
Betalain Content and Morphological Characteristics of Table Beet Accessions: Their Interplay with Abiotic Factors



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Agricultural Details
2.2. Quantification of Betalains
2.3. Statistical Analyses
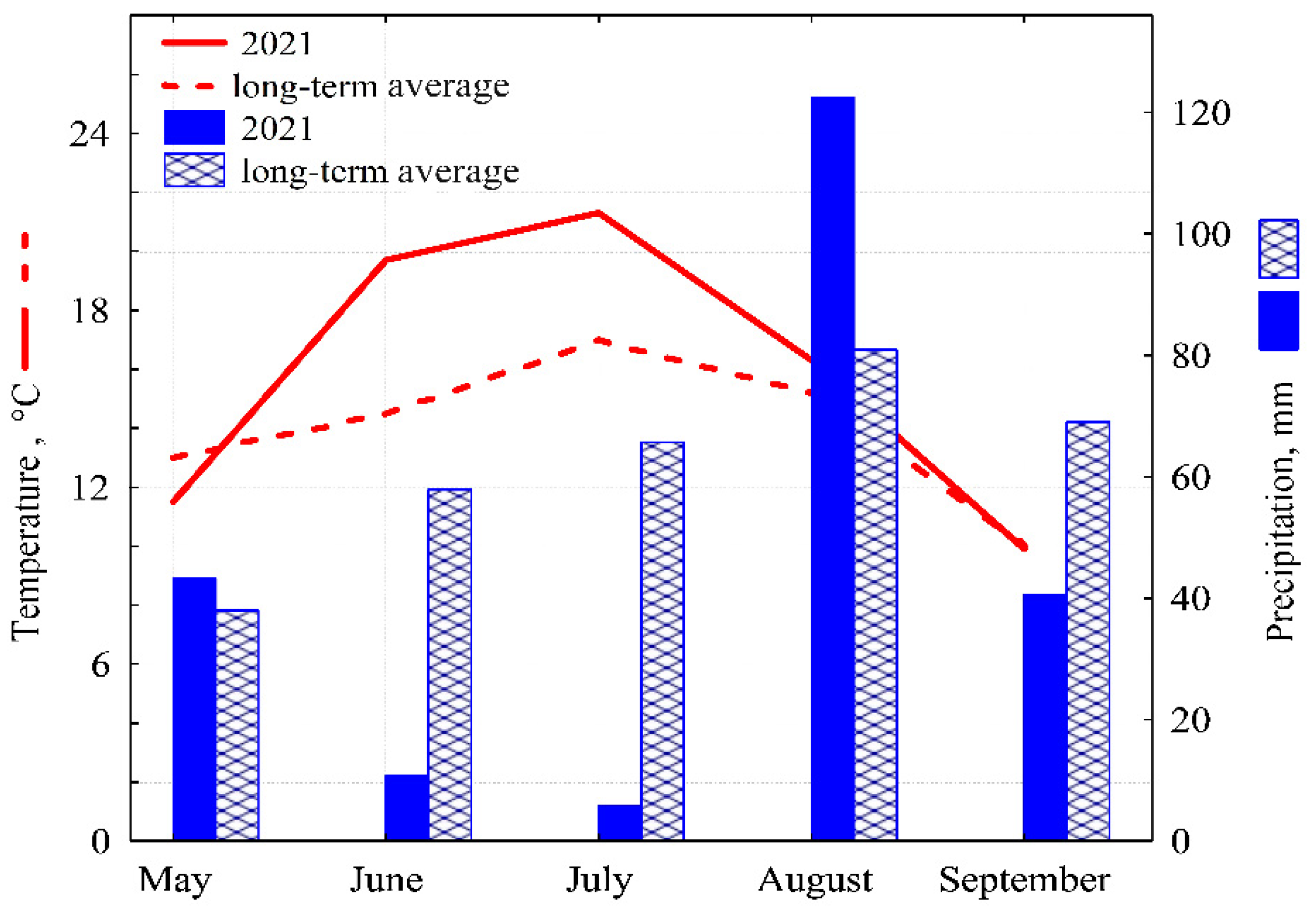
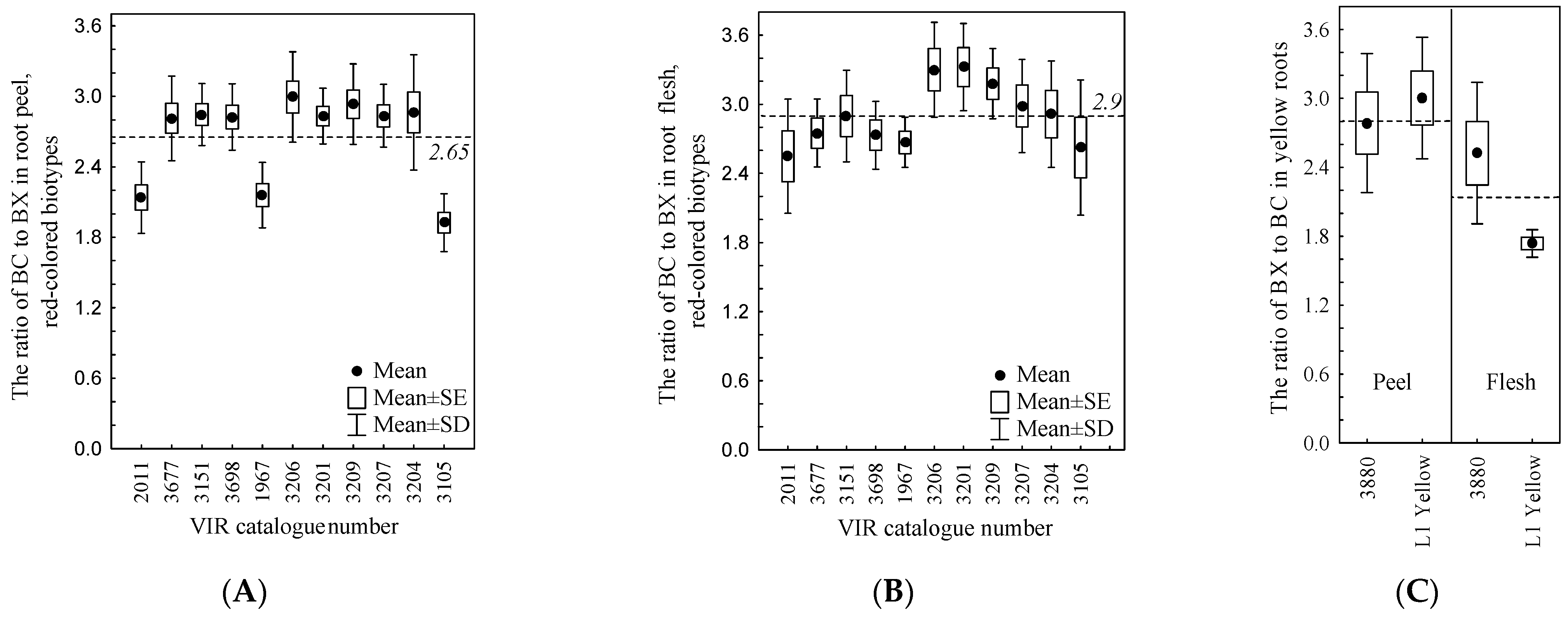
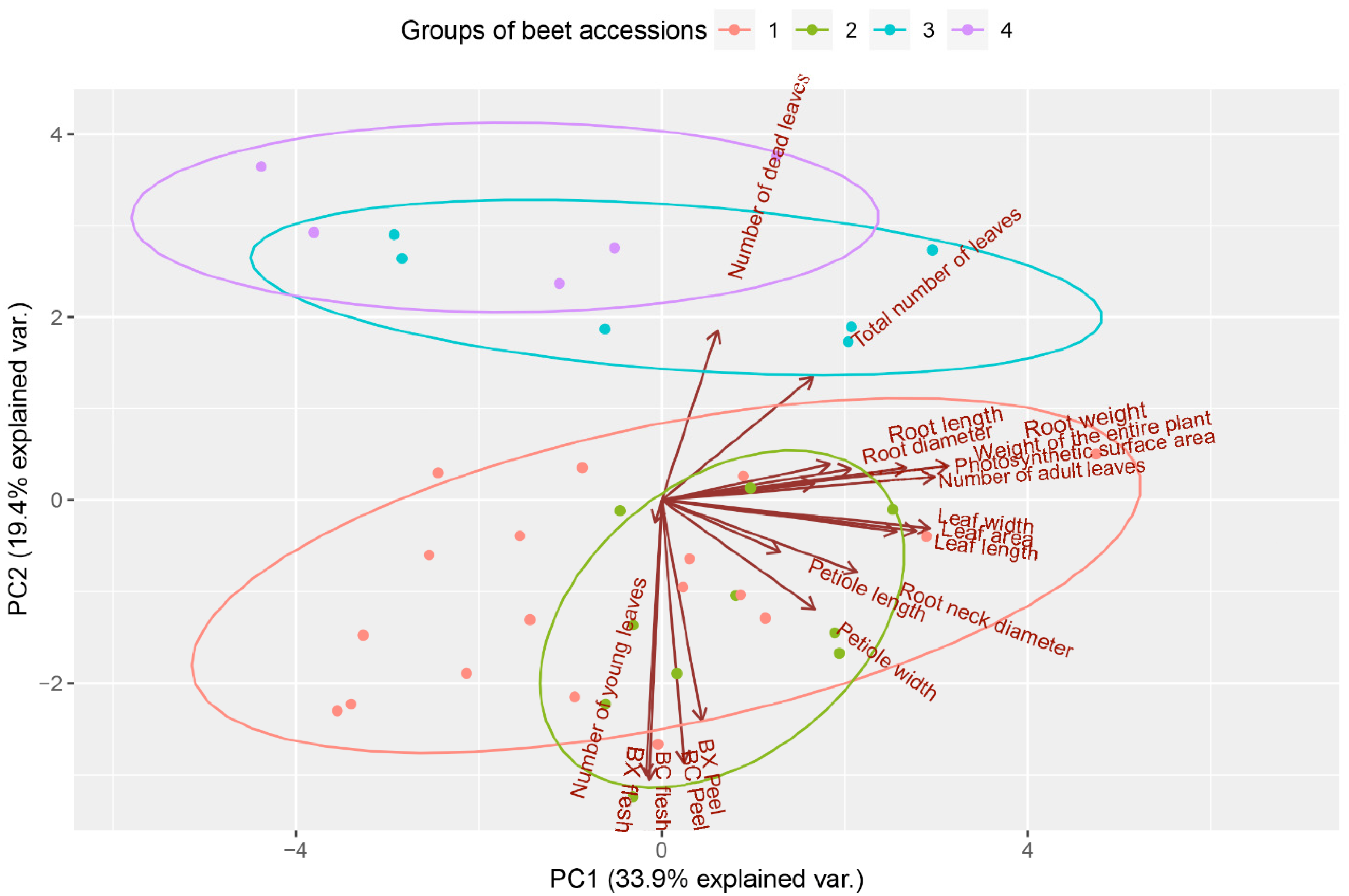
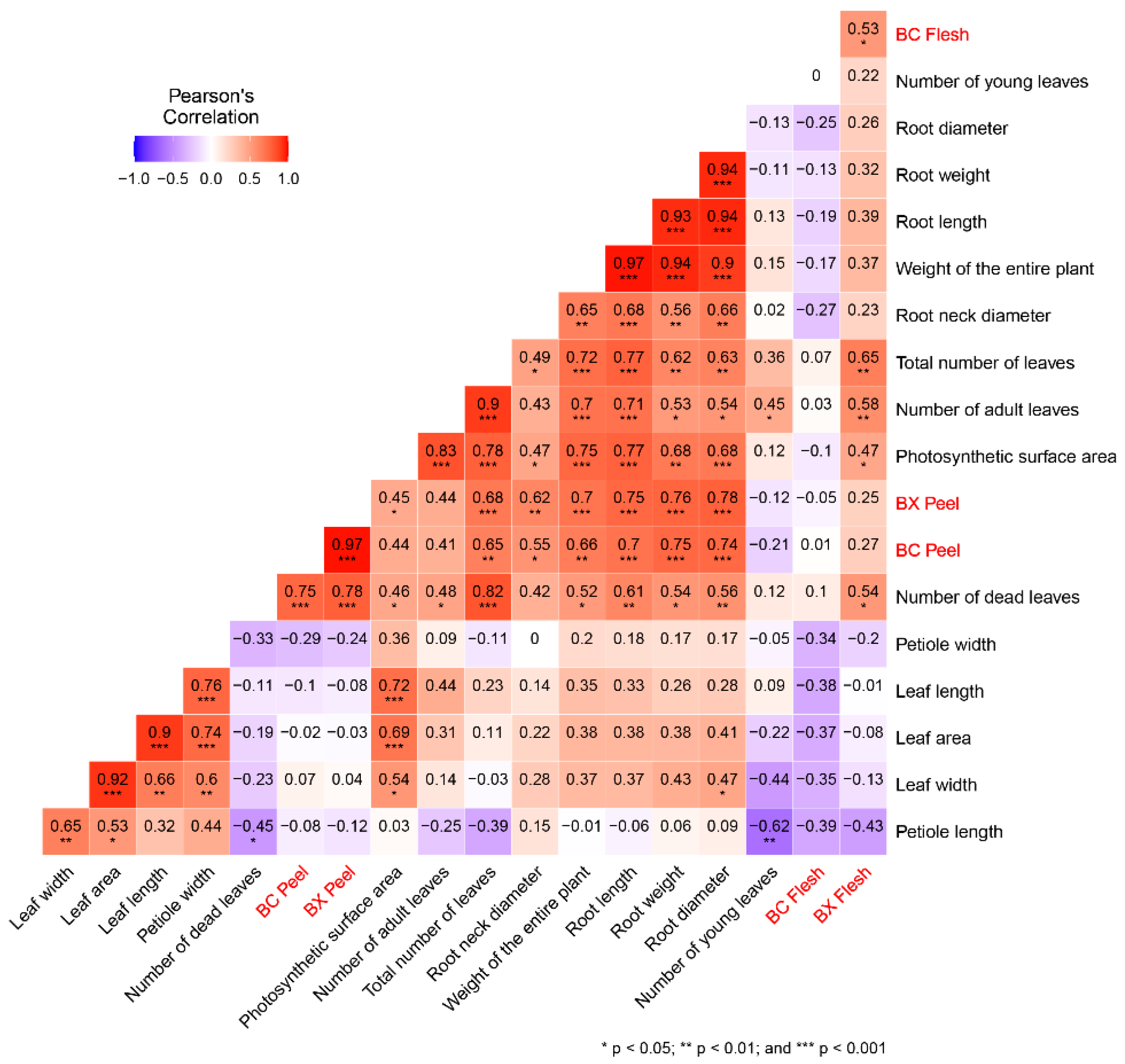
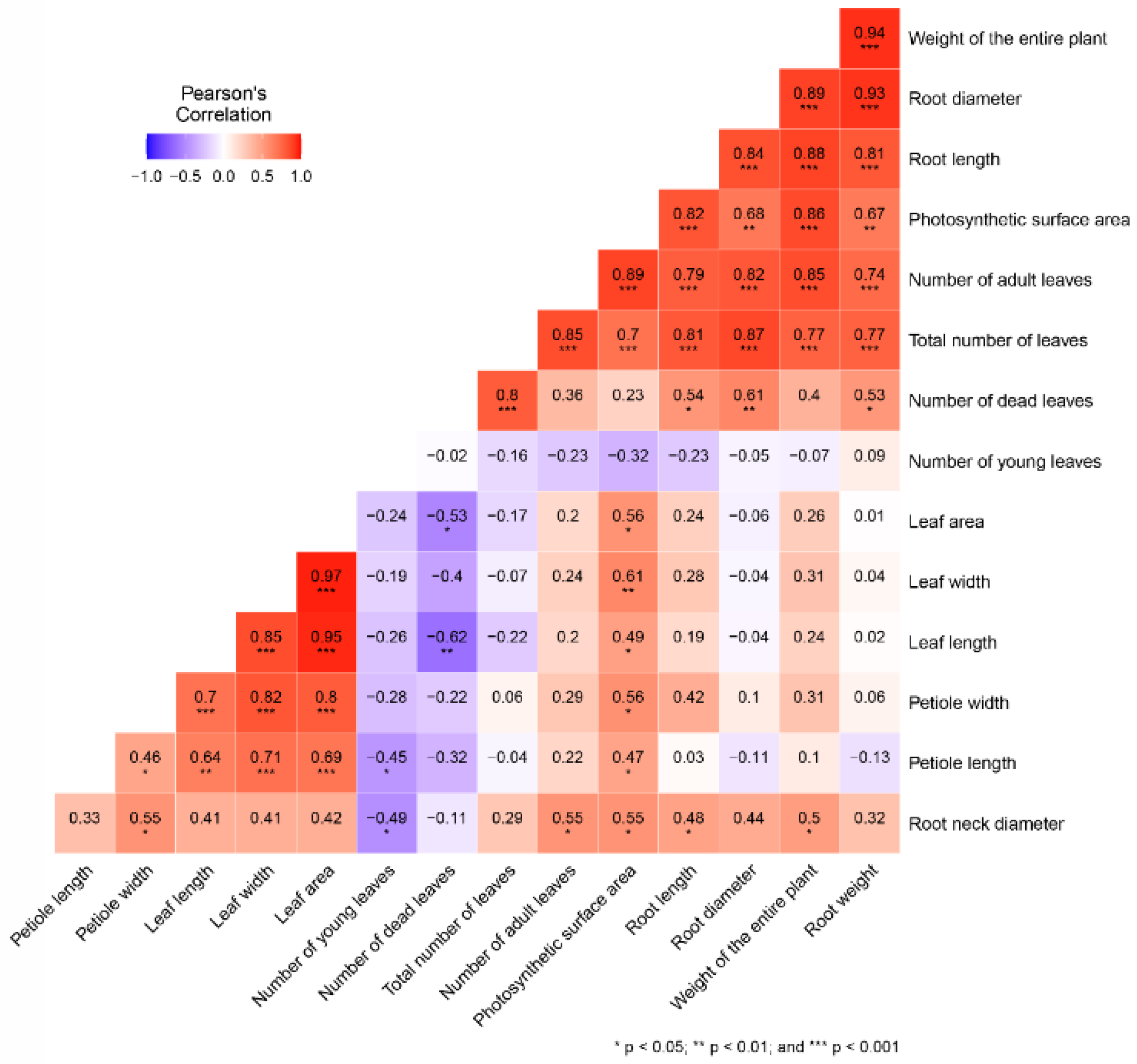
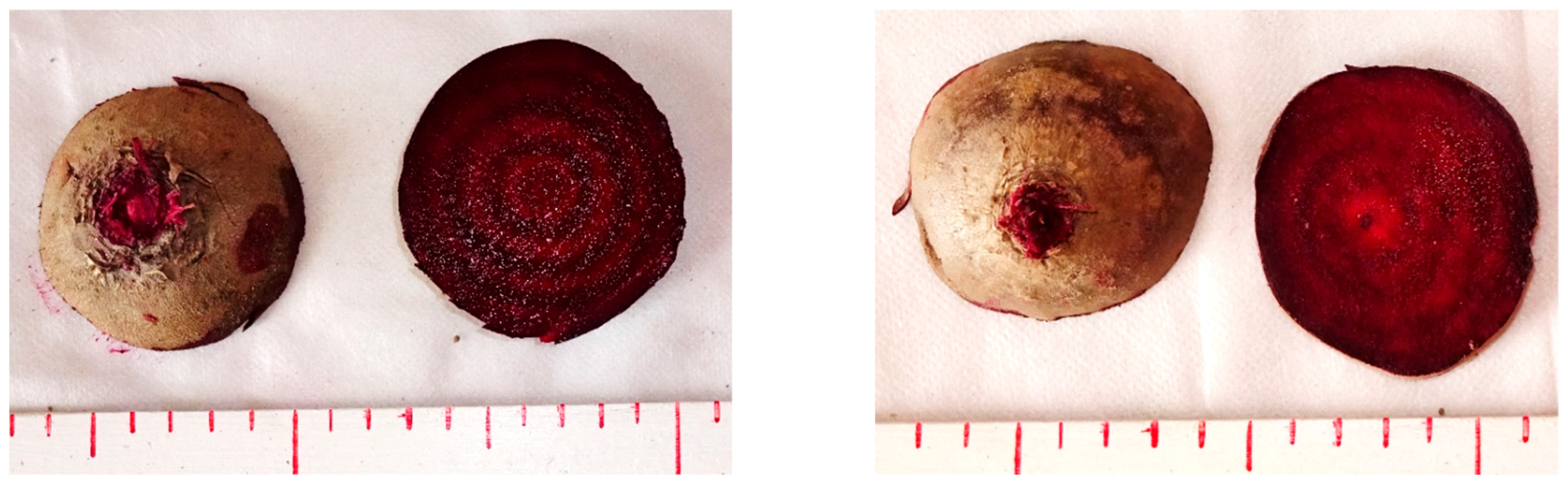
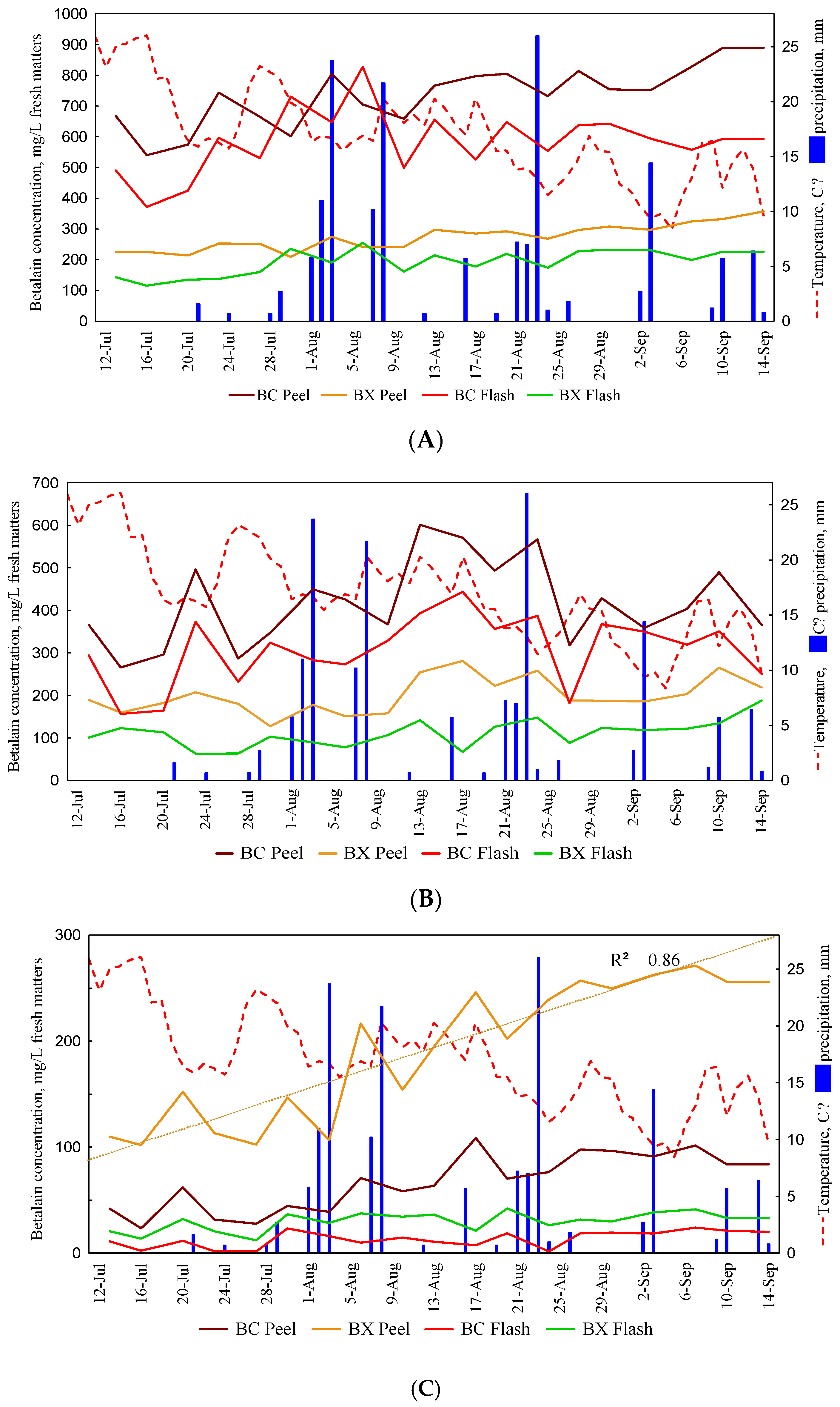
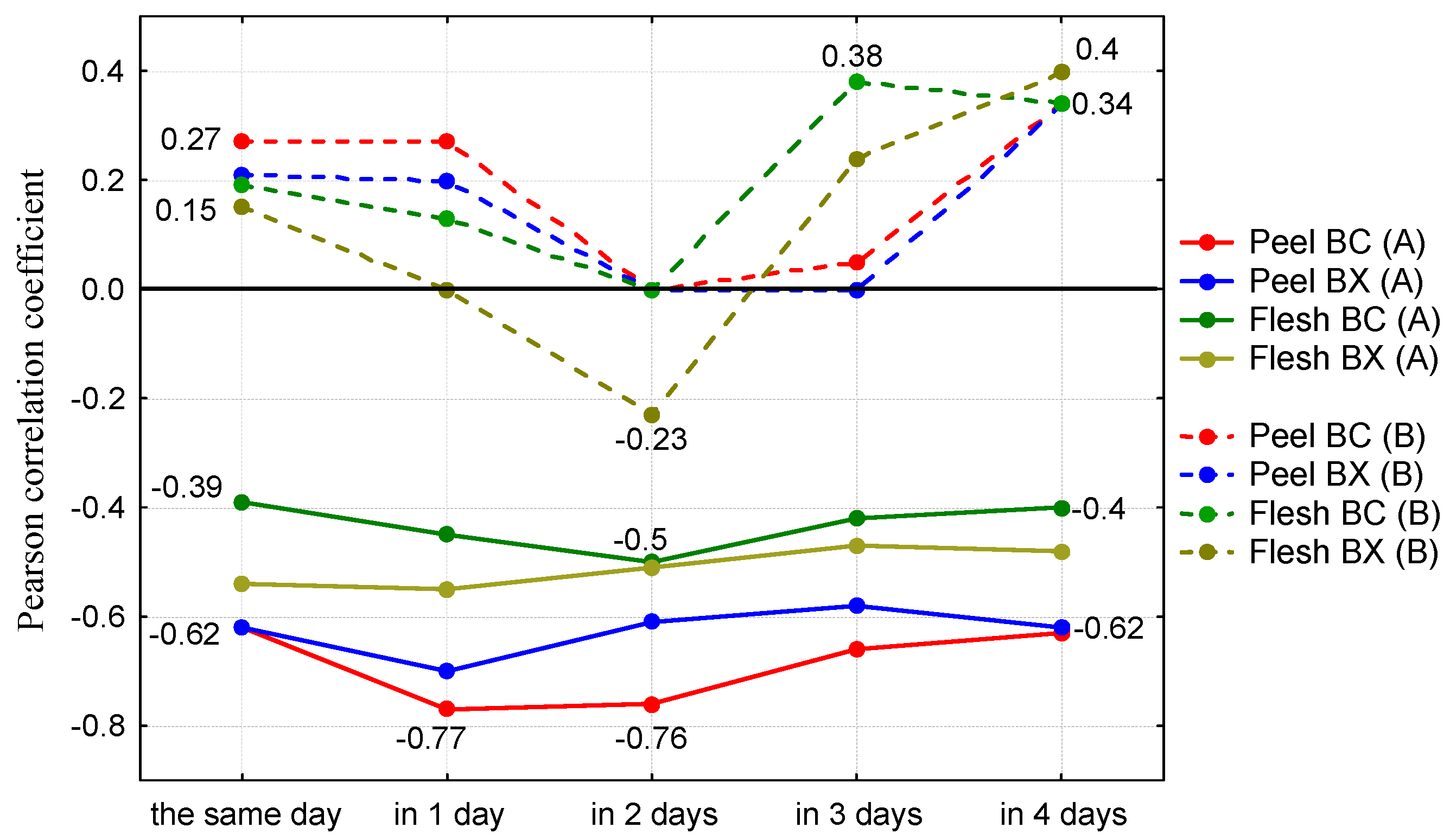
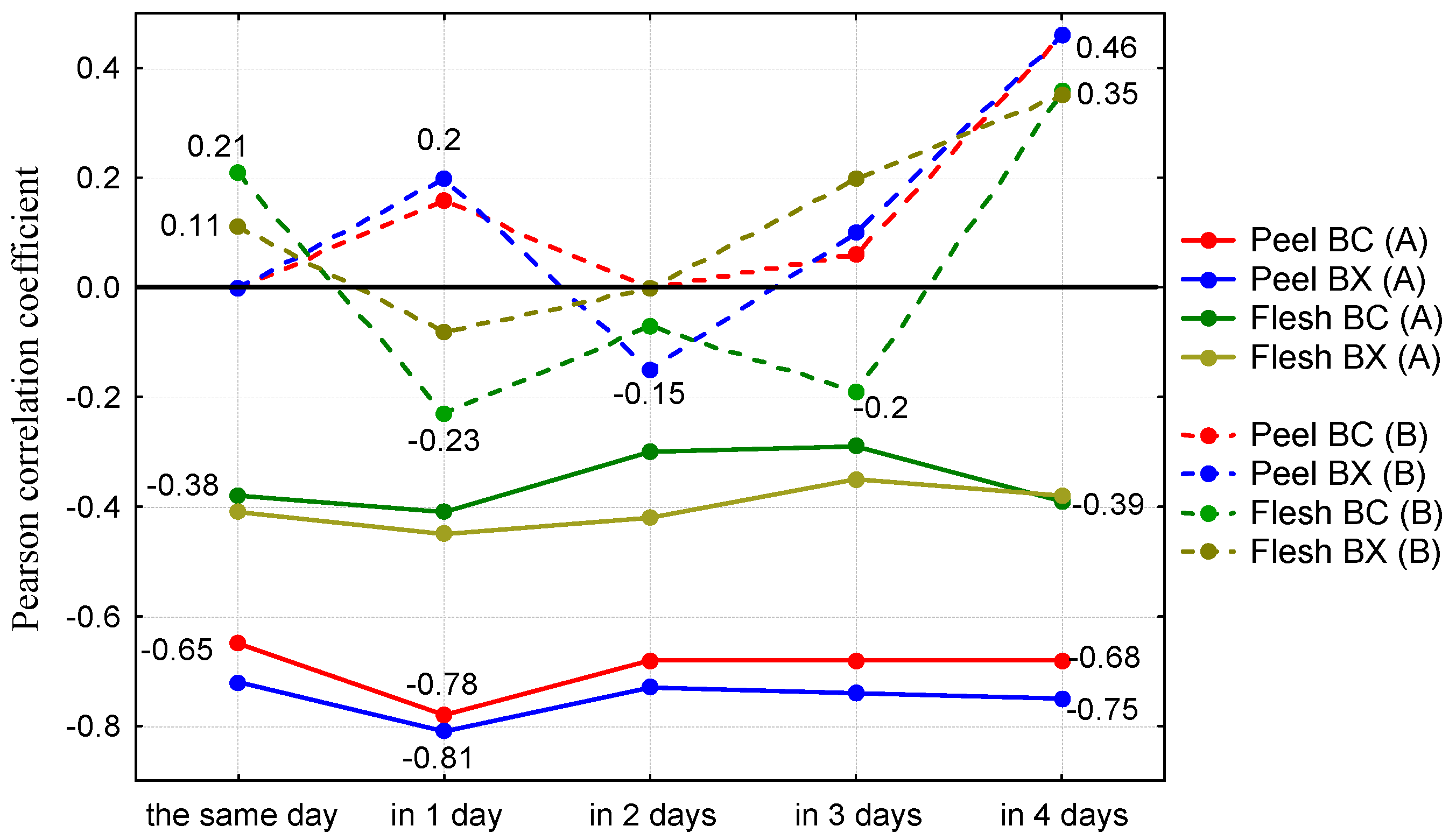
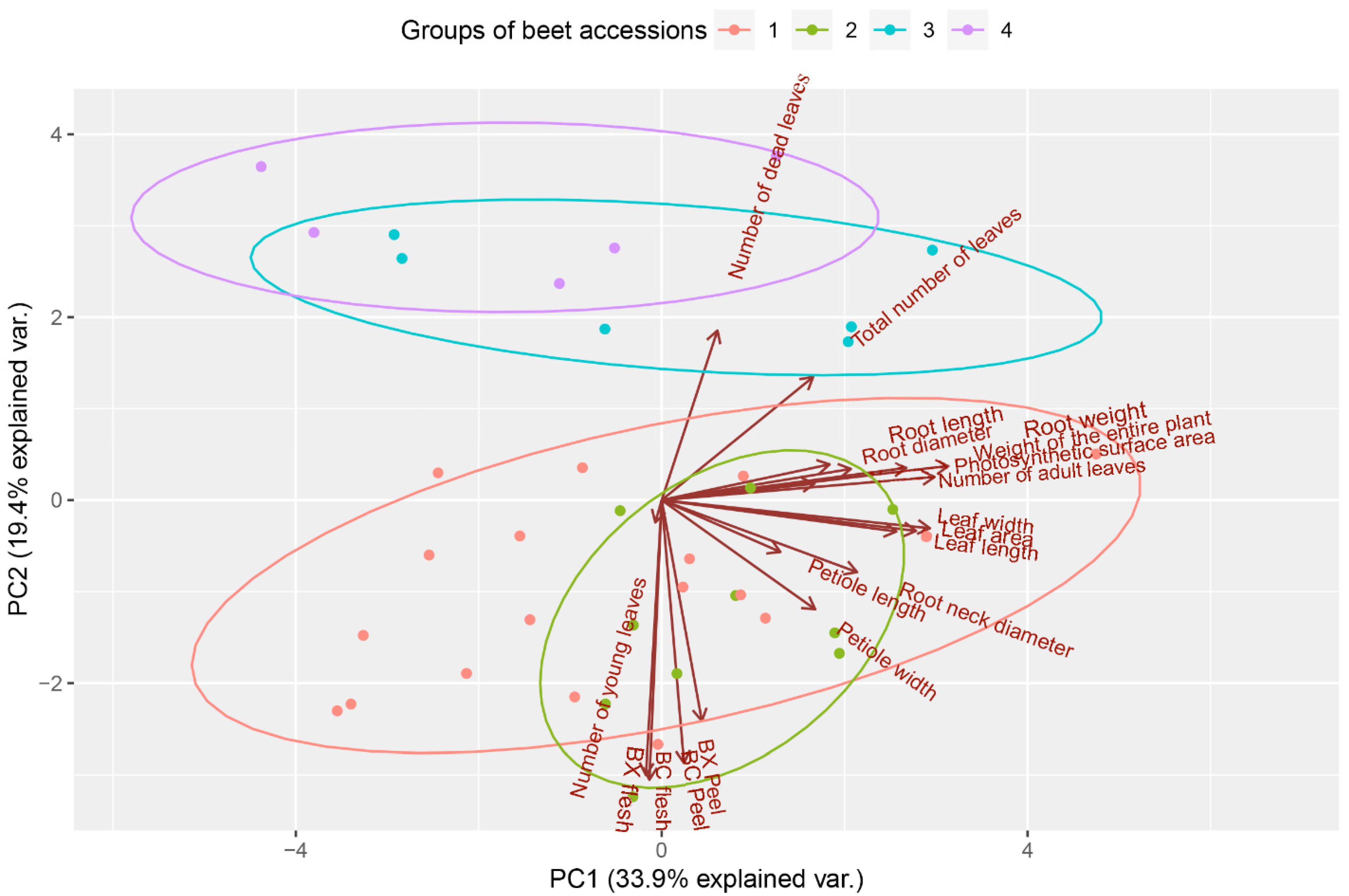
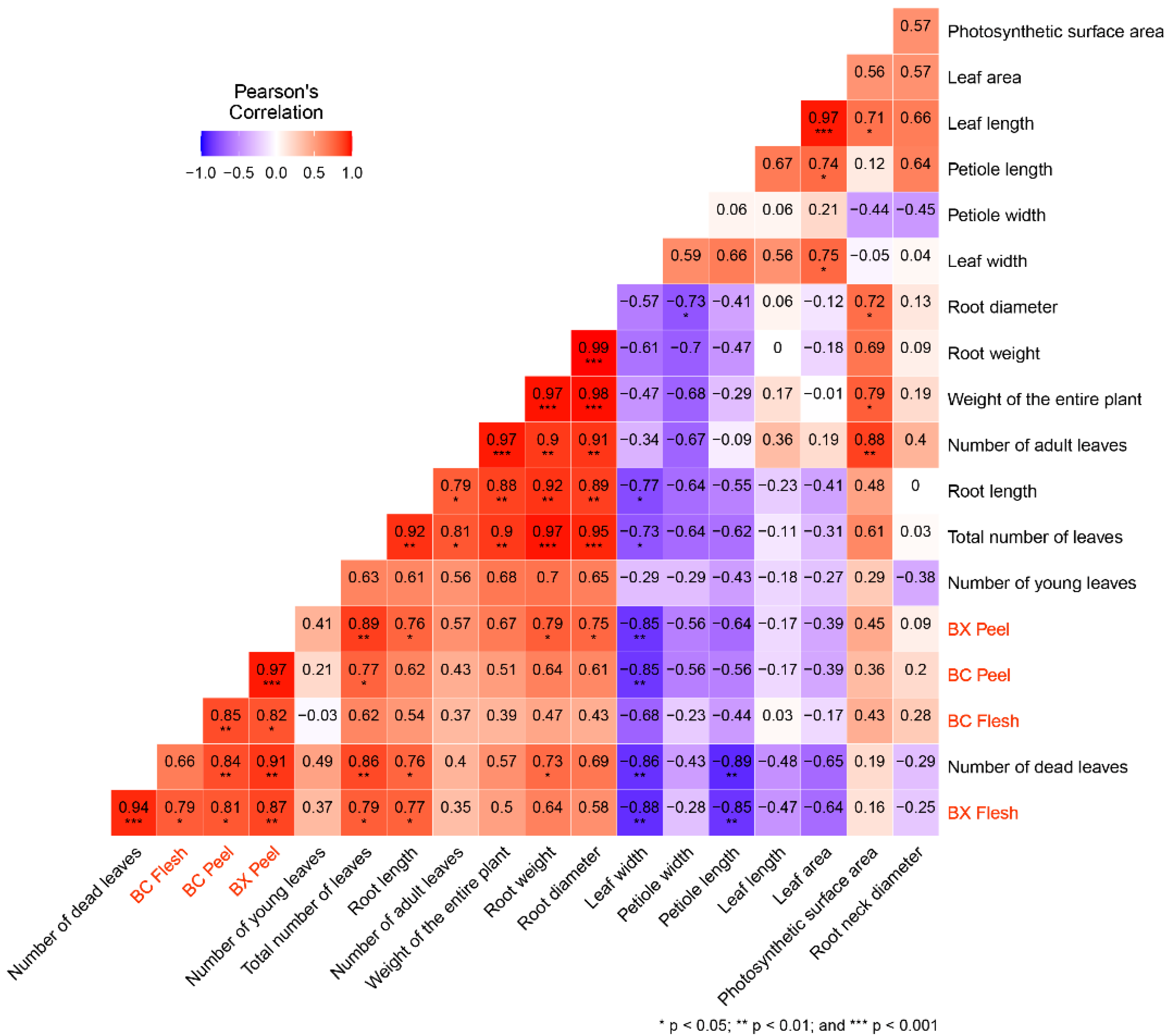
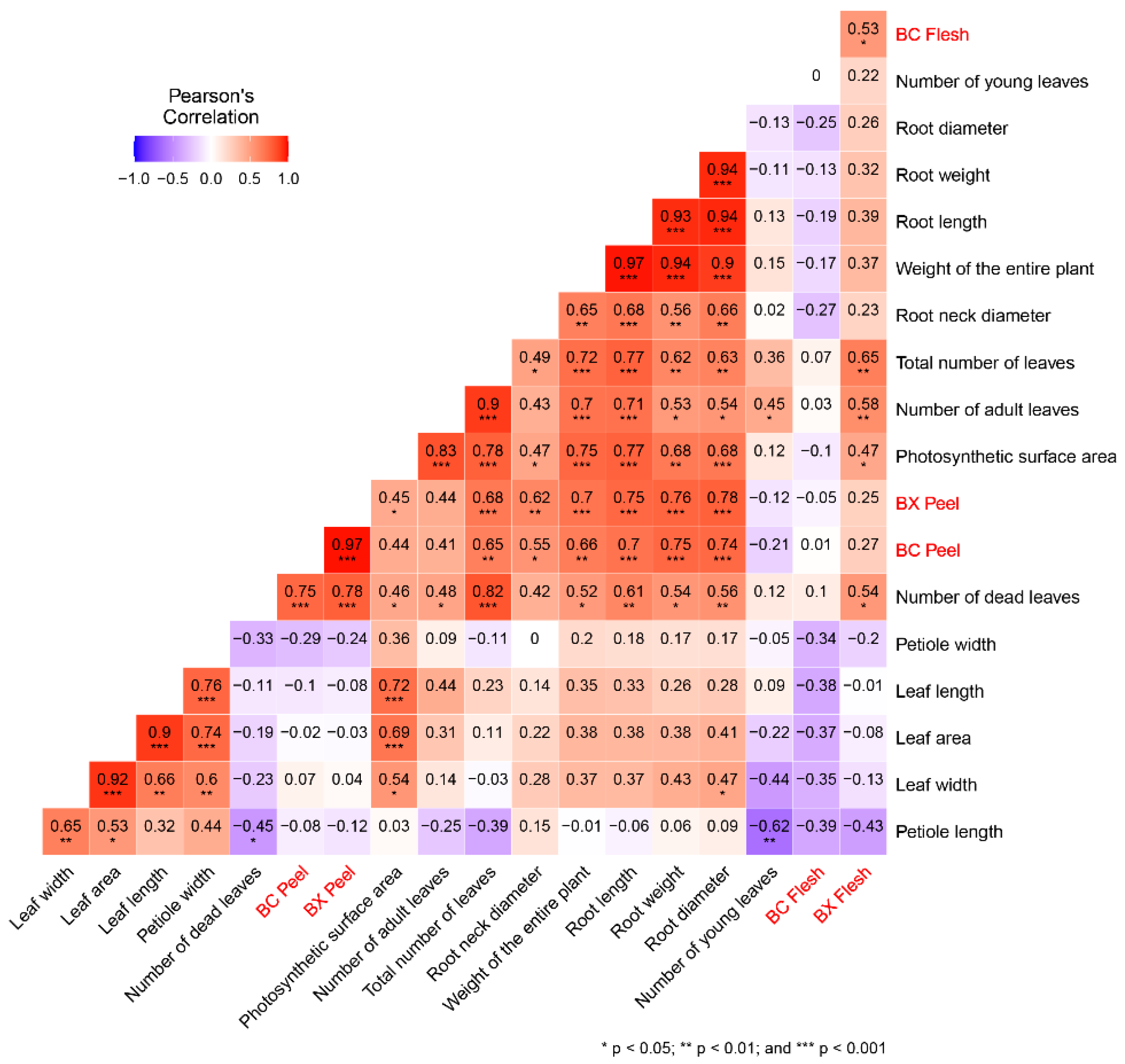
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stintzing, F.C.; Schieber, A.; Carle, R. Rote Bete als Färbendes Lebensmittel- eine Bestandsaufnahme. Obst.-Gemüse-Kartoff. 2000, 85, 196–204. [Google Scholar]
- Strack, D.; Vogt, T.; Schliemann, W. Recent advances in betalain research. Phytochemistry 2003, 62, 247–269. [Google Scholar] [CrossRef]
- Sokolova, D.V. Formation of the trait-specific group in VIR’s table beet collection: Environmental plasticity and stability. Proc. Appl. Bot. Genet. Breed. 2018, 179, 106–117. [Google Scholar] [CrossRef]
- Cai, Y.; Corke, H. Amaranthus betacyanin pigments applied in model food systems. J. Food Sci. 1999, 64, 869–873. [Google Scholar] [CrossRef]
- Castellanos-Santiago, E.; Yahia, E.M. Identification and quantification of betalains from the fruits of 10 mexican prickly pear cultivars by high-performance liquid chromatography and electrospray ionization mass spectrometry. J. Agric. Food Chem. 2008, 56, 5758–5764. [Google Scholar] [CrossRef]
- Sumaya-Martínez, M.T.; Cruz-Jaime, S.; Madrigal-Santillán, E.; García-Paredes, J.D.; Cariño-Cortés, R.; Cruz-Cansino, N.; Valadez-Vega, C.; Martinez-Cardenas, C.; Alanís-García, E. Betalain, acid ascorbic, phenolic contents and antioxidant properties of purple, red, yellow and white cactus pears. Int. J. Mol. Sci. 2011, 12, 6452–6468. [Google Scholar] [CrossRef] [Green Version]
- Sawicki, T.; Bączek, N.; Wiczkowski, W. Betalain profile, content and antioxidant capacity of red beetroot dependent on the genotype and root part. J. Funct. Foods 2016, 27, 249–261. [Google Scholar] [CrossRef]
- Vogt, T.; Ibdah, M.; Schmidt, J.; Wray, V.; Nimtz, M.; Strack, D. Light-induced betacyanin and flavonol accumulation in bladder cells of Mesembryanthemum crystallinum. Phytochemistry 1999, 52, 583–592. [Google Scholar] [CrossRef]
- Wang, C.Q.; Zhao, J.Q.; Chen, M.; Wang, B.S. Identification of betacyanin and effects of environmental factors on its accumulation in halophyte Suaeda salsa L. J. Plant Physiol. Mol. Biol. 2006, 32, 195–201. [Google Scholar]
- Hayakawa, K.; Agarie, S. Physiological roles of betacyanin in a halophyte, Suaeda japonica Makino. Plant Prod. Sci. 2010, 13, 351–359. [Google Scholar] [CrossRef]
- Sepúlveda-Jiménez, G.; Rueda-Benítez, P.; Porta, H.; Rocha-Sosa, M. Betacyanin synthesis in red beet (Beta vulgaris) leaves induced by wounding and bacterial infiltration is preceded by an oxidative burst. Physiol. Mol. Plant Pathol. 2004, 64, 125–133. [Google Scholar] [CrossRef]
- Casique-Arroyo, G.; Martínez-Gallardo, N.; González de la Vara, L.; Délano-Frier, J.P. Betacyanin biosynthetic genes and enzymes are differentially induced by (a)biotic stress in Amaranthus hypochondriacus. PLoS ONE 2014, 9, e99012. [Google Scholar] [CrossRef] [PubMed]
- Kapadia, G.J.; Tokuda, H.; Konoshima, T.; Nishino, H. Chemoprevention of lung and skin cancer by Beta vulgaris (beet) root extract. Cancer Lett. 1996, 100, 211–214. [Google Scholar] [CrossRef]
- Azeredo, H.M.C. Betalains: Properties, sources, applications, and stability—A review. Int. J. Food Sci. Technol. 2008, 44, 2365–2376. [Google Scholar] [CrossRef] [Green Version]
- Vulić, J.J.; Ćebović, T.N.; Čanadanović, V.M.; Ćetković, G.S.; Djilas, S.M.; Čanadanović-Brunet, J.M.; Tumbas, V.T. Antiradical, antimicrobial and cytotoxic activities of commercial beetroot pomace. Food Funct. 2013, 4, 713. [Google Scholar] [CrossRef]
- Ninfali, P.; Antonini, E.; Frati, A.; Scarpa, E.S. C-glycosyl flavonoids from Beta vulgaris cicla and betalains from Beta vulgaris rubra: Antioxidant, anticancer and antiinflammatory activities—A review. Phytother. Res. 2017, 31, 871–884. [Google Scholar] [CrossRef]
- De Oliveira, S.P.A.; do Nascimento, H.M.A.; Sampaio, K.B.; de Souza, E.L. A review on bioactive compounds of beet (Beta vulgaris L. subsp. vulgaris) with special emphasis on their beneficial effects on gut microbiota and gastrointestinal health. Crit. Rev. Food Sci. Nutr. 2020, 61, 2022–2033. [Google Scholar] [CrossRef]
- Da Silva, D.V.t.; Baião, D.D.S.; Ferreira, V.F.; Flosi Paschoalin, V.M. Betanin as a multipath oxidative stress and inflammation modulator: A beetroot pigment with protective effects on cardiovascular disease pathogenesis. Crit. Rev. Food Sci. Nutr. 2021, 62, 539–554. [Google Scholar] [CrossRef]
- Vinson, J.A.; Hao, Y.; Su, X.; Zubik, L. Phenol antioxidant quantity and quality in foods: Vegetables. J. Agric. Food Chem. 1998, 46, 3630–3634. [Google Scholar] [CrossRef]
- Pedreño, M.A.; Escribano, J. Studying the oxidation and the antiradical activity of betalain from beetroot. J. Biol. Educ. 2000, 35, 49–51. [Google Scholar] [CrossRef]
- Wettasinghe, M.; Bolling, B.; Plhak, L.; Parkin, K. Screening for phase II enzyme-inducing and antioxidant activities of common vegetables. J. Food Sci. 2002, 67, 2583–2588. [Google Scholar] [CrossRef]
- Hadipour, E.; Taleghani, A.; Tayarani-Najaran, N.; Tayarani-Najaran, Z. Biological effects of red beetroot and betalains: A review. Phytother. Res. 2020, 34, 1847–1867. [Google Scholar] [CrossRef]
- Nizioł-Łukaszewska, Z.; Gawęda, M. Changes in quality of selected red beet (Beta vulgaris L.) cultivars during the growing season. Folia Hortic. 2014, 26, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Nizioł-Łukaszewska, Z.; Gawęda, M. Selected indicators of the root quality of fifteen cultivars of red beet (Beta vulgaris L.). J. Hortic. Res. 2015, 23, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Montes-Lora, S.; Rodríguez-Pulido, F.J.; Cejudo-Bastante, M.J.; Heredia, F.J. Implications of the red beet ripening on the colour and betalain composition relationships. Plant Foods Hum. Nutr. 2018, 73, 216–221. [Google Scholar] [CrossRef]
- Sokolova, D.V. Dynamic changes in red beet betanin content during the growing season: Their interplay with abiotic factors. Vavilovskii Zhurnal Genet. I Sel. 2022, 26, 30–39. [Google Scholar] [CrossRef]
- Herbach, K.M.; Stintzing, F.C.; Carle, R. Impact of thermal treatment on color and pigment pattern of red beet (Beta vulgaris L.) preparations. J. Food Sci. 2006, 69, 491–498. [Google Scholar] [CrossRef]
- Kayın, N.; Atalay, D.; Türken Akçay, T.; Erge, H.S. Color stability and change in bioactive compounds of red beet juice concentrate stored at different temperatures. J. Food Sci. Technol. 2019, 56, 5097–5106. [Google Scholar] [CrossRef]
- Herbach, K.M.; Stintzing, F.C.; Carle, R. Stability and color changes of thermally treated betanin, phyllocactin, and hylocerenin solutions. J. Agric. Food Chem. 2006, 54, 390–398. [Google Scholar] [CrossRef]
- Wybraniec, S. Formation of decarboxylated betacyanins in heated purified betacyanin fractions from red beet root (Beta vulgaris L.) monitored by LC−MS/MS. J. Agric. Food Chem. 2005, 53, 3483–3487. [Google Scholar] [CrossRef]
- Sutor-Świeży, K.; Antonik, M.; Proszek, J.; Nemzer, B.; Pietrzkowski, Z.; Popenda, Ł.; Wybraniec, S. Dehydrogenation of betacyanins in heated betalain-rich extracts of red beet (Beta vulgaris L.). Int. J. Mol. Sci. 2022, 23, 1245. [Google Scholar] [CrossRef]
- Mglinets, A.V.; Osipova, Z.A. Formation of root color and its genetic control in fodder beet. Vavilovskii Zhurnal Genet. I Sel. 2010, 14, 720–728. [Google Scholar]
- Azeredo, H.M.C.; Pereira, A.C.; de Souza, A.C.R.; Gouveia, S.T.; Mendes, K.C.B. Study on efficiency of betacyanin extraction from red beetroots. Int. J. Food Sci. Technol. 2009, 44, 2464–2469. [Google Scholar] [CrossRef]
- Burak, L.C.H.; Zavaley, A.P. Influence of the method of processing table beets on the antioxidant activity of juice and fruit and vegetable juices. Tekhnologii Pishchevoy I Pererabat. Promyshlennosti APK-Prod. Zdorovogo Pitan. Technol. Food Process. Ind. AIC Healthy Food 2020, 4, 51–61. (In Russian) [Google Scholar] [CrossRef]
- Frolov, V.L.; Chizhik, J.L. Method of Preparing Food Dye from Beet. Invention Patent. RU 2081136 C1. 1997. Available online: https://new.fips.ru/registers-doc-view/fips_servlet?DB=RUPAT&DocNumber=2081136&TypeFile=html (accessed on 23 March 2022). (In Russian).
- Burenin, V.I. (Ed.) Methodological Guidelines for the Study and Maintenance of the World Collection of Root Crops; VIR Publ.: Leningrad, Russia, 1989; Available online: https://search.rsl.ru/ru/record/01001503638 (accessed on 23 March 2022). (In Russian)
- Stintzing, F.C.; Schieber, A.; Carle, R. Evaluation of colour properties and chemical quality parameters of cactus juices. Eur. Food Res. Technol. 2003, 216, 303–311. [Google Scholar] [CrossRef]
- Sokolova, D.V.; Solovieva, A.E. Promising starting material for selection of beet varieties with a high content of betanin. Agrar. Russ. 2019, 8, 26–32. [Google Scholar] [CrossRef]
- Krasochkin, V.T. Beet, 1st ed.; Selkhozgiz: Leningrad, Russia, 1960; pp. 42–92. [Google Scholar]
- Kujala, T.S.; Vienola, M.S.; Klika, K.D.; Loponen, J.M.; Pihlaja, K. Betalain and phenolic compositions of our beetroot (Beta vulgaris) cultivars. Eur. Food Res. Technol. 2002, 214, 505–510. [Google Scholar] [CrossRef]
- Wolyn, D.J.; Gabelman, W.H. Selection for betalain pigment concentrations and total dissolved solids in red table beets. J. Am. Soc. Hortic. Sci. 1990, 115, 165–169. [Google Scholar] [CrossRef]
- Slatnar, A.; Stampar, F.; Veberic, R.; Jakopic, J. HPLC-MSn Identification of betalain profile of different beetroot (Beta vulgaris L.ssp. vulgaris) parts and cultivars. J. Food Sci. 2015, 80, 1952–1958. [Google Scholar] [CrossRef]
- Czapski, J.; Mikołajczyk, K.; Kaczmarek, M. Relationship between antioxidant capacity of red beet juice and contents of its betalain pigments. Pol. J. Food Nutr. Sci. 2009, 59, 119–122. [Google Scholar]
- Wruss, J.; Waldenberger, G.; Huemer, S.; Uygun, P.; Lanzerstorfer, P.; Müller, U.; Höglinger, O.; Weghuber, J. Compositional characteristics of commercial beetroot products and beetroot juice prepared from seven beetroot varieties grown in Upper Austria. J. Food Compos. Anal. 2015, 42, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Zhuchenko, A.A. Adaptive Potential of Cultivated Plants (Genetic and Ecological Bases), 1st ed.; Shtiintsa: Chisinau, Moldova, 1988; pp. 17–26. [Google Scholar]
- Burenin, V.I. Genetic Resources of Genus Beta L. (Beet); Scientific Edition: Saint-Petersburg, Russia, 2007; pp. 156–158. [Google Scholar]
- Lee, Y.N.; Wiley, R.C. Betalaine yield from a continuous solid-liquid extraction system as influenced by raw product, post-harvest and processing variables. J. Food Sci. 1981, 46, 421–424. [Google Scholar] [CrossRef]
- Timakova, L.N.; Borisov, V.A.; Filroze, N.A.; Uspenskaya, O.N.; Sokolova, L.M. Assessment of the quality of beet varieties in the Moscow region. Potato Veg. 2020, 7, 28–32. [Google Scholar] [CrossRef]
- Sokolova, D.V. Environmental and geographic study of betanin accumulation in promising red beet accessions from the VIR Collection. Proc. Appl. Bot. Genet. Breed. 2019, 180, 66–74. [Google Scholar] [CrossRef]
- Kishima, Y.; Shimaya, A.; Adachi, T. Evidence that blue light induces betalain pigmentation in Portulaca callus. Plant Cell Tissue Organ Cult. 1995, 43, 67–70. [Google Scholar] [CrossRef]
- Schliemann, W.; Strack, D. Intramolecular stabilization of acylated betacyanins. Phytochemistry 1998, 49, 585–588. [Google Scholar] [CrossRef]
- Herbach, K.; Stintzing, F.C.; Carle, R. Thermal degradation of betacyanins in juices from purple pitaya [Hylocereus polyrhizus (Weber) Britton & Rose]. monitored by high-performance liquid chromatography–tandem mass spectometric analyses. Eur. Food Res. Technol. 2004, 219, 377–385. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Wagner, A.E.; Schini-Kerth, V.B.; Rimbach, G. Betanin—A food colorant with biological activity. Mol. Nutr. Food Res. 2015, 59, 36–47. [Google Scholar] [CrossRef]
- Wolyn, D.J.; Gabelman, W.H. Effects of planting and harvest date on betalain pigment concentrations in three table beet genotypes. Hortic. Sci. 1986, 21, 1339–1340. [Google Scholar]
- Sadowska-Bartosz, I.; Bartosz, G. Biological properties and applications of betalains. Molecules 2021, 26, 2520. [Google Scholar] [CrossRef]
- Stintzing, F.C.; Carle, R. Betalains—Emerging prospects for food scientists. Trends Food Sci. Technol. 2007, 18, 514–525. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
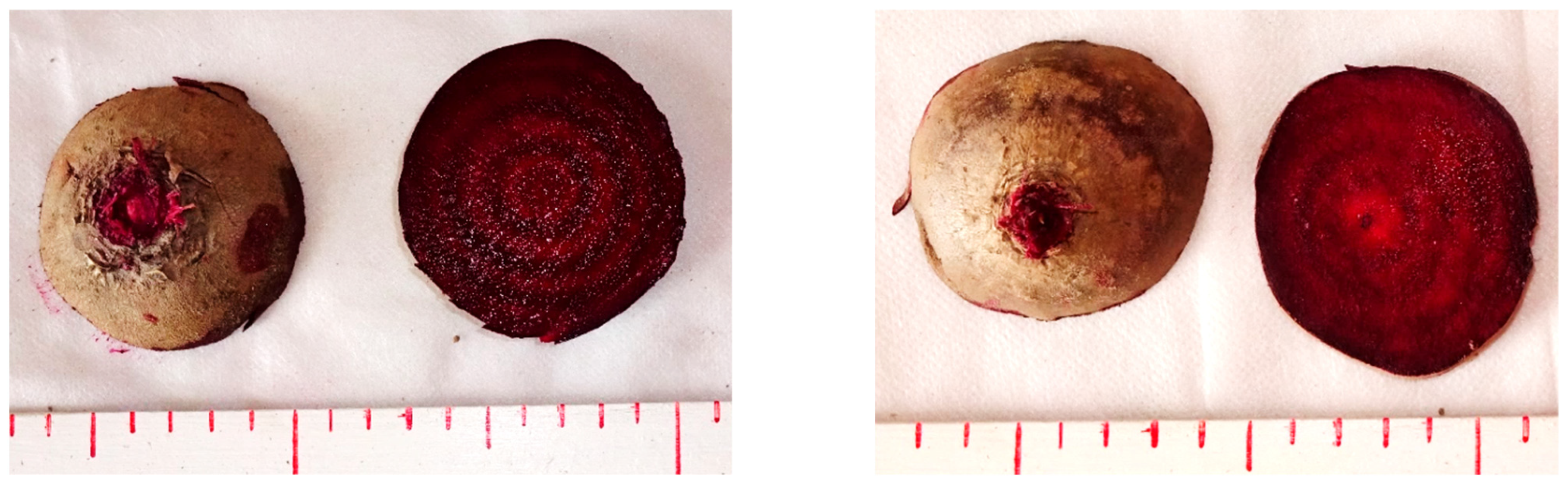{kind=link}
{kind=link}
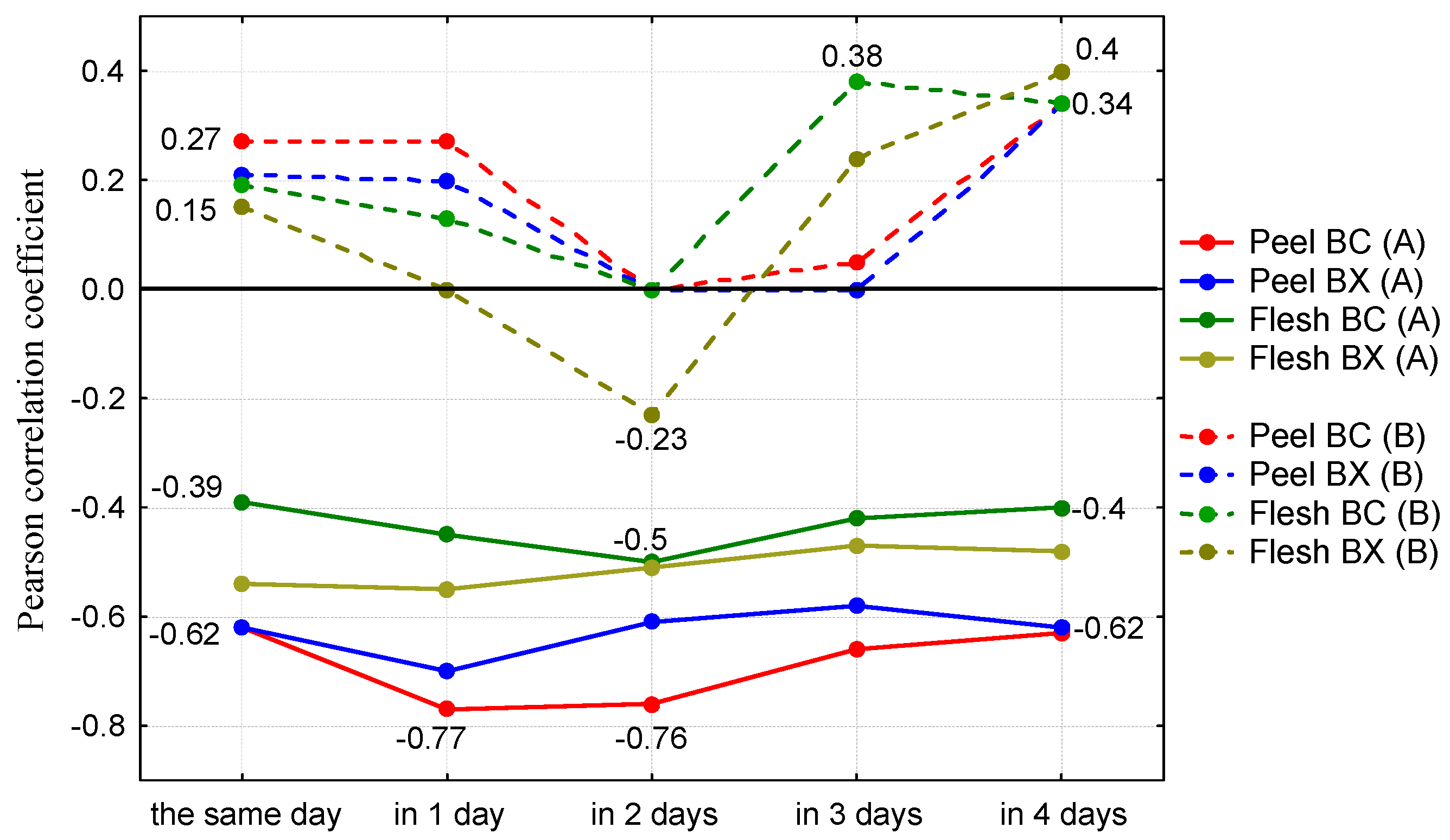{kind=link}
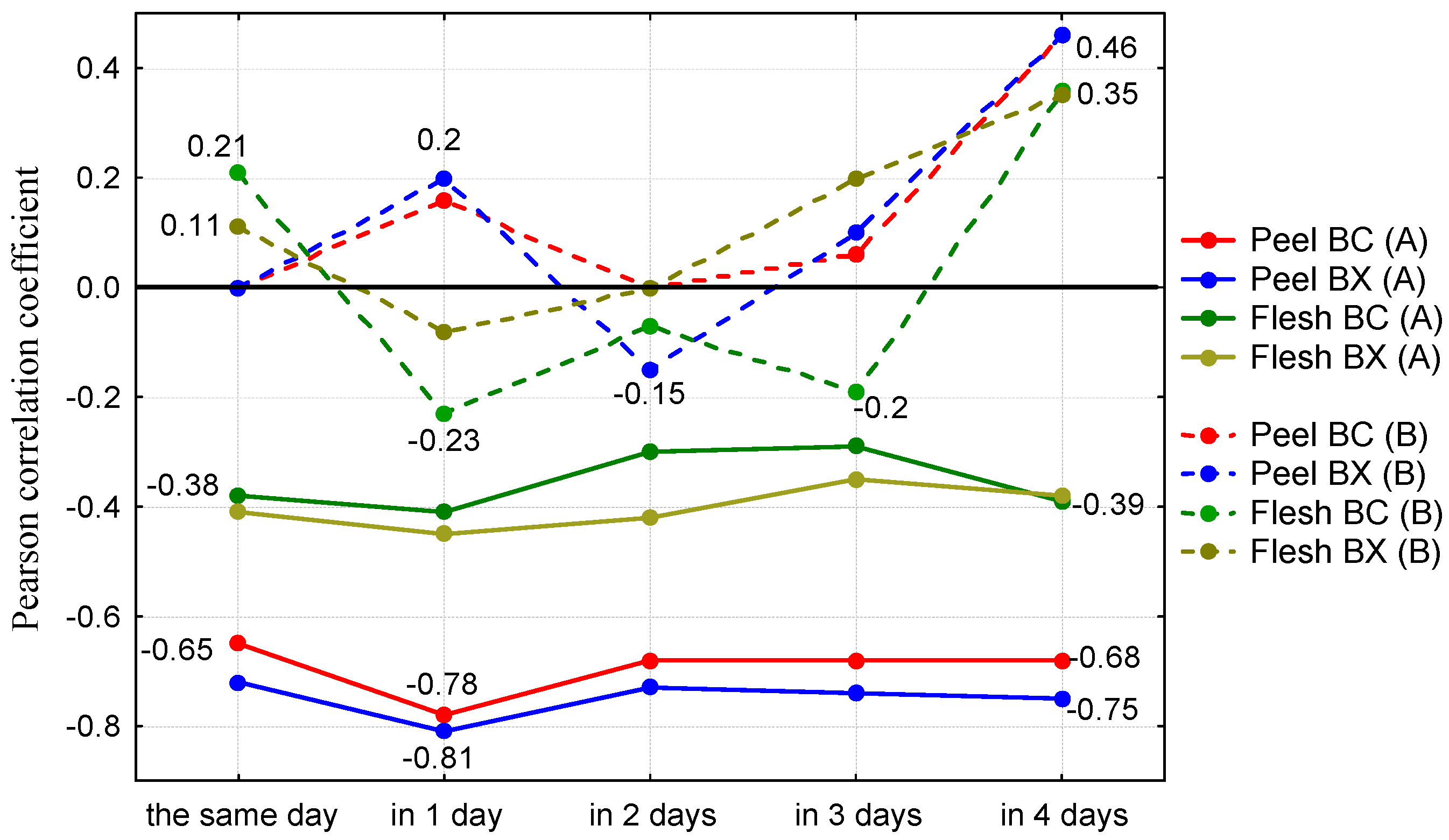{kind=link}
| VIR Catalogue No. | Accession Name | Origin | Photosynthetic Surface Area, cm2 | Root Color Group | Yield, kg/10 m² | Root Weight, g ** | |
|---|---|---|---|---|---|---|---|
| Mean ± SE | % CV | ||||||
| 2011 | Betterowe Potagere *** | Algeria | 2070.0 | 1 | 33.6 | 263.2 ± 100.9 | 96.3 |
| 3677 | Detroit rubinovy | Russia | 700.10 | 1 | 24.5 | 136.0 ± 36.2 | 65.2 |
| 3151 | Bordo odnosemyannaya | Russia | 868.47 | 1 | 31.2 | 121.5 ± 15.1 | 30.4 |
| 3698 | Russkaya odnosemyannaya | Russia | 873.14 | 1 | 28.9 | 158.5 ± 36.4 | 56.2 |
| 3206 | Joijaj | Lithuania | 561.46 | 1 | 16.5 | 91.8 ± 11.2 | 27.2 |
| 3207 | Red Cloud | Netherlands | 693.58 | 1 | 24.3 | 135.0 ± 19.6 | 35.5 |
| 3204 | Rubidius *** | Hungary | 884.00 | 1 | 23.5 | 130.7 ± 34.7 | 65.1 |
| 3209 | Mona | Russia | 1685.81 | 1 | 22.3 | 123.8 ± 25.3 | 45.6 |
| 1967 | Kubanskaya borshchevaya | Russia | 1041.25 | 2 | 15.0 | 83.2 ± 11.7 | 14.0 |
| 3201 | Long Canner *** | Botswana | 1292.00 | 2 | 30.9 | 171.6 ± 69.6 | 90.7 |
| 3105 | Dvusemyannaya 4-53 | Ukraine | 1143.89 | 2 | 10.5 | 47.3 ± 6.8 | 25.2 |
| 3880 | Boldor | Netherlands | 840.13 | 3 | 31.4 | 174.3 ± 25.8 | 36.3 |
| - | L1 yellow * | Russia | 707.69 | 3 | 25.5 | 141.5 ± 31.7 | 54.9 |
| 3881 | Avalanch | Netherlands | 785.94 | 4 | 24.6 | 141.0 ± 27.3 | 22.2 |
| - | L1 white * | Russia | 555.21 | 4 | 25.2 | 140.0 ± 46.5 | 61.4 |
| Mean ± SD | 980.2 ± 422.2 | 23.9 ± 7.0 | 137.3 ± 12.4 | ||||
| LSD05 | 238.7 | 4.6 | 29.9 | ||||
| Root Color Groups | Peel Pigments | Flesh Pigments | ||||||
|---|---|---|---|---|---|---|---|---|
| BC * | BX ** | Ʃ | BC | BX | Ʃ | |||
| 1 |  | Mean ± SE | 889.0 ± 58.7 | 357.1 ± 23.5 | 1246.2 ± 81.4 | 593.1 ± 62.7 | 225.9 ± 27.3 | 819.1 ± 89.1 |
| Min ÷ Max | 647.2 ÷ 1077.1 | 270.8 ÷ 435.9 | 941.1 ÷ 1498.6 | 386.7 ÷ 824.7 | 143.1 ÷ 354.1 | 529.7 ÷ 1178.8 | ||
| %CV | 18.7 | 23.5 | 18.4 | 29.1 | 32.2 | 30.8 | ||
| 2 |  | Mean ± SE | 665.9 ± 163.8 a | 318.7 ± 33.4 | 984.6 ± 195.2 a | 450.5 ± 143.6 a | 188.0 ± 48.9 a | 638.5 ± 189.2 a |
| Min ÷ Max | 384.1 ÷ 951.5 | 178.9 ÷ 385.1 | 563.0 ÷ 1336.6 | 241.8 ÷ 725.6 | 128.1 ÷ 184.9 | 392.7 ÷ 1010.5 | ||
| %CV | 42.6 | 18.1 | 34.3 | 55.2 | 45.1 | 51.3 | ||
| 3 |  | Mean ± SE | 83,8 ± 25.1 | 256.0 ± 65.1 | 339.8 ± 90.1 | 20,1 ± 10.7 | 33.3 ± 8.0 | 53.4 ± 18.3 |
| Min ÷ Max | 58.7 ÷ 108.8 | 191.0 ÷ 321.1 | 249.7 ÷ 430.0 | 9.4 ÷ 30.8 | 25.3 ÷ 41.3 | 34.7 ÷ 72.1 | ||
| %CV | 42.3 | 35.9 | 37.5 | 75.4 | 34.0 | 49.65 | ||
| 4 |  | Mean | not detected | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokolova, D.V.; Shvachko, N.A.; Mikhailova, A.S.; Popov, V.S. Betalain Content and Morphological Characteristics of Table Beet Accessions: Their Interplay with Abiotic Factors. Agronomy 2022, 12, 1033. https://doi.org/10.3390/agronomy12051033
Sokolova DV, Shvachko NA, Mikhailova AS, Popov VS. Betalain Content and Morphological Characteristics of Table Beet Accessions: Their Interplay with Abiotic Factors. Agronomy. 2022; 12(5):1033. https://doi.org/10.3390/agronomy12051033
Chicago/Turabian StyleSokolova, Diana V., Natalia A. Shvachko, Aleksandra S. Mikhailova, and Vitaliy S. Popov. 2022. "Betalain Content and Morphological Characteristics of Table Beet Accessions: Their Interplay with Abiotic Factors" Agronomy 12, no. 5: 1033. https://doi.org/10.3390/agronomy12051033