 0.015 * | |
| 1 | 1.061 | 1.061 | 16.49 | 0.002 ** | |
| 1 | 5.103 | 5.103 | 79.33 | 0.0001 ** | |
| 1 | 6.109 | 6.109 | 94.97 | 0.0001 ** | |
| 1 | 2.717 | 2.717 | 42.23 | 0.0001 ** | |
| 1 | 2.486 | 2.486 | 38.65 | 0.0001 ** | |
| 1 | 0.609 | 0.609 | 9.47 | 0.011 * | |
| 1 | 0.061 | 0.061 | 0.95 | 0.352 | |
| 1 | 0.352 | 0.352 | 5.47 | 0.039 * | |
| Regression | 11 () | 26.225 () | 2.384 | = 37.06 | 0.0001 ** |
| Residual error | 11 () | 0.708 () | 0.064 | ||
| Lack-of-fit | 5 () | 0.546 () | 0.109 | = 4.04 | 0.059 |
| Error | 6 () | 0.162 () | 0.027 | ||
| Sum | 22 | 26.225 |
| Code | ||||||||
|---|---|---|---|---|---|---|---|---|
| t | f | t | f | t | f | t | f | |
| 1.682 | 36 | 0.279 | 25 | 0.194 | 25 | 0.194 | 18 | 0.140 |
| 1 | 51 | 0.395 | 46 | 0.357 | 36 | 0.279 | 41 | 0.318 |
| 0 | 39 | 0.302 | 49 | 0.380 | 39 | 0.302 | 41 | 0.318 |
| −1 | 3 | 0.023 | 9 | 0.070 | 25 | 0.194 | 18 | 0.140 |
| −1.682 | 0 | 0 | 0 | 0 | 4 | 0.031 | 0 | 0 |
| total | 129 | 1 | 129 | 1 | 129 | 1 | 129 | 1 |
| Weighted average | 0.843 | 0.621 | 0.369 | 0.565 | ||||
| 95% Confidence interval | 0.720–0.965 | 0.486–0.756 | 0.196–0.542 | 0.411–0.720 | ||||
| Application rate | 658.00–694.75 mm | 123.44–129.51 kg/hm2 | 233.82–249.39 kg/hm2 | 13.13–14.54 kg/hm2 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, W.; Duan, X.; Wang, Q.; Wei, X.; Wei, K.; Li, Y.; Guo, X.
Effect of Drip Irrigation, N, K, and Zn Coupling on
Mu W, Duan X, Wang Q, Wei X, Wei K, Li Y, Guo X.
Effect of Drip Irrigation, N, K, and Zn Coupling on
Mu, Weiyi, Xiaoxian Duan, Quanjiu Wang, Xuesong Wei, Kai Wei, Yuanxin Li, and Xin Guo.
2023. "Effect of Drip Irrigation, N, K, and Zn Coupling on