


Stem Characteristic Associated with Lodging Resistance of Rice Changes with Varied Alternating Drought and Flooding Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Condition and Plant Material
2.2. Treatments and Experimental Design
2.3. Shoot Morphological and Mechanical Properties
2.3.1. Shoot Morphological Properties
2.3.2. Physical Properties of Culms
2.4. Self-Weight Mass Moment and Safety Factor
2.4.1. Self-Weight Mass Moment of the Individual Tiller
2.4.2. Safety Factor against Stem Breakage
2.5. Analysis of Stem Chemical Components in Rice Culms
2.5.1. Cellulose Content and Lignin Content of Culm
2.5.2. N Content in Leaf Blade
2.5.3. Ash Content of Culms
2.6. Gap Fraction
2.7. Statistical Analyses
3. Results
3.1. Weather Conditions
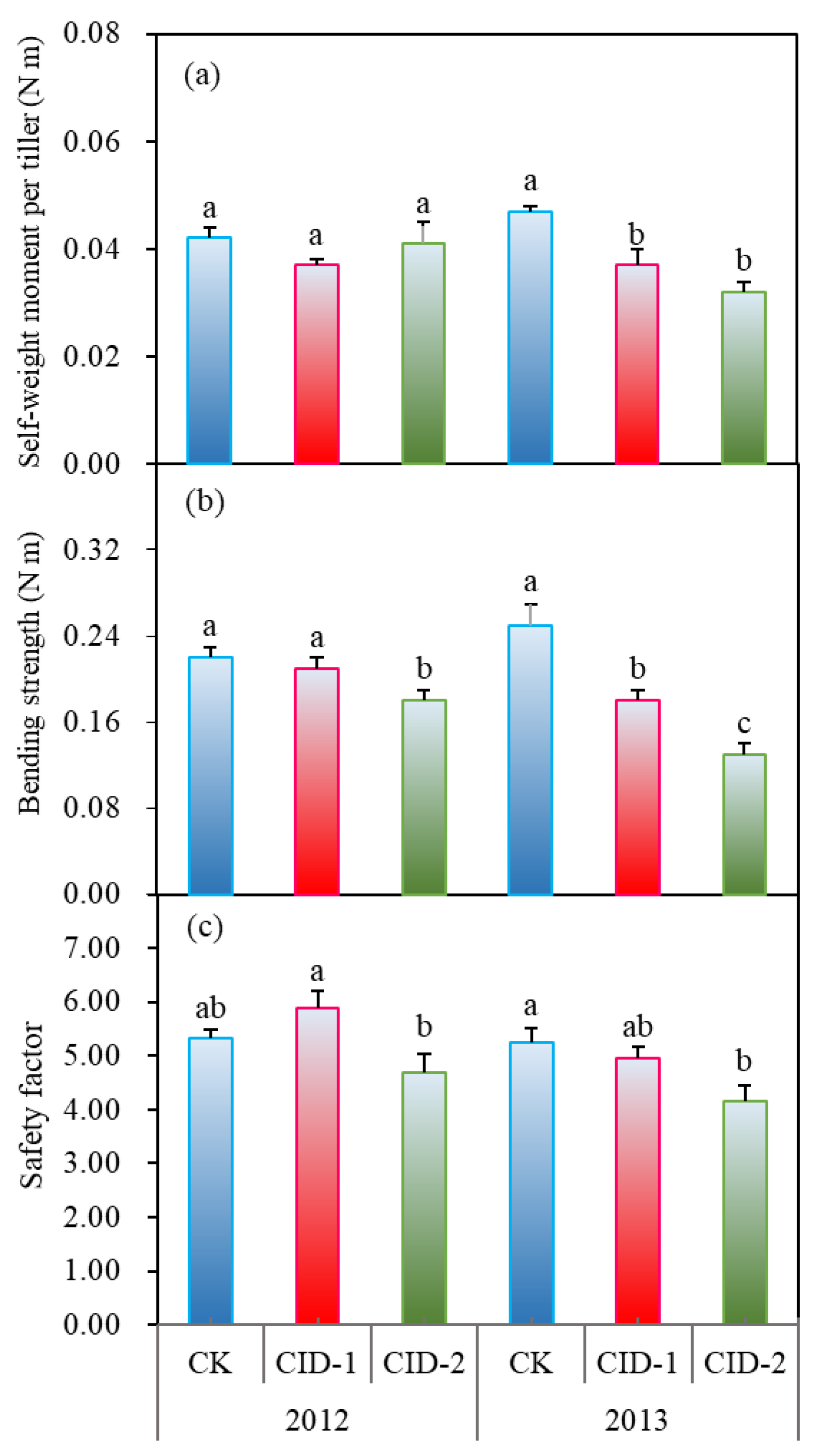
3.2. Bending Stress of Culms, Section Modulus of Culms, and the Safety Factor against Stem Breakage
3.3. Components of the Lower Culms and N Content in Flag Leaves
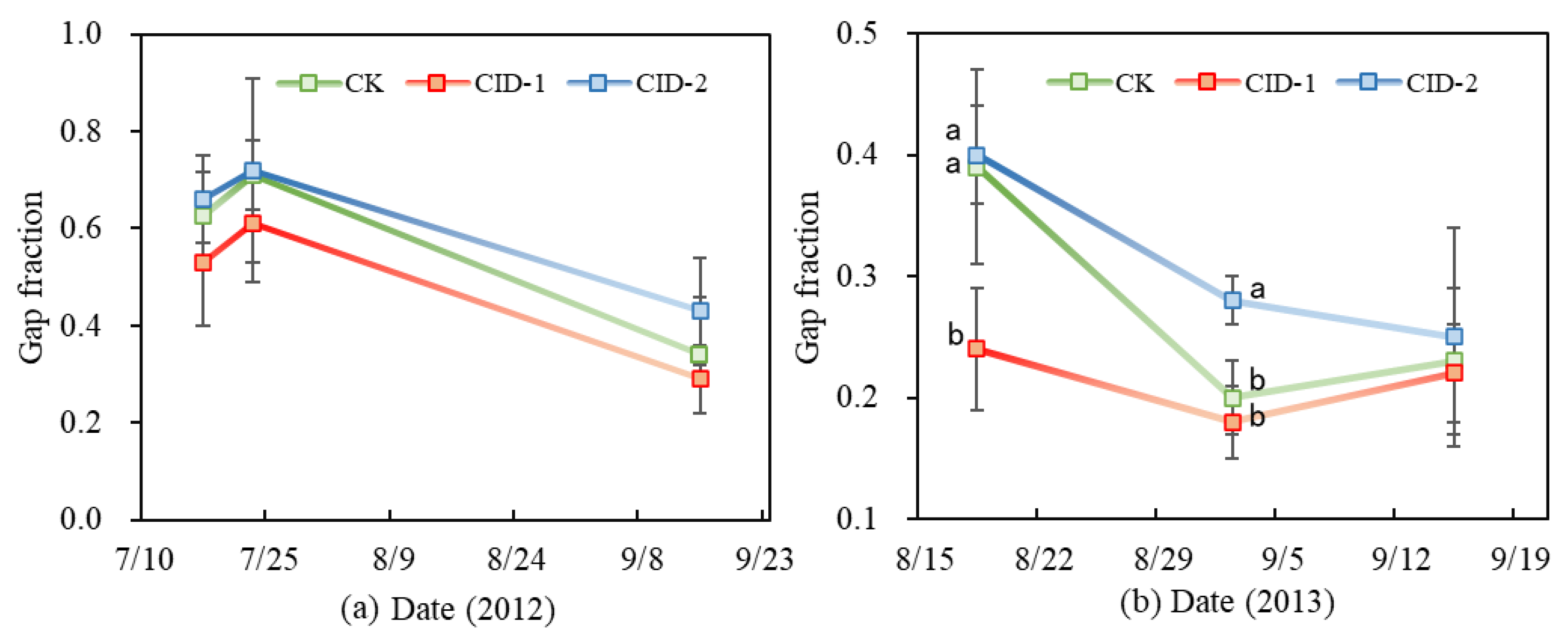
3.4. Gap Fraction (fGPAR)
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shah, L.; Yahya, M.; Shah, S.M.A.; Nadeem, M.; Ali, A.; Ali, A.; Wang, J.; Riaz, M.W.; Rehman, S.; Wu, W.; et al. Improving Lodging Resistance: Using Wheat and Rice as Classical Examples. Int. J. Mol. Sci. 2019, 20, 4211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghuman, L.; Ram, H. Enhancing Wheat (Triticum aestivum, L.) Grain Yield and Quality by Managing Lodging with Growth Regulators under Different Nutrition Levels. J. Plant Nutr. 2021, 44, 1916–1929. [Google Scholar] [CrossRef]
- Setter, T.L.; Laureles, E.V.; Mazaredo, A.M. Lodging Reduces Yield of Rice by Self-Shading and Reductions in Canopy Photosynthesis. Field Crop. Res. 1997, 49, 95–106. [Google Scholar] [CrossRef]
- Kashiwagi, T.; Hirotsu, N.; Ujiie, K.; Ishimaru, K. Lodging Resistance Locus Prl5 Improves Physical Strength of the Lower Plant Part under Different Conditions of Fertilization in Rice (Oryza sativa, L.). Field Crop. Res. 2010, 115, 107–115. [Google Scholar] [CrossRef]
- Zhang, J.; Li, G.; Song, Y.; Liu, Z.; Yang, C.; Tang, S.; Zheng, C.; Wang, S.; Ding, Y. Lodging Resistance Characteristics of High-Yielding Rice Populations. Field Crop. Res. 2014, 161, 64–74. [Google Scholar] [CrossRef]
- Robertson, D.J.; Brenton, Z.W.; Kresovich, S.; Cook, D.D. Maize Lodging Resistance: Stalk Architecture Is a Stronger Predictor of Stalk Bending Strength than Chemical Composition. Biosyst. Eng. 2022, 219, 124–134. [Google Scholar] [CrossRef]
- Wang, X.; Xu, L.; Li, X.; Yang, G.; Wang, F.; Peng, S. Grain Yield and Lodging-Related Traits of Ultrashort-Duration Varieties for Direct-Seeded and Double-Season Rice in Central China. J. Integr. Agric. 2022, 1–23. [Google Scholar] [CrossRef]
- Peng, Y.; Hu, Y.; Qian, Q.; Ren, D. Progress and Prospect of Breeding Utilization of Green Revolution Gene Sd1 in Rice. Agriculture 2021, 11, 611. [Google Scholar] [CrossRef]
- Chen, L.; Yi, Y.; Wang, W.; Zeng, Y.; Tan, X.; Wu, Z.; Chen, X.; Pan, X.; Shi, Q.; Zeng, Y. Innovative Furrow Ridging Fertilization under a Mechanical Direct Seeding System Improves the Grain Yield and Lodging Resistance of Early Indica Rice in South China. Field Crop. Res. 2021, 270, 108184. [Google Scholar] [CrossRef]
- Zhang, W.; Yao, X.; Duan, X.; Liu, Q.; Tang, Y.; Li, J.; Li, G.; Ding, Y.; Liu, Z. Foliar Application Uniconazole Enhanced Lodging Resistance of Hybrid Indica Rice by Altering Basal Stem Quality under Poor Light Stress. Agron. J. 2022, 114, 524–544. [Google Scholar] [CrossRef]
- Guo, X.; Huang, S.; Wang, Z.; Wang, F.; Chen, B. Impact of Different Irrigation Methods on Resistance of Rice Against Bending and Breaking. J. Irrig. Drain. 2017, 36, 1–5. [Google Scholar]
- Wu, H.; Liu, D.; Zhong, M.; Wang, Y.Y. Effects of Water Management and Potash Application on Grain Yield and Lodging Resistance of Rice. Crops 2019, 1, 127–133. [Google Scholar]
- Li, Q.; Fu, C.; Liang, C.; Ni, X.; Zhao, X.; Chen, M.; Ou, L. Crop Lodging and The Roles of Lignin, Cellulose, and Hemicellulose in Lodging Resistance. Agronomy 2022, 12, 1795. [Google Scholar] [CrossRef]
- Kaveri, K.; Kumar, R.; Ismail, A.M. Elongation Ability and Non-Structural Carbohydrate Levels in Relation to Submergence Tolerance in Rice. Plant Sci. 2005, 168, 131–136. [Google Scholar]
- Ghoneim, A. Soil Nutrients Availability, Rice Productivity and Water Saving under Deficit Irrigation Conditions. J. Plant Prod. 2020, 11, 7–16. [Google Scholar] [CrossRef]
- Hamoud, Y.A.; Guo, X.; Wang, Z.; Shaghaleh, H.; Chen, S.; Hassan, A.; Bakour, A. Effects of Irrigation Regime and Soil Clay Content and Their Interaction on the Biological Yield, Nitrogen Uptake and Nitrogen-Use Efficiency of Rice Grown in Southern China. Agric. Water Manag. 2019, 213, 934–946. [Google Scholar] [CrossRef]
- Niu, Y.; Chen, T.; Zhao, C.; Zhou, M. Improving Crop Lodging Resistance by Adjusting Plant Height and Stem Strength. Agronomy 2021, 11, 2421. [Google Scholar] [CrossRef]
- Zhang, W.; Wu, L.; Wu, X.; Ding, Y.; Li, G.; Li, J.; Weng, F.; Liu, Z.; Tang, S.; Ding, C.; et al. Lodging Resistance of Japonica Rice (Oryza Sativa, L.): Morphological and Anatomical Traits Due to Top-Dressing Nitrogen Application Rates. Rice 2016, 9, 31. [Google Scholar] [CrossRef] [Green Version]
- Kashiwagi, T. Novel QTL for Lodging Resistance, PRL4, Improves Physical Properties with High Non-Structural Carbohydrate Accumulation of Basal Culms in Rice (Oryza sativa, L.). Euphytica 2022, 218, 1–12. [Google Scholar] [CrossRef]
- Yano, K.; Ookawa, T.; Aya, K.; Ochiai, Y.; Hirasawa, T.; Ebitani, T.; Takarada, T.; Yano, M.; Yamamoto, T.; Fukuoka, S.; et al. Isolation of a Novel Lodging Resistance QTL Gene Involved in Strigolactone Signaling and Its Pyramiding with a QTL Gene Involved in Another Mechanism. Mol. Plant 2015, 8, 303–314. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Chen, Q. Rice Research in China in the Early 21st Century. Chinese Rice Res. Newsl. 2000, 8, 14–16. [Google Scholar]
- Hanjra, M.A.; Qureshi, M.E. Global Water Crisis and Future Food Security in an Era of Climate Change. Food Policy 2010, 35, 365–377. [Google Scholar] [CrossRef]
- Bouman, B.A.M. A Conceptual Framework for the Improvement of Crop Water Productivity at Different Spatial Scales. Agric. Syst. 2007, 93, 43–60. [Google Scholar] [CrossRef]
- Oliver, V.; Cochrane, N.; Magnusson, J.; Brachi, E.; Monaco, S.; Volante, A.; Courtois, B.; Vale, G.; Price, A.; Teh, Y.A. Effects of Water Management and Cultivar on Carbon Dynamics, Plant Productivity and Biomass Allocation in European Rice Systems. Sci. Total Environ. 2019, 685, 1139–1151. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Shu, K.; Zhu, T.; Wang, L.; Liu, X.; Cai, W.; Qi, Z.; Feng, S. Effects of Alternate Wetting and Drying Irrigation on Yield, Water and Nitrogen Use, and Greenhouse Gas Emissions in Rice Paddy Fields. J. Clean. Prod. 2022, 349, 131487. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, J.; Hamoud, Y.A.; Wang, Y.; Qiu, R.; Agathokleous, E.; Hong, C.; Shaghaleh, H. Natural 15N Abundance as an Indicator of Nitrogen Utilization Efficiency in Rice under Alternate Wetting and Drying Irrigation in Soils with High Clay Contents. Sci. Total Environ. 2022, 838, 156528. [Google Scholar] [CrossRef] [PubMed]
- Song, T.; Das, D.; Hu, Q.; Yang, F.; Zhang, J. Alternate Wetting and Drying Irrigation and Phosphorus Rates Affect Grain Yield and Quality and Heavy Metal Accumulation in Rice. Sci. Total Environ. 2021, 752, 141862. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Zhu, Z.; Guo, Q.; Ma, H.; Zhu, L. Alternate Wetting and Drying Irrigation-Mediated Changes in the Growth, Photosynthesis and Yield of the Medicinal Plant Tulipa Edulis. Ind. Crop. Prod. 2015, 66, 81–88. [Google Scholar] [CrossRef]
- Shao, G.; Deng, S.; Liu, N.; Yu, S.E.; Wang, M.H.; She, D.L. Effects of Controlled Irrigation and Drainage on Growth, Grain Yield and Water Use in Paddy Rice. Eur. J. Agron. 2014, 53, 1–9. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Niu, H. Effects of Temperature, Precipitation and Carbon Dioxide Concentrations on the Requirements for Crop Irrigation Water in China under Future Climate Scenarios. Sci. Total Environ. 2019, 656, 373–387. [Google Scholar] [CrossRef]
- Wang, Z.; Shao, G.; Lu, J.; Zhang, K.; Gao, Y.; Ding, J. Effects of Controlled Drainage on Crop Yield, Drainage Water Quantity and Quality: A Meta-Analysis. Agric. Water Manag. 2020, 239, 106253. [Google Scholar] [CrossRef]
- Yu, Y.; Xu, J.; Zhang, P.; Meng, Y.; Xiong, Y. Controlled Irrigation and Drainage Reduce Rainfall Runoff and Nitrogen Loss in Paddy Fields. Int. J. Environ. Res. Public Health 2021, 18, 3348. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Zhang, Z.; Hao, S.; Chen, W.; Pan, Y.; Wang, Z. Agronomic Growth Performance of Super Rice under Water-Saving Irrigation Methods with Different Water-Controlled Thresholds in Different Growth Stages. Agronomy 2020, 10, 293. [Google Scholar] [CrossRef] [Green Version]
- Chu, G.; Chen, T.; Chen, S.; Xu, C.; Wang, D.; Zhang, X. Agronomic Performance of Drought-Resistance Rice Cultivars Grown under Alternate Wetting and Drying Irrigation Management in Southeast China. Crop J. 2018, 6, 482–494. [Google Scholar] [CrossRef]
- Li, H.; Mei, X.; Wang, J.; Huang, F.; Hao, W.; Li, B. Drip Fertigation Significantly Increased Crop Yield, Water Productivity and Nitrogen Use Efficiency with Respect to Traditional Irrigation and Fertilization Practices: A Meta-Analysis in China. Agric. Water Manag. 2021, 244, 106534. [Google Scholar] [CrossRef]
- Guo, X.; Zhen, B.; Wang, Z. Increasing Lodging Resistance Performance of Rice by Alternating Drought and Flooding Stress. Trans. Chin. Soc. Agric. Eng. 2013, 29, 130–135. [Google Scholar]
- Loreti, E.; van Veen, H.; Perata, P. Plant Responses to Flooding Stress. Curr. Opin. Plant Biol. 2016, 33, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.I.R.; Trivellini, A.; Chhillar, H.; Chopra, P.; Ferrante, A.; Khan, N.A.; Ismail, A.M. The Significance and Functions of Ethylene in Flooding Stress Tolerance in Plants. Environ. Exp. Bot. 2020, 179, 104188. [Google Scholar] [CrossRef]
- Bashar, K.K.; Tareq, M.Z.; Amin, M.R.; Honi, U.; Ul-Arif, M.T.; Sadat, M.A.; Mosaddeque Hossen, Q.M. Phytohormone-Mediated Stomatal Response, Escape and Quiescence Strategies in Plants under Flooding Stress. Agronomy 2019, 9, 43. [Google Scholar] [CrossRef] [Green Version]
- Shao, G.; Cui, J.; Yu, S.; Lu, B.; Brian, B.J.; Ding, J.; She, D. Impacts of Controlled Irrigation and Drainage on the Yield and Physiological Attributes of Rice. Agric. Water Manag. 2015, 149, 156–165. [Google Scholar] [CrossRef]
- Crook, M.J.; Ennos, A.R. Stem and Root Characteristics Associated with Lodging Resistance in Four Winter Wheat Cultivars. J. Agric. Sci. 1994, 123, 167–174. [Google Scholar] [CrossRef]
- Hoshikawa, K.A.; Wang, S.B. Studies on Lodging in Rice Plants. I. A General Observation on Lodged Rice Culms. Jpn. J. Crop Sci. 1990, 59, 809–814. [Google Scholar] [CrossRef] [Green Version]
- Shao, Z.; Mu, H.; Zhao, Y. Impacts of Ozone Stress on Stem Lodging Resistance of Rice Genotypes with Varying Ozone Sensitivities. J. Agro-Environ. Sci. 2017, 36, 1703–1711. [Google Scholar]
- Shi, S.; He, F. Analysis and Detection of Pulping and Papermaking; Chinese Light Ind. Press: Beijing, China, 2003. [Google Scholar]
- Song, W.; Wang, J.; Wang, M.; Wang, J. Study on Main Chemical Compositions of Rice Straw in Three Different Areas of Jiangsu Province. Agric. Sci. Technol. 2016, 17, 405–408. [Google Scholar]
- Zhao, X.; Zhou, N.; Lai, S.; Frei, M.; Wang, Y.; Yang, L. Elevated CO2 Improves Lodging Resistance of Rice by Changing Physicochemical Properties of the Basal Internodes. Sci. Total Environ. 2019, 647, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Sato, K.; Wada, K. Internode Elongation of Rice Plant. IV. Effects of Gibberellic Acid at Different Stages of Growth on the Elongation of Internode. Jpn. J. Crop Sci. 1972, 41, 449–453. [Google Scholar] [CrossRef] [Green Version]
- Kamiji, Y.; Hayashi, S.; Horie, T. Influences on Nitrogen Nutrient and Solar Radiation in the Canopy on Length of Lower Internodes of Rice [Oryza sativa]. Jpn. J. Crop Sci. 1993, 62, 164–171. [Google Scholar] [CrossRef]
- Ishimaru, K.; Togawa, E.; Ookawa, T.; Kashiwagi, T.; Madoka, Y.; Hirotsu, N. New Target for Rice Lodging Resistance and Its Effect in a Typhoon. Planta 2008, 227, 601–609. [Google Scholar] [CrossRef]
- Sreeja, R.; Balaji, S.; Arul, L.; Nirmala Kumari, A.; Kannan Bapu, J.R.; Subramanian, A. Association of Lignin and FLEXIBLE CULM 1 (FC1) Ortholog in Imparting Culm Strength and Lodging Resistance in Kodo Millet (Paspalum scrobiculatum, L.). Mol. Breed. 2016, 36, 149. [Google Scholar] [CrossRef]
- Ookawa, T.; Todokoro, Y.; Ishihara, K. Changes in Physical and Chemical Characteristics of Culm Associated with Lodging Resistance in Paddy Rice [Oryza sativa] under Different Growth Conditions and Varietal Difference of Their Changes. Jpn. J. Crop Sci. 1993, 62, 525–533. [Google Scholar] [CrossRef] [Green Version]
- Idris, M.; Hossain, M.M.; Choudhury, F.A. The Effect of Silicon on Lodging of Rice in Presence of Added Nitrogen. Plant Soil 1975, 43, 691–695. [Google Scholar] [CrossRef]
- Dorairaj, D.; Ismail, M.R.; Sinniah, U.R.; Kar Ban, T. Influence of Silicon on Growth, Yield, and Lodging Resistance of MR219, a Lowland Rice of Malaysia. J. Plant Nutr. 2017, 40, 1111–1124. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. Silicon Uptake and Accumulation in Higher Plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Merah, O.; Deléens, E.; Monneveux, P. Grain Yield, Carbon Isotope Discrimination, Mineral and Silicon Content in Durum Wheat under Different Precipitation Regimes. Physiol. Plant. 1999, 107, 387–394. [Google Scholar] [CrossRef]
- Martens, S.N.; Ustin, S.L.; Rousseau, R.A. Estimation of Tree Canopy Leaf Area Index by Gap Fraction Analysis. For. Ecol. Manag. 1993, 61, 91–108. [Google Scholar] [CrossRef]
- Michel, A.J.; Teixeira, E.I.; Brown, H.E.; Dellow, S.J.; Maley, S.; Gillespie, R.N.; Richards, K.K. Water Stress Responses of Three Potato Cultivars. Christch. N. Z. Inst. Plant Food Res. Ltd. 2019, 25, 37. [Google Scholar]
- Zhang, J.; Li, G.; Huang, Q.; Liu, Z.; Ding, C.; Tang, S.; Chen, L.; Wang, S.; Ding, Y.; Zhang, W. Effects of Culm Carbohydrate Partitioning on Basal Stem Strength in a High-Yielding Rice Population. Crop J. 2017, 5, 478–487. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Treatment | T a | J/B | H/F | M/R | |
|---|---|---|---|---|---|
| IT | MT/LT | ||||
| CK | θs~30~70 b | θs~30~90 | θs~30~120 | θs~30~100 | θs~30~60 |
| CID-1 | 0.8θs~θs ~80 | 0.7θs~θs~100 | 0.7θs~θs~100 | 0.8θs~θs~150 | 0.8θs~θs~80 |
| CID-2 | 0.8θs~θs ~100 | 0.7θs~θs~120 | 0.7θs~θs~200 | 0.8θs~θs~200 | 0.8θs~θs~80 |
| Year | Irrigation | Plant Height (cm) | Shoot fw (g) | Outer Diameter of the Major Axis of Internode II (mm) | Outer Diameter of the Minor Axis of Internode II (mm) | Length of Internode II (mm) |
|---|---|---|---|---|---|---|
| 2012 | CK a | 81.3 ± 0.9 a | 25.6 ± 1.2 a | 5.52 ± 0.31 a | 4.45 ± 0.25 a | 7.80 ± 0.63 a |
| CID-1 | 82.3 ± 1.1 a | 21.9 ± 0.2 b | 5.11 ± 0.25 a | 4.52 ± 0.13 a | 7.60 ± 0.41 a | |
| CID-2 | 80.6 ± 2.7 a | 23.0 ± 1.7 ab | 5.09 ± 0.21 a | 4.07 ± 0.12 a | 8.36 ± 0.40 a | |
| 2013 | CK | 81.4 ± 1.8 a | 27.0 ± 0.7 a | 5.69 ± 0.13 a | 5.48 ± 0.12 a | 6.73 ± 0.37 b |
| CID-1 | 80.0 ± 1.9 a | 21.5 ± 1.4 b | 5.26 ± 0.11 a | 4.87 ± 0.13 b | 6.56 ± 0.32 b | |
| CID-2 | 77.8 ± 1.0 a | 19.2 ± 0.9 b | 4.69 ± 0.19 b | 4.28 ± 0.20 c | 8.83 ± 0.92 a | |
| Irrigation | n.s. | ** | * | * | * | |
| Year | n.s. | n.s. | n.s. | n.s. | n.s. | |
| Irrigation × Year | n.s. | n.s. | n.s. | n.s. | n.s. | |
| Irrigation | Position of Internode | Section Modulus (mm3) | Bending Stress (N mm−2) | Bending Strength at Breaking (N m) |
|---|---|---|---|---|
| CK | N2 | 15.41 ± 1.17 a | 16.55 ± 0.49 d | 0.254 ± 0.016 a |
| N3 | 9.37 ± 1.18 bc | 12.17 ± 0.91 cd | 0.112 ± 0.013 cd | |
| N4 | 7.64 ± 1.90 bc | 14.36 ± 1.65 bcd | 0.103 ± 0.015 cd | |
| CID-1 | N2 | 10.96 ± 0.66 b | 17.11 ± 0.47 bcd | 0.186 ± 0.007 b |
| N3 | 8.26 ± 0.87 bc | 13.97 ± 0.38 bcd | 0.116 ± 0.015 c | |
| N4 | 5.94 ± 1.49 c | 18.87 ± 3.02 abcd | 0.104 ± 0.022 cd | |
| CID-2 | N2 | 10.94 ± 1.65 b | 13.03 ± 1.35 abc | 0.137 ± 0.012 c |
| N3 | 7.20 ± 0.68 bc | 15.43 ± 0.55 ab | 0.110 ± 0.011 cd | |
| N4 | 5.93 ± 0.77 c | 11.57 ± 0.76 a | 0.067 ± 0.008 d | |
| Irrigation | * | * | ** | |
| Position of internode | ** | n.s. | ** | |
| Irrigation × Position of internode | n.s. | * | ** | |
| Components of the Lower Culms | Treatment | ||
|---|---|---|---|
| CK | CID-1 | CID-2 | |
| Cellulose (DW%) | 29.6 ± 0.5 a | 30.0 ± 0.4 a | 25.1 ± 0.2 b |
| Lignin (DW%) | 11.7 ± 0.1 b | 9.3 ± 0.2 c | 13.7 ± 0.4 a |
| Ash (DW%) | 14.3 ± 0.1 a | 11.1 ± 0.1 b | 9.9 ± 0.5 c |
| N content in leaf blade (mg g−1) | 11.07 ± 0.16 a | 10.42 ± 0.18 a | 10.75 ± 0.26 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Wang, Z.; Guo, X. Stem Characteristic Associated with Lodging Resistance of Rice Changes with Varied Alternating Drought and Flooding Stress. Agronomy 2022, 12, 3070. https://doi.org/10.3390/agronomy12123070
Liu J, Wang Z, Guo X. Stem Characteristic Associated with Lodging Resistance of Rice Changes with Varied Alternating Drought and Flooding Stress. Agronomy. 2022; 12(12):3070. https://doi.org/10.3390/agronomy12123070
Chicago/Turabian StyleLiu, Jinjing, Zhenchang Wang, and Xiangping Guo. 2022. "Stem Characteristic Associated with Lodging Resistance of Rice Changes with Varied Alternating Drought and Flooding Stress" Agronomy 12, no. 12: 3070. https://doi.org/10.3390/agronomy12123070
APA StyleLiu, J., Wang, Z., & Guo, X. (2022). Stem Characteristic Associated with Lodging Resistance of Rice Changes with Varied Alternating Drought and Flooding Stress. Agronomy, 12(12), 3070. https://doi.org/10.3390/agronomy12123070