
Allelopathic Effects of the Invasive Prosopis juliflora (Sw.) DC. on Native Plants: Perspectives toward Agrosystems


1
Department of Biological and Environmental Sciences, College of Arts and Sciences, Qatar University, Doha P.O. Box 2713, Qatar
2
Environmental Science Program, Department of Biological and Environmental Sciences, College of Arts and Sciences, Qatar University, Doha P.O. Box 2713, Qatar
3
Biological Science Program, Department of Biological and Environmental Sciences, College of Arts and Sciences, Qatar University, Doha P.O. Box 2713, Qatar
*
Author to whom correspondence should be addressed.
Agronomy 2023, 13(2), 590; https://doi.org/10.3390/agronomy13020590
Submission received: 14 January 2023
/
Revised: 10 February 2023
/
Accepted: 13 February 2023
/
Published: 18 February 2023
(This article belongs to the Special Issue Application of Allelopathy in Sustainable Agriculture)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Prosopis juliflora (Sw.) DC. Is highly recognized for its invasive character and its detrimental effects on different plant species across the world. In this study, a field survey was performed to assess the impact of P. juliflora on the surrounding native plants, followed by the effect of different concentrations of its aqueous leaf extract on seed germination and seedling growth of selected Qatari flora. Our field observations indicated a high prevalence of new recruitments of P. juliflora than other native-associated plants in the study site. Germination percentage and/or seed radicle length of the test species: Acacia tortilis (Forssk.) Hayne, Prosopis cineraria (L.) Druce, Sueda aegyptica (Hasselq.) Aellen, Halopeplis perfoliata (Forssk.) Bunge ex Schweinf. & Asch, and P. juliflora were significantly (p ≤ 0.05) reduced by the extracts at different concentration levels. Moreover, the germination rate of A. tortilis was not affected; however, the seed germination of P. cineraria was significantly affected at concentrations of 6 and 8 mg mL−1. The seed radicle lengths at 2 mg mL−1 were significantly reduced for both plant species. The seed germination and radicle length of S. aegyptica and H. perfoliata were significantly decreased at 2 mg mL−1 of extract of P. juliflora. Furthermore, the height and biomass of the seedlings of Aeluropus lagopoides (L.) Trin. ex Thwaites, Caroxylon imbricatum (Forssk.), and Tetraena qatarensis (Hadidi) Beier & Thulin were significantly reduced by the aqueous extracts at 6 and 8 mg mL−1. The aqueous extract reduced the seed germination and seedling growth at 6 and 8 mg mL−1, indicating an autotoxic effect. Such allelopathic behavior of P. juliflora designates the potential investigation of using its allelochemicals as naturally based herbicides in agrosystems.
1. Introduction
Allelopathic plants are ecologically important as they are key players in disturbing and re-shaping the natural biodiversity of plants growing close to them. Allelopathic plants are also involved in shaping the composition of native vegetation and improving species richness by developing microenvironments that are suitable for the growth of certain species and their interactions [1]. Allelochemicals have different roles in the environment; some might be involved in the plant defense mechanism, in chelating nutrients available for plants, and in regulating populations of soil fauna and flora, while others might have effects on the decomposition cycles, and consumption of plants by other organisms, thus affecting the nutrient cycling and interactions at different trophic levels [2]. Allelochemicals, in general, have been known to affect the germination of seeds and pollen, the seedling growth, cell division, uptake of nutrients, photosynthesis, and function of enzymes negatively [3].
P. juliflora (Sw.) DC is one such allelopathic plant. It is a xerophyte and has adapted strategies to grow in dry and arid regions with alkaline soils [4]. It is also able to grow on sand dunes and in regions with low vegetation; hence, it can be appropriate in fighting against desertification [5]. It is reported that an area invaded by P. juliflora showed a greater species richness, diversity, and dominance as compared to regions with no P. juliflora invasion [6].
The plant grows into a relatively large canopy, which makes the growth of associated plants difficult. P. juliflora is an inhabitant plant of Central and South America. Despite its capability to decline the growth of other native plant species, over the past 100–150 years, the species of Prosopis have been widely introduced by human beings for different purposes resulting in the reduction of other plant diversity [7]. Due to its invasive nature, Prosopis has spread to different regions like Africa, Asia, and Australia [8]. P. juliflora was introduced as early as the 1970s and 1980s for different purposes, such as in regions vulnerable to desertification or deforestation. Furthermore, it was also introduced as a source of fuelwood and fodder in rural areas [9]. P. juliflora is now considered to be one of the 100 world’s most invasive species [10] and is extensively studied for its allelopathy on different plant types. Its extracts were used to test the impact of allelochemicals on the germination of the seeds as well as the growth of rice [11]. P. juliflora is composed of different allelochemicals, such as derivatives of phenolic compounds, alkaloids, and water-soluble compounds. Some of the allelochemicals are juliflorine, julifloricine, juliprosopine, mesuitol, syringin, L-tryptophan, (-)-lariciresinol, and juliprosopinal [10]. Almost all parts of the plant produce metabolites with allelopathic effects. For example, pollen [12], fruits [13], bark [14], leaves [15,16], pods [17], roots, and seeds [18]. The composition of the metabolites plays a major role in determining their role in an environment. For instance, none of the metabolites have the same composition, nor do they have concentrations that are similar; they differ from species to species. Due to this variation in the complexity of metabolites, these types of plants could be used for different purposes in agriculture [19].
Considering the allelopathic nature of P. juliflora, it is safe to mention here that due to the presence of efficient bioactive secondary metabolites in this plant, it could be exploited for its use as an efficient bio-herbicide. For instance, L-tryptophan, syringing, and (-)- lariciersinol isolated from P. juliflora were used to inhibit the growth of weeds [20,21,22]. Similarly, phenolics isolated from P. juliflora showed inhibition against weed plants [23]. These allelochemicals are known to have a similar mode of action as studied in the synthetic herbicides, thereby having the ability to develop novel herbicides by fusing efficient allelochemicals and modes of action from the herbicides [24].
It was brought to Qatar, like other Arabian Gulf countries, for eco-friendly purposes like greenery or desertification control [25,26]. P. juliflora is distributed in Southern and Northern Qatar in regions that include Mazra’ at Turayna, Al-Ghuwayriyah, Sidriyat Al Kheesah, Abu Al Dhalouf, and Al Khor [27]. Several research projects have been carried out to investigate the allelopathic effects of this invasive plant on other plants. In one research, it was found that the seed germination along with the radicle length of wheat was negatively affected when compared with the control [28]. In another study, extracts of leaves, flowers, and fruits inhibited the seed germination and seedlings growth of Zea mays, Triticum aestivum, and Albizia lebbeck [29]. Furthermore, P. juliflora is also reported to significantly reduce the seed germination of Bauhinia racemose, Drypetes sepiaria, Flueggea leucopyrus, Salvadora persica, Ziziphus mauritiana, Hordeum vulgare, and Medicago sativa [30,31]. In the same experiment, the seeds were allowed to germinate in soil from a place where the allelopathic plant P. juliflora was present. A significant reduction in the percentage of germination compared with the germination in natural forest soil indicated the presence of allelochemicals or secondary plant metabolites in the soil taken from a region with P. juliflora.
P. juliflora can also be toxic to itself (autotoxicity). It is reported that roots, shoots, and fruit extracts of P. juliflora are involved in the autotoxicity process at higher concentrations [32]. Other researchers concluded that the autotoxicity of P. juliflora is very little or does not exist because of the higher growth rate of seedlings of the same species under the canopy of P. juliflora [33]. The contradictory results could be justified by different environmental factors that might affect the biological and chemical properties of the bioactive compounds.
Studying the allelopathic effects of P. juliflora on the local Qatari plant species is an important first step in evaluating its harmful effects on the conservation of the existing plant community. No previous studies have been performed in Qatar to evaluate the allelopathic effects of P. juliflora on native plant species. Since P. juliflora is seen growing abundantly in different regions of Qatar, this research could be a good initiative to have the basic knowledge for future studies. Keeping in mind the endangered status of certain local native plants like P. cineraria and the arid land growth limitation, other researchers can use this knowledge to plan ways to protect rare species like P. cineraria and other species from the invasion of P. juliflora. Therefore, the current research was established to achieve the following objectives: (1) conduct field surveys to investigate the survival of seedlings of certain native species encountered in the protected field (inside Qatar University (QU) Campus), (2) investigate the consequences of water leaf extract of P. juliflora on seed germination of selected native plant species, and (3) investigate the effects of leaf leachate of P. juliflora on seedling survival of selected native plant species.
2. Materials and Methods
2.1. Study Site
The fieldwork was accomplished in one of the protected fields (number III) inside the QU campus. QU is located on the northern side of the capital Doha at 25.3747° N, 51. 4903° E. The main habitat and associated vegetation are characterized by compact soils. The soil is sandy clay and alkaline in nature with an electrical conductivity (µS/cm) of 5.59 ± 1.59 with a mean organic matter of 12.3% [34]. For experiments, the plant materials of P. juliflora were collected from the trees grown in the protected field.
2.2. Seeds Collection
Field trips were arranged to collect seeds of several Qatari native plants, in addition to seeds of P. juliflora. Collected seeds were cleaned and dried before storing them in the refrigerator.
From earlier preliminary field visits, the following plant species were observed to be encountered in the same area around P. juliflora (Sw.) DC: Prosopis cineraria (L.) Druce, Acacia nilotica (L.) Delile, Acacia tortilis (Forssk.) Hayne, Limonium axillare (Forssk.) Kuntze, Sueda aegyptiaca (Hasselq.) Aellen, Tetraena qatarense (Hadidi) Beier & Thulin, and Halopeplis perfoliata (Forssk.) Bunge ex Schweinf. & Asch. The seeds of these plants were collected and screened for their quality and viability. Few seeds from each batch were selected for a test seed germination experiment in Petri plates with sterile moist filter paper. If the germination percentage of seeds was above 80%, the seeds were considered viable [35].
2.3. A Preliminary Field Survey
To detect the magnitude of the invasiveness of P. juiliflora and its impact on associated plant species, we conducted a field survey. Eight line transects, each 100 m long, were used. The line transects started at a random point and were directed from there in 4 different directions: north, south, east, and west. The line transects were maintained at a 10 m distance from each other. The survey targeted seedlings of species with tree or shrubby tree growth habits. The seedlings of plant species that touch the line transects have been visually identified and counted. The survey was conducted in November 2018 and repeated in March 2019. The average total counts of each plant species were calculated.
2.4. Preparation of Aqueous Extract of Leaves of Prosopis juliflora
To prepare the aqueous extract, a similar method as described by Siddique et al. (2009) and Omer et al. (2017), where the powder of P. juliflora was dissolved in water was used, the aqueous extract of P. juliflora leaves was prepared based on the following method: the leaves of P. juliflora were collected from QU Campus; they were air-dried and were ground to a fine powder. The crude water extract of plant leaves was made by soaking dried plant parts in distilled water (10 g L−1). The mixture was incubated for 1 day in a water bath (OLS200; Grant Instruments Cambridge Ltd., England) set at 50 °C with 102 shakes per minute [28,36]. The mixture was filtered using Whatman filter paper after 1 day. The collected brownish filtrate was considered stock extract (10 mg mL−1). The full-strength solution was diluted to achieve the following concentrations: 8 mg mL−1, 6 mg mL−1, 4 mg mL−1, and 2 mg mL−1 under sterile conditions. Distilled water was used as untreated control. For smaller seeds, the following dilutions from the stock solutions were prepared: 0.5 mg mL−1, 1 mg mL−1, 2 mg mL−1, 4 mg mL−1, and 6 mg mL−1. The full-strength stock solution was left in the refrigerator until being used for experimental work within 1 week.
2.5. Surface Sterilization and Preparation of Seeds Prior to Treatment
Prior to any experiment, the seeds were surface sterilized using 3–5% sodium hypochlorite (NaClO) (Peros, Turkey) for 1–2 min based on the seed size. The small seeds (<2 mm length) were subjected to 3% at a 1 min duration while large seeds (>2 mm length) were subjected to 5% NaClO (Peros, Turkey) and 2 min duration [37,38]. The seeds were then washed with sterile distilled water twice, followed by drying with filter paper. Due to hard seed coats, seeds of A. tortilis, P. cineraria, and P. juliflora were subjected to a cut at the bottom near the emerging radicle site using a sterile scalpel (Sigma-Aldrich, Germany) [39]. This method was tested and experienced prior to the experimental work.
2.6. Effect of Aqueous Extracts on Seed Germination and Seedling Growth
2.6.1. Effect on Seed Germination of Native Plants
Aqueous extract concentrations (2 mg mL−1, 4 mg mL−1, 6 mg mL−1, 8 mg mL−1, and 10 mg mL−1) were tested on germination seeds of A. tortilis, P. cineraria, S. aegyptiaca, H. perfoliata, and P. juliflora. Two Whatman No. 1 filter papers were positioned in Petri dishes (9 cm diameter). The filter papers were moistened with 3 mL of extracts. Distilled water was used as untreated control. Five replicates were designed for each treatment. The experimental design was completely random and was repeated once. The plates were incubated at a temperature of 24 ± 2 °C with a 15 hr day and 9 hr night cycle (photon flux density minimum of 350 ± 50 µmol m−2 s−1). The germination was observed and counted daily for a maximum period of two weeks. Accumulative germination was reported for all treatments. Radicle lengths were measured after two weeks using a digital caliper [40,41].
2.6.2. Effect of Leaf Leachates of P. juliflora on Seedlings Growth of Native Plants
Fresh leaves of P. juliflora were collected from the dropped leaves under the trees located at the study site, which represents the upper part of the topsoil. The protocol was modified before applying it [42]. The leaves were air-dried and then used to prepare leachates. Of air-dried leaves, 10 g were immersed in 1 L of distilled water (Milli-Q ultra pure water system, Sigma-Aldrich, Germany) for 1 day and then filtered by double-layered cheesecloth [42,43]. The obtained filtrate was considered full-strength leachate (10 mg mL−1). It was further diluted to 4 mg mL−1, 6 mg mL−1, and 8 mg mL−1. Seedlings of A. lagopoides, C. imbricatum, and T. qatarensis with the same growth size and height were collected with a bunch of soil to keep the same natural soil around the roots. One specimen from each species were deposited as voucher specimens at Qatar University Herbarium. Seedlings were transplanted into 15-cm plastic pots filled with soil to the lower rim. The pots were labeled as per the dilution from T1 (stock) to T3 (least concentrated) and were irrigated with tap water. The seedlings were left under greenhouse conditions for two weeks. For each plant species, 10 replicates were performed. The labeled pots were organized randomly under greenhouse conditions (24 ± 2 °C with 15-hr day and 9-hr night cycle at photon flux density minimum of 350 ± 50 µmol m−2 s−1). Control pots were prepared and irrigated with only 50 mL water each. Other pots were treated with 50 mL of the allocated extract concentration thrice a week for two weeks, followed by irrigation with tap water. The pots were regularly checked for any sign of damage, weak growth, and/or disease. After one month, the roots of the plants were cut, cleaned, and dried in an oven (Gallenkamp, England) for 3 days at 80 °C and then weighed to calculate the dry matter biomass.
2.6.3. Autotoxicity of the Aqueous Extract
Aqueous extracts of leaves of P. juliflora were prepared with five concentrations (0, 2, 4, 6, and 8 mg mL−1) and were applied to seeds of P. juliflora. A total of 10 seeds were distributed in each Petri plate, with five replicates in total. Seed germination was monitored daily for two weeks, and accumulative germination (%) was calculated. After two weeks, the radicle of the seed was measured using a digital caliper.
2.7. Data Analysis
3. Results
3.1. Field Survey
In the field survey, recruitment seedlings of four plant species were found, namely Ziziphus spp., Tamarix sp., A. tortilis, and P. juliflora. The survey was conducted to preliminary investigate the recruitments of seedlings for the four encountered species that have the same growth habits (shrubby trees). A prominent increase in the recruitment of P. juliflora seedlings has been recorded (Figure 1).
3.2. Effect of Aqueous Extract of P. juliflora on Seed Germination Potential
3.2.1. Effect of Aqueous Extract of P. juliflora on Seed Germination of Native Plants
The seed germination of A. tortilis was not significantly affected by any concentration of the aqueous extract of P. juliflora (Figure 2A). Furthermore, the effect of the extract on the seed germination of P. cineraria also showed no significant differences at 2 and 4 mg mL−1 as compared to the control. However, a slight (about 10%) but significant reduction was exerted on seed germination at higher concentrations of 6 and 8 mg mL−1 (Figure 2B). Moreover, the seed germination percentage of S. aegyptiaca was negatively affected by the extract. A significant reduction in germination started at 2 mg mL−1, which significantly decreased with the application of an increasing extract concentration (Figure 2C). Lastly, the seed germination of H. perfoliata showed a significant difference as compared to the control under 4 mg mL−1 and at higher concentrations (Figure 2D).
3.2.2. Effect of Aqueous Extract of P. juliflora on Radicle Length of Native Plants
The effect of extracts of P. juliflora showed a significant difference in the radicle length of the seedlings of A. tortilis starting from a concentration as low as 2 mg mL−1 compared to control and with stronger significant effects at higher concentrations (Figure 3A). Furthermore, the radicle length of P. cineraria was significantly reduced, starting from the lowest concentration (2 mg mL−1) and with greater reduction at higher concentrations (Figure 3B). The seed radicle length of S. aegyptiaca was also negatively affected by the extract (Figure 3C). A significant reduction in the parameter started at the lowest concentration and continued to decrease with the application of an increasing extract concentration.
Figure 4 shows pictures of the affected emerging radicles of seeds of P. cineraria (A) and S. aegyptiaca (B) after two weeks of treatment.
3.2.3. Autotoxicity; Effect of Aqueous Extract of P. juliflora on Germination Potential of P. juliflora
The auto allelopathy results showed significant differences at higher concentrations (6 and 8 mg mL−1) as compared to the control, as shown in Figure 5A. Similarly, the radicle lengths of P. juliflora were significantly reduced at higher concentrations (Figure 5B).
The effect of the extract of P. juliflora on the germination of their own seeds is illustrated in Figure 6. At 2 and 4 mg mL−1, the seeds among replicates responded differently. This heterogeneity of data exerted less significance compared to the control.
3.3. Effect of Leaf Leachate of P. juliflora on Seedling Growth of Selected Plant Species
3.3.1. Effect of Leaf Leachate of P. juliflora on Seedling Growth of A. lagopoides
Although no significant differences in plant height occurred due to treatments (Figure 7A), a significant reduction in dry matter biomasses was found at concentrations of 6 mg mL−1 and 8 mg mL−1 as compared to control plants (Figure 7B). Figure 7C shows pictures of seedlings of A. lagopoides at the time of harvest post-one-month treatment.
3.3.2. Effect of Leaf Leachate of P. juliflora on Seedling Growth of C. imbricatum
The leaf leachate of P. juliflora significantly declined the seedling growth and dry matter biomass of C. imbricatum only under the highest concentration treatment (8 mg mL−1), as shown in Figure 8A,B. Pictures of the seedlings of C. imbricatum at harvesting time post-treatment of one month are shown in Figure 8C.
3.3.3. Effect of Leaf Leachate of P. juliflora on Seedling Growth of T. qatarensis
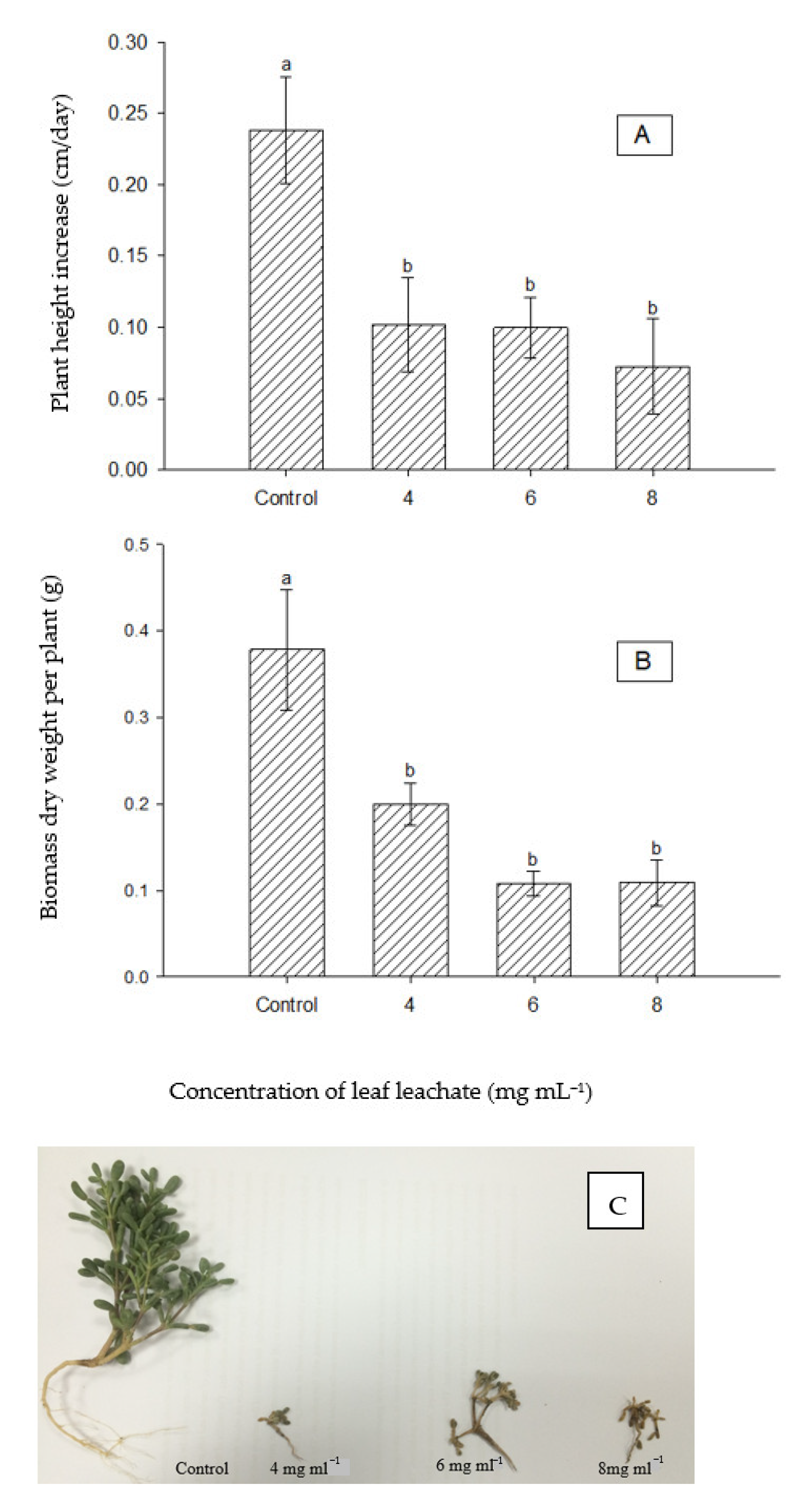
The leaf leachate of P. juliflora significantly reduced the seedling’s growth of T. qatarensis at all treatment levels as compared to the control. The growth in plant height of control plants was about 0.25 cm per day compared to less than 0.1 cm per day under all extract treatments (Figure 9A). The dry matter biomass was also significantly reduced to half or lower under any of the extract treatments compared to control plants (Figure 9B). Pictures of the plants post-one-month treatment at harvesting time are shown in Figure 9C.
4. Discussion
As per the field survey investigation, the recruitment seedlings of P. juliflora predominated the area as compared to other plants encountered in the protected field. This field survey is just a preliminary investigation giving an idea about the new recruitments of P. juliflora. A higher number of recruitments of P. juliflora could be supported by its characteristics of high seed germination rate, efficient seed dispersal mechanisms, and broad ecological adaptability [46]. In the future, a survey considering variables such as pH, soil moisture, elevation, and others could help in obtaining the relation between species richness and occurrence [47].
It is important to keep in mind that different high or low concentrations of secondary metabolites in an extract of any allelopathic plant will be a factor in inhibiting the germination potential of seeds and the healthy growth of seedlings. In certain cases, higher concentrations are observed to inhibit seed germination and the growth of the seedlings, while in other cases, the same is achieved with low extract concentrations.
The allelopathic effect of P. juliflora on germination is generally reported as dose-dependent, which means the allelopathic effect normally increases with extract concentration. One of the publications reported the reduction of seed germination of Cynodon dactylon with increasing aqueous extract concentration. At higher concentrations (60 g L−1) of the extract, seed germination completely declined [48]. Another publication reported a reduction in seed germination of Triticum aestivum by increasing the concentration of extract. At the highest concentration (0.1 g/L), the seed germination percentage was inhibited by 40.75% [28].
Further experiments on native Qatari plants collected from the same site where P. juliflora is encountered indicated a range of allelopathic effects on different selected plants. The inhibitory effect of the aqueous extract could not be seen significantly on the seed germination of A. tortilis and with a minor effect on P. cineraria (Figure 2A,B). However, the inhibitory and detrimental effects of the aqueous extract could be seen in the radicle lengths of both species. Radicle lengths were significantly reduced, starting at the lowest concentration (Figure 3A,B). Since A. tortilis and P. cineraria have relatively large seeds and can imbibe more water/extract than smaller seeds, therefore, the embryos of the large seeds will have less contact with the active allelopathic chemicals compared to small-sized seeds; hence, higher seed germination with the least allelopathic impact of the aqueous extract. Another reason that reduced the allelopathic effect of the extract might be the hard seed coat of both species, which might hinder the imbibition of the extract and then delay or prevent the germination process. On the other side, as the germinated radicle is expected to be more exposed to the extract, the radicle length of germinated seeds of both species significantly declined compared to control plants. A similar effect of the aqueous extract of P. juliflora on larger and smaller seeds was reported. One of the publications reported the inability of extract to inhibit the seed germination of A. nilotica (large seeds) even at a higher percentage of 6% [49].
Aqueous extract of P. juliflora affected the germination percentage of all studied smaller seeds. A significant decline in the germination of seeds and radicle length of S. aegyptiaca with an increase in extract concentration indicated their sensitivity to the allelochemicals present in the leaf extract of P. juliflora (Figure 2C and Figure 3C). Research conducted on species of genus Suaeda reported a similar decline in germination percentage with increasing extract concentration due to an invasive plant Solidago canadensis, which inhibited the germination of Suaeda glauca at a concentration of 1:60 (M:M) prepared by diluting ethanolic extracts (25 g/L) in distilled water [50].
In comparison to S. aegyptiaca, the inhibitory effect of the aqueous extract of P. juliflora on the germination percentage of H. perfoliata was minor (Figure 2D). Unfortunately, the effect of aqueous extract on the radicle length of H. perfoliata was not monitored due to extremely smaller radicle lengths. Hence, it is doubtful to conclude whether the radicle length has any detrimental effect due to extract treatment or not. Halophytes and their interactions with allelochemicals are not widely studied, but some publications have reported an inhibitory effect of allelopathic plants on halophytes. One such publication has reported that P. juliflora limited the growth and distribution of Suaeda fructicosa. The publication reported two possible reasons for inhibition, one being the presence of allelochemicals and the other being the large canopy of P. juliflora covering the halophyte [51]. Similarly, in another research, halophyte Salsola baryosma growth was suppressed under the canopy of P. juliflora as compared to the marginal regions [33]. The relationship between halophytes and allelopathic plants is not clear as it is difficult to assume whether a certain growth inhibition in plants is caused by allelochemicals or by the accumulation of a specific ion that results in toxicity [52].
The allelochemicals not only inhibit or decline the growth of other plant species but also their own. This auto-toxic property of P. juliflora was observed in experimental work to a certain extent. The seed germination and radicle length showed slight variations in the treatment groups as compared to the control, but higher concentrations of 6 and 8 mg mL−1 exerted significant reductions in both growth parameters (Figure 5 and Figure 6). Therefore, a greater extent of allelochemicals might appear under higher concentrations of the extract. Coinciding with our findings, research found that the self-inhibitory action of P. juliflora occurred at higher extract concentrations of 75% and 100% by the reduction in seed germination and root and shoot lengths, respectively [32]. Autotoxicity might inhibit the growth of the same kind of plants, but certain allelopathic plant species have adopted this phenomenon to occupy a competitive ecological position with respect to other plant species [53]. It is reported in another research that P. juliflora is a key factor in inhibiting the growth of native plants of Eritrea. It is reported in the literature that the native species of Eritrea are at high risk of getting affected by P. juliflora as it occupies a strong ecological position [54]. The similarly arid and semi-arid environment also enhances the invasion risk of such plant species when compared with moist environments. Allelochemicals responsible for degrading or completely inhibiting the growth of seeds and seedlings are always available under the canopy of the tree. They accumulate in the soil and make it hard for other seeds to germinate. Successfully germinated seeds undergo stress that lowers their healthy growth due to the presence of allelochemicals in the soil [36]. Importantly, the two initial stages of growth; seed germination and seedlings are necessary for any species survival as they add new recruitments to populations in a biotic community. In the present study, we also investigated the consequences of the leaf leachate of P. juliflora on seedling growth of a monocot plant, A. lagopoides and two dicot plants: C. imbricatum and T. qatarensis.
The growth height of A. Lagopoides showed no significant differences at any treatment level as compared to the control group (Figure 7A,C). Looking at the concentrations of 4, 6, and 8 mg mL−1, the plant height is more at 6 mg mL−1 as compared to 4 and 8 mg mL−1. It indicates that there is heterogeneity among the treatments; therefore, in this case, plant height cannot be used as an accurate estimate of the allelopathic effects of P. juliflora. On the other hand, the dry matter biomass showed a significant decline in the healthy growth of the seedlings with increasing extract concentration throughout one-month post-treatment (Figure 7B); hence, biomass is a more reliable approximation of the allelopathic effect of P. juliflora. Different research on A. lagopoides provided evidence that this perennial grass can withstand different environmental stresses that appear to be present for a longer time [55]. Therefore, the growth of seedlings to a greater height in the presence of extract accumulated in the soil might be a physiological response to the stress through the production of long stolons, which allow the plant to spread in an area where the concentration of allelochemicals is less. Despite the ability of A. lagopoides to overcome allelochemical stresses by increasing plant height, the growth of seedlings declined due to higher concentrations of extract which may indicate negative allelopathic stress on the growth of A. lagopoides. Supporting our findings, a report states that allelochemicals usually affect grasses more than perennials. The publication further reported a significant inhibition in the germination potential of grasses, Cenchrus ciliaris and Enteropogon rupestris, at a concentration of 6 mg mL−1 [49].
Inhibition and degradation of seedlings growth of C. imbricatum were observed at the concentrated extract of 8 mg mL−1. The variations in growth were evident in both height and dry matter biomass (Figure 8A,B), which reduces the significance of results and may explain why the effect was only using the highest concentration. Compared with A. lagopoides and C. imbricatum, the consequence of leaf leachate on seedling growth of T. qatarensis was severe. Both parameters of growth significantly declined, starting from a concentration of 2 mg mL−1 (Figure 9A,B). Seedlings of C. imbricatum and T. qatarensis are shown in Figure 8C and Figure 9C, respectively. It has been reported that the leaf extract and leachates of P. juliflora have inhibitory effects as compared to its live root extracts [56]. Similarly, another research reports the negative effects of P. juliflora leaf leachate on the root growth of crops [57]. Comparatively, the allelopathic effect of soil leachate of P. juliflora, however, is highly dependent on the canopy size of the plant, age of the plant, microbial community, and characteristics of the soil. It is reported that the allelochemical L-tryptophan degrades quickly in the soil; hence, it is less effective [58]. Similarly, another research confirms the degradation process of allelochemicals in the soil and reports that these inhibitors of plant growth can stay active in the soil for only up to two months [56]. Here, it is important to mention that since the accumulation of the allelochemicals in the soil is a continuous process, it cannot be concluded which type of leachate, either soil or leaf, would be more effective in terms of its allelopathic impact. It is mentioned that the leaf leachate of P. juliflora showed neutral to negative effects on the growth of roots of certain plants [59]. Halophytes, being of severe socio-economic importance, might face severe threat by the presence of invasive P. juliflora. Similar halophytes present in the Farasan Islands in the Arabian Peninsula of Saudi Arabia are threatened by the presence of invasive P. juliflora and its allelochemicals [60].
P. juliflora not only poses a threat to other plants but also has beneficial uses. Upon screening of phytochemicals, similar compounds (alkaloids, flavonoids, coumarins, and anthraquinone glycoside) were reported to have antifungal and antimicrobial activities [61]. Certain laboratory experiments have proved the worth of these allelochemicals by inhibiting certain pathogens like Escherichia coli, Micrococcus luteus, and Pseudomonas aeruginosa [62]. In our laboratory, we used the water-soluble leaf ethanolic extracts of P. juliflora (PJ-WS-LE) against post-harvest pathogens of mango fruits and inhibited the growth of Alternaria alternata, Colletotrichum gloeosporioides, Botrytis cinerea, Bacillus subtilis, Staphylococcus aureus, and Candida albicans [63,64,65]. Similarly, we used PJ-WS-LE as a potential coating agent for preserving cucumber and strawberries and extending their shelf life [66].
Taking into consideration the vast applications of allelopathic plants, they can also be used as biological herbicides instead of synthetic ones. Allelochemicals being used as bioherbicides is an environment-friendly and sustainable approach as they have no residual toxic effects and are biodegradable, leading to low environmental pollution [67,68]. In the near future, we will explore the allelopathic nature of P. juliflora in terms of weed management in agriculture with the hope of utilizing the allelochemicals to protect the crops.
5. Conclusions
This report concludes that the invasive and allelopathic P. juliflora affects the Qatari local plant species based on laboratory experiments. The capability of P. juliflora to inhibit the germination potential of test species and its negative effects on the health of seedlings at the lowest concentration indicates a threat to the existing Qatari native plant species in a region where P. juliflora is inhabited. Keeping in mind the invasive and allelopathic nature of P. juliflora, it is important to protect the already existing Qatari flora and to take precautionary steps for the future. This research could be used as a baseline for further studies where higher concentrations of aqueous extract of P. juliflora could be used to test the germination potential of plant species. It is also important to study the allelopathic effects along with other factors like soil biota or climatic variations to have an in-depth knowledge of the topic. Furthermore, P. juliflora, with its allelopathic nature, could also be investigated and utilized in the future for the purpose of crop protection in agrosystems as a potential natural herbicide.
Author Contributions
Conceptualization, S.B. and M.H.A.-D.; Methodology, S.B., A.B. and M.H.A.-D.; Validation, M.A.A.-G. and M.H.A.-D.; Formal analysis, S.B., A.B., M.A.A.-G. and M.H.A.-D.; Investigation, S.B.; Data curation, S.B. and A.B.; Writing—original draft, S.B.; Writing—review & editing, M.A.A.-G. and M.H.A.-D.; Supervision, M.H.A.-D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Acknowledgments
The research was made possible by Qatar University Graduate Assistantship Program. Open Access funding provided by the Qatar National Library.
Conflicts of Interest
The authors declare that they have no known competing financial or non-financial interests.
References
- Ehlers, B.K.; Charpentier, A.; Grøndahl, E. An allelopathic plant facilitates species richness in the Mediterranean garrigue. J. Ecol. 2014, 102, 176–185. [Google Scholar] [CrossRef]
- Inderjit; Wardle, D.A.; Karban, R.; Callaway, R.M. The ecosystem and evolutionary contexts of allelopathy. Trends Ecol. Evol. 2011, 26, 655–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, J.J.; Rathinasabapathi, B.; Chase, C.A. Allelopathy: How Plants Suppress Other Plants. EDIS 2013, 2013. [Google Scholar] [CrossRef]
- Balcha, L.D. Prosopis juliflora Distribution, Impacts, and Control Methods Available in Ethiopia. Int. J. Nat. Resour. Ecol. Manag. 2022, 7, 132–144. [Google Scholar]
- Howari, F.M.; Sharma, M.; Nazzal, Y.; El-Keblawy, A.; Mir, S.; Xavier, C.M.; Salem, I.B.; Al-Taani, A.A.; Alaydaroos, F. Changes in the Invasion Rate of Prosopis juliflora and Its Impact on Depletion of Groundwater in the Northern Part of the United Arab Emirates. Plants 2022, 11, 682. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Mathur, M. Impact of invasion by Prosopis juliflora on plant communities in arid grazing lands Network Project on Guggul View project Assessing adaptability and utilization potential of Opuntia ficus-indica in arid and semi-arid regions of India V. Trop. Ecol. 2014, 55, 33–46. [Google Scholar]
- Pasiecznik, N.M.; Felker, P.; Harris, P.J.C.; Harsh, L.N.; Cruz, G.; Tewari, J.C.; Cadoret, K.; Maldonado, L.J. The Prosopis juliflora-Prosopis pallida Complex: A Monograph; HDRA: Coventry UK, 2002; p. 172. [Google Scholar]
- Shiferaw, W.; Demissew, S.; Bekele, T.; Aynekulu, E.; Pitroff, W. Invasion of Prosopis juliflora and its effects on soil physicochemical properties in Afar region, Northeast Ethiopia. Int. Soil Water Conserv. Res. 2021, 9, 631–638. [Google Scholar] [CrossRef]
- Mehari, Z.H. The invasion of Prosopis juliflora and Afar pastoral livelihoods in the Middle Awash area of Ethiopia. Ecol. Process. 2015, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- De Brito Damasceno, G.A.; Souto, A.L.; da Silva, I.B.; Roque, A.d.A.; Ferrari, M.; Giordani, R.B. Prosopis juliflora: Phytochemical, Toxicological, and Allelochemicals. Ref. Ser. Phytochem. 2020, 521–541. [Google Scholar] [CrossRef]
- Kumar Mehar, S. Assessment of effect of Prosopis juliflora litter extract on seed germination and growth of rice. Food Sci. Qual Manag. 2011, 2. [Google Scholar]
- Almaraz-Abarca, N.; da Graça Campos, M.; Avila-Reyes, J.A.; Naranjo-Jimenez, N.; Corral, J.H.; Gonzalez-Valdez, L.S. Antioxidant activity of polyphenolic extract of monofloral honeybee-collected pollen from mesquite (Prosopis juliflora, Leguminosae). J. Food Compos. Anal. 2007, 20, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Wassel, G.; Rizk, A.; Abdel-Bary, E. Phytochemical investigation of Prosopis juliflora DCI Flavonoids and free sugars. Qual. Plant. Et Mater. Veg. 1972, 22, 119–121. [Google Scholar] [CrossRef]
- Nee’Shukla, R.V.; Misra, K. Two flavonoid glycosides from the bark of Prosopis juliflora. Phytochemistry 1981, 20, 339–340. [Google Scholar] [CrossRef]
- Ibrahim, M.; Nadir, M.; Ali, A.; Ahmad, V.U.; Rasheed, M. Phytochemical analyses of Prosopis juliflora Swartz DC. Pak. J. Bot 2013, 45, 2101–2104. [Google Scholar]
- Sharmila, S.; Jeyanthi, R.; Saduzzaman, M. Biodegradation of tannery effluent using Prosopis juliflora. Int. J. ChemTech Res. 2013, 5, 2186–2192. [Google Scholar]
- Tesoriere, L.; Butera, D.; Allegra, M.; Fazzari, M.; Livrea, M.A. Distribution of betalain pigments in red blood cells after consumption of cactus pear fruits and increased resistance of the cells to ex vivo induced oxidative hemolysis in humans. J. Agric. Food Chem. 2005, 53, 1266–1270. [Google Scholar] [CrossRef]
- Bhatt, S.; Chovatiya, S.; Shah, A. Evaluation of raw and hydrothermically processed Prosopis juliflora seed meal as supplementary feed for the growth of Labeo rohita fingerlings. Aquac. Nutr. 2011, 17, e164–e173. [Google Scholar] [CrossRef]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef]
- Nakano, H.; Fujii, Y.; Suzuki, T.; Yamada, K.; Kosemura, S.; Yamamura, S.; Suzuki, T.; Hasegawa, K. A growth-inhibitory substance exuded from freeze-dried mesquite (Prosopis juliflora (Sw.) DC.) leaves. Plant Growth Regul. 2001, 33, 165–168. [Google Scholar] [CrossRef]
- Nakano, H.; Fujii, Y.; Yamada, K.; Kosemura, S.; Yamamura, S.; Hasegawa, K.; Suzuki, T. Isolation and identification of plant growth inhibitors as candidate (s) for allelopathic substance (s), from aqueous leachate from mesquite (Prosopis juliflora (Sw.) DC.) leaves. Plant Growth Regul. 2002, 37, 113–117. [Google Scholar] [CrossRef]
- Nakano, H.; Nakajima, E.; Fujii, Y.; Yamada, K.; Shigemori, H.; Hasegawa, K. Leaching of the allelopathic substance,-tryptophan from the foliage of mesquite (Prosopis juliflora (Sw.) DC.) plants by water spraying. Plant Growth Regul. 2003, 40, 49–52. [Google Scholar] [CrossRef]
- Inderjit; Seastedt, T.R.; Callaway, R.M.; Pollock, J.L.; Kaur, J. Allelopathy and plant invasions: Traditional, congeneric, and bio-geographical approaches. Biol. Invasions 2008, 10, 875–890. [Google Scholar] [CrossRef]
- Pardo-Muras, M.; Puig, C.G.; Pedrol, N. Complex Synergistic Interactions among Volatile and Phenolic Compounds Underlie the Effectiveness of Allelopathic Residues Added to the Soil for Weed Control. Plants 2022, 11, 1114. [Google Scholar] [CrossRef] [PubMed]
- Ghazanfar, S.A. Invasive Prosopis in sultanate of Oman. Aliens 1996, 3, 10. [Google Scholar]
- Western, A.R. The Flora of the United Arab Emirates: An Introduction; United Arab Emirates University: Al-Ain, United Arab Emirates, 1989. [Google Scholar]
- Sergeev, A.V. Prosopis juliflora—Flora of Qatar. 2008. Available online: https://www.floraofqatar.com/prosopis_juliflora.htm (accessed on 5 October 2022).
- Siddiqui, S.; Bhardwar, S.; Khan, S.K.; Meghvanshi, M.K. Allelopathic effect of different conentration of water extract of Prosopis Juliflora leaf on seed germination and radicle length of wheat. J. Sci. Res. 2009, 4, 81–84. [Google Scholar]
- Noor, M.; Salam, U.; Khan; Ajmal, M. Allelopathic effects of Prosopis juliflora Swartz. J. Arid. Environ. 1995, 31, 83–90. [Google Scholar] [CrossRef]
- Jayasinghe, A.G.C.S.; Perera, G.A.D. Allelopathic Effect of Prosopis juliflora (Mesquite) on Seed Germination of Native Coastal Dry Forest Species. In Proceedings of the Peradeniya University Research Sessions, Peradeniya, Sri Lanka, 24 November 2011; Volume 16, p. 180. [Google Scholar]
- Saadaoui, E.; Martin, J.J.; Ghazel, N.; Ben Romdhane, C.; Massoudi, N.; Cervantes, E. Allelopathic Effects of Aqueous Extracts of Ricinus communis L. on the Germination of Six Cultivated Species. Int. J. Plant Soil Sci. 2015, 7, 220–227. [Google Scholar] [CrossRef]
- Khan, D.; Shahid Shaukat, S. Phytotoxic effects of P. juliflora Swartz. DC. against some of its field associates and the cultivated species. Int. J. Biol Biotech 2006, 3, 353–366. [Google Scholar]
- El-Keblawy, A.; Al-Rawai, A. Impacts of the invasive exotic Prosopis juliflora (Sw.) D.C. on the native flora and soils of the UAE. Plant Ecol. 2007, 190, 23–35. [Google Scholar] [CrossRef]
- Iffat Alam, S.; Hammoda, H.; Khan, F.; Al Enazi, R.; Goktepe, I. Electrical Conductivity, pH, Organic Matter and Texture of Selected Soils Around the Qatar University Campus. Res. Agric. Livest. Fish. 2020, 7, 403–409. [Google Scholar] [CrossRef]
- Amin, S.; Mansur, I.; Pamoengkas, P.; Yusmur, A. Germination Test of Two Different Types of Seed to Accelerate Reclamation and Rehabilitation of Ex-limestone Mining Land. Proc. IOP Conf. Ser. Earth Environ. Sci. 2021, 959, 012050. [Google Scholar] [CrossRef]
- Omer, H.H.M.; Mohammed, I.S. Allelopathic effects of mesquite (Prosopis juliflora) aqueous extracts on seeds germination and seedlings growth of alfalfa, sesame and sorghum. Cell Biol. Development. 2017, 1, 51–54. [Google Scholar] [CrossRef]
- Ramandi, A.; Javan, I.Y.; Tazehabadi, F.M.; Asl, G.I.; Khosravanian, R.; Ebrahimzadeh, M.H. Improvement in seed surface sterilization and in vitro seed germination of ornamental and medicinal plant-Catharanthus roseus (L.). Chiang Mai J. Sci. 2019, 46, 1107–1112. [Google Scholar]
- Alhaddad, F.A.; Abu-Dieyeh, M.H.; ElAzazi, E.S.M.; Ahmed, T.A. Salt tolerance of selected halophytes at the two initial growth stages for future management options. Sci. Rep. 2021, 11, 10194. [Google Scholar] [CrossRef]
- Statwick, J.M. Germination pretreatments to break hard-seed dormancy in Astragalus cicer L. (Fabaceae). PeerJ 2016, 4, e2621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najib, T.; Heydari, M.M.; Meda, V. Combination of germination and innovative microwave-assisted infrared drying of lentils: Effect of physicochemical properties of different varieties on water uptake, germination, and drying kinetics. Appl. Food Res. 2022, 2, 100040. [Google Scholar] [CrossRef]
- Setia, R.; Dai, Z.; Nickerson, M.T.; Sopiwnyk, E.; Malcolmson, L.; Ai, Y. Impacts of short-term germination on the chemical compositions, technological characteristics and nutritional quality of yellow pea and faba bean flours. Food Res. Int. 2019, 122, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Wymore, A.S.; Compson, Z.G.; McDowell, W.H.; Potter, J.D.; Hungate, B.A.; Whitham, T.G.; Marks, J.C. Leaf-litter leachate is distinct in optical properties and bioavailability to stream heterotrophs. Freshw. Sci. 2015, 34, 857–866. [Google Scholar] [CrossRef] [Green Version]
- Strauss, E.A.; Lamberti, G.A. Effect of dissolved organic carbon quality on microbial decomposition and nitrification rates in stream sediments. Freshw. Biol. 2002, 47, 65–74. [Google Scholar] [CrossRef]
- Gianinetti, A. Basic features of the analysis of germination data with generalized linear mixed models. Data 2020, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Bertsouklis, K.; Vlachou, G.; Trigka, M.; Papafotiou, M. In Vitro Studies on Seed Germination of the Mediterranean Species Anthyllis barba-jovis to Facilitate Its Introduction into the Floriculture Industry. Horticulturae 2022, 8, 889. [Google Scholar] [CrossRef]
- Haregeweyn, N.; Tsunekawa, A.; Tsubo, M.; Meshesha, D.; Melkie, A. Analysis of the invasion rate, impacts and control measures of Prosopis juliflora: A case study of Amibara District, Eastern Ethiopia. Environ. Monit. Assess. 2013, 185, 7527–7542. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.M.; Soliman, W.S.; Mahmoud, M.; Youssif, M.F.; Mansour, A.; Taher, E.; Hassan, I.N.; Mansour, H.; Abdelkareem, M. Predicting the Spatial Spread of Invasive Prosopis juliflora (SW.) D.C along Environmental Gradients in Gabel Elba National Park, Egypt. Int. J. Sci. Eng. Res. 2016, 7, 596–599. [Google Scholar] [CrossRef]
- Al-Humaid, A.I.; Warrag, M.O.A. Allelopathic effects of mesquite (Prosopis juliflora) foliage on seed germination and seedling growth of bermudagrass (Cynodon dactylon). J. Arid Environ. 1998, 38, 237–243. [Google Scholar] [CrossRef]
- Getachew, S.; Demissew, S.; Woldemariam, T.; Dana, E. Allelopathic effects of the invasive Prosopis juliflora (Sw.) DC. on selected native plant species in Middle Awash, Southern Afar Rift of Ethiopia. Manag. Biol. Invasions 2012, 3, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Mei, L.; Tang, J. Allelopathic effects of invasive Solidago canadensis on germination and root growth of native Chinese plants. In Proceedings of the 4th World Congress on Allelopathy, Wagga Wagga, NSW, Australia, 21–26 August 2005; Charles Sturt University: Wagga Wagga, NSW, Australia, 2005; pp. 43–49. [Google Scholar]
- Saxena, S.K. Banni grassland and halophytes. In Halophytes as a Resource for Livestock and for Rehabilitation of Degraded Lands; Springer: Dordrecht, The Netherlands, 1994; Volume 32, pp. 217–222. [Google Scholar] [CrossRef]
- Ungar, I.A. Are Biotic Factors Significant in Influencing the Distribution of Halophytes in Saline Habitats? Bot. Review. 1998, 64, 176–199. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Autotoxicity: Concept, Organisms, and Ecological Significance. Crit. Rev. Plant Sci. 2010, 18, 757–772. [Google Scholar] [CrossRef]
- Bokrezion, H. The Ecological and Socio Economic Role of Prosopis juliflora in Eritrea an Analytical Assessment within the Context of Rural Development in the Horn of Africa; Johannes Gutenberg-Universität Mainz: Mainz, Germany, 2008. [Google Scholar]
- Mohsenzadeh, S.; Malboobi, M.A.; Razavi, K.; Farrahi-Aschtiani, S. Physiological and molecular responses of Aeluropus lagopoides (Poaceae) to water deficit. Environ. Exp. Bot. 2006, 56, 314–322. [Google Scholar] [CrossRef]
- Goel, U.; Saxena, D.B.; Kumar, B. Comparative study of allelopathy as exhibited byProsopis Juliflora swartz and Prosopis cineraria (L) druce. J. Chem. Ecol. 1989, 15, 591–600. [Google Scholar] [CrossRef]
- Kaur, R.; Gonzáles, W.L.; Llambi, L.D.; Soriano, P.J.; Callaway, R.M.; Rout, M.E.; Gallaher, T.J.; Inderjit. Community Impacts of Prosopis juliflora Invasion: Biogeographic and Congeneric Comparisons. PLoS ONE 2012, 7, e44966. [Google Scholar] [CrossRef]
- Kaur, R.; Callaway, R.M.; Inderjit. Soils and the conditional allelopathic effects of a tropical invader. Soil Biol. Biochem. 2014, 78, 316–325. [Google Scholar] [CrossRef]
- Mustafa, H.F.; Bushra, R.A. Effect of Leaf Aqueous Extract of Prosopis juliflora (Swartz) DC and Acacia raddiana (Brenan) on Early Growth of Sorghum sudanese (Piper Stapf). IOSR J. Agric. Vet. Sci. 2018, 11, 26–31. [Google Scholar]
- Hall, M.; Llewellyn, O.A.; Miller, A.G.; Al-Abbasi, T.M.; Al-Wetaid, A.H.; Al-Harbi, R.J.; Al-Shammari, K.F. Important plant areas in the arabian peninsula: 2. farasan archipelago. Edinb. J. Bot. 2010, 67, 189–208. [Google Scholar] [CrossRef]
- Badri, A.M.; Garbi, M.I.; Gmaraldeen, S.M.; Magzoub, A.A.; Ibrahim, I.T.; Saleh, M.S.; Kabbashi, A.S.; Mohamed, S.G. Antioxidant activity and phytochemical screening of Prosopis juliflora leaves extract. Adv. Med. Plant Res 2017, 5, 37–40. [Google Scholar] [CrossRef] [Green Version]
- Sathiya, M.; Muthuchelian, K. Investigation of Phytochemical Profile and Antibacterial Potential of Ethanolic Leaf Extract of Prosopis juliflora DC. Ethnobot. Leafl. 2008, 2008, 167. [Google Scholar]
- Saleh, I.; Ahmed, T.; Halboosi, R.; Abu-Dieyeh, M. Genetic diversity of Prosopis juliflora in the state of Qatar and its valuable use against postharvest pathogen of mango fruits. Sci. Rep. 2022, 12, 1–13. [Google Scholar] [CrossRef]
- Saleh, I.; Abu-Dieyeh, M.H. Novel Prosopis juliflora leaf ethanolic extract as natural antimicrobial agent against food spoiling microorganisms. Sci. Rep. 2021, 11, 1–17. [Google Scholar] [CrossRef]
- Saleh, I.; Abu-Dieyeh, M. Novel Prosopis juliflora leaf ethanolic extract coating for extending postharvest shelf-life of strawberries. Food Control. 2022, 133, 108641. [Google Scholar] [CrossRef]
- Saleh; Abu-Dieyeh. Evaluation of novel Prosopis juliflora water soluble leaf ethanolic extract as preservation coating material of cucumber. J. Food Process. Preserv. 2022, 46, e16352. [Google Scholar] [CrossRef]
- Palanivel, H.; Tilaye, G.; Belliathan, S.K.; Benor, S.; Abera, S.; Kamaraj, M. Allelochemicals as Natural Herbicides for Sustainable Agriculture to Promote a Cleaner Environment. In Strategies and Tools for Pollutant Mitigation: Avenues to a Cleaner Environment; Aravind, J., Kamaraj, M., Prashanthi Devi, M., Rajakumar, S., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 93–116. [Google Scholar]
- Ullah, R.; Aslam, Z.; Attia, H.; Sultan, K.; Alamer, K.H.; Mansha, M.Z.; Althobaiti, A.T.; Al Kashgry, N.A.T.; Algethami, B.; Zaman, Q.U. Sorghum Allelopathy: Alternative Weed Management Strategy and Its Impact on Mung Bean Productivity and Soil Rhizosphere Properties. Life 2022, 12, 1359. [Google Scholar] [CrossRef]
Figure 1.
The number of seedlings of inhabitants Ziziphus spp., Tamarix sp., A. tortilis, and P. juliflora observed in the field survey.
Figure 1.
The number of seedlings of inhabitants Ziziphus spp., Tamarix sp., A. tortilis, and P. juliflora observed in the field survey.
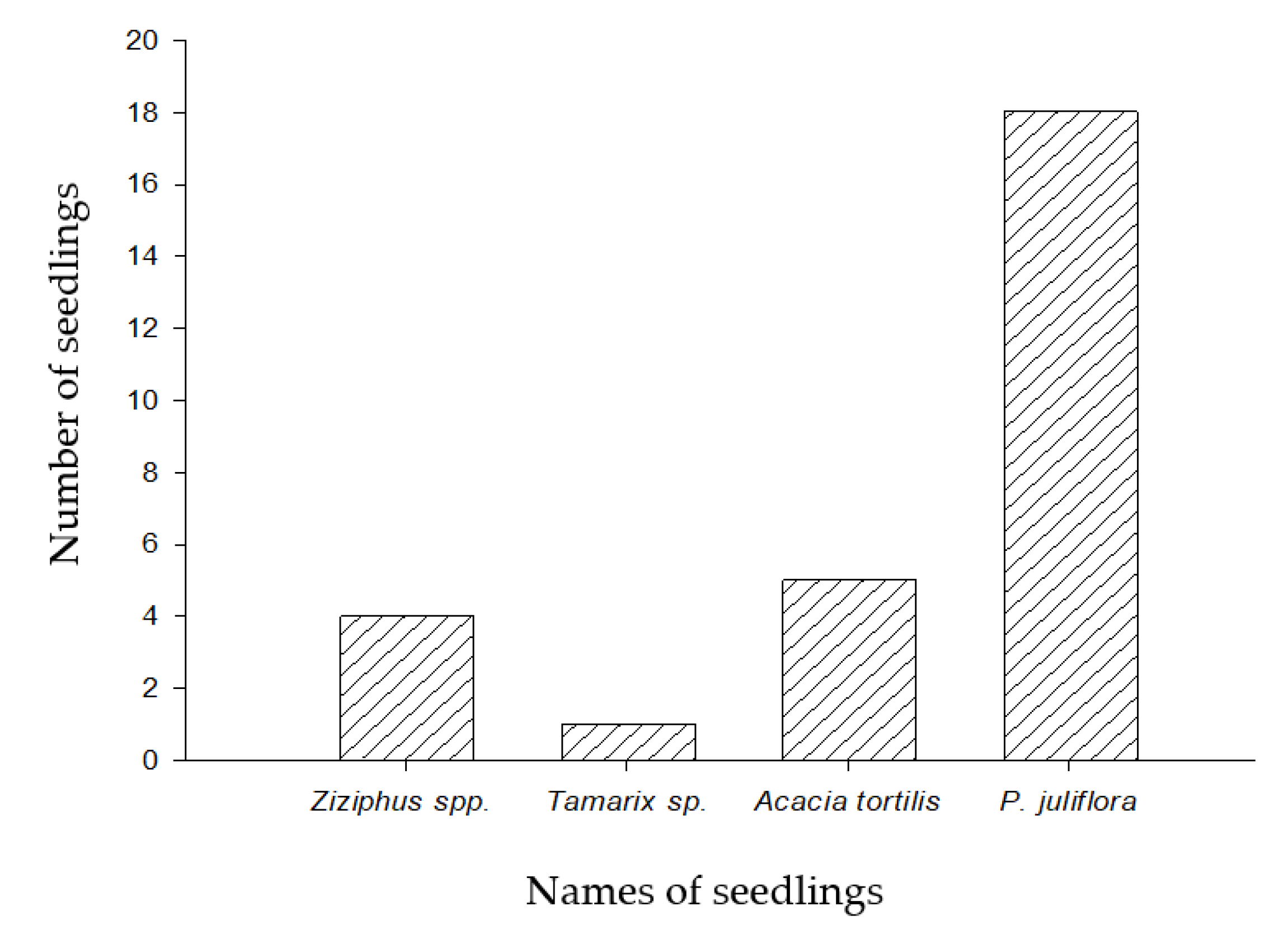
Figure 2.
Effect of aqueous extract of P. juliflora on seed germination of A. tortilis (A), P. cinereria (B), S. aegyptiaca (C), and H. perfoliata (D). According to Tukey’s test, mean values having the same letters show no significant difference at p ≤ 0.05 (when comparing means of the same species). Error bars represent the standard error of the means (n = 5).
Figure 2.
Effect of aqueous extract of P. juliflora on seed germination of A. tortilis (A), P. cinereria (B), S. aegyptiaca (C), and H. perfoliata (D). According to Tukey’s test, mean values having the same letters show no significant difference at p ≤ 0.05 (when comparing means of the same species). Error bars represent the standard error of the means (n = 5).
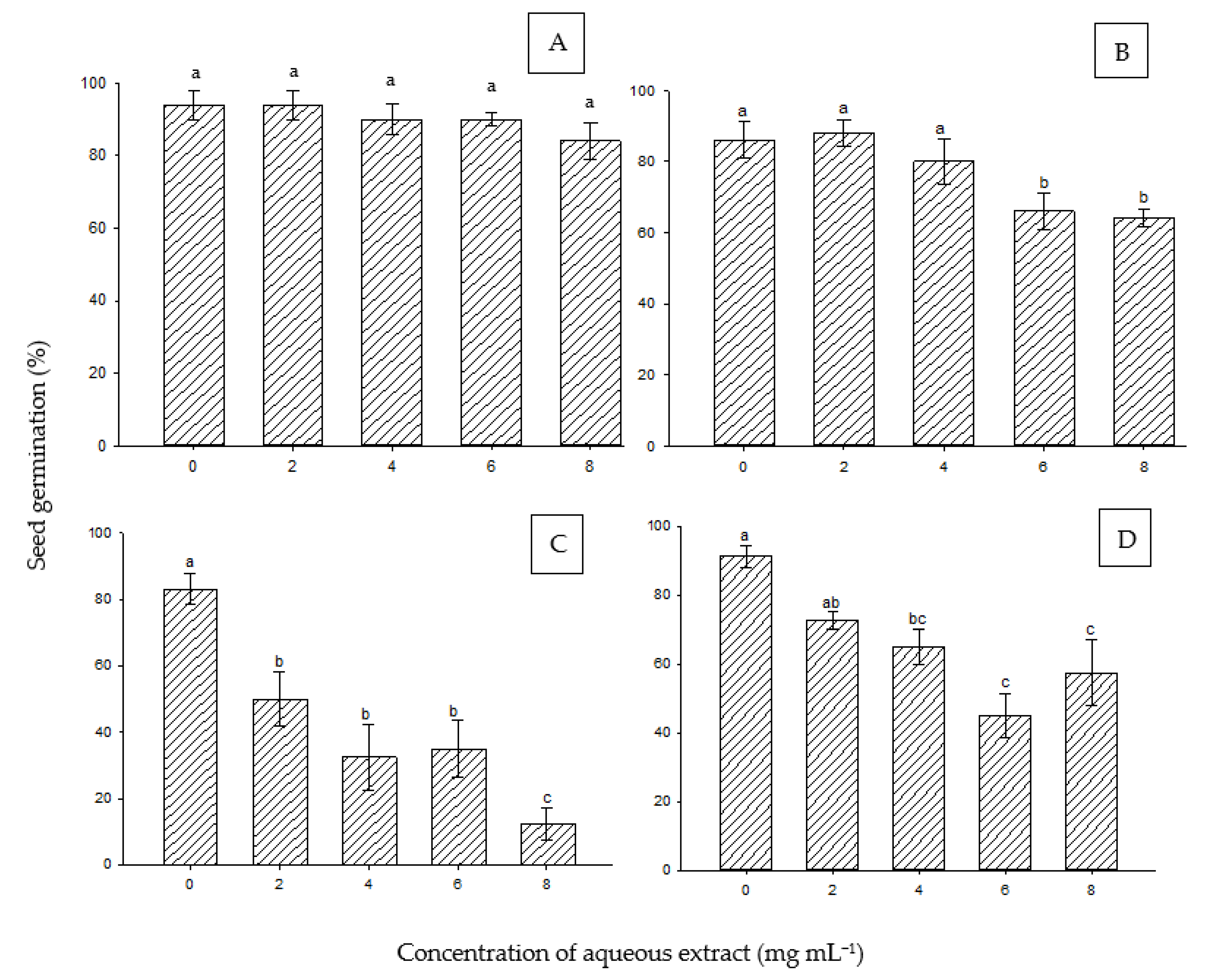
Figure 3.
Effect of aqueous extract of P. juliflora on radicle length of A. tortilis (A), P. cinereria (B), and S. aegyptiaca (C). According to Tukey’s test, mean values having the same letters show no significant difference at p ≤ 0.05. Error bars represent the standard error of the means (n = 5).
Figure 3.
Effect of aqueous extract of P. juliflora on radicle length of A. tortilis (A), P. cinereria (B), and S. aegyptiaca (C). According to Tukey’s test, mean values having the same letters show no significant difference at p ≤ 0.05. Error bars represent the standard error of the means (n = 5).
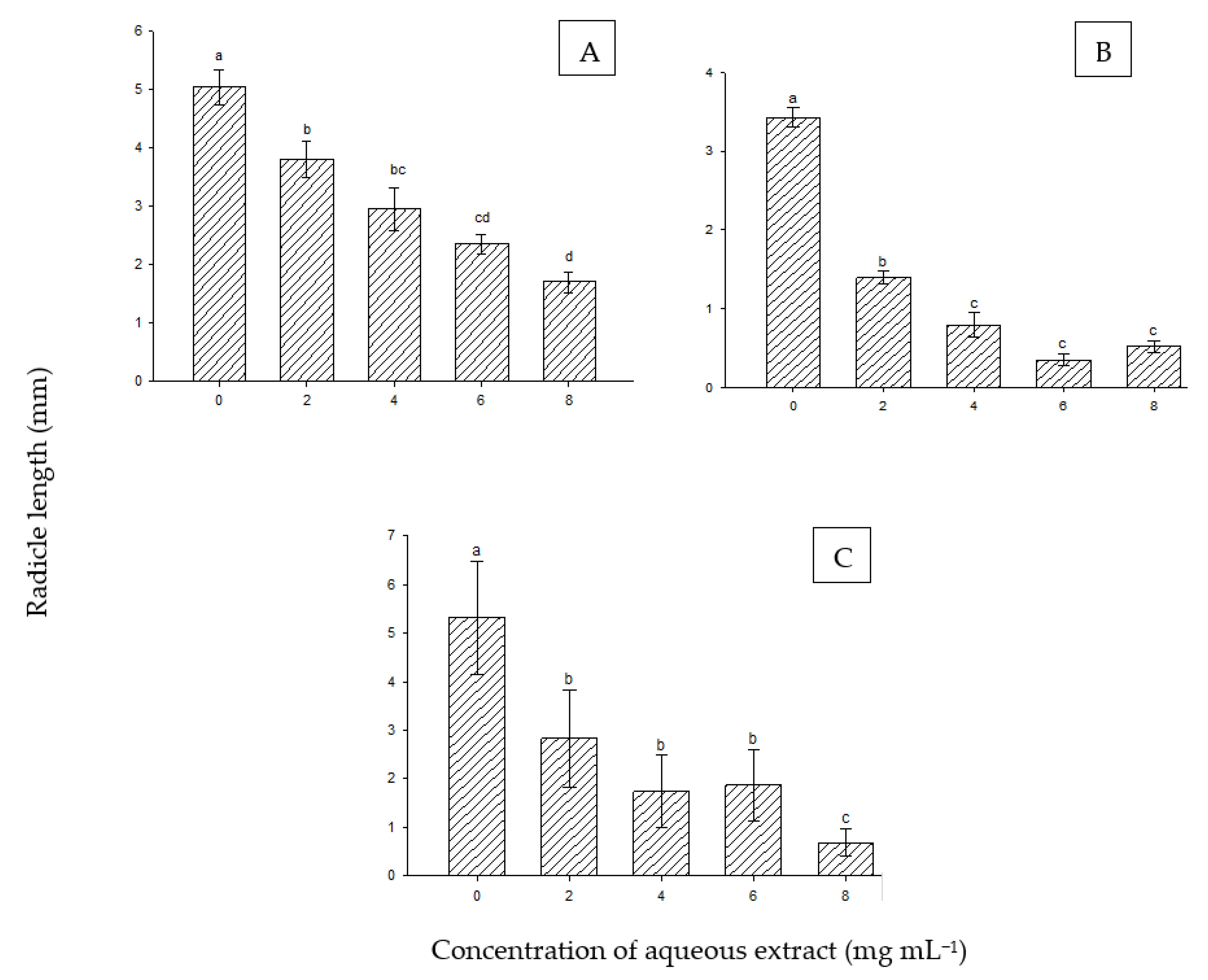
Figure 4.
Effect of aqueous extract of P. juliflora on radicle length of P. cineraria (A) and S. aegyptiaca (B) after two weeks of treatment.
Figure 4.
Effect of aqueous extract of P. juliflora on radicle length of P. cineraria (A) and S. aegyptiaca (B) after two weeks of treatment.

Figure 5.
Effect of aqueous extract of P. juliflora on seed germination (A) and radicle length (B) of P. juliflora. According to Tukey’s test, mean values having the same letters show no significant difference at p ≤ 0.05. Error bars represent the standard error of the means (n = 5).
Figure 5.
Effect of aqueous extract of P. juliflora on seed germination (A) and radicle length (B) of P. juliflora. According to Tukey’s test, mean values having the same letters show no significant difference at p ≤ 0.05. Error bars represent the standard error of the means (n = 5).
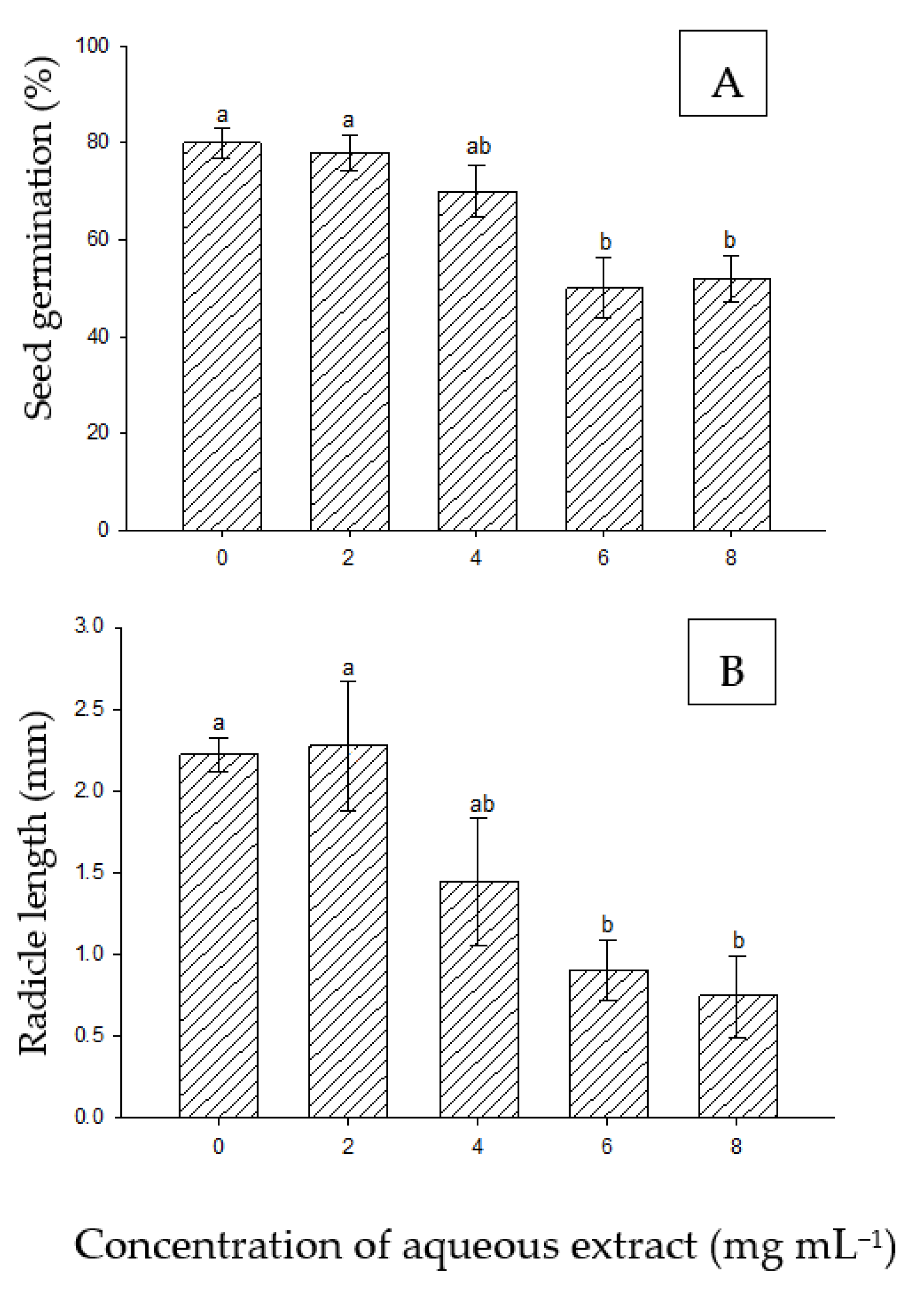
Figure 6.
Effect of aqueous extract of P. juliflora on radicle length of their seeds with a clear effect of auto allelopathy on seed germination).
Figure 6.
Effect of aqueous extract of P. juliflora on radicle length of their seeds with a clear effect of auto allelopathy on seed germination).

Figure 7.
Effect of leaf leachate of P. juliflora on seedling growth of A. lagopoides (A) Effect on plant height. (B) Effect on dry matter biomass. (C) Photos of seedlings of A. lagopoides at harvesting time, one-month post-treatment. According to Tukey’s test, mean values having same letters show no significant difference at p ≤ 0.05. Error bars represent the standard error of the means (n = 5).
Figure 7.
Effect of leaf leachate of P. juliflora on seedling growth of A. lagopoides (A) Effect on plant height. (B) Effect on dry matter biomass. (C) Photos of seedlings of A. lagopoides at harvesting time, one-month post-treatment. According to Tukey’s test, mean values having same letters show no significant difference at p ≤ 0.05. Error bars represent the standard error of the means (n = 5).


Figure 8.
Effect of leaf leachate of P. juliflora on seedling growth of C. imbricatum (A) Effect on plant height, (B) Effect on dry matter biomass. (C) Photos of seedlings of C. imbricatum at harvesting time, one-month post-treatment. According to Tukey’s test, mean values having the same letters show no significant difference at p ≤ 0.05. Error bars represent the standard error of the means (n = 10).
Figure 8.
Effect of leaf leachate of P. juliflora on seedling growth of C. imbricatum (A) Effect on plant height, (B) Effect on dry matter biomass. (C) Photos of seedlings of C. imbricatum at harvesting time, one-month post-treatment. According to Tukey’s test, mean values having the same letters show no significant difference at p ≤ 0.05. Error bars represent the standard error of the means (n = 10).
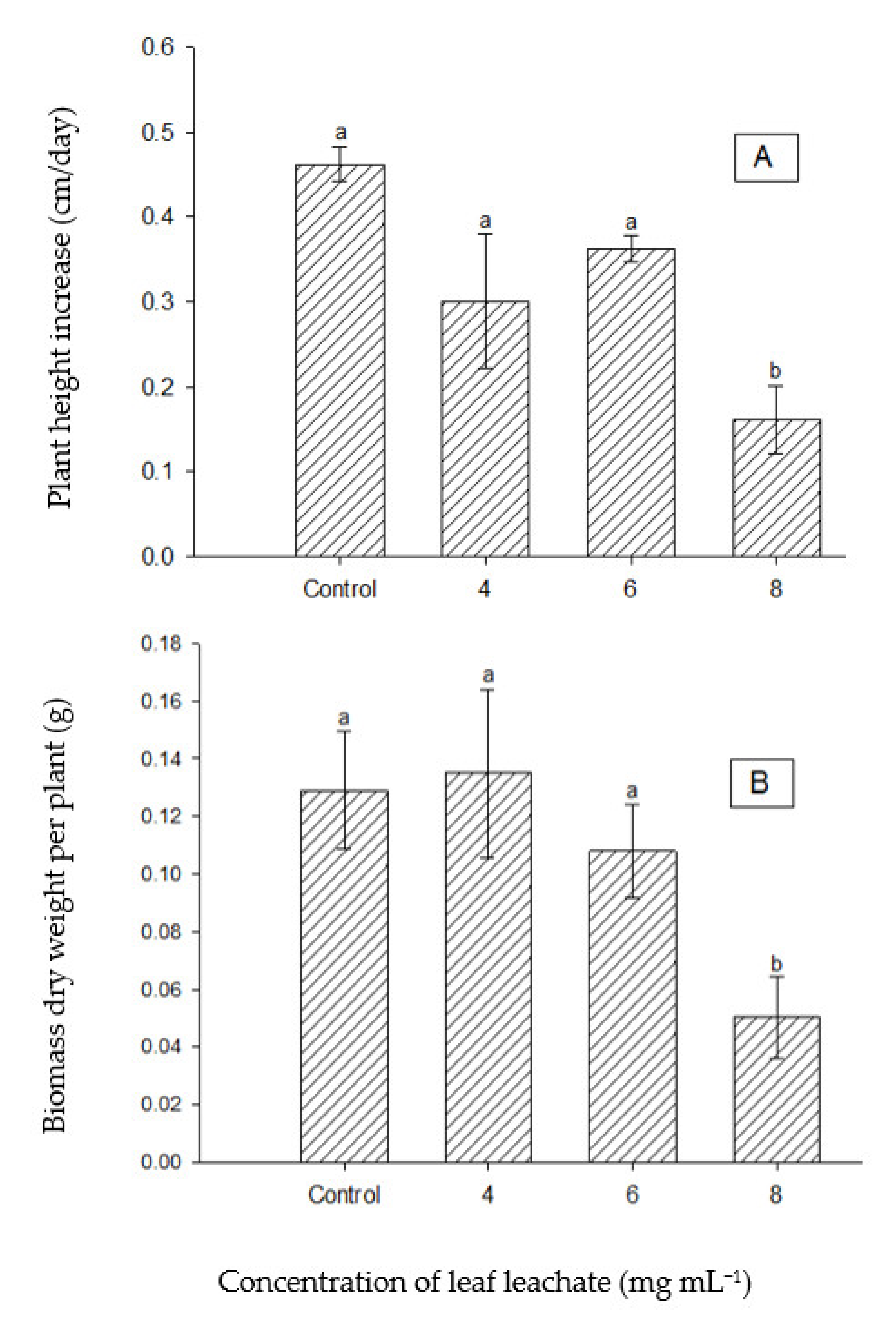

Figure 9.
Effect of leaf leachate of P. juliflora on seedling growth of T. qatarensis (A) Effect on plant height, (B) Effect on dry matter biomass. (C) Photos of seedlings of T. qatarensis at harvesting time, one-month post-treatment. According to Tukey’s test, mean values having the same letters show no significant difference at p ≤ 0.05. Error bars represent the standard error of the means (n = 10).
Figure 9.
Effect of leaf leachate of P. juliflora on seedling growth of T. qatarensis (A) Effect on plant height, (B) Effect on dry matter biomass. (C) Photos of seedlings of T. qatarensis at harvesting time, one-month post-treatment. According to Tukey’s test, mean values having the same letters show no significant difference at p ≤ 0.05. Error bars represent the standard error of the means (n = 10).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bibi, S.; Bibi, A.; Al-Ghouti, M.A.; Abu-Dieyeh, M.H. Allelopathic Effects of the Invasive Prosopis juliflora (Sw.) DC. on Native Plants: Perspectives toward Agrosystems. Agronomy 2023, 13, 590. https://doi.org/10.3390/agronomy13020590
AMA Style
Bibi S, Bibi A, Al-Ghouti MA, Abu-Dieyeh MH. Allelopathic Effects of the Invasive Prosopis juliflora (Sw.) DC. on Native Plants: Perspectives toward Agrosystems. Agronomy. 2023; 13(2):590. https://doi.org/10.3390/agronomy13020590
Chicago/Turabian StyleBibi, Shazia, Amina Bibi, Mohammad A. Al-Ghouti, and Mohammed H. Abu-Dieyeh. 2023. "Allelopathic Effects of the Invasive Prosopis juliflora (Sw.) DC. on Native Plants: Perspectives toward Agrosystems" Agronomy 13, no. 2: 590. https://doi.org/10.3390/agronomy13020590
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.