

Soil Nematodes as a Means of Conservation of Soil Predatory Mites for Biocontrol

1
Ecology Group, Institute of Biology, Humboldt Universität zu Berlin, Philippstraße 13, 10115 Berlin, Germany
2
Newe Ya’ar Research Center, Agricultural Research Organization (ARO), P.O. Box 1021, Ramat Yishay 3009500, Israel
*
Author to whom correspondence should be addressed.
Agronomy 2023, 13(1), 32; https://doi.org/10.3390/agronomy13010032
Submission received: 14 November 2022
/
Revised: 12 December 2022
/
Accepted: 16 December 2022
/
Published: 22 December 2022
(This article belongs to the Special Issue Nematodes: Drivers of Agricultural Ecosystem Performance)
Abstract
:Numerous lab and field studies have reported the potential of soil predatory mites for the biological control of plant-parasitic nematodes and arthropods pests. Most of these studies have utilized biocontrol agents in augmentative releases, essentially controlling the pest with the released predators. While this may be a valid approach, we hypothesize that conservation of soil mite predators with available, suitable, and accessible free-living nematodes as prey, will provide better agricultural ecosystem performance and long-range sustainability. In this manuscript, we review the relevant studies on soil predatory mite–nematode interactions and highlight their potential for conservation biological control of soil-borne pests. Additionally, we emphasize the importance of implementing environmentally sound soil management practices for the sustainability and conservation of functional soil food webs.
1. Introduction
Biological pest control by natural enemies is a key ecosystem service for soil and plant health. Soil predatory mites feed on a diverse diet making them excellent candidates as biocontrol agents in conservation biocontrol programs in agriculture. However, the contribution of soil microarthropods in regulating pest populations is poorly known [1].
Within mites, Mesostigmata, and especially the family Phytoseiidae, are already widely used for control of above-ground agricultural pests. The most well-known are specialist species, having one prey (e.g., Phytoseiulus persimilis Athias-Henriot), while some generalists (e.g., Amblyseius swirskii Athias-Henriot) have gained prominence because they can be used for several prey species and also perform well with additional alternative food [2,3,4]. The same cannot be said for below-ground systems. Despite their known diversity and predatory capacity, only a few soil mite species are used in the biological control of pests, even though below-ground pests, in particular plant-parasitic nematodes, are responsible for severe crop damage in agricultural systems [5]. While soil Mesostigmata mites are mostly generalists, many of them have a preference for free-living nematodes [6,7,8].
Here we review the trophic interactions between mites and nematodes in soil, how they affect mite fitness and how we can take advantage of these interactions to control plant pests above- as well as below-ground.
1.1. Trophic Interactions in Soil
Soil organisms and the structure and function of their food webs play a key role in nutrient cycling and litter decomposition, soil processes which are crucial for above-ground primary production [9,10]. Their trophic interactions significantly impact the composition and activity of microbial assemblages, and in turn, plant growth and fitness [11]. The microfauna grazers, protists and nematodes, act as regulators by organizing the community structure and by driving nutrient mineralization of their microbial prey. Moreover, the microfauna releases in mineral forms part of the nutrients captured by grazing [12,13], which for ammonium is estimated at 32–38% of the annual N mineralization in arable soil [14]. Taken together, the multi-trophic interactions in the soil food web are key determinants for plant development and health.
Besides nutrient cycling, another desirable function of a healthy soil is pest suppression. Diverse functions of the soil fauna, such as improving plant resistance and tolerance, are widely acknowledged [15,16,17], while trophic relationships receive less attention [1,18]. This is mainly due to the fact that predator–prey interactions are difficult to predict in highly diverse soil food webs, settled in a cryptic habitat with a mosaic of abiotic and biotic conditions. Moreover, the two major food chains in soil, the herbivore (“green”) and the detritivore (“brown”) chains, are traditionally considered as different entities (Figure 1) [19,20], and predators that cross-link these pathways are seldom investigated. However, organisms with a wide diet range, such as predatory mites, have regulatory impact in both the green and brown compartments, and connect carbon and energy transfer in between.
Microarthropods with their high degree of omnivory and multi-channel feeding across energy sources, are key components of sustainable biological control strategies [18]. In particular, mites function as trophic-level omnivores and form a feeding gradient rather than a discrete trophic group [1]. Generally, biocontrol approaches are designed as top–down control of the prey (pest) by the predator (or parasite). The opposite way, i.e., bottom–up forces along the food chain, leading to increased fitness and improved biocontrol function of a predator, have received very little attention. Only recently, the trophic link between predatory mites and free-living nematodes (FLN) as complementary prey has been shown to improve the efficacy of mites as biocontrol agents [22,23,24]. This highlights that biological pest control by soil fauna predators should be viewed within the framework of such interactivity.
1.2. Nematodes in Soil Food Webs
Nematodes form one of the largest phyla in the animal kingdom, with more than 25,000 species described to date, and an estimated ≈10 million species [25]. They are the most abundant metazoans, e.g., in soils 3.5–5 million individuals per square meter are common [26]. A recent global survey estimated a total nematode biomass of approximately 0.3 gigatonnes are available as resources for higher predators in surface soils across the world [27]. Soil nematodes are either free-living (FLN) or parasites of plants (PPN) and animals (APN) [28]. The FLN taxa consume bacteria, fungi, protozoa, algae, small rotifers, enchytraeids, as well as other nematodes. Additionally, “facultative root and fungal feeders” change resource depending on availability, and thus display considerable resilience to, e.g., crop removal at harvest [29]. Overall, nematodes have important roles as primary, secondary, and tertiary consumers [30] and, moreover, are prey of the micro- and mesofauna, forming an important node and transition in the soil food web.
With functional groups in both food chains, the herbivore and the detritivore, nematodes can cross-link the pathways of energy and nutrients (Figure 1). In the herbivore food chain, more than 4100 PPN species are responsible for an estimated economic loss to the agricultural sector of nearly $125 billion annually [31]. The detrital food chain comprises the bacterial and fungal grazers and the related decomposition pathways, with the latter pronounced in forest soil due to frequently recalcitrant plant litter [32]. Finally, rhizodeposition fosters microorganisms and the associated bacterial channel, the dominant pathway in arable soils [33]. Taken together, nematodes are widely involved in ecosystem-level processes such as soil energy flow and nutrient cycling (e.g., [34,35]).
Their high abundance, species, and trophic diversity make nematodes good indicators of food web conditions [36,37,38]. Nematode life history traits and trophic groups provide a framework of indices to determine food web disturbance, succession, carbon pathways, enrichment, and structure [37]. Further, to assign the magnitude of ecosystem response, Ferris [39] established “metabolic footprints” as functional attributes by integrating nematode body mass and activity (i.e., respiration). These approaches are used, e.g., as functional descriptors of land use [40] or as models towards identifying sustainable soil health management [41]. Comparable to the famous model Caenorhabditis elegans, nematode trophic networks have developed into a model providing insights into the nature of ecosystem functions and services as well as soil and plant health [42,43].
1.3. Mites in Soil Food Webs
Mites are one of the most abundant groups of the mesofauna (reaching up to 85% of the soil invertebrate fauna: [44]), and are also diverse, with numerous species mainly from Mesostigmata, Oribatida and Prostigmata [45,46,47,48]. Mites are characterized by the trophic levels they occupy, the multiplicity of reproductive strategies they exhibit, and their diverse forms of dispersal [45,49]. Due to their importance in edaphic processes and their response to perturbation, e.g., in agricultural systems [50], mites have been used as indicators of environmental disturbance, particularly taxa of Oribatida (excluding the Astigmatina cohort) [51,52] and Mesostigmata [53,54,55].
Edaphic Mesostigmata are an abundant and diverse group, universally present in soil. Although fungal and detritivorous species are known, these mites are largely predators of other invertebrates, including plant pests [6,45,46,56,57,58]. Many species, especially the sub-orders Dermanyssina and Parasitina, are considered aggressive predators that have very active prey-seeking behavior in leaf litter and soil pores [47]. Soil surface- and litter-dwelling Mesostigmata are large, generally well-sclerotized, and predators of arthropods, whereas taxa inhabiting greater soil depths are relatively small, poorly sclerotized and mainly feed on small invertebrates such as nematodes [45,59].
Oribatid mites have been recognized as a characteristic soil group due to their high abundance, juvenile polymorphism and slow reproduction. In addition, oribatids are an important group involved in several processes occurring in the soil, such as decomposition of organic matter, control of microorganism and nematode populations, and facilitation of microbial colonization [45,60,61]. Oribatids are sensitive to changes in soil ecological quality, as has been proven by studies such as Maraun et al. [61], Arroyo and Iturrondobeitia [62], Minor and Cianciolo [63], Illig et al. [64], Kardol et al. [65] and Nielsen et al. [66]. Oribatids are the dominant group in the upper soil layers and reach up to 150 species in a single location [67], and between 50,000 and 500,000 individuals per square meter [45].
2. What Has Prevented the Broader Use of Soil Predatory Mites and Why Should This Change in the near Future?
To date there are several reasons for the limited use of soil predatory mites in agricultural systems for the control of soil pests: (1) Pre-plant soil sterilization by fumigation, solar energy or steam as well as pesticides applied to the soil are still recommended [70,71]. (2) Agricultural soils lack structure and conditions limit the mobility and ability of mites to forage for prey [72], and they are deficient in organic matter [73], which supports soil biota growth. (3) Only limited species of predatory mites are commercially available (Table 1), and distributing novel generalist biocontrol agents (BCAs) across borders is becoming more and more problematic due to legislation requiring specific risk assessments for each species [74]. (4) The effect of prey species’ suitability on life table parameters are usually tested separately for each prey, but predator fitness is likely to be better when different prey types are offered together [22]. (5) Identification of species must be performed by experts in soil predatory mite taxonomy, an expertise in shortage on a global scale [75]. (6) Knowledge is lacking on the predation capacity of predatory mites in agricultural soils with given abiotic and biotic attributes, because most studies are conducted in artificial laboratory set-ups. Of the 177 publications we reviewed with reports of trophic relationships between mites and nematodes, 149 were exclusively laboratory experiments, especially in Petri dishes; 17 were conducted in potted plants either in the laboratory or in greenhouses; and only 24 included observations or experiments in the field (Table S1).
While soil sterilization is still being used, mostly in conventional agriculture, for the control of soil pests, this cannot be considered to be a sustainable solution as it needs to be routinely repeated and negatively impacts soil diversity [76]. Additionally, pesticides applied to the soil pose a potential hazard to the beneficial soil meso- and macrofauna [77]. However, the availability of these pesticides is decreasing as registrations are being reviewed and revoked by the respective authorities [78,79,80].
Similar to the scientific and public awareness of the need for the reduction of pesticide use [81,82], there are initiatives across the globe, all with the common objective of caring for soils [83], aimed at enhancing soil fertility and preserving and conserving soil biodiversity [84] including: the EU Soil and Biodiversity Strategies, EU Soil Observatory, FAO UN Global Soil Partnership (GSP), the Intergovernmental Technical Panel on Soils (ITPS), the Convention on Biological Diversity (CBD) and the Global Soil Biodiversity Initiative (GSBI). Ultimately, these initiatives will lead to environmentally friendly soil practices, supporting functional soil food webs capable of providing the necessary ecosystem services.
The drawback of a low number of commercialized species and the legislation required for registering new species will likely continue to restrict their use for augmentative biological control. However, these limitations do not apply to the use of known and novel BCAs in conservation biological control programs. This is especially true when soil amendments, such as cover crops [85,86] and various composts [87,88], are utilized for enhancing abiotic and biotic soil attributes pertaining to the conservation of multiple species of arthropods and FLN.
Finally, embracing soil biodiversity for the conservation biological control of soil pests is challenging because it requires specialized taxonomists, an expertise that is in short supply across the globe. In recent studies [89,90], we have met this challenge by applying an integrative taxonomic approach, utilizing the Barcode of Life Database system (BOLD) platform. This entailed a combined effort where agronomists and/or ecologists were responsible for collecting and extracting samples, specialized personnel for DNA barcoding and expert taxonomists for morphological identification. These studies contributed novel sequences of expertly identified soil predatory mites to BOLD and provided new information on the species diversity of soil mites in farming systems.
3. Nematophagous Mites
Despite the large number of published studies showing interactions between mites and nematodes (177 publications, Table S1; mostly in laboratory experiments), the understanding of this interaction is still neglected, as stated by Walter and Proctor [47]. According to these authors, there are two main reasons for the lack of study. The first is related to the conception that mites are restricted to air-filled soil pore spaces, while nematodes are restricted to water-filled pore spaces, limiting their encounter and interaction. However, this argument is not entirely accurate, because it is known that nematodes can move between air-filled pore spaces, as well as that some mites are able to extend their chelicerae into water films to extract nematodes. The second mis-assumption is that the size and soft elongated body of nematodes allows them to enter small soil pores, not accessible for large mites with sclerotized and inflexible shields. However, nematophagous mites, such as many Rhodacaridae [8] and several Ascidae [93], have an elongated and flexible body, allowing them to move through small spaces, or have chelicerae capable of reaching these small pores [47,94] (for details see Section 5.2).
3.1. Oribatida (Non-Astigmatina)
Few studies have been conducted on the interaction between Oribatida and nematodes. Of the 177 references reviewed, only 13 studies investigated Oribatida (Table S1), which corresponds to approximately 60 species or morphospecies of Oribatida, related to about 20 species of nematodes or mixtures of different nematodes. The first studies showing a relationship between Oribatida and nematodes were conducted by Rockett and Woodring [95,96] who observed Pergalumna (P.) emarginata (Banks) (cited as Pergalumna omniphagous Rockett and Woodring) feeding on a FLN and a PPN. Based on their observations, these authors suggested that oribatids could be an important regulating factor for nematode populations. Following these studies, nematode consumption was shown for various oribatids, yet only in a few cases (e.g., [6,97,98,99]) species were reported to have succeeded in developing on a nematode-based diet. As for the type of nematodes, most studies reveal FLN as an important prey, but feeding and reproduction on PPNs and APNs takes place too (Table S1). Recently, DNA gut content analyses revealed FLN as a frequent diet of 14 Oribatida species in forest soil [100,101,102] (see Section 4). This indicates that nematodes are part of the diet in the field, despite the availability of other resources.
3.2. Astigmatina
While Astigmatina is a group of mites that is not well-represented in soil, Acaridae may be especially abundant in leaf litter and the upper soil layers rich in organic matter [68]. Although scarce (19 out of 177 published studies reviewed), there are some reports of nematophagy. The first mention of this interaction was made by Karg [103], reporting eleven- and five-times higher populations of Rhizoglyphus echinopus (Fumouze and Robin) and Tyrophagus similis Volgin (cited as Tyrophagus dimidiatus (Hermann), Tyrophagus infestans Berlese), respectively, in fields contaminated with Globodera rostochiensis (Wollenweber) Skarbilovich (cited as Heterodera rostochiensis Wollenweber) compared to non-infested fields. Even though no direct trophic relationship between nematode populations and Astigmatina was established in this study, it is possible that the mites may have increased their populations because of the large availability of nematodes as food source. Tyrophagus similis was later reported as a consumer also of FLN [97,98], while R. echinopus has only been reported as a consumer of PPNs [104]. The latter is also true for Rhizoglyphus robini Claparede, where Meloidogyne incognita (Kofoid and White) Chitwood was observed as prey [105]. The latter two species are considered important soil-dwelling pests of plants such as bulbs [106] that under special conditions can have a regulating effect on soil nematodes [68]. The Astigmatina species with the most reports of nematophagy is Tyrophagus putrescentiae (Schrank), using 14 different species of FLNs [107,108], 17 species of PPNs [108,109,110,111] and one species of APN [112] as prey. Moreover, Sancassania species are consumers of entomopathogenic nematodes, reducing their populations in the field [113].
The effect of a nematode-based diet compared to other diets has not been extensively studied for Astigmatina species. There are only reports for Sancassania manure (Eraky & Osman) and Sancassania sp. (cited as Caloglyphus sp.). In the first case, fecundity was higher with an unidentified yeast and cheese, than with Meloidogyne sp. [114]. For Sancassania sp., a slight delay in development was observed when feeding on Cephalobus sp. compared to an unidentified yeast [115]. Further experiments are needed to determine whether a mixed diet including nematodes can be beneficial for these mites.
3.3. Prostigmata
For Prostigmata only 11 publications recorded consumption of nematodes (Table S1). More studies have been performed, but the results show that this mite group does not seem to have a nematophagous habit for the most part. A total of 16 species, mainly of the family Cunaxidae, are reported to consume nematodes, with most able to be reared or developed on nematode-based diets [116,117,118,119,120]. Walter [7] found that several species of Eupodidae are capable of feeding on nematodes, but at low frequency and with a preference for microphytes.
Comparison of the effect of a nematode-based diet and other diets on mite fitness has only been conducted for Cunaxa capreolus (Berlese) and Pulaeus pseudominutus (Shiba). The fecundity of C. capreolus was higher when feeding on J2 of M. incognita than when feeding on J2 of Tylenchulus semipenetrans Cobb [120] and was higher when feeding on the colembolan Entomobrya musatica Stach than on the nematode Mesorhabditis scanica (Allgén) Sudhaus (cited as Rhabditis scanica Allgén) [119]. In the case of P. pseudominutus, no differences in fecundity were observed when mites were fed with the fungi Fusarium oxysporum Schltdl. and Pythium spinosum Sawada compared to the nematode Rhabditella muscicola Andrássy [118]. As with Oribatida, further studies are needed to establish the relationship these mites have with nematodes in the soil and how this interaction may be beneficial in the control of other organisms, including plant pests.
3.4. Endeostigmata
In the suborder Endeostigmata, nematophagous mites have been reported [6,47,121] in five species belonging to two families (Table S1). Most studies with these mites have been conducted with FLN; however, Alicorhagia fragilis Berlese and Alycus roseus Koch were shown to consume the entomopathogenic nematode Steinernema feltiae (Filipjev) Wouts, Mráček, Gerdin and Bedding. For most of the reported species of this family, reproduction and development have been observed with a nematode-based diet.
3.5. Mesostigmata
Mesostigmata is the group with the most studies (148 publications out of 177 reviewed) of interactions with nematodes (Table S1). However, only four were conducted on communities of mites and not on a specific species. In these studies, either nematode consumption was observed [47] or that the presence of these predatory mites coincided with the reduction of PPNs [105,122,123] (cited by [124]). Among these studies, Stirling [105] reported that Mesostigmata mites were present in all sawdust-based amendments added to pots in field and greenhouse, which coincided with a reduction in M. incognita density and a reduction in root galls compared to sterilized soil and sand.
3.5.1. Gamasina
Within Gamasina, nematophagy has been reported in 250 species within 19 families, by 134 studies performed with about 106 different species of nematodes, 19 unidentified nematodes and several species mixtures (Figure 2, Table S1). Azevedo et al. [125], Castilho et al. [8], McMurtry et al. [126], Moreira and Moraes [127], and Moraes et al. [93] reviewed the dietary habits (including nematodes) up to 2015 for Gamasina mites including Ascidae, Blattisociidae, Digamasellidae, Eviphididae, Heatherellidae, Laelapidae, Leptolaelapidae, Macrochelidae, Melicharidae, Ologamasidae, Parasitidae, Parholaspididae, Phytoseiidae, and Rhodacaridae.
The family with the highest number of nematophagous species is Ascidae (46 species, Figure 2). Species in this family consume FLNs, APNs and PPNs. The ascid species consuming the most nematode species is Gamasellodes vermivorax Walter (11 FLN species, two PPN species, two APN species, one species of unidentified nematode and four mixtures of different nematodes), followed by Protogamasellus mica (Athias-Henriot) (nine FLN species, five PPN species, one species of unidentified nematode and four mixtures of different nematodes). Generally, reproduction and development have been observed with a nematode-based diet, and in several cases, reproductive parameters have been better with FLNs than with PPNs [128,129]. The few comparisons of nematode prey with other diets showed that effects depended on the mite species. While in some cases shorter development times were reported with a nematode diet (e.g., G. vermivorax; [7,130]), in others higher fecundity occurred with other diets (e.g., Arctoseius cetratus (Sellnick) [131]; Protogamasellus minutus Nasr [132]; Protogamasellus similis Genis, Loots and Ryke [133]). In the greenhouse, the colonization of Ascidae species has been observed on PPN-infested pots ([117,134]) and also reduction of both FLN and PPN populations when, e.g., P. mica were present [135]. Several species of this family are considered to belong to the small-pore nematophagous guild because of their size and body shape [6,47,59] (see Section 5.2).
Laelapidae and Macrochelidae are the families with the second-highest number of nematophagous species (both with 30 species, Figure 2). For both families, most of the reports involve FLN. However, there are more reports of species consuming APN and PPN in Laelapidae than in Macrochelidae (Table S1, Figure 2).
Laelapidae is well-represented in soil and leaf litter, although it is also found in association with animals [136]. Cosmolaelaps indicus Bhattacharyya (cited as Hypoaspis calcuttaensis Bhattacharyya) consumed many different nematodes (16 FLN species, 17 PPN species) [137] (Table S1), followed by Gaeolaelaps aculeifer (Canestrini) (cited as Hypoaspis aculeifer Canestrini) (four species of FLN, five species of PPN, two species of APN, two species of unidentified nematodes and two mixes of FLN and PPN). The latter is a commercially produced species for the control of agricultural pests that have at least part of their life cycle in the soil [136]. Several field studies and greenhouse experiments for PPN control have been conducted with the commercially available species G. aculeifer, Stratiolaelaps miles (Berlese) and Stratiolaelaps scimitus Womersley demonstrating reductions in nematode abundances [24,138,139,140,141].
Comparison of a nematode-based diet with other diets has been conducted in species such as Cosmolaelaps claviger (Berlese) [142], Cosmolaelaps jaboticabalensis Moreira, Klompen & Moraes [143], Cosmolaelaps simplex Berlese [144], Cosmolaelaps vacuus (Michael) (cited as Hypoaspis (Cosmolaelaps) vacua (Michael)) [7,130], G. aculeifer [145,146,147], Gaeolaelaps oreithyiae Walter and Oliver [148], S. miles [149], and S. scimitus (Figure 3) [24,141]. In many cases, a positive effect on the reproductive parameters and developmental times of mite species has been observed with a nematode-based diet, especially FLN (e.g., [7,130,143,146,147,148]). On one hand, such positive effects of a nematode-based diet can hamper predation on a target pest, e.g., Frankliniella occidentalis (Pergande), an insect pest of great importance in various crops [143]. On the other hand, a study by Azevedo et al. [24] shows that the effect of FLN as supplementary food can result in a decrease in damage caused by pests such as root-knot nematode, M. incognita, likely mediated by an increased predator fitness.
Macrochelidae is a family especially associated with temporary habitats such as excreta and decaying animals, so efforts to use this family as a biological control agent have largely focused on pest organisms associated with these environments such as different species of flies [125]. It is also in these habitats, or on substrates simulating these environments, that the trophic relationship with nematodes, especially FLN, has been observed (Table S1). Macrocheles muscaedomesticae (Scopoli) consumed most nematode species (11 FLN species, one PPN species and four unidentified FLN species), followed by Glyptholaspis confusa (Foà) (five FLN species). For many mites in this family, reproduction and development have been observed on a nematode-based diet. Also, the comparison with other diets was performed and in several cases preference or higher fecundity has been found when mites feed on fly eggs or larvae (e.g., [150,151,152,153]). However, higher fecundity has been observed also with nematode diets (e.g., [150,154,155]) and even the addition of nematodes to other diets has led to higher reproduction (e.g., [154,156,157]). Although there may be a decrease in predation of the target pest with nematodes as an alternative diet, mite fecundity increases [23,154]. Ultimately, such a higher predator population can control the pest of interest in the medium to long term [23]. Another important point to consider is that adults of several species of this family seem to prefer fly eggs or larvae for feeding, but in immature stages the preference may be towards nematodes (e.g., [156,157,158]). This suggests that a diverse diet including nematodes may favor the pre-adult development and sustainability of Macrochelidae to perform as BCAs (see also Section 5.1).
In Ologamasidae and Blattisociidae, 25 species have been shown to consume nematodes for each family (Figure 2, Table S1). For Ologamasidae only FLN consumption was observed (e.g., [115,159,160]), while for Blattisociidae predation on both FLN and PPN was reported (Table S1). For the latter, a nematode-based diet was also compared to other diets. For Lasioseius africanus Nasr [161], Lasioseius berlesei (Oudemans) [134], and Lasioseius floridensis Berlese [162], higher fecundity occurred with a nematode diet compared to other food resources. However, in other species such as Lasioseius queenslandicus (Womersley) (cited as Lasioseius athiasae Nawar & Nasr) higher fecundity has been observed with other diets [163,164]. In experiments with PPN, species such as Blattisocius dolichus Ma [124,165] and Lasioseius penicilliger Berlese [138] have been observed to reduce nematode populations.
In the case of Parasitidae, 18 nematophagous species have been reported (Figure 2). The species consuming the highest number of nematode species is Pergamasus septentrionalis (Oudemans) (four FLN species and two APN species) [100,101,102]. When comparing a nematode-based diet with others, higher fecundity with nematodes has been observed in Parasitus bituberosus Karg [22,166], and Porrhostaspis lunulata Müller [167]. Rueda-Ramírez et al. [22] further showed that daily oviposition was 1.4 times higher when the target prey, F. occidentalis, was mixed with the FLN Rhabditella axei (Cobbold), than with a solely nematode diet. In this family, as in Macrochelidae, a varied diet seems important as it has been observed that immature stages may prefer nematodes, while feeding behavior may be different in the adult stage [22,158].
Diagamasellidae, Rhodacaridae and Melicharidae have reports of 16, 14 and 12 nematophagous species, respectively (Figure 2, Table S1). Digamasellidae and Melicharidae were shown to feed on APN, FLN, and PPN, while in Rhodacaridae there are only reports of consumption of FLN and PPN. This is not unusual as species of the first two families are associated with animals, especially arthropods, while Rhodacaridae mostly inhabit soil [8]. In the latter family several species are considered generalist predators, but they reproduce and develop very well on a nematode-based diet [6,59,168]. Moreover, their shape and size give them the ability to reach spaces in the soil where nematodes are present, comparable to Ascidae (see Section 5.2) [6,47,59]. In these three families, species capable of developing and reproducing on a nematode-based diet have been observed, which in several cases, had a more positive effect on the reproductive parameters of the mite population than other diets (Table S1).
For Eviphididae, only eight nematophagous species have been reported (Figure 2, Table S1) despite being considered a family with commonly nematode specialist species [6,7,47], which may be because this family is relatively small (about 108 species) [169] compared to Macrochelidae and Laelapidae, or owing to the few studies conducted (12 publications). For most, reproduction (e.g., [170,171]), development (e.g., [6,7]) or preference for nematodes (e.g., [7,145]; Table S1) has been observed. In the case of Alliphis siculus (Oudemans), populations increase in fields contaminated with PPN [145,172] and the addition of nematodes to the diet considerably increases their abundance [173]. Eviphididae is a family that seems to be closely related to nematodes, suggesting their regulatory potential on populations of these organisms.
Other families (see Figure 2) have fewer reports of nematophagous species (five or less), possibly because they are less common or less-studied families, e.g., Zerconidae, or due to different feeding preferences e.g., Veigaiidae [6,8,59]. Among these, Zerconidae deserve highlighting, as species specialized in consuming nematodes have been reported [7] and, moreover, in microcosm experiments their presence coincides with low nematode populations [174].
3.5.2. Uropodina
Within Uropodina, nematophagy was shown in 15 species within 5 families (15 publications; Table S1). The family with the most species reported is Uropodidae, with Uropoda cassidea (Hermann) consuming the most nematode species in this group (six FLN species and one APN species). Reports of the nematophagy of U. cassidea, as well as that of Trachytes aegrota (Koch), are based on the detection of nematode genetic material in the gut of the mites [100,101,102]. Other studies have shown that different uropodid species prefer nematodes (e.g., [152,158,175]) and that addition of nematodes in the diet may increase reproduction (e.g., [176,177]).
3.5.3. Other Mesostigmata Groups
Nematophagy has also been reported in one species of Microgyniina, four species of Sejida and ten species of Trigynaspida (Table S1). Further studies are needed to better understand the effect of nematodes on different biological parameters of these mites.
4. Nematodes as a Food Source for Mites in Ecosystems
Nematodes serve as a diet for many predatory soil animals, particularly other nematodes [28,178] as well as several groups of arthropods such as Acari, Coleoptera, Collembola, and Symphyla [59,174,179,180] (for mites see Section 3). Although early soil food web models assigned mites as top predators [181] (Figure 1), the trophic link to nematodes has not received attention for a long time, with exception of Mesostigmata, as worm-like prey was indicated by their mouthpart morphology [58,182,183].
A suite of advanced methodological approaches now provides deeper insight into mite feeding behavior in the field. Using stable isotopes (15N/14N) Schneider et al. [184] showed the first evidence that oribatid mites span over four trophic levels, from phycophages over primary and secondary decomposers to carnivores. The combination of 15N isotopic signals from forest soil mesostigmatids and oribatids, with data from laboratory feeding experiments, further pointed to a central role in regulating the population dynamics of the microfauna [185]. Finally, Klarner et al. [186] integrated the variation in 13C/12C and 15N/14N of 40 mesostigmatid mites from an old growth beech forest soil, which revealed that the diet of the dominant species consisted in large part of nematode prey.
A more fine-tuned approach is molecular gut content analysis, which characterizes feeding interactions at a high level of taxonomic resolution [187]. Heidemann et al. [100] established the first primers for binary trophic links between nematodes and mites in laboratory no-choice experiments. Subsequently, for forest soil food webs, nematode predation was assigned for species in Uropodina, Gamasina, and Oribatida [102]. Thus, FLN form an important prey for mites, also in groups commonly regarded as detritivores such as Oribatida. In a follow up study, Heidemann et al. [101] showed that nematode consumption by mites differed due to microhabitat, i.e., litter, grass, and moss, indicating distinct trophic niches in this predator–prey interaction.
This considerable top–down control of predatory and omnivore mites on nematodes can impose biocontrol on PPN in the rhizosphere [123,188], but also affect bacterial and fungal feeders (up to 50% less biomass [189]) and the activity of their microbial resources [180]. This trophic linkage even results in the fact that ecological indices based on nematode community structure (see Section 1.2) are reliable predictors of the trophic composition and functional characteristics of soil mite assemblages [50]. For example, bacterial-feeding and predatory nematodes, together with predatory mites, were associated with high food web enrichment and structure. This highlights the close relationship between nematodes and mites in soil ecosystems.
5. Variables Important for Trophic Interactions
5.1. Nematodes as an Important Source of Fatty Acids in Soil Food Webs
Nutritional quality is a key determinant for population development in soil microarthropods and can affect, e.g., fitness traits such as body mass [190], offspring and longevity [146] or even male reproductive behavior and mating success [191]. Using the oribatid Archegozetes longisetosus as model, Brückner et al. [192] showed that diet quality affected nine different traits related to life history, morphology, and exocrine gland chemistry. These effects were mainly based on element stoichiometry in the diet, with offspring correlated to the C/N ratio, while body mass was linked to the C/P ratio [193]. To avoid nutritional imbalances active prey mixing is common, also in predatory mites [194]. For mass rearing of mites as BCAs, dietary supplements as well as improved prey quality were shown to have beneficial impact on biocontrol parameters, i.e., the robustness of the predatory mite population via its target pest [195,196].
For a long time, soil microarthropods were assumed to optimize food intake by selection according to C/N ratio [192,197,198], while other nutrients were rarely taken into account. Only recently, the lipid composition of the diet was established, in particular the content of ω3 long-chain polyunsaturated fatty acids (ω3 LC-PUFA). Eicosapentaenoic acid (EPA, 20:5 ω3) and docosahexaenoic acid (DHA, 22:6 ω3) are considered as semi-essential biomolecules in food webs, due to their key physiological functions in many animals [199]. However, ω3 LC-PUFAs are not widely available in terrestrial food webs [200]. Early work reports the synthesis of EPA in four species of the FLN Caenorhabditis [201,202] as well as EPA occurrence in Turbatix aceti [203]. Recently, the ability to synthesize EPA was described in the taxa Panagrolaimus and Mesorhabditis [204]. This suggests nematodes are an important source for ω3 LC-PUFA in the soil food web.
Mites are known to perform the initial step of PUFA biosynthesis, the desaturation from oleic acid to linoleic acid [205,206]. Experiments with 13C glucose as substrate showed that linoleic acid is stably converted into 6,9, heptadecadiene, a major secretory component [207]. To date, in the few lipid patterns available for mites, no ω3 LC-PUFAs are reported for body lipids, oil glands or cuticular surface [208,209,210]. That the provision of EPA and DHA by nematode prey enhances consumers’ life cycle, health, and fitness, was shown for Collembola [211]. Similarly, nematodes can provide these nutritionally valuable biomolecules to their major predators, the mites, fostering overall fitness and thereby biocontrol function.
5.2. Size Matters?
At the micro-scale, soil structure constrains food accessibility. As stated by Walter and Ikonnen [59] and observed by other authors [8,212] small Rhodacaridae-type mites (species of Ascidae, Digamasellidae and Rhodacaridae, especially) have a small, elongated body with a split dorsal shield and a flexibility that allows them to access deeper soil layers. There, they can reach depths [8,59] or pore spaces [47] populated by nematodes and feed on them. These mites were assigned by Walter and Ikonnen [59] as the nematophagous small-pore guild (Figure 4a).
Besides the many reports of nematode preference and ability to control populations in the small-pore nematophagous guild (see Table S1), also larger mites (Laelapidae, Macrochelidae, and Parasitidae, especially, Figure 4b) voraciously feed on nematodes (e.g., [22,23,150,155]). It can be assumed that medium-to-large sized mites, mostly inhabiting the upper soil layers, predominantly feed on nematode populations associated with the organic matter present (e.g., all trophic groups of FLN and PPN, attacking the roots of young plants), whereas smaller mites accessing greater depths and small pores mainly feed on nematodes that occupy these spaces (e.g., bacterial- and fungal-feeding FLN and juvenile PPN).
Size and shape may confer the ability of certain mites to reach spaces where nematodes are present in the soil. However, this is not always associated with being exclusively nematophagous. For example, Walter et al. [6] found several species of Rhodacaridae-type mites (e.g., Gamasellodes vermivorax Walter and Rhodacarellus silesiacus Willmann) that, although they fed and developed very well on nematodes, they also fed on other prey such as Collembola and mites. These same authors classified some species of Eviphididae (Mesostigmata) and Endeostigmata, whose body format is not slender and without a divided dorsal shield, as nematode specialists. This food flexibility within guilds or morphotypes of mites can be viewed as an adaptation to the accessibility of nematode resources in the soil matrix.
6. Enhancing Conservation and Efficacy of Soil Acarine Biocontrol Agents (ABA)
Beretta et al. [4] recently reviewed the use of soil predatory mites, mostly applied as augmentative biological control agents for the control of edaphic stages of thrips, flies, mites, and PPN in greenhouse and screenhouse systems. While a considerable number of studies were cited, most were conducted on the two commercially available laelapid species, G. aculeifer and S. scimitus (Table 1). To improve the efficacy and establishment of augmentatively released soil predatory mites, the authors encouraged the provisioning of alternative foods and the addition of soil mulches. For many greenhouse systems these approaches are certainly valid since they are often soilless. In contrast to augmentative biological control in such protected agrosystems, conservation biological control in outdoors cropping systems promotes the ‘modification of the environment or existing practices to protect and enhance specific natural enemies or other organisms to reduce the effect of pests’ [213].
Ecofriendly soil management of abiotic and biotic parameters is crucial for the conservation of ABA. Abiotic interrelated physical attributes such as soil structure, texture, and water content [214] can be limiting factors for ABA foraging and survival [72] (see also Section 5.2). Abiotic soil attributes, especially soil organic matter, are essential for the growth of microbiota, upon which free-living nematodes graze [215,216], the latter serving as essential prey (as discussed above) for many soil predatory mite species. Conservation agriculture, promoted by the FAO as an ecofriendly approach, includes three principles: minimum mechanical soil disturbance, permanent soil organic cover, and species diversification [217]. Compared to the conventional practices of tillage, bare soil between crops and minimal crop rotation, conservation agriculture provides a solid foundation for re-generating functional soil food webs. Minimal mechanical soil disturbances help to conserve and preserve soil structure and texture, while permanent soil organic cover generates soil organic matter, moderates temperatures and retains soil moisture. Finally, species diversification by crop rotation [218] and the introduction of service crops [219] enhances soil microbiota biodiversity.
In a study conducted in a Long Term Research Agricultural Systems (LTRAS) project (12 years after it was established), the abundance of bacterial-feeding and predatory nematodes, as well as predatory and omnivorous mites, were greater in organic plots, predominantly under no till, and lowest in the conventional standard tillage [50]. In contrast, in a more recent study in olives and vineyards in semi-arid Mediterranean agroecosystems in Spain, the soil food webs of the organic and conventional systems were degraded, with similar low structure and enrichment nematode indices [220]. The authors suggested that organic management in itself is not enough and that soil conservation practices such as reduced tillage and cover cropping are necessary to enhance agroecosystem sustainability. Wang et al. [221] evaluated in a two-year study the effects of strip till cover cropping. In the second year, yield was significantly higher in the sun hemp cover crop treatment, as were the nematode structure index and abundance of predatory mites. In a long-term field study assessing the effects of tillage and cover crop over 16 years, management systems using cover crops strongly enhanced the nematode assemblages in the field soil [222]. Recently, using a soil amendment, Udalova et al. [223] demonstrated that the addition of insect-compost enhanced the abundance of bacteriophagous, polyphagous, and predatory nematodes, and reduced root-knot nematodes, M. incognita, in tomato.
Just as many soil amendment studies focused on nematodes (some cited above) have not monitored mite communities, likewise, several mite studies have not taken into account FLN. Messelink and Kogel [224] applied a biowaste compost for the introduction of predatory mites and Grosman et al. [225] used a mulch layer and the provisioning of Astigmatina prey to enhance establishment, abundance and performance of the predatory mites Macrocheles robustulus and Gaeolaelaps aculeifer. Esteca et al. [226] applied dehydrated coffee husk and pulp as a mulch for the conservation biological control of Tetranychus urticae. In all three cases, these soil amendments probably altered FLN abundances, which could have in turn affected predatory mite populations and biocontrol efficacy (Figure 5).
The time and taxonomic skills required to morphologically identify mesofauna communities from soil samples are most likely the reasons why soil amendment manuscripts are usually focused on either mites or nematodes, but not both. Future studies on the effects of soil conservation practices on ecosystem services could consider using metabarcoding of soil samples, which have recently been used for identifying mite [227] and nematode communities [228].
7. Future Research Needed
Although numerous studies have been published on soil mite–nematode interactions, most have been conducted in the lab in small arenas with a single species of mite and nematode, assessing the effects on mite fitness and/or nematode control (Table S1). An important next step is to transfer laboratory research to semi-field and field conditions. Modern molecular approaches can support this in both taxonomic (i.e., barcoding) and ecological (i.e., gut content analyses) studies, allowing for a more realistic picture of the trophic linkages between mites and nematodes and its significance for biological control.
Future research should study soil mite and nematode assemblages in different cropping systems, soils, and climate zones. Soil degradation is recognized as a global threat and soil organic matter depletion is a significant factor in this. Novel conservation soil management strategies will aim at increasing soil organic matter and reducing soil erosion and compaction. Future research should assess how these management strategies will improve soil health and biodiversity, which largely include mites and nematodes. Harnessing the local biodiversity can favor the increase and sustainability of biological control agents and favor the restoration of soil food webs (Figure 5). Strategies may include the use of organic amendments derived from animal feed production and or composting, which increase the accumulation of soil organic matter and in turn foster soil life as well as biocontrol.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/agronomy13010032/s1, Table S1: Reports of mite species per group and family reported consuming nematodes [229,230,231,232,233,234,235,236,237,238,239,240,241,242,243,244,245,246,247,248,249,250,251,252,253,254,255,256,257,258,259,260,261,262,263,264,265,266,267,268,269,270,271,272,273,274,275,276,277,278,279,280,281,282,283,284,285,286,287,288,289,290,291,292,293,294,295,296,297,298,299,300,301,302,303,304,305,306,307,308,309,310,311,312,313,314,315,316,317,318,319,320,321,322,323,324].
Author Contributions
Conceptualization, D.R.-R., E.P. and L.R.; methodology, D.R.-R.; validation, D.R.-R., E.P. and L.R.; formal analysis, D.R.-R., E.P. and L.R.; investigation, D.R.-R., E.P. and L.R.; writing—original draft preparation, D.R.-R., E.P. and L.R.; writing—review and editing, D.R.-R., E.P. and L.R.; visualization, D.R.-R., E.P. and L.R.; supervision, E.P. and L.R.; project administration, L.R.; funding acquisition, E.P. and L.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the grant RU780/20-1 (‘Harnessing the soil food web for the biological control of root-knot nematodes’) received from the German Research Foundation (DFG).
Acknowledgments
To Gary Bauchan, Joseph Mowery, and Ronald Ochoa, U.S. Department of Agriculture (USDA), for their help with taking the amazing images of the mites feeding on nematodes. To Lynn Carta, U.S. Department of Agriculture (USDA), for providing the nematodes for photographs and identification of nematodes.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Neher, D.A.; Barbercheck, M.E. Soil microarthropods and soil health: Intersection of decomposition and pest suppression in agroecosystems. Insects 2019, 10, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, A.; Sabelis, M.W. Alternative food and biological control by generalist predatory mites: The case of Amblyseius swirskii. Exp. Appl. Acarol. 2015, 65, 413–418. [Google Scholar] [CrossRef]
- Muñoz-Cárdenas, K.; Ersin, F.; Pijnakker, J.; van Houten, Y.; Hoogerbrugge, H.; Leman, A.; Pappas, M.L.; Duarte, M.V.A.A.; Messelink, G.J.; Sabelis, M.W.; et al. Supplying high-quality alternative prey in the litter increases control of an above-ground plant pest by a generalist predator. Biological. Control. 2017, 105, 19–26. [Google Scholar] [CrossRef]
- Beretta, G.M.; Deere, J.A.; Messelink, G.J.; Muñoz-Cárdenas, K.; Janssen, A. Review: Predatory soil mites as biocontrol agents of above and below-ground plant pests. Exp. Appl. Acarol. 2022, 87, 143–162. [Google Scholar] [CrossRef] [PubMed]
- Nicol, J.M.; Turner, S.J.; Coyne, D.L.; den Nijs, L.; Hockland, S.; Maafi, Z.T. Current Nematode Threats to World Agriculture. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Jones, J., Gheysen, G., Fenoll, C., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 21–43. ISBN 978-94-007-0434-3. [Google Scholar]
- Walter, D.E.; Hunt, H.W.; Elliott, E.T. Guilds or functional groups? an analysis of predatory arthropods from a shortgrass steppe soil. Pedobiologia 1988, 31, 247–260. [Google Scholar]
- Walter, D.E. Nematophagy by soil arthropods from the shortgrass steppe, Chihuahuan desert and Rocky Mountains of the central United States. Agric. Ecosyst. Environ. 1988, 24, 307–316. [Google Scholar] [CrossRef]
- Castilho, R.C.; Venancio, R.; Narita, J.P.Z. Mesostigmata as biological control agents, with emphasis on Rhodacaroidea and Parasitoidea. In Prospects for Biological Control of Plant Feeding Mites and Other Harmful Organisms; Carrillo, D., de Moraes, G.J., Peña, J.E., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–31. ISBN 978-3-319-15042-0. [Google Scholar]
- Gessner, M.O.; Swan, C.M.; Dang, C.K.; McKie, B.G.; Bardgett, R.D.; Wall, D.H.; Hättenschwiler, S. Diversity meets decomposition. Trends Ecol. Evol. 2010, 25, 372–380. [Google Scholar] [CrossRef]
- de Vries, F.T.; Thébault, E.; Liiri, M.; Birkhofer, K.; Tsiafouli, M.A.; Bjørnlund, L.; Bracht Jørgensen, H.; Brady, M.V.; Christensen, S.; de Ruiter, P.C.; et al. Soil food web properties explain ecosystem services across European land use systems. Proc. Natl. Acad. Sci. USA 2013, 110, 14296–14301. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef]
- Bonkowski, M.; Villenave, C.; Griffiths, B. Rhizosphere fauna: The functional and structural diversity of intimate interactions of soil fauna with plant roots. Plant Soil 2009, 321, 213–233. [Google Scholar] [CrossRef]
- Gebremikael, M.T.; Steel, H.; Buchan, D.; Bert, W.; De Neve, S. Nematodes enhance plant growth and nutrient uptake under C and N-rich conditions. Sci. Rep. 2016, 6, 32862. [Google Scholar] [CrossRef] [PubMed]
- Whalen, J.K.; Kernecker, M.L.; Thomas, B.W.; Sachdeva, V.; Ngosong, C. Soil food web controls on nitrogen mineralization are influenced by agricultural practices in humid temperate climates. CABI Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2013, 2013, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Whalen, J.K. Managing Soil Biota-Mediated Decomposition and Nutrient Mineralization in Sustainable Agroecosystems. Adv. Agric. 2014, 2014, 384604. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Roy, M.M.; Jaiswal, A.K.; Baitha, A. Soil Arthropods in Maintaining Soil Health: Thrust Areas for Sugarcane Production Systems. Sugar. Tech. 2018, 20, 376–391. [Google Scholar] [CrossRef]
- Samaddar, S.; Karp, D.S.; Schmidt, R.; Devarajan, N.; McGarvey, J.A.; Pires, A.F.A.; Scow, K. Role of soil in the regulation of human and plant pathogens: Soils’ contributions to people. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200179. [Google Scholar] [CrossRef]
- Kergunteuil, A.; Bakhtiari, M.; Formenti, L.; Xiao, Z.; Defossez, E.; Rasmann, S. Biological Control beneath the Feet: A Review of Crop Protection against Insect Root Herbivores. Insects 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Wolkovich, E.M. Reticulated channels in soil food webs. Soil Biol. Biochem. 2016, 102, 18–21. [Google Scholar] [CrossRef]
- de Vries, F.T.; Wallenstein, M.D. Below-ground connections underlying above-ground food production: A framework for optimising ecological connections in the rhizosphere. J. Ecol. 2017, 105, 913–920. [Google Scholar] [CrossRef] [Green Version]
- Pausch, J.; Hünninghaus, M.; Kramer, S.; Scharroba, A.; Scheunemann, N.; Butenschoen, O.; Marhan, S.; Bonkowski, M.; Kandeler, E.; Scheu, S.; et al. Carbon budgets of top- and subsoil food webs in an arable system. Pedobiologia 2018, 69, 29–33. [Google Scholar] [CrossRef]
- Rueda-Ramírez, D.; Rios-Malaver, D.; Varela-Ramírez, A.; Moraes, G.J. de Biology and predation capacity of Parasitus bituberosus (Acari: Mesostigmata: Parasitidae) on Frankliniella occidentalis (Thysanoptera: Thripidae) and free-living nematodes as complementary diet. Pest Manag. Sci. 2019, 75, 1819–1830. [Google Scholar] [CrossRef]
- Azevedo, L.H.; Leite, L.G.; Chacon-Orozco, J.G.; Moreira, M.F.P.; Ferreira, M.P.; González-Cano, L.M.; Borges, V.; Rueda-Ramírez, D.; Moraes, G.J.; Palevsky, E. Free living nematodes as alternative prey for soil predatory mites: An interdisciplinary case study of conservation biological control. Biol. Control. 2019, 132, 128–134. [Google Scholar] [CrossRef]
- Azevedo, L.H.; Moreira, M.F.P.; Pereira, G.G.; Borges, V.; de Moraes, G.J.; Inomoto, M.M.; Vicente, M.H.; de Siqueira Pinto, M.; Peres, L.E.P.; Rueda-Ramírez, D.; et al. Combined releases of soil predatory mites and provisioning of free-living nematodes for the biological control of root-knot nematodes on ‘Micro Tom tomato. ’ Biol. Control. 2020, 146, 104280. [Google Scholar] [CrossRef]
- Blaxter, M. Nematodes: The Worm and Its Relatives. PLOS Biol. 2011, 9, e1001050. [Google Scholar] [CrossRef]
- Yeates, G.W. Nematodes as soil indicators: Functional and biodiversity aspects. Biol. Fertil. Soils 2003, 37, 199–210. [Google Scholar] [CrossRef]
- van den Hoogen, J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.A.; de Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.S.; et al. Soil nematode abundance and functional group composition at a global scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Yeates, G.W.; Bongers, T.; De Goede, R.G.; Freckman, D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera-an outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- Ewald, M.; Glavatska, O.; Ruess, L. Effects of resource manipulation on nematode community structure and metabolic footprints in an arable soil across time and depth. Nematology 2020, 22, 1025–1043. [Google Scholar] [CrossRef]
- Lu, Q.; Liu, T.; Wang, N.; Dou, Z.; Wang, K.; Zuo, Y. A review of soil nematodes as biological indicators for the assessment of soil health. Front. Agr. Sci. Eng. 2020, 7, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Mesa-Valle, C.M.; Garrido-Cardenas, J.A.; Cebrian-Carmona, J.; Talavera, M.; Manzano-Agugliaro, F. Global Research on Plant Nematodes. Agronomy 2020, 10, 1148. [Google Scholar] [CrossRef]
- Ruess, L. Nematode soil faunal analysis of decomposition pathways in different ecosystems. Nematology 2003, 5, 179–181. [Google Scholar] [CrossRef]
- Pausch, J.; Hofmann, S.; Scharroba, A.; Kuzyakov, Y.; Ruess, L. Fluxes of root-derived carbon into the nematode micro-food web of an arable soil. Food Webs 2016, 9, 32–38. [Google Scholar] [CrossRef]
- Nieminen, J.K. Soil animals and ecosystem processes: How much does nutrient cycling explain? Pedobiologia 2008, 51, 367–373. [Google Scholar] [CrossRef]
- Neher, D.A. Ecology of plant and free-living nematodes in Natural and Agricultural Soil. Annu. Rev. Phytopathol. 2010, 48, 371–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Ferris, H.; Bongers, T.; de Goede, R.G.M. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Du Preez, G.; Daneel, M.; De Goede, R.; Du Toit, M.J.; Ferris, H.; Fourie, H.; Geisen, S.; Kakouli-Duarte, T.; Korthals, G.; Sánchez-Moreno, S.; et al. Nematode-based indices in soil ecology: Application, utility, and future directions. Soil Biol. Biochem. 2022, 169, 108640. [Google Scholar] [CrossRef]
- Ferris, H. Form and function: Metabolic footprints of nematodes in the soil food web. Eur. J. Soil Biol. 2010, 46, 97–104. [Google Scholar] [CrossRef]
- Mulder, C.; Maas, R. Unifying the functional diversity in natural and cultivated soils using the overall body-mass distribution of nematodes. BMC Ecol. 2017, 17, 36. [Google Scholar] [CrossRef] [Green Version]
- Melakeberhan, H.; Bonito, G.; Kravchenko, A.N. Application of Nematode Community Analyses-Based Models towards Identifying Sustainable Soil Health Management Outcomes: A Review of the Concepts. Soil Syst. 2021, 5, 32. [Google Scholar] [CrossRef]
- Ferris, H.; Tuomisto, H. Unearthing the role of biological diversity in soil health. Soil Biol. Biochem. 2015, 85, 101–109. [Google Scholar] [CrossRef]
- Lazarova, S.; Coyne, D.G.; Rodríguez, M.; Peteira, B.; Ciancio, A. Functional Diversity of Soil Nematodes in Relation to the Impact of Agriculture—A Review. Diversity 2021, 13, 64. [Google Scholar] [CrossRef]
- Morais, J.W.; Oliveira, F.L.; Braga, R.F.; Korasaki, V. Mesofauna. In O Ecossistema Solo: Componentes, Relações Ecológicas e Efeitos na Produção Vegetal; Moreira, F.M.S., Cares, J.E., Zanetti, R., Stürmer, S.L., Eds.; Universidade Federal de Lavras: Lavras, Brazil, 2013; pp. 183–200. [Google Scholar]
- Coleman, D.C.; Crossley, D.A., Jr.; Hendrix, P.F. Secondary production: Activities of heterotrophic organisms—the soil fauna. In Fundamentals of Soil Ecology; Coleman, D.C., Crossley, D.A., Jr., Hendrix, P.F., Eds.; Academic Press: Burlington, VT, USA, 2004; pp. 79–185. ISBN 978-0-12-179726-3. [Google Scholar]
- Coleman, D.C.; Callaham, M.A.; Crossley, D.A. Chapter 4 - Secondary Production: Activities of Heterotrophic Organisms—The Soil Fauna. In Fundamentals of Soil Ecology, 3rd ed.; Coleman, D.C., Callaham, M.A., Crossley, D.A., Eds.; Academic Press: Burlington, VT, USA, 2018; pp. 77–171. ISBN 978-0-12-805251-8. [Google Scholar]
- Walter, D.E.; Proctor, H.C. Mites: Ecology, Evolution & Behaviour. Life at a microscale, 2nd ed.; Springer: Dordrecht, The Netherlands, 2013; ISBN 978-94-007-7163-5. [Google Scholar]
- Nielsen, U.N. Soil Fauna Assemblages: Global to Local Scales, 1st ed.; Cambridge University Press: Cambridge, UK, 2019; ISBN 978-1-108-12351-8. [Google Scholar]
- Lavelle, P.; Spain, A. Soil Ecology; Dordrecht Kluwer Academic Publishers: Boston, MA, USA, 2001; ISBN 978-0-7923-7123-6. [Google Scholar]
- Sánchez-Moreno, S.; Nicola, N.L.; Ferris, H.; Zalom, F.G. Effects of agricultural management on nematode-mite assemblages: Soil food web indices as predictors of mite community composition. Appl. Soil Ecol. 2009, 41, 107–117. [Google Scholar] [CrossRef]
- Ruf, A.; Beck, L. The use of predatory soil mites in ecological soil classification and assessment concepts, with perspectives for oribatid mites. Ecotoxicol. Environ. Saf. 2005, 62, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Gergócs, V.; Hufnagel, L. Application of oribatid mites as indicators. Appl. Ecol. Environ. Res. 2009, 7, 79–98. [Google Scholar] [CrossRef]
- Ruf, A. A maturity index for predatory soil mites (Mesostigmata: Gamasina) as an indicator of environmental impacts of pollution on forest soils. Appl. Soil Ecol. 1998, 9, 447–452. [Google Scholar] [CrossRef]
- Ruf, A.; Beck, L.; Dreher, P.; Hund-Rinke, K.; Römbke, J.; Spelda, J. A biological classification concept for the assessment of soil quality: “Biological soil classification scheme” (BBSK). Agric. Ecosyst. Environ. 2003, 98, 263–271. [Google Scholar] [CrossRef]
- Bedano, J.C.; Ruf, A. Sensitivity of different taxonomic levels of soil Gamasina to land use and anthropogenic disturbances. Agric. For. Entomol. 2010, 12, 203–212. [Google Scholar] [CrossRef]
- Krantz, G.W.; Ainscough, B.D. Acarina: Mesostigmata (Gamasida). In Soil Biology Guide; Dindal, D.L., Ed.; A Wiley-Interscience Publication: New York, NY, USA, 1990; pp. 583–666. ISBN 0-471-04551-9. [Google Scholar]
- Koehler, H.H. Mesostigmata (Gamasina, Uropodina), efficient predators in agroecosystems. Agric. Ecosyst. Environ. 1997, 62, 105–117. [Google Scholar] [CrossRef]
- Köhler, H.H. Predatory mites (Gamasina, Mesostigmata). Agric. Ecosyst. Environ. 1999, 74, 395–410. [Google Scholar] [CrossRef]
- Walter, D.E.; Ikonen, E.K. Species, guilds and functional groups: Taxonomy and behaviour in nematophagous arthropods. J. Nematol. 1989, 21, 315–327. [Google Scholar]
- Norton, R.A. Acarina: Oribatida. In Soil Biology Guide; Dindal, D.L., Ed.; A Wiley-Interscience Publication: New York, NY, USA, 1990; pp. 779–803. ISBN 0-471-04551-9. [Google Scholar]
- Maraun, M.; Visser, S.; Scheu, S. Oribatid mites enhance the recovery of the microbial community after a strong disturbance. Appl. Soil Ecol. 1998, 9, 175–181. [Google Scholar] [CrossRef]
- Arroyo, J.; Iturrondobeitia, J.C. Differences in the diversity of oribatid mite communities in forests and agrosystems lands. Eur. J. Soil Biol. 2006, 42, 259–269. [Google Scholar] [CrossRef]
- Minor, M.A.; Cianciolo, J.M. Diversity of soil mites (Acari: Oribatida, Mesostigmata) along a gradient of land use types in New York. Appl. Soil Ecol. 2007, 35, 140–153. [Google Scholar] [CrossRef]
- Illig, J.; Schatz, H.; Scheu, S.; Maraun, M. Decomposition and colonization by micro-arthropods of two litter types in a tropical montane rain forest in southern Ecuador. J. Trop. Ecol. 2008, 24, 157–167. [Google Scholar] [CrossRef]
- Kardol, P.; Newton, J.S.; Bezemer, T.M.; Maraun, M.; van der Putten, W.H. Contrasting diversity patterns of soil mites and nematodes in secondary succession. Acta Oecologica 2009, 35, 603–609. [Google Scholar] [CrossRef]
- Nielsen, U.N.; Osler, G.H.R.; Campbell, C.D.; Burslem, D.F.R.P.; van der Wal, R. The influence of vegetation type, soil properties and precipitation on the composition of soil mite and microbial communities at the landscape scale. J. Biogeogr. 2010, 37, 1317–1328. [Google Scholar] [CrossRef]
- Norton, R.A.; Behan-Pelletier, V.M. Order Oribatida. In A Manual of Acarology; Krantz, G.W., Walter, D.E., Eds.; Texas Tech University Press: Lubbock, TX, USA, 2009; pp. 421–564. [Google Scholar]
- Gerson, U.; Smiley, R.L.; Ochoa, R. Mites (Acari) for pest control; Blackwell Science Ltd.: Oxford, UK, 2003; ISBN 0-632-05658-4. [Google Scholar]
- Lindquist, E.E.; Krantz, G.W.; Walter, D.E. Classification. In A Manual of Acarology; Krantz, G.W., Walter, D.E., Eds.; Texas Tech University Press: Lubbock, TX, USA, 2009; pp. 97–103. [Google Scholar]
- Hajihassani, A. Chemical Nematicides for Control of Plant-Parasitic Nematodes in Georgia Vegetable Crops 2018. Available online: https://extension.uga.edu/publications/detail.html?number=B1502#:~:text=The%20most%20common%20fumigants%20used,(dimethyl%20disulfide%20or%20DMDS) (accessed on 5 July 2022).
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for Management of Soilborne Diseases in Crop Production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Erktan, A.; Or, D.; Scheu, S. The physical structure of soil: Determinant and consequence of trophic interactions. Soil Biol. Biochem. 2020, 148, 107876. [Google Scholar] [CrossRef]
- Hijbeek, R.; Cormont, A.; Hazeu, G.; Bechini, L.; Zavattaro, L.; Janssen, B.; Werner, M.; Schlatter, N.; Guzmán, G.; Bijttebier, J.; et al. Do farmers perceive a deficiency of soil organic matter? A European and farm level analysis. Ecol. Indic. 2017, 83, 390–403. [Google Scholar] [CrossRef]
- Loomans, A.J.M. Every generalist biological control agent requires a special risk assessment. BioControl 2021, 66, 23–35. [Google Scholar] [CrossRef]
- Young, M.R.; Moraza, M.L.; Ueckermann, E.; Heylen, D.; Baardsen, L.F.; Lima-Barbero, J.F.; Gal, S.; Gavish-Regev, E.; Gottlieb, Y.; Roy, L.; et al. Linking morphological and molecular taxonomy for the identification of poultry house, soil, and nest dwelling mites in the Western Palearctic. Sci. Rep. 2019, 9, 5784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrascosa, M.; Sánchez-Moreno, S.; Alonso-Prados, J.L. Relationships between nematode diversity, plant biomass, nutrient cycling and soil suppressiveness in fumigated soils. Eur. J. Soil Biol. 2014, 62, 49–59. [Google Scholar] [CrossRef]
- Gunstone, T.; Cornelisse, T.; Klein, K.; Dubey, A.; Donley, N. Pesticides and Soil Invertebrates: A Hazard Assessment. Front. Environ. Sci. 2021, 9, 122. [Google Scholar] [CrossRef]
- European Food Safety Authority. Administrative guidance on submission of dossiers and assessment reports for the peer-review of pesticide active substances and on the maximum residue level (MRL) application procedure. EFSA J. 2021, 18, EN-6464. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Restrictions and Bans; FAO headquarters, United Nations: Rome, Italy, 2021. [Google Scholar]
- United States Environmental Protection Agency Reregistration and Other Review Programs Predating Pesticide Registration Review 2021. Available online: https://www.epa.gov/pesticide-reevaluation/reregistration-and-other-review-programs-predating-pesticide-registration (accessed on 2 August 2022).
- Mahmood, I.; Imadi, S.R.; Shazadi, K.; Gul, A.; Hakeem, K.R. Effects of Pesticides on Environment. In Plant, Soil and Microbes: Volume 1: Implications in Crop Science; Hakeem, K.R., Akhtar, M.S., Abdullah, S.N.A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 253–269. ISBN 978-3-319-27455-3. [Google Scholar]
- Schaub, S.; Huber, R.; Finger, R. Tracking societal concerns on pesticides – a Google Trends analysis. Environ. Res. Lett. 2020, 15, 084049. [Google Scholar] [CrossRef]
- European Commission; Directorate-General for Research and Innovation; Veerman, C.; Pinto Correia, T.; Bastioli, C.; Biro, B.; Bouma, J.; Cienciala, E.; Emmett, B.; Frison, E.; et al. Caring for Soil Is Caring for Life: Ensure 75% of Soils are Healthy by 2030 for Healthy Food, People, Nature and Climate: Interim Report of the Mission Board for Soil Health and Food; Publications Office: Brussels, Belgium, 2020. [Google Scholar]
- Geisen, S.; Wall, D.H.; van der Putten, W.H. Challenges and Opportunities for Soil Biodiversity in the Anthropocene. Curr. Biol. 2019, 29, R1036–R1044. [Google Scholar] [CrossRef]
- McSorley, R.; Seal, D.R.; Klassen, W. Non-target effects of sunn hemp and marigold cover crops on the soil invertebrate community. Nematropica 2009, 39, 11. [Google Scholar]
- Wang, K.-H.; Radovich, T.; Pant, A.; Cheng, Z. Integration of cover crops and vermicompost tea for soil and plant health management in a short-term vegetable cropping system. Appl. Soil Ecol. 2014, 82, 26–37. [Google Scholar] [CrossRef]
- Leroy, B.L.M.M.; Bommele, L.; Reheul, D.; Moens, M.; De Neve, S. The application of vegetable, fruit and garden waste (VFG) compost in addition to cattle slurry in a silage maize monoculture: Effects on soil fauna and yield. Eur. J. Soil Biol. 2007, 43, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Han, X.; Hu, C.; Chen, J.; Zhang, D.; Steinberger, Y. Changes in the abundance and structure of a soil mite (Acari) community under long-term organic and chemical fertilizer treatments. Appl. Soil Ecol. 2011, 49, 131–138. [Google Scholar] [CrossRef]
- Rueda-Ramírez, D.; Santos, J.C.; Young, M.R.; Mowery, J.; Bauchan, G.; Ochoa, R.; Palevsky, E. In memory of Gary Bauchan: Utilizing an integrated taxonomy approach for the description of a new species of Gamasellodes (Mesostigmata: Ascidae). Syst. Appl. Acarol. 2022, 27, 165–180. [Google Scholar] [CrossRef]
- Rueda-Ramírez, D.; Carta, L.; Mowery, J.; Bauchan, G.; Ochoa, R.; Young, M.; Santos, J.C.; Palevsky, E. In memory of Gary Bauchan: Integrated taxonomy of soil mites in farming systems. Syst. Appl. Acarol. 2022, 27, 181–208. [Google Scholar] [CrossRef]
- van Lenteren, J.C. The state of commercial augmentative biological control: Plenty of natural enemies, but a frustrating lack of uptake. BioControl 2011, 57, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Knapp, M.; van Houten, Y.; van Baal, E.; Groot, T. Use of predatory mites in commercial biocontrol: Current status and future prospects. Acarologia 2018, 58, 7282. [Google Scholar] [CrossRef]
- de Moraes, G.J.; Venancio, R.; dos Santos, V.L.V.; Paschoal, A.D. Potential of Ascidae, Blattisociidae and Melicharidae (Acari: Mesostigmata) as biological control agents of pest organisms. In Prospects for biological control of plant feeding mites and other harmful organisms; Carrillo, D., de Moraes, G.J., Peña, J.E., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 33–75. ISBN 978-3-319-15042-0. [Google Scholar]
- Walter, D.E.; Kaplan, D.T. A guild of thelytokous mites associated with citrus roots in Florida. Environ. Entomol. 1990, 19, 1338–1343. [Google Scholar]
- Rockett, C.L.; Woodring, J.P. Biological investigations on a new species of Ceratozetes and of Pergalumna (Acarina: Cryptostigmata). Acarologia 1966, VIII, 511–520. [Google Scholar]
- Rockett, C.L.; Woodring, J.P. Oribatid mites as predators of soil nematodes. Ann. Entomol. Soc. Am. 1966, 59, 669–671. [Google Scholar] [CrossRef]
- Walter, D.E. Belowground arthropods of semiarid grasslands. In Integrated pest management on rangeland: A shortgrass prairie perspective; Capinera, J.L., Ed.; Westview Press: Boulder, CO, USA, 1987; pp. 271–290. [Google Scholar]
- Walter, D.E. Trophic Behavior of “Mycophagous” Microarthropods. Ecology 1987, 68, 226–229. [Google Scholar] [CrossRef]
- Elmoghazy, M.; El-Kawas, H.; Salman, M. Biological Aspects of Scheloribates laevigatus (Acari: Oribatida) when fed on mixture of the free-living nematode, Eudiplogaster phlagemcaudatus and potato in the Laboratory. Acarines J. Egypt. Soc. Acarol. 2012, 6, 21–24. [Google Scholar] [CrossRef]
- Heidemann, K.; Scheu, S.; Ruess, L.; Maraun, M. Molecular detection of nematode predation and scavenging in oribatid mites: Laboratory and field experiments. Soil Biol. Biochem. 2011, 43, 2229–2236. [Google Scholar] [CrossRef]
- Heidemann, K.; Ruess, L.; Scheu, S.; Maraun, M. Nematode consumption by mite communities varies in different forest microhabitats as indicated by molecular gut content analysis. Exp. Appl. Acarol. 2014, 64, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Heidemann, K.; Hennies, A.; Schakowske, J.; Blumenberg, L.; Ruess, L.; Scheu, S.; Maraun, M. Free-living nematodes as prey for higher trophic levels of forest soil food webs. Oikos 2014, 123, 1199–1211. [Google Scholar] [CrossRef]
- Karg, W. Untersuchungen über die Veränderungen und Wechselbeziehungen der Microarthropoden in Kartoffelnematoden verseuchten Flächen. Nachr. Des Dtsch. Pflanzenschutzd. 1962, 16, 187–195. [Google Scholar]
- Sturhan, D.; Hampel, G. Pflanzenparasitische Nematoden als Beute der Wurzelmilbe Rhizoglyphus echinopus (Acarina Tyroglyphidae). Anz. Schadlingskde. Pflanzenschutz Umweltschutz 1977, 50, 115–118. [Google Scholar] [CrossRef]
- Stirling, G.R. Surrounding the swollen roots of sweetpotato with a decomposing band of an organic amendment enhances nematode-suppressive services and reduces damage caused by root-knot nematode. Australas. Plant Pathol. 2021, 50, 151–168. [Google Scholar] [CrossRef]
- Palevsky, E.; Konopická, J.; Rueda-Ramírez, D.; Zemek, R. A Review of prospective biocontrol agents and sustainable soil practices for bulb mite (Acari: Acaridae) management. Agronomy 2022, 12, 1491. [Google Scholar] [CrossRef]
- Walter, D.E.; Hudgens, R.A.; Freckman, D.W. Consumption of nematodes by fungivorous mites, Tyrophagus spp. (Acarina: Astigmata: Acaridae). Oecologia 1986, 70, 357–361. [Google Scholar] [CrossRef]
- Bilgrami, A.L. Predatory behaviour of a nematode feeding mite Tyrophagus putrescentiae (Sarcoptiformes: Acaridae). Fundam. Appl. Nematol. 1994, 17, 293–297. [Google Scholar]
- Walia, K.K.; Mathur, S. Predatory behavior of two nematophagous mites, Tyrophagus putrescentiae and Hypoaspis calcuttaensis, on root-knot nematode, Meloidogyne javanica. Nematol. Mediterr. 1995, 23, 255–261. [Google Scholar]
- Abou El-Atta, D.A.E.-M.; Osman, M.A. Development and reproductive potential of Tyrophagus putrescentiae (Acari: Acaridae) on plant-parasitic nematodes and artificial diets. Exp. Appl. Acarol. 2016, 68, 477–483. [Google Scholar] [CrossRef]
- Abou El-Atta, D.A.E.-M.; Habashy, M.G.; Mesbah, A.E.; Tawfik, A.A. Life History of Caloglyphus manure, Sancassania (Caloglyphus) berlesei and Tyroghagus putrescentiae (Acari:Acaridae) Feeding on Root-Knot Nematodes, Meloidogyne incognita. J. Plant Prot. Pathol. 2017, 8, 69–72. [Google Scholar] [CrossRef]
- Epsky, N.D.; Walter, D.E.; Capinera, J.L. Potential Role of Nematophagous Microarthropods as Biotic Mortality Factors of Entomogenous Nematodes (Rhabditida: Steinernematidae, Heterorhabditidae). J. Econ. Entomol. 1988, 81, 821–825. [Google Scholar] [CrossRef]
- Karagoz, M.; Gulcu, B.; Cakmak, I.; Kaya, H.K.; Hazir, S. Predation of entomopathogenic nematodes by Sancassania sp. (Acari: Acaridae). Exp. Appl. Acarol. 2007, 43, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Eraky, S.A.; Osman, M.A. Some biological aspects and life table parameters of Caloglyphus manuri Eraky & Osman (Acaridida: Acaridae) fed on different kinds of food. ACARINES J. Egypt. Soc. Acarol. 2008, 2, 45–48. [Google Scholar]
- Muraoka, M.; Ishibashi, N. Nematode-feeding mites and their feeding behavior. Appl. Entomol. Zool. 1976, 11, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Walter, D.E.; Kaplan, D.T. Observations on Coleoscirus simplex (Acarina: Prostigmata), a predatory mite that colonizes greenhouse cultures of rootknot nematode (Meloidogyne spp.), and a review of feeding behavior in the Cunaxidae. Exp. Appl. Acarol. 1991, 12, 47–59. [Google Scholar] [CrossRef]
- Walter, D.E.; Kaplan, D.T.; Davis, E.L. Colonization of greenhouse nematode cultures by nematophagous mites and fungi. Suppl. J. Nematol. 1993, 25, 789–794. [Google Scholar]
- Khalil, A. Biological aspects of the cunaxid mite, Pulaeus pseudominutus when fed on different diets at different temperatures. Egypt. Acad. J. Biol. Sciences. A Entomol. 2016, 9, 49–55. [Google Scholar] [CrossRef]
- Mostafa, M.; Abdel-Rahman, A.; Younis, A.; Yassin, E.; Saber, R. Life History of the Predaceous Mite Cunaxa capreolus (Berlese) (Acari: Prostigmata: Cunaxidae) When Fed on Different Diets at Different Temperatures. Egypt. Acad. J. Biol. Sciences. A Entomol. 2016, 9, 1–6. [Google Scholar] [CrossRef]
- Al-Azzazy, M.M.; Al-Rehiayani, S.M. The soil mite Cunaxa capreolus (Acari: Cunaxidae) as a predator of the root-knot nematode, Meloidogyne incognita and the citrus Nematode, Tylenchulus semipenetrans: Implications for biological control. Acarologia 2022, 62, 174–185. [Google Scholar] [CrossRef]
- Walter, D.E. Predation and mycophagy by endeostigmatid mites (Acariformes: Prostigmata). Exp. Appl. Acarol. 1988, 4, 159–166. [Google Scholar] [CrossRef]
- Amin, A.W.; Mowafe, M.H.; Fatma, S.A. Effect of predaceous mesostigmatid mites in the control of Meloidogyne javanica root-knot nematode on kidney bean. Pak. J. Nematol. 1999, 17, 91–96. [Google Scholar]
- El-Banhawy, E.M.; Osman, H.A.; El-Sawaf, B.M.; Afia, S.I. Interactions of soil predacious mites and citrus nematodes (parasitic and saprophytic), in citrus orchard under different regime of fertilizers. Effect on the population densities and citrus yield. Anz. Schadlingskde. Pflanzenschutz Umweltschutz 1997, 70, 20–23. [Google Scholar] [CrossRef]
- Xu, C.-L.; Chen, Y.-L.; Xu, X.-N.; Wang, D.-W.; Xie, H.; Wang, E.-D.; Li, D.-S.; Zhang, B.-X.; Qin, H.-G. Evaluation of Blattisocius dolichus (Acari: Blattisociidae) for biocontrol of root-knot nematode, Meloidogyne incognita (Tylenchida: Heteroderidae). BioControl 2014, 59, 617–624. [Google Scholar] [CrossRef]
- de Azevedo, L.H.; Emberson, R.M.; de Esteca, F.C.N.; de Moraes, G.J. Macrochelid mites (Mesostigmata: Macrochelidae) as biological control agents. In Prospects for Biological Control of Plant Feeding Mites and Other Harmful Organisms; Carrillo, D., de Moraes, G.J., Peña, J.E., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 103–132. ISBN 978-3-319-15042-0. [Google Scholar]
- McMurtry, J.A.; Sourassou, N.F.; Demite, P.R. The Phytoseiidae (Acari: Mesostigmata) as Biological Control Agents. In Prospects for Biological Control of Plant Feeding Mites and Other Harmful Organisms; Carrillo, D., de Moraes, G.J., Peña, J.E., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 133–149. ISBN 978-3-319-15042-0. [Google Scholar]
- Moreira, G.F.; Moraes, G.J. The potential of free-living laelapid mites (Mesostigmata: Laelapidae) as biological control agents. In Prospects for Biological Control of Plant Feeding Mites and Other Harmful Organisms; Carrillo, D., de Moraes, G.J., Peña, J.E., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 77–102. ISBN 978-3-319-15042-0. [Google Scholar]
- Saber, S.A.; EI-Laithy, A.Y.M.; Korayem, A.M. Biology of the predaceous mite Protogamasellus denticus Nasr (Mesostigmata: Ascidae) feeding on different nematode diets. Int. J. Nematol. 2007, 17, 17–20. [Google Scholar]
- Manwaring, M.; Nahrung, H.F.; Wallace, H. Attack rate and prey preference of Lasioseius subterraneous and Protogamasellus mica on four nematode species. Exp. Appl. Acarol. 2020, 80, 29–41. [Google Scholar] [CrossRef]
- Walter, D.E.; Hunt, H.W.; Elliott, E.T. The influence of prey type on the development and reproduction of some predatory soil mites. Pedobiologia 1987, 30, 419–424. [Google Scholar]
- Binns, E.S. Notes on the biology of Arctoseius cetratus (Sellnick) (Mesostigmata: Ascidae). Acarologia 1974, XVI, 577–582. [Google Scholar]
- Afifi, A.M.; Hassan, M.F.; Nawar, M.S. Notes on the biology feeding habits of Protogamasellus minutus Hafez, El-Badry & Nasr (Acari: Gamasida: Ascidae). Bull. De La Société Entomol. D’égypte 1986, 66, 251–259. [Google Scholar]
- Nawar, M.S.; Nasr, A.K. Biology of the ascid mite Protogamasellus primitivus similis Genis Loots and Ryke with description of immature stages (Acari: Mesostigmata: Ascidae). Bull.-Zool. Soc. Egypt 1989, 68, 85–94. [Google Scholar]
- Walter, D.E.; Lindquist, E.E. The distributions of parthenogenetic ascid mites (Acari: Parasitiformes) do not support the biotic uncertainty hypothesis. Exp. Appl. Acarol. 1995, 19, 423–442. [Google Scholar]
- Stirling, G.R.; Stirling, A.M.; Walter, D.E. The Mesostigmatid mite Protogamasellus mica, an effective predator of free-living and plant-parasitic nematodes. J. Nematol. 2017, 49, 327–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Moraes, G.J.; Moreira, G.F.; Freire, R.A.P.; Beaulieu, F.; Klompen, H.; Halliday, B. Catalogue of the free-living and arthropod-associated Laelapidae Canestrini (Acari: Mesostigmata), with revised generic concepts and a key to genera. Zootaxa 2022, 5184, 001–509. [Google Scholar] [CrossRef]
- Bilgrami, A.L. Evaluation of the predation abilities of the mite Hypoaspis calcuttaensis, predaceous on plant and soil nematodes. Fundam. Appl. Nematol. 1997, 20, 96–98. [Google Scholar]
- Sharma, R.D. Studies of the parasitic nematode Tylenchorhynchus dubius. Meded. Van De Fac. Landbouwwet. 1971, 71, 1–154. [Google Scholar]
- Van de Bund, C.F. Some observations on predatory action of mites on nematodes. Zesz. Probl. Postep. Nauk. Rol. 1972, 129, 103–110. [Google Scholar]
- Salehi, A.; Ostovan, H.; Modarresi, M. Evaluation of the efficiency of Gaeolaelaps aculeifer in control of plant parasitic nematode Tylenchulus semipenetrans under greenhouse conditions. J. Entomol. Nematol. 2014, 6, 150–153. [Google Scholar]
- Yang, S.-H.; Wang, D.; Chen, C.; Xu, C.-L.; Xie, H. Evaluation of Stratiolaelaps scimitus (Acari: Laelapidae) for controlling the root-knot nematode, Meloidogyne incognita (Tylenchida: Heteroderidae). Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Afifi, A.M.; Van Der Geest, L.P.S. Notes on the development and biology of the predaceous soil mite Cosmolaelaps claviger (Berlese 1883) (Gamasida: Laelapidae). In Acarology VI; Griffiths, D.A., Bowman, C.E., Eds.; Ellis Horwood Limited Publishers: West Sussex, UK, 1984; pp. 585–590. [Google Scholar]
- Moreira, G.F.; de Morais, M.R.; Busoli, A.C.; Moraes, G.J. de Life cycle of Cosmolaelaps jaboticabalensis (Acari: Mesostigmata: Laelapidae) on Frankliniella occidentalis (Thysanoptera: Thripidae) and two factitious food sources. Exp. Appl. Acarol. 2015, 65, 219–226. [Google Scholar] [CrossRef]
- Al Rehiayani, S.M.; Fouly, A.H. Cosmolaelaps simplex (Berlese), a Polyphagous Predatory Mite Feeding on Root-knot Nematode Meloidogyne javanica and Citrus Nematode Tylenchulus semipenetrans. Pak. J. Biol. Sci. 2005, 8, 168–174. [Google Scholar] [CrossRef]
- Karg, W. Über die Beziehungen von edaphischen Raubmilben zur Arthropoden- und Nematoden fauna des Bodens. In Tagungsberichte Nr. 45. Bericht über die 9. Wanderversammlung Deutscher Entomologen; Deutche Akademie der Landwirtschaftswissenschaften zu Berlin: Berlin, Germany, 1962; pp. 311–327. [Google Scholar]
- Murphy, P.W.; Sardar, M.A. Resource allocation and utilization contrasts in Hypoaspis aculeifer (Can.) and Alliphis halleri (G. & R. Can.) (Mesostigmata) with emphasis on food source. In The Acari. Reproduction, development and life-history strategies; Schuster, R., Murphy, P.W., Eds.; Chapman & Hall Publications: New York, NY, USA, 1991; pp. 301–311. [Google Scholar]
- Heckmann, L.-H.; Ruf, A.; Nienstedt, K.M.; Krogh, P.H. Reproductive performance of the generalist predator Hypoaspis aculeifer (Acari: Gamasida) when foraging on different invertebrate prey. Appl. Soil Ecol. 2007, 36, 130–135. [Google Scholar] [CrossRef]
- Walter, D.E.; Oliver, J.H. Geolaelaps oreithyiae, n. sp. (Acari, Laelapidae), a thelytokous predator of arthropods and nematodes, and a discussion of clonal reproduction in the Mesostigmata. Acarologia 1989, 30, 293–303. [Google Scholar]
- Berndt, O.; Meyhöfer, R.; Poehling, H.M. Propensity towards cannibalism among Hypoaspis aculeifer and H. miles, two soil-dwelling predatory mite species. Exp. Appl. Acarol. 2003, 31, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Filipponi, A.; Dojmi di Delupis, G. Sul regime dietetico di alcuni macrochelidi (Acari: Mesostigmata), associati in natura a muscidi di interesse sanitario. Riv. Di Parassitol. 1963, XXIV, 277–287. [Google Scholar]
- Filipponi, A.; Petrelli, M.G. Autoecologia e capacita moltiplicativa di Macrocheles muscadomesticae (Scopoli) (Acari: Mesostigmata). Riv. Di Parassitol. 1967, XXVIII, 129–156. [Google Scholar]
- Karg, W. Ökologische Untersuchungen an Milben aus Komposterden im Freiland und unter Glas besonders im Hinblick auf die Uroobovella marginata C.L. Koch. Arch. Für Pflanzenschutz 1968, 4, 93–122. [Google Scholar] [CrossRef]
- Afifi, A.M. Laboratory studies on the biology and feeding habits of three Macrochelid mites (Acari: Gamasida: Macrochelidae). Bull. De La Société Entomol. D’égypte 1989, 68, 169–174. [Google Scholar]
- Ito, Y. The effects of nematode feeding on the predatory efficiency for house fly eggs and reproduction rate of Macrocheles muscaedomesticae (Acarina: Mesostigmata). Jpn. Soc. Med. Entomol. Zool. 1973, 23, 209–213. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, L.H.; Ferreira, M.P.; de Castilho, R.C.; Cançado, P.H.D.; de Moraes, G.J. Potential of Macrocheles species (Acari: Mesostigmata: Macrochelidae) as control agents of harmful flies (Diptera) and biology of Macrocheles embersoni Azevedo, Castilho and Berto on Stomoxys calcitrans (L.) and Musca domestica. Biol. Control. 2018, 123, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, J.G.; Wade, C.F.; Wells, C.N. Nematodes as a natural food for Macrocheles muscaedomesticae (Acarina: Macrochelidae), a predator of the house fly egg. Ann. Entomol. Soc. Am. 1962, 55, 507–511. [Google Scholar] [CrossRef]
- Axtell, R.C. Macrochelidae (Acarina: Mesostigmata) as Biological Control Agents for Synanthropic Flies. In Proceedings of the 2nd International Congress of Acarology, Sutton Bonington, UK, 19–25 July 1969; pp. 401–416. [Google Scholar]
- Ito, Y. Predatory activity of mesostigmatid mites (Acarina: Mesostigmata) for house fly eggs and larvae under feeding of nematodes. Med. Entomol. Zool. 1977, 28, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.C. Rhodacaridae (Acari: Mesostigmata) from near Adelaide, Australia iii. Behaviour and development. Acarologia 1974, XVI, 21–44. [Google Scholar]
- Beaulieu, F.; Walter, D.E. Predation in suspended and forest floor soils: Observations on Australian mesostigmatic mites. Acarologia 2007, XLVII, 43–54. [Google Scholar]
- Mowafi, M.H. Effect of food type on development and reproduction of mite species Lasioseius africanus Nasr (Acari: Ascidae). Egypt. J. Biol. Pest Control. 2005, 15, 93–95. [Google Scholar]
- Britto, E.P.J.; Gago, E.; de Moraes, G.J. How promising is Lasioseius floridensis as a control agent of Polyphagotarsonemus latus? Exp. Appl. Acarol. 2012, 56, 221–231. [Google Scholar] [CrossRef]
- Abou-Awad, B.A.; Korayem, A.M.; Hassan, M.F.; Abou-Elela, M.A. Life history of the predatory mite Lasioseius athiasae (Acari, Ascidae) on various kinds of food substances: A polypeptide analysis of prey consideration. J. Appl. Entomol. 2001, 125, 125–130. [Google Scholar] [CrossRef]
- Tawfik, A.A.; Mesbah, A.E.; Abou El-Atta, D.A.; Abou Zaid, W.R. Effect of food type on life tables and feeding behavior of the ascid mite, Lasioseius athiasae (Acari: Ascidae). J. Plant Prot. Pathol. 2017, 8, 65–68. [Google Scholar] [CrossRef]
- Chen, Y.L.; Xu, C.L.; Xu, X.N.; Xie, H.; Zhang, B.X.; Qin, H.G.; Zhou, W.Q.; Li, D.S. Evaluation of predation abilities of Blattisocius dolichus (Acari: Blattisociidae) on a plant-parasitic nematode, Radopholus similis (Tylenchida: Pratylenchidae). Exp. Appl. Acarol. 2013, 60, 289–298. [Google Scholar] [CrossRef]
- Szafranek, P.; Lewandowski, M.; Kozak, M. Prey preference and life tables of the predatory mite Parasitus bituberosus (Acari: Parasitidae) when offered various prey combinations. Exp. Appl. Acarol. 2013, 61, 53–67. [Google Scholar] [CrossRef]
- Afifi, A.M.; Nasr, A.K. Biology and feeding habits of the predatory mite, Porrhostaspis lunulata Muller (Acari-Gamsida-Parasitidae). Bull. Fac. Agric. Univ. Cairo 1984, 35, 1741–1746. [Google Scholar]
- Rossini, L.A.C.J.; Prado, T.J.; Ferreira, R.J.; Soares, P.L.M.; de Moraes, G.J.; Castilho, R.C. Suitability of the soybean cyst nematode as prey to Protogamasellopsis zaheri (Acari: Mesostigmata: Rhodacaridae). Biol. Control. 2022, 170, 104905. [Google Scholar] [CrossRef]
- Beaulieu, F.; Dowling, A.P.G.; Klompen, H.; de Moraes, G.J.; Walter, D.E. Superorder Parasitiformes Reuter, 1909. In: Zhang, Z.-Q. (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa 2011, 3148, 123–128. [Google Scholar] [CrossRef]
- Sardar, M.A.; Murphy, P.W. Feeding tests of grassland soil-inhabiting gamasine predators. Acarologia 1987, XXVIII, 117–121. [Google Scholar]
- Royce, L.A.; Krantz, G.W. A new rearing method for nematophagous mites. In Modern Acarology; Dusbábek, F., Bukva, V., Eds.; Academia and SPB Academic Publishing BV: Prague, Czech Republic; The Hague, The Netherlands, 1991; Volume 2, pp. 619–622. [Google Scholar]
- Karg, W. Ökologische Untersuchungen von edaphischen Gamasiden (Acarina, Parasitiformes). Pedobiologia 1961, 1, 77–98. [Google Scholar] [CrossRef]
- Habersaat, U. Die Bedeutung der Bodenraubmilben als Prädatoren von landwirtschaftlichen Schädlingen am Beispiel von Hypoaspis angusta KARG, 1965 (Acari: Gamasina). Ph.D. Thesis, ETH Zurich, Zurich, Switzerland, 1989; p. 87S. [Google Scholar]
- Martikainen, E.; Huhta, V. Interactions between nematodes and predatory mites in raw humus soil: A microcosm experiment. Rev. D’écologie Et De Biol. Du Sol 1990, 27, 13–20. [Google Scholar]
- Ito, Y. Predation by manure-inhabiting mesostigmatids (Acarina: Mesostigmata) on some free-living nematodes. Appl. Entomol. Zool. 1971, 6, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Willis, R.R.; Axtell, R.C. Mite predators of the house fly: A comparison of Fuscuropoda vegetans and Macrocheles muscaedomesticae. J. Econ. Entomol. 1968, 61, 1669–1674. [Google Scholar] [CrossRef]
- Jalil, M.; Rodriguez, J.G. Biology of and Odor Perception by Fuscuropoda vegetans (Acarina: Uropodidae), a Predator of the House Fly. Ann. Entomol. Soc. Am. 1970, 63, 935–938. [Google Scholar] [CrossRef]
- Yeates, G.W. Nematodes in Ecological Webs. In Encyclopedia of Life Science Chichester; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2010; pp. 1–10. ISBN 978-0-470-01590-2. [Google Scholar]
- Lee, Q.; Widden, P. Folsomia candida, a “fungivorous” collembolan, feeds preferentially on nematodes rather than soil fungi. Soil Biol. Biochem. 1996, 28, 689–690. [Google Scholar] [CrossRef]
- Zaitsev, A.S.; Birkhofer, K.; Ekschmitt, K.; Wolters, V. Belowground Tritrophic Food Chain Modulates Soil Respiration in Grasslands. Pedosphere 2018, 28, 114–123. [Google Scholar] [CrossRef]
- Moore, J.C.; McCann, K.; de Ruiter, P.C. Modeling trophic pathways, nutrient cycling, and dynamic stability in soils. Pedobiologia 2005, 49, 499–510. [Google Scholar] [CrossRef]
- Perdomo, G.; Evans, A.; Maraun, M.; Sunnucks, P.; Thompson, R. Mouthpart morphology and trophic position of microarthropods from soils and mosses are strongly correlated. Soil Biol. Biochem. 2012, 53, 56–63. [Google Scholar] [CrossRef]
- Bowman, C.E. Feeding design in free-living mesostigmatid chelicerae (Acari: Anactinotrichida). Exp. Appl. Acarol. 2021, 84, 1–119. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.; Migge, S.; Norton, R.A.; Scheu, S.; Langel, R.; Reineking, A.; Maraun, M. Trophic niche differentiation in soil microarthropods (Oribatida, Acari): Evidence from stable isotope ratios (15N/14N). Soil Biol. Biochem. 2004, 36, 1769–1774. [Google Scholar] [CrossRef]
- Díaz-Aguilar, I.; Quideau, S.A. Trophic ecology of mesostigmatan and oribatid mites in harvested and control coniferous and deciduous stands of the boreal mixedwood forest determined using 15N stable isotopes. Soil Biol. Biochem. 2013, 67, 147–154. [Google Scholar] [CrossRef]
- Klarner, B.; Maraun, M.; Scheu, S. Trophic diversity and niche partitioning in a species rich predator guild – Natural variations in stable isotope ratios (13C/12C, 15N/14N) of mesostigmatid mites (Acari, Mesostigmata) from Central European beech forests. Soil Biol. Biochem. 2013, 57, 327–333. [Google Scholar] [CrossRef]
- Traugott, M.; Kamenova, S.; Ruess, L.; Seeber, J.; Plantegenest, M. Chapter Three - Empirically Characterising Trophic Networks: What Emerging DNA-Based Methods, Stable Isotope and Fatty Acid Analyses Can Offer. In Advances in Ecological Research; Woodward, G., Bohan, D.A., Eds.; Academic Press: Burlington, VT, USA, 2013; Volume 49, pp. 177–224. ISBN 0065-2504. [Google Scholar]
- Mcsorley, R.; Wang, K.-H.; Kokalis-Burelle, N.; Church, G. Effects of soil type and steam on nematode biological control potential of the rhizosphere community. Nematropica 2006, 36, 197–214. [Google Scholar]
- Laakso, J.; Setala, H. Sensitivity of Primary Production to Changes in the Architecture of Belowground Food Webs. Oikos 1999, 87, 57. [Google Scholar] [CrossRef]
- Kühn, J.; Ruess, L. Effects of resource quality on the fitness of collembola fed single and mixed diets from the green and brown food chain. Soil Biol. Biochem. 2021, 154, 108156. [Google Scholar] [CrossRef]
- Łukasiewicz, A. Juvenile diet quality and intensity of sexual conflict in the mite Sancassania berlesei. BMC Evol. Biol. 2020, 20, 35. [Google Scholar] [CrossRef]
- Brückner, A.; Schuster, R.; Wehner, K.; Heethoff, M. Nutritional quality modulates trait variability. Front. Zool. 2018, 15, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brückner, A.; Schuster, R.; Wehner, K.; Heethoff, M. Effects of nutritional quality on the reproductive biology of Archegozetes longisetosus (Actinotrichida, Oribatida, Trhypochthoniidae). Soil Org. 2018, 90, 1–12. [Google Scholar]
- Marques, R.V.; Sarmento, R.A.; Lemos, F.; Pedro-Neto, M.; Sabelis, M.W.; Venzon, M.; Pallini, A.; Janssen, A. Active prey mixing as an explanation for polyphagy in predatory arthropods: Synergistic dietary effects on egg production despite a behavioural cost. Funct. Ecol. 2015, 29, 1317–1324. [Google Scholar] [CrossRef]
- Hubert, J.; Nesvorná, M.; Palevsky, E.; Smrz, J. Diseases of Prey Mites Used for Mass Rearing Predatory Mites. In Proceedings of the Acta Horticulturae; International Society for Horticultural Science (ISHS), Leuven, Belgium, 10 July 2014; pp. 177–185. [Google Scholar]
- Delisle, J.F.; Brodeur, J.; Shipp, L. Evaluation of various types of supplemental food for two species of predatory mites, Amblyseius swirskii and Neoseiulus cucumeris (Acari: Phytoseiidae). Exp. Appl. Acarol. 2015, 65, 483–494. [Google Scholar] [CrossRef]
- Scheu, S.; Folger, M. Single and mixed diets in Collembola: Effects on reproduction and stable isotope fractionation. Funct. Ecol. 2004, 18, 94–102. [Google Scholar] [CrossRef]
- Haubert, D.; Langel, R.; Scheu, S.; Ruess, L. Effects of food quality, starvation and life stage on stable isotope fractionation in Collembola. Pedobiologia 2005, 49, 229–237. [Google Scholar] [CrossRef]
- Ruess, L.; Müller-Navarra, D.C. Essential Biomolecules in Food Webs. Front. Ecol. Evol. 2019, 7, 269. [Google Scholar] [CrossRef] [Green Version]
- Gladyshev, M.I.; Sushchik, N.N.; Makhutova, O.N. Production of EPA and DHA in aquatic ecosystems and their transfer to the land. Prostaglandins Other Lipid Mediat. 2013, 107, 117–126. [Google Scholar] [CrossRef]
- Rothstein, M.; Götz, P. Biosynthesis of fatty acids in the free-living nematode, Turbatrix aceti. Arch. Biochem. Biophys. 1968, 126, 131–140. [Google Scholar] [CrossRef]
- Watts, J.L.; Browse, J. Genetic dissection of polyunsaturated fatty acid synthesis in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2002, 99, 5854–5859. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, C.L.; Krusberg, L.R. Investigation of some lipids from Turbatrix aceti. Comp. Biochem. Physiol. Part B Comp. Biochem. 1973, 45, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Menzel, R.; von Chrzanowski, H.; Tonat, T.; van Riswyck, K.; Schliesser, P.; Ruess, L. Presence or absence? Primary structure, regioselectivity and evolution of Δ12/ω3 fatty acid desaturases in nematodes. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2019, 1864, 1194–1205. [Google Scholar] [CrossRef]
- Malcicka, M.; Visser, B.; Ellers, J. An evolutionary perspective on linoleic acid synthesis in animals. Evol. Biol. 2018, 45, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Brückner, A.; Heethoff, M. Fatty acid metabolism in an oribatid mite: De novo biosynthesis and the effect of starvation. Exp. Appl. Acarol. 2020, 81, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, N.; Naito, M.; Mori, N.; Kuwahara, Y. De novo biosynthesis of linoleic acid and its conversion to the hydrocarbon (Z,Z)-6,9-heptadecadiene in the astigmatid mite, Carpoglyphus lactis: Incorporation experiments with 13C-labeled glucose. Insect Biochem. Mol. Biol. 2014, 45, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Raspotnig, G.; Krisper, G. Fatty acids as cuticular surface components in oribatid mites (Acari: Oribatida). Arthropod Biology: Contributions to Morphology, Ecology and Systematics. Biosyst. Ecol. Ser. 1998, 14, 215–243. [Google Scholar]
- Raspotnig, G.; Schuster, R.; Krisper, G.; Fauler, G.; Leis, H.-J. Chemistry of the oil gland secretion of Collohmannia gigantea (Acari: Oribatida). Exp. Appl. Acarol. 2001, 25, 933–946. [Google Scholar] [CrossRef]
- Dmitryjuk, M.; Zalewski, K.; Raczkowski, M.; Żółtowska, K. Composition of fatty acids in the Varroa destructor mites and their hosts, Apis mellifera drone-prepupae. Ann. Parasitol. 2015, 61, 21–26. [Google Scholar]
- Menzel, R.; Geweiler, D.; Sass, A.; Simsek, D.; Ruess, L. Nematodes as important source for Omega-3 Long-Chain fatty acids in the soil food web and the impact in nutrition for higher trophic levels. Front. Ecol. Evol. 2018, 6, 96. [Google Scholar] [CrossRef] [Green Version]
- Stirling, G.R. Biological control of plant-parasitic nematodes: Soil ecosystem management in sustainable agriculture, 2nd ed.; CABI Pub.: Croydon, UK, 2014; ISBN 978-1-78064-415-8. [Google Scholar]
- Eilenberg, J.; Hajek, A.; Lomer, C. Suggestions for unifying the terminology in biological control. Bio. Control. 2001, 46, 387–400. [Google Scholar]
- Barbercheck, M.E. Effect of soil physical factors on biological control agents of soil insect pests. Fla. Entomol. 1992, 75, 539–548. [Google Scholar] [CrossRef]
- Yadav, S.; Patil, J.; Kanwar, R.S. The role of free living nematode population in the organic matter recycling. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 2726–2734. [Google Scholar] [CrossRef]
- Treonis, A.M.; Sutton, K.A.; Unangst, S.K.; Wren, J.E.; Dragan, E.S.; McQueen, J.P. Soil organic matter determines the distribution and abundance of nematodes on alluvial fans in Death Valley, California. Ecosphere 2019, 10, e02659. [Google Scholar] [CrossRef]
- FAO Conservation Agriculture 2022. Available online: https://www.fao.org/publications/card/en/c/CB8350EN/ (accessed on 10 September 2022).
- Venter, Z.S.; Jacobs, K.; Hawkins, H.-J. The impact of crop rotation on soil microbial diversity: A meta-analysis. Pedobiologia 2016, 59, 215–223. [Google Scholar] [CrossRef]
- Abán, C.L.; Verdenelli, R.; Gil, S.V.; Huidobro, D.J.; Meriles, J.M.; Brandan, C.P. Service crops improve a degraded monoculture system by changing common bean rhizospheric soil microbiota and reducing soil-borne fungal diseases. FEMS Microbiol. Ecol. 2021, 97, fiaa258. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Moreno, S.; Cano, M.; López-Pérez, A.; Rey Benayas, J.M. Microfaunal soil food webs in Mediterranean semi-arid agroecosystems. Does organic management improve soil health? Appl. Soil Ecol. 2018, 125, 138–147. [Google Scholar] [CrossRef]
- Wang, K.-H.; Hooks, C.R.R.; Marahatta, S.P. Can using a strip-tilled cover cropping system followed by surface mulch practice enhance organisms higher up in the soil food web hierarchy? Appl. Soil Ecol. 2011, 49, 107–117. [Google Scholar] [CrossRef]
- Zhang, X.; Ferris, H.; Mitchell, J.; Liang, W. Ecosystem services of the soil food web after long-term application of agricultural management practices. Soil Biol. Biochem. 2017, 111, 36–43. [Google Scholar] [CrossRef]
- Udalova, Z.; Ushakova, N.; Butorina, N.; Zinovieva, S. Influence of insectocomposts through Hermetia illucens larvae on nematodes of various ecological-trophic groups. Res. Crops 2021, 22, 150–157. [Google Scholar] [CrossRef]
- Messelink, G.J.; De Kogel, W.J. Impact of chrysanthemum cultivar, fertilization and soil-dwelling predatory mites on Frankliniella occidentalis. Proc. Neth. Entomol. Soc. Meet. 2005, 16, 101–107. [Google Scholar]
- Grosman, A.H.; Messelink, G.J.; Groot, E.B. de Combined use of a mulch layer and the soil-dwelling predatory mite Macrocheles robustulus (Berlese) enhance the biological control of sciarids in potted plants. IOBC/WPRS Bull. 2011, 68, 51–54. [Google Scholar]
- de Esteca, F.C.N.; Rodrigues, L.R.; de Moraes, G.J.; Júnior, I.D.; Klingen, I. Mulching with coffee husk and pulp in strawberry affects edaphic predatory mite and spider mite densities. Exp. Appl. Acarol. 2018, 76, 161–183. [Google Scholar] [CrossRef] [PubMed]
- Young, M.R.; Hebert, P.D.N. Unearthing soil arthropod diversity through DNA metabarcoding. Peer. J. 2022, 10, e12845. [Google Scholar] [CrossRef] [PubMed]
- Schenk, J.; Geisen, S.; Kleinboelting, N.; Traunspurger, W. Metabarcoding data allow for reliable biomass estimates in the most abundant animals on earth. MBMG 2019, 3, e46704. [Google Scholar] [CrossRef]
- Santos, P.F.; Whitford, W.G. The effects of microarthropods on litter decomposition in a Chihuahuan desert ecosystem. Ecology 1981, 62, 654–663. [Google Scholar] [CrossRef]
- Santos, P.F.; Phillips, J.; Whitford, W.G. The role of mites and nematodes in early stages of buried litter decomposition in a desert. Ecology 1981, 62, 664–669. [Google Scholar] [CrossRef]
- Elkins, N.Z.; Whitford, W.G. The role of microarthropods and nematodes in decomposition in a semi-arid ecosystem. Oecologia 1982, 55, 303–310. [Google Scholar] [CrossRef]
- Parker, L.W.; Santos, P.F.; Phillips, J.; Whitford, W.G. Carbon and nitrogen dynamics during the decomposition of litter and roots of a Chihuahuan desert annual, Lepidium lasiocarpum. Ecol. Monogr. 1984, 54, 339–360. [Google Scholar] [CrossRef]
- Hyvönen, R.; Persson, T. Effects of fungivorous and predatory arthropods on nematodes and tardigrades in microcosms with coniferous forest soil. Biol Fertil Soils 1996, 21, 121–127. [Google Scholar] [CrossRef]
- Mcsorley, R.; Wang, K.-H. Possibilities for biological control of root-knot nematodes by natural predators in Florida soils. Proc. Fla. State Hortic. Soc. 2009, 122, 421–425. [Google Scholar]
- Stirling, G.R.; Mankau, R. Biological control of nematode parasited of citrus by natural enemies. Proc. Int. Soc. Citric. 1977, 3, 843–847. [Google Scholar]
- Walter, D.E.; Proctor, H.C. Predatory mites in Tropical Australia: Local species richness complementarity. Biotropica 1998, 30, 72–81. [Google Scholar] [CrossRef]
- El-Banhawy, E.M.; Nasr, A.K.; Afia, S.I. Survey of predacious soil mites (Acari: Mesostigmata) in citrus orchards of the Nile Delta and Middle Egypt with notes on the abundance of the citrus parasitic nematode Tylenchulus semipenetrans (Tylenchida: Tylenchulidae). JTI 2006, 26. [Google Scholar] [CrossRef]
- Lindquist, E.E.; Walter, D.E. Antennoseius ( Vitzthumia ) janus n.sp. (Acari: Ascidae), a mesostigmatic mite exhibiting adult female dimorphism. Can. J. Zool. 1989, 67, 1291–1310. [Google Scholar] [CrossRef]
- Halliday, R.B.; Walter, D.E.; Lindquist, E.E. Revision of the Australian Ascidae (Acarina: Mesostigmata). Invert. Syst. 1998, 12, 1–54. [Google Scholar] [CrossRef]
- Binns, E.S. Arctoseius cetratus (Sellnick) (Acarina: Ascidae) phoretic on mushroom sciarid flies. Acarologia 1972, XIV, 350–356. [Google Scholar]
- Binns, E.S. Three predator mites from mushroom beds. Mushroom J. 1973, 7, 319–320. [Google Scholar]
- Binns, E.S. Predatory mites - neglected allies? Mushroom J. 1973, 12, 540–544. [Google Scholar]
- Karg, W. Acari (Acarina), Milben. Unterordnung Anactinochaeta (Parasitiformes). Die freilebenden Gamasina (Gamasides), Raubmilben. Die Tierwelt Dtschl. 1971, 59, 1–475. [Google Scholar]
- Chaires-Grijalva, M.P.; Estrada-Venegas, E.G.; Equihua-Martinez, A.; Moser, J.C.; Blomquist, S.R. Trophic habits of mesostigmatid mites associated with bark beetles in Mexico. J. Acarol. Soc. Jpn. 2016, 25, S161–S167. [Google Scholar] [CrossRef] [Green Version]
- Rudzińska, M. Life history of the phoretic predatory mite Arctoseius semiscissus (Acari: Ascidae) on a diet of sciarid fly eggs. Exp. Appl. Acarol. 1998, 22, 643–648. [Google Scholar] [CrossRef]
- Walter, D.E. The genus Gamasellodes (Acari: Mesostigmata: Ascidae): New Australian and North American species. Syst. Appl. Acarol. Spec. Publ. 2003, 15, 1–10. [Google Scholar] [CrossRef]
- Walter, D.E. Life history, trophic behavior, and description of Gamasellodes vermivorax n. sp. (Mesostigmata: Ascidae), a predator of nematodes and arthropods in semiarid grassland soils. Can. J. Zool. 1987, 65, 1689–1695. [Google Scholar] [CrossRef]
- Walter, D.E.; Lindquist, E.E. Arrhenoseius gloriosus n. g., n. sp. (Acari: Mesostigmata: Ascidae), an arrhenotokous mite from rainforests in Queensland, Australia. Acarologia 2000, 41, 53–68. [Google Scholar]
- Walter, D.E.; Lindquist, E.E. Life history and behavior of mites in the genus Lasioseius (Acari: Mesostigmata: Ascidae) from grassland soils in Colorado, with taxonomic notes and description of a new species. Can. J. Zool. 1989, 67, 2797–2813. [Google Scholar] [CrossRef]
- Walter, D.E.; Lindquist, E.E. Australian species of Lasioseius (Acari: Mesostigmata: Ascidae): The porulosus group and other species from rainforest canopies. Invert. Syst. 1997, 11, 525. [Google Scholar] [CrossRef]
- Mankau, R.; Imbriani, J.L. Studies on the mite Lasioseius scapulatusa predator on soil nematodes. Nematropica 1978, 8, 15. [Google Scholar]
- Imbriani, J.L.; Mankau, R. Studies on Lasioseius scapulatus, a Mesostigmatid mite predaceous on nematodes. J. Nematol. 1983, 15, 523–528. [Google Scholar]
- Fouly, A.H. Influence of different nourishment on the biology of Lasioseius dentatus (Fox), a new record from Egypt (Acari: Gamasida: Ascidae). Egypt. J. Biol. Pest Control. 1997, 7, 1–6. [Google Scholar]
- Van de Bund, C.F. Mesofauna of arable soil. In Integrated control of pests in the Netherlands; Minks, A.K., Gruys, P., Eds.; Centre for Agricultural Publishing and Documentation: Pudoc, Wageningen, 1980; pp. 87–92. [Google Scholar]
- Aguilar-Marcelino, L.; Quintero-Martínez, M.T.; Mendoza de Gives, P.; López-Arellano, M.E.; Liébano-Hernández, E.; Torres-Hernández, G.; González-Camacho, J.M.; Cid del Prado, I. Evaluation of predation of the mite Lasioseius penicilliger (Aracnida: Mesostigmata) on Haemonchus contortus and bacteria-feeding nematodes. J. Helminthol. 2014, 88, 20–23. [Google Scholar] [CrossRef]
- Nasr, A.K.; Nawar, M.S.; Mowafi, M.H.A. Biological studies and feeding habits of Lastoseius [sic] athiasae Nawar and Nasr (Acari: Mesostigmata: Ascidae) in Egypt. Bull.-Zool. Soc. Egypt 1990, 39. [Google Scholar]
- Cayrol, J.C. Action des autres composants de la biocénose du champignon de couche sur le nématode mycophage, Ditylenchus myceliophagus J. B. Goodey, 1958, et étude de son anabiose: Forme de survie en conditions défavorables. Rev. D’écologie Et De Biol. Du Sol 1970, VII, 409–440. [Google Scholar]
- Karg, W. Verbreitung und Bedeutung Von Raubmilben der Cohors Gamasina als Antagonisten von Nematoden. Pedobiologia 1983, 25, 419–432. [Google Scholar]
- Binns, E.S. Digamasellus fallax Leitner (Mesostigmata: Digamasellidae) phoretic on mushroom sciarid flies. Acarologia 1973, 15, 10–17. [Google Scholar] [PubMed]
- Kinn, D.N. Mutualism Between Dendrolaelaps neodisetus and Dendroctonus frontalis1. Environ. Entomol. 1980, 9, 756–758. [Google Scholar] [CrossRef]
- Abou-El-Soud, A.B.; Shoeib, A.A. Root-knot nematode Meloidogyne javanica and acarid mite, Acarus siro L. predation by digamasellid mite, Dendrolaelaps rasmii Nasr and Mersal. Al-Azhar J. Agric. Res. 2000, 31, 45–50. [Google Scholar]
- Krantz, G.W.; Poinar, G.O., Jr. Mites, nematodes and the multimillion dollar weevil. J. Nat. Hist. 2004, 38, 135–141. [Google Scholar] [CrossRef]
- Kinn, D.N. Notes on the life cycle and habitats of Digamasellus quadrisetus (Mesostigmata: Digamasellidae). Ann. Entomol. Soc. Am. 1967, 60, 862–865. [Google Scholar] [CrossRef]
- Hirschmann, W.; Rühm, W. Ein „Haustier" des Buchdruckers? Mikrokosmos 1955, 44, 234–236. [Google Scholar]
- Enda, N.; Tamura, H. Nematophagous mites found on adults of the Japanese pine sawyer. Shinrin Boecki 1970, 26, 188–190. [Google Scholar]
- Enda, N.; Tamura, H. Nematode feeding mites found on adults of the Japanese pine sawyer. Shinrin Boecki 1980, 29, 17–20. [Google Scholar]
- Walter, D.E. Heatherellidae — a new family of Mesostigmata (Acari: Parasitiformes) based on two new species from rainforest litter in Australia. Int. J. Acarol. 1997, 23, 167–175. [Google Scholar] [CrossRef]
- Inserra, R.N.; Davis, D.W. Hypoaspis nr. aculeifer: A mite predacious on Root-knot and Cyst nematodes. J. Nematol. 1983, 15, 324–325. [Google Scholar] [PubMed]
- Walter, D.E.; Campbell, N.J.H. Exotic vs endemic biocontrol agents: Would the real Stratiolaelaps miles (Berlese) (Acari: Mesostigmata: Laelapidae), please stand up? Biol. Control. 2003, 26, 253–269. [Google Scholar] [CrossRef]
- Brodsgaard, H.F.; Sardar, M.A.; Enkegaard, A. Prey preference of Hypoaspis miles (Berlese) (Acarina: Hypoaspididae): Non-interference with other beneficials in glasshouse crops. IOBC/WPRS Bull. 1996, 19, 23–26. [Google Scholar]
- Singer, G.; Krantz, G.W. The use of nematodes and oligochaetes for rearing predatory mites. Acarologia 1967, IX, 485–487. [Google Scholar]
- Krantz, G.W.; Royce, L.A. Observations on the biology and behavior of Macrocheles mycotrupetes Krantz and Mellott (Acari: Macrochelidae). Int. J. Acarol. 1994, 20, 115–121. [Google Scholar] [CrossRef]
- Manning, M.J.; Halliday, R.B. Biology and reproduction of some australian species of Macrochelidae (Acarina). Aust. Entomol. 1994, 21, 89–94. [Google Scholar]
- Krantz, G.W. A new species of Macrocheles (Acarina: Macrochelidae) associated with bark beetles of the genera Ips and Dendroctonus. J. Kans. Entomol. Soc. 1965, 38, 145–153. [Google Scholar]
- Kinn, D.N.; Witcosky, J.J. The life cycle and behaviour of Macrocheles boudreauxi Krantz. Z. Für Angew. Entomol. 1977, 84, 136–144. [Google Scholar] [CrossRef]
- Cicolani, B. The influence of temperature on the speed of development, from egg to adult, of Macrocheles matrius (Acarina: Mesostigmata). Acarologia 1977, XIX, 384–394. [Google Scholar]
- Filipponi, A.; Cicolani, B. Influenza della temperatura sulla fecondita’ longevita’ e capacita’ moltiplicativa nello intervallo ottimale di Macrocheles matrius (Acarina, Mesostigmata). Riv. Di Parassitol. 1974, XXXV, 291–306. [Google Scholar]
- Singh, P.; Rodriguez, J.G. Food for macrochelid mites (Acarina) by an improved method for mass rearing of a nematode, Rhabditella leptura. Acarologia 1966, VIII, 549–550. [Google Scholar]
- Filipponi, A.; Francaviglia, G. Larviparita Facoltativa in alguni Macrochelidi (Acari: Mesostigmata) associati a muscidi di interesse sanitario. Parassitologia 1964, VI, 291–301. [Google Scholar]
- Geden, C.J.; Axtell, R.C. Predation by Carcinops pumilio (Coleoptera: Histeridae) and Macrocheles muscaedomesticae (Acarina: Macrochelidae) on the house fly (Diptera: Muscidae): Functional response, effects of temperature, and availability of alternative prey. Environ. Entomol. 1988, 17, 739–744. [Google Scholar] [CrossRef] [Green Version]
- Heikal, H.M. Parasitus fimetorum and Macrocheles muscaedomesticae (Acarina:Parasitidae, Macrochelidae) as Natural Predators of the Root Knot Nematode, Meloidogyne javanica Treub. Egypt J. Biol. Pest Control. 2020, 30, 33. [Google Scholar] [CrossRef]
- Ho, C.-C.; Cromroy, H.L.; Patterson, R.S. Mass production of the predaceous mite, Macrocheles muscaedomesticae (Scopoli) (Acarina: Macrochelidae), a predator of the house fly. In Biocontrol of arthropods affecting livestock and poultry; Rutz, D.A., Patterson, R.S., Eds.; Westview Press: Boulder, CO, USA, 1990; pp. 201–213. [Google Scholar]
- Filipponi, A. Influence of Temperature on Population Increase of Four Dung-Living Macrochelid Mites within Optimum Range. In Proceedings of the 3rd International Congress of Acarology, Prague; Rosický B, D.M., Ed.; S. Junk, The Hague and Academia: Prague, Czech Republic, 1971; pp. 775–779. [Google Scholar]
- Filipponi, A.; Mosna, B.; Petrelli, G. L’ottimo di temperatura di Macrocheles muscaedomesticae (Scopoli), come attributo di popolazione. Riv. Di Parassitol. 1971, 32, 193–218. [Google Scholar]
- Costa, M. Notes on macrochelids associated with manure and coprid beetles in Israel. II. Three new species of the Macrocheles pisentii complex, with notes on their biology. Acarologia 1967, IX, 304–329. [Google Scholar]
- Krantz, G.W. Nature of the association between pisentii group mites (Acari, Macrochelidae, Macrocheles) and dung beetles of the genus Scarabaeus (Coleoptera, Scarabaeidae) in Southern France. Acarologia 1991, XXXII, 3–11. [Google Scholar]
- Costa, M. Notes on Macrochelids associated with manure and coprid beetles in Israel. I Macrocheles robustulus (Berlese, 1904). Development and biology. Acarologia 1966, VIII, 532–548. [Google Scholar]
- Nasr, A.K.; Nawar, M.S.; Mowafi, M.H.A. Biological studies on Proctolaelaps bickleyi Bram (Acari: Gamasida: Ascidae). Bull.-Zool. Soc. Egypt 1990, 39, 89–100. [Google Scholar]
- Nawar, M.S. Life tables of Proctolaelaps deleoni Nawar, Childers and Abou-Setta (Gamasida: Ascidae) at different temperatures. Exp. Appl. Acarol. 1992, 13, 281–285. [Google Scholar] [CrossRef]
- Kinn, D.N. The life cycle of Proctolaelaps dendroctoni lindquist and hunter (acari: Ascidae): A mite associated with pine bark beetles. Int. J. Acarol. 1983, 9, 205–210. [Google Scholar] [CrossRef]
- Evans, G.O. A revision of the British Aceosejinae (Acarina: Mesostigmata). Proc. Zool. Soc. Lond. 1958, 131, 177–229. [Google Scholar] [CrossRef]
- Hirschmann, W.; Rühm, W. Milben und Fadenwürmer als Symphonsten und Parasiten des Buchdruckers (Ips typographus). Nachr. Des Nat. Mus. Der Stadt Aschaffenbg. 1954, 43, 41–50. [Google Scholar]
- Kinn, D.N. The life cycle and behaviour of Cercoleipus coleonotus (Acarina: Mesostigmata) including a survey of phoretic mite associates of California Scolytidae. University of California Publications in Entomology; University of California Press: Berkeley, CA, USA, 1971; Volume 65. [Google Scholar]
- Lindquist, E.E.; Hunter, P.E. Some Mites of the Genus Proctolaelaps Berlese (Acarina: Blattisociidae) Associated with Forest Insect Pests. Can. Entomol. 1965, 97, 15–32. [Google Scholar] [CrossRef]
- Ahmed, M.M.; Mowafi, M.H. Life tables of Proctolaelaps pygmaeus (Muller) at different temperatures. Al-Azhar J. Agric. Res. 1998, 27, 338–348. [Google Scholar]
- Metwalli, A.M.; Abbasy, M.R.A.; Montaser, S.A. Life history of the ascid mite Proctolaelaps pygmaeus Muller when fed on two different preys. Al-Azhar J. Agric. Res. 1991, 13, 237–246. [Google Scholar]
- Ibrahim, G.A.; Abdel-Samedi, M.A.; El-Gazzar, H.F. Biological aspects of predator mite Proctolaelaps subcorticalis Lindquist (Ascidae: Mesostigmata) with description to its immature stages. Minufiya J. Agric. Res. 1992, 17, 2015–2023. [Google Scholar]
- Pugh, P.J.A.; King, P.E. Feeding in intertidal Acari. J. Exp. Mar. Biol. Ecol. 1985, 94, 269–280. [Google Scholar] [CrossRef]
- Costa, M. Descriptions of the juvenile stages of Pachylaelaps hispani Berlese (Acari: Mesostigmata). Acarologia 1966, VIII, 9–22. [Google Scholar]
- Ishikawa, K. Taxonomic and ecological studies in the family Parholaspidae from Japan (Part 4). Rep. Res. Matsuyama Shinonome Jr. Col 1980, 11, 183–190. [Google Scholar]
- Lindquist, E.E.; Krantz, G.W.; Walter, D.E. Order Mesostigmata. In A Manual of Acarology; Krantz, G.W., Walter, D.E., Eds.; Texas Tech University Press: Lubbock, TX, USA, 2009; pp. 124–232. [Google Scholar]
- Hse, S.H.H. Biological Studies of Mites Associated with Bark Beetles. M.S. Thesis, Lousiana State University, Baton Rouge, LA, USA, 1964. [Google Scholar]
- Karg, W. Zur Kenntnis der Gattung Parasitus Latreille, 1795 (Acarina, Parasitiformes) aus Komposterden und Gurkenkulturen. Dtsch. Entomol. Z. 1972, 19, 55–63. [Google Scholar] [CrossRef]
- Al-Amidi, A.H.K.; Downes, M.J. Parasitus bituberosus (Acari: Parasitidae), a possible agent for biological control of Heteropeza pygmaea (Diptera: Cecidomyiidae) in mushroom compost. Exp. Appl. Acarol. 1990, 8, 13–25. [Google Scholar] [CrossRef]
- Costa, M. Descriptions of the hitherto unknown stages of Parasitus copridis Costa (Acari: Mesostigmata) with notes on its biology. Zool. J. Linn. Soc. 1964, 45, 209–222. [Google Scholar] [CrossRef]
- Muma, M.H.; Denmark, H.A. Biological Studies on Macroseius biscutatus (Acarina: Phytoseiidae). Fla. Entomol. 1967, 50, 249–255. [Google Scholar] [CrossRef]
- Taha, H.A.; Shereef, G.M.; Soliman, Z.R.; Wafaa, O.G. Effect of different prey types and temperature on biological aspects of predatory mite Protogamasellus discorus (Acari: Gamasida: Ascidae) with special references of chemical analysis of prey. Egypt. J. Plant Prot. Res. Inst. 2019, 2, 480–487. [Google Scholar]
- Castilho, R.C.; de Moraes, G.J.; Silva, E.S.; Silva, L.O. Predation potential and biology of Protogamasellopsis posnaniensis Wisniewski & Hirschmann (Acari: Rhodacaridae). Biol. Control. 2009, 48, 164–167. [Google Scholar] [CrossRef]
- Bunt, J.A. Effect and mode of action of some systemic nematicides; Mededelingen Landbouwhogeschool Wageningen Nederland: Wageningen, The Netherlands, 1975; Volume 75-10. [Google Scholar]
- Walter, D.E.; Proctor, H.C. Feeding behavior and phylogeny: Observations on early derivative Acari. Exp. Appl. Acarol. 1998, 22, 39–50. [Google Scholar] [CrossRef]
- El-Banhawy, E.M.; El-Borolossy, M.A.; El-Sawaf, B.M.; Afia, S.I. Biological aspects and feeding behaviour of the predacious soil mite Nenteria hypotrichus (Uropodina: Uropodidae). Acarologia 1997, XXXVIII, 357–360. [Google Scholar]
- Walter, D.E. From the subantarctic to the subtropics: A revision of the Davacaridae Kethley, 1977 (Acari: Trigynaspida: Mesostigmata) with the description of a new genus and three new species. J. Nat. Hist. 2004, 38, 2033–2049. [Google Scholar] [CrossRef]
- Seeman, O.D. Life cycle, development, feeding and immature life stages of the Fedrizziidae (Mesostigmata: Fedrizzioidea). Acarologia 2000, 41, 39–52. [Google Scholar]
- Butler, L.; Hunter, P.E. Redescription of Megisthanus floridanus with observations on its biology (Acarina: Megisthanidae). Fla. Entomol. 1968, 51, 187. [Google Scholar] [CrossRef]
- Walter, D.E. A jumping mesostigmatan mite, Saltiseius hunteri n. g., n. sp. (Acari: Mesostigmata: Trigynaspida: Saltiseiidae, n. fam.) from Australia. Int. J. Acarol. 2000, 26, 25–31. [Google Scholar] [CrossRef]
- Seeman, O.D.; Walter, D.E. A new species of Triplogyniidae (Mesostigmata: Celaenopsoidea) from Australian rainforests. Int. J. Acarol. 1997, 23, 49–59. [Google Scholar] [CrossRef]
- Rockett, C.L. Nematode predation by oribatid mites (Acari: Oribatida). Int. J. Acarol. 1980, 6, 219–224. [Google Scholar] [CrossRef]
- Oliveira, A.R.; De Moraes, G.J.; Ferraz, L.C.C.B. Consumption rate of phytonematodes by Pergalumna sp. (Acari: Oribatida: Galumnidae) under laboratory conditions determined by a new method. Exp. Appl. Acarol. 2007, 41, 183–189. [Google Scholar] [CrossRef]
- Abou El-Atta, D.A.E.-M.; Ghazy, N.A.; Osman, M.A. Effects of temperature on the life-history traits of Sancassania (Caloglyphus) berlesei (Acari: Astigmatina: Acaridae) feeding on root-knot nematodes, Meloidogyne spp. (Nematoda: Meloidogynidae). Exp. Appl. Acarol. 2014, 64, 299–307. [Google Scholar] [CrossRef]
- Sell, P. Caloglyphus sp. (Acarina: Acaridae), an effective nematophagous mite on root-knot nematodes (Meloidogyne spp.). Nematol 1988, 34, 246–248. [Google Scholar] [CrossRef]
- Walter, D.E.; Kaplan, D.T. Feeding observations on two astigmatic mites, Schwiebea rocketti (Acaridae) and Histiostoma bakeri (Histiostomatidae) associated with Citrus feeder roots. Pedobiologia 1990, 34, 281–286. [Google Scholar]
- Taha, H.A.; El-Naggar, M.E.E.; Abou-El-Ngaga, M.M.; Soliman, S.M. Effect of different prey species on the development and fecundity of the predacious mite, Neocunaxoides andrei Baker and Hoff. (Acari: Cunaxidae). Agric. Res. Rev. 1988, 66, 129–135. [Google Scholar]
- Hernandes, F.A.; de Castro, T.M.M.G.; Venancio, R. Prostigmata (Acari: Trombidiformes) as Biological Control Agents. In Prospects for Biological Control of Plant Feeding Mites and Other Harmful Organisms; Carrillo, D., de Moraes, G.J., Peña, J.E., Eds.; Springer International Publishing: Cham, Switzerland, 2015; ISBN 978-3-319-15042-0. [Google Scholar]
- Fan, Q.-H.; Walter, D.E. Genus Caligohomus Habeeb (Acari: Prostigmata: Stigmaeidae). Syst. Appl. Acarol. 2004, 9, 77–88. [Google Scholar] [CrossRef]
Figure 1.
Herbivore (green) and detritivore (brown) food chain in an arable field cropped with maize at 0–10 cm depth. Scheme on C flow and bottom–up and top–down forces in the food web. BF–bacterial feeders, FF–fungal feeders, PF–plant feeders, OMN–omnivores, PRE–predators. Biomass of the soil fauna in brackets in g C m−2, data from Pausch et al. [21].
Figure 1.
Herbivore (green) and detritivore (brown) food chain in an arable field cropped with maize at 0–10 cm depth. Scheme on C flow and bottom–up and top–down forces in the food web. BF–bacterial feeders, FF–fungal feeders, PF–plant feeders, OMN–omnivores, PRE–predators. Biomass of the soil fauna in brackets in g C m−2, data from Pausch et al. [21].

Figure 2.
Number of mite species per family of Gamasina (Mesostigmata) reported consuming nematodes. APN = animal parasitic nematodes, FLN = free-living nematodes, PPN = plant parasitic nematodes.
Figure 2.
Number of mite species per family of Gamasina (Mesostigmata) reported consuming nematodes. APN = animal parasitic nematodes, FLN = free-living nematodes, PPN = plant parasitic nematodes.

Figure 3.
Low temperature scanning electron microscope images of Stratiolaelaps scimitus Womersley sensing and feeding nematodes. (a) Adult female senses the free-living nematode Pristionchus aerivorus (Cobb in Merrill and Ford) Chitwood by touching the nematode with the tarsus of leg I. (b) Adult female grasping a J2 of the root-knot nematode Meloidogyne incognita (Kofoid and White) between palps just at the level of the apotele. (c) Immature holding P. aerivorus with its chelicerae and initiating the feeding process. (d) Close-up of the mite gnathosoma in dorsal view of an adult female holding between the chelicera and feeding on the free-living nematode Mesorhabditis inarimensis (Meyl) Dougherty. Images: Joe Mowery, Gary Bauchan, Ron Ochoa, Lynn Carta, Eric Palevsky.
Figure 3.
Low temperature scanning electron microscope images of Stratiolaelaps scimitus Womersley sensing and feeding nematodes. (a) Adult female senses the free-living nematode Pristionchus aerivorus (Cobb in Merrill and Ford) Chitwood by touching the nematode with the tarsus of leg I. (b) Adult female grasping a J2 of the root-knot nematode Meloidogyne incognita (Kofoid and White) between palps just at the level of the apotele. (c) Immature holding P. aerivorus with its chelicerae and initiating the feeding process. (d) Close-up of the mite gnathosoma in dorsal view of an adult female holding between the chelicera and feeding on the free-living nematode Mesorhabditis inarimensis (Meyl) Dougherty. Images: Joe Mowery, Gary Bauchan, Ron Ochoa, Lynn Carta, Eric Palevsky.

Figure 4.
(a) Adult female of Protogamasellopsis zaheri Abo-Shnaf, Castilho and Moraes (Mesostigmata: Rhodacaridae) from the small-pore guild feeding on the free-living nematode Acrobeloides sp. (b) Adult female Gaeolaelaps aculeifer (Canestrini) (Mesostigmata: Laelapidae) feeding on the free-living nematode Panagrellus sp.
Figure 4.
(a) Adult female of Protogamasellopsis zaheri Abo-Shnaf, Castilho and Moraes (Mesostigmata: Rhodacaridae) from the small-pore guild feeding on the free-living nematode Acrobeloides sp. (b) Adult female Gaeolaelaps aculeifer (Canestrini) (Mesostigmata: Laelapidae) feeding on the free-living nematode Panagrellus sp.

Figure 5.
Schematic illustration of our hypothesis: conservation of soil mite predators with available, suitable, and accessible free-living nematodes as prey will provide better agricultural ecosystem performance and long-range sustainability.
Figure 5.
Schematic illustration of our hypothesis: conservation of soil mite predators with available, suitable, and accessible free-living nematodes as prey will provide better agricultural ecosystem performance and long-range sustainability.
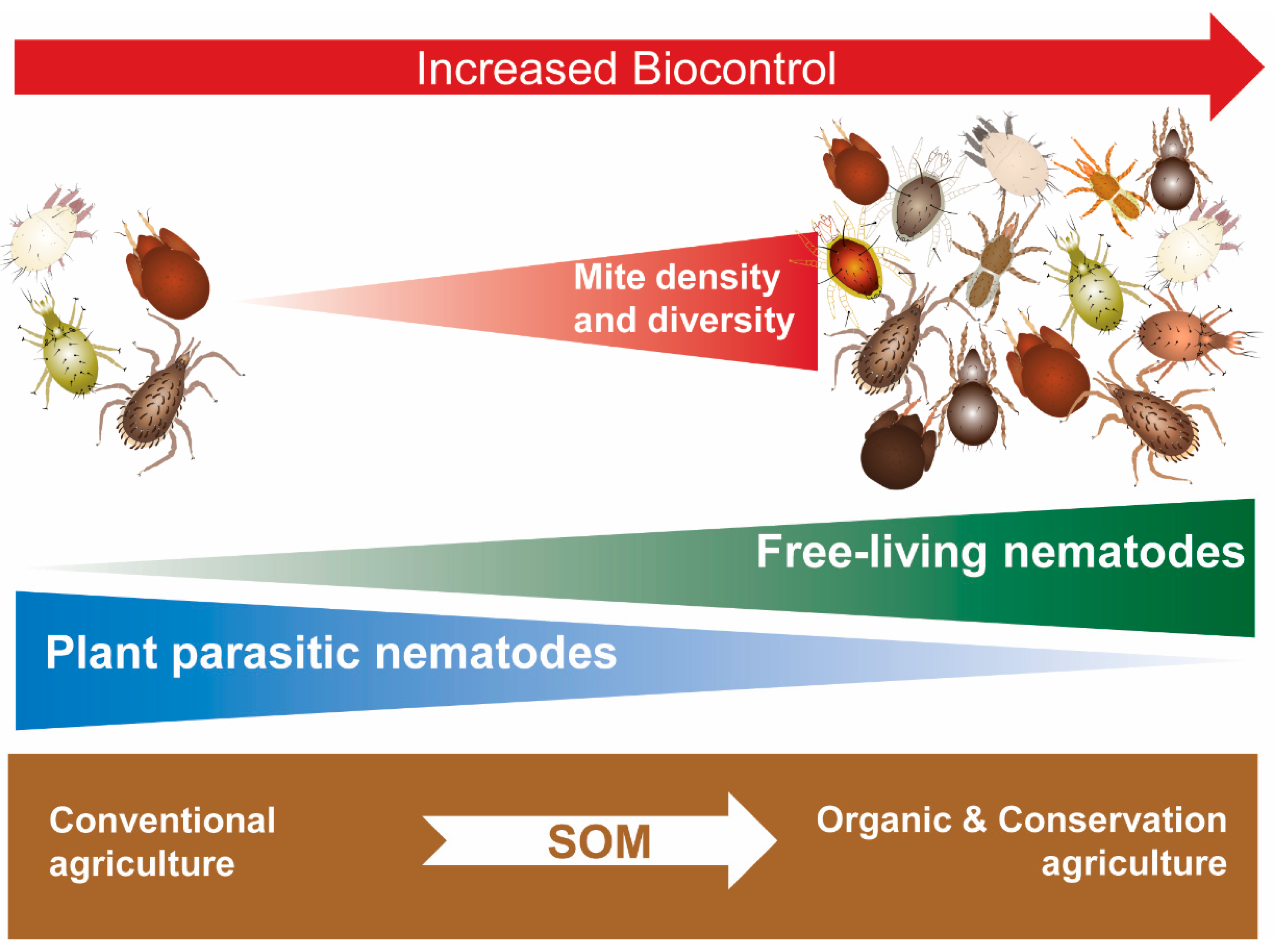

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Soil mites commercialized for biological control (modified after Van Lenteren [91] and Knapp et al. [92]).
| Species | Target | Region of Application | First Commercial Use |
|---|---|---|---|
| Androlaelaps casalis (Berlese, 1887) | Dermanyssus | Europe | 2012 |
| Stratiolaelaps miles (Berlese, 1882) | Sciaridae | Europe, North America, Asia, Aus/NZ | 1995 |
| Stratiolaelaps scimitus (Womersley, 1956) | Sciaridae, thrips | Europe, Latin America | 1990 |
| Gaeolaelaps aculeifer (Canestrini, 1884) | Diptera, thrips, mites | Europe, North Africa, North America, Asia, Aus/NZ | 1995 |
| Gaeolaelaps gillespiei Beaulieu | Sciaridae, thrips | North America (Canada) | 2009 |
| Macrocheles robustulus (Berlese, 1904) | Diptera, thrips, Lepidoptera? | Europe | 2010 |
| Pergamasus quisquiliarum (Canestrini, 1882) | Symphyla | Europe | 2000 |
| Cheyletus eruditus (Schrank, 1781) | Dermanyssus, pests of stored products | Europe | 1985 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rueda-Ramírez, D.; Palevsky, E.; Ruess, L. Soil Nematodes as a Means of Conservation of Soil Predatory Mites for Biocontrol. Agronomy 2023, 13, 32. https://doi.org/10.3390/agronomy13010032
AMA Style
Rueda-Ramírez D, Palevsky E, Ruess L. Soil Nematodes as a Means of Conservation of Soil Predatory Mites for Biocontrol. Agronomy. 2023; 13(1):32. https://doi.org/10.3390/agronomy13010032
Chicago/Turabian StyleRueda-Ramírez, Diana, Eric Palevsky, and Liliane Ruess. 2023. "Soil Nematodes as a Means of Conservation of Soil Predatory Mites for Biocontrol" Agronomy 13, no. 1: 32. https://doi.org/10.3390/agronomy13010032
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.