Climate Changes Challenges to the Management of Mediterranean Montado Ecosystem: Perspectives for Use of Precision Agriculture Technologies

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
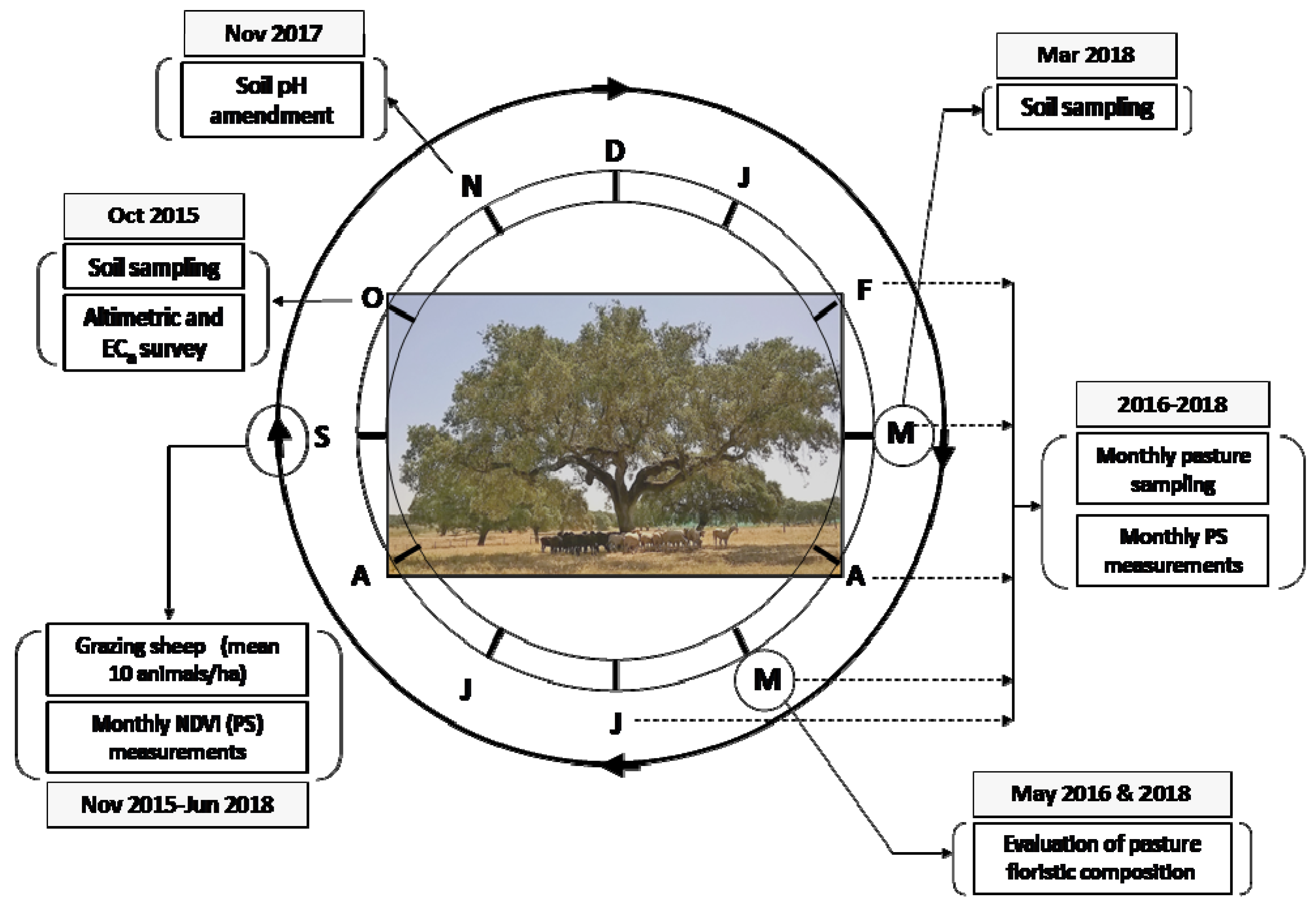
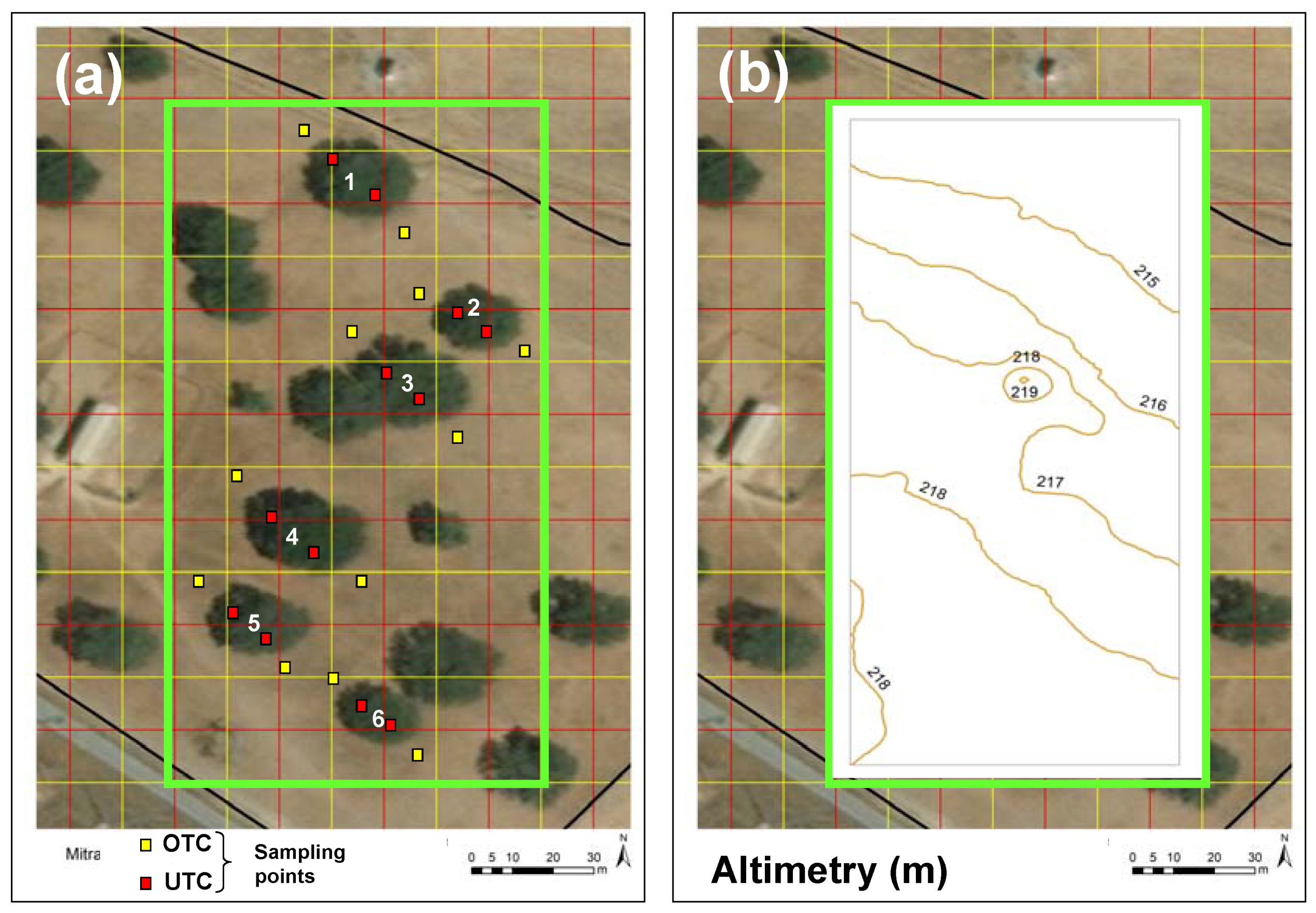
2.1. Experimental Field
2.2. Soil Measurements
2.2.1. Laboratory Determination of Soil Characteristics
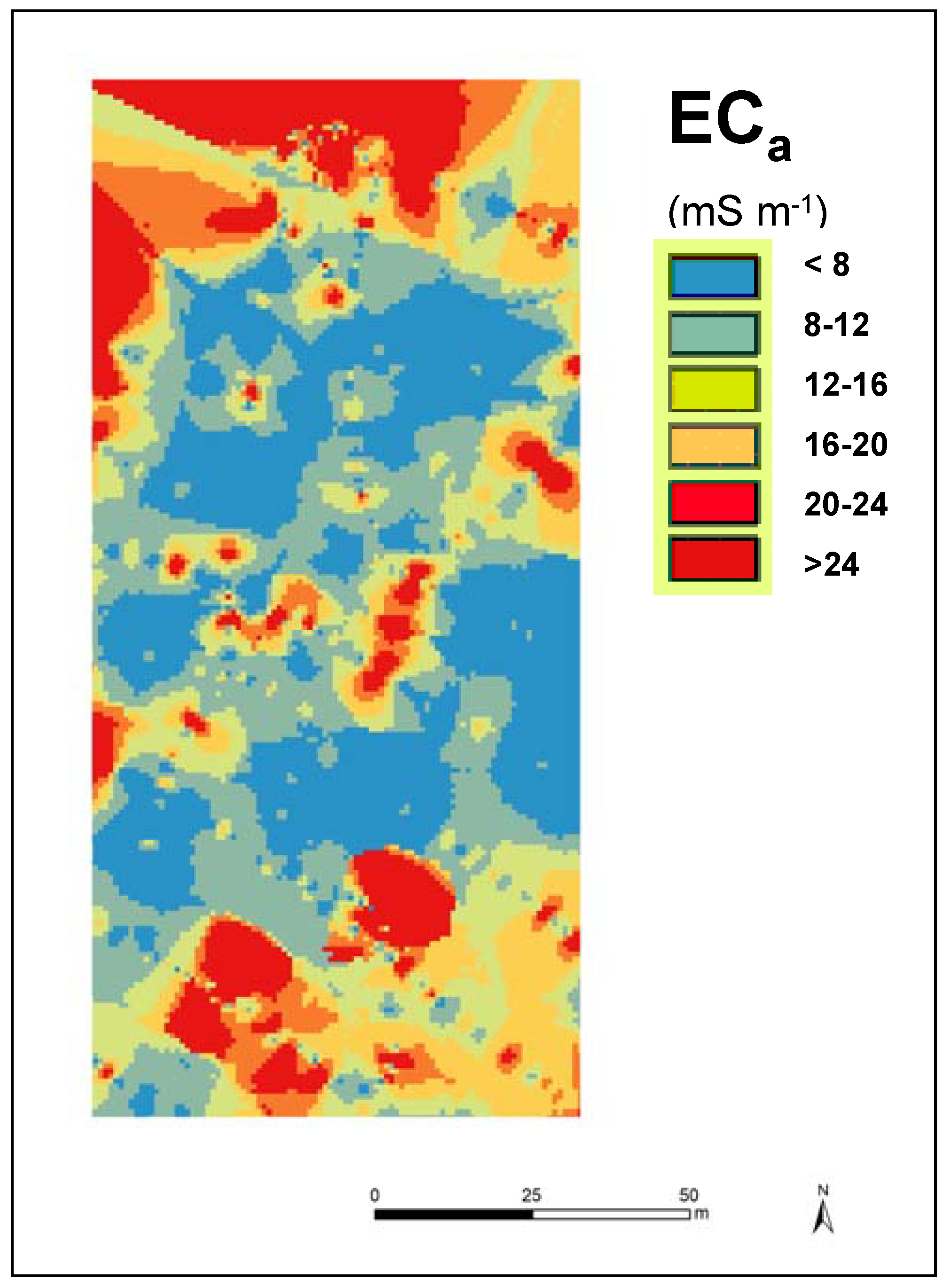
2.2.2. Soil Apparent Electrical Conductivity Survey
2.2.3. Soil Moisture Content and Cone Index Measurements
2.3. Pasture Measurements
2.3.1. Proximal Sensors Measurements
2.3.2. Pasture Samples Collection and Analysis
2.4. Statistical Analysis of the Data
3. Results
3.1. Seasonality and Interannual Climate Variability
3.2. Soil Spatial and Temporal Variability
3.3. Spatial and Temporal Variability of Pasture Productivity and Quality
3.4. Spatial and Temporal Variability of Pasture Floristic Composition
3.5. Spatial Variability of Apparent Soil Electrical Conductivity
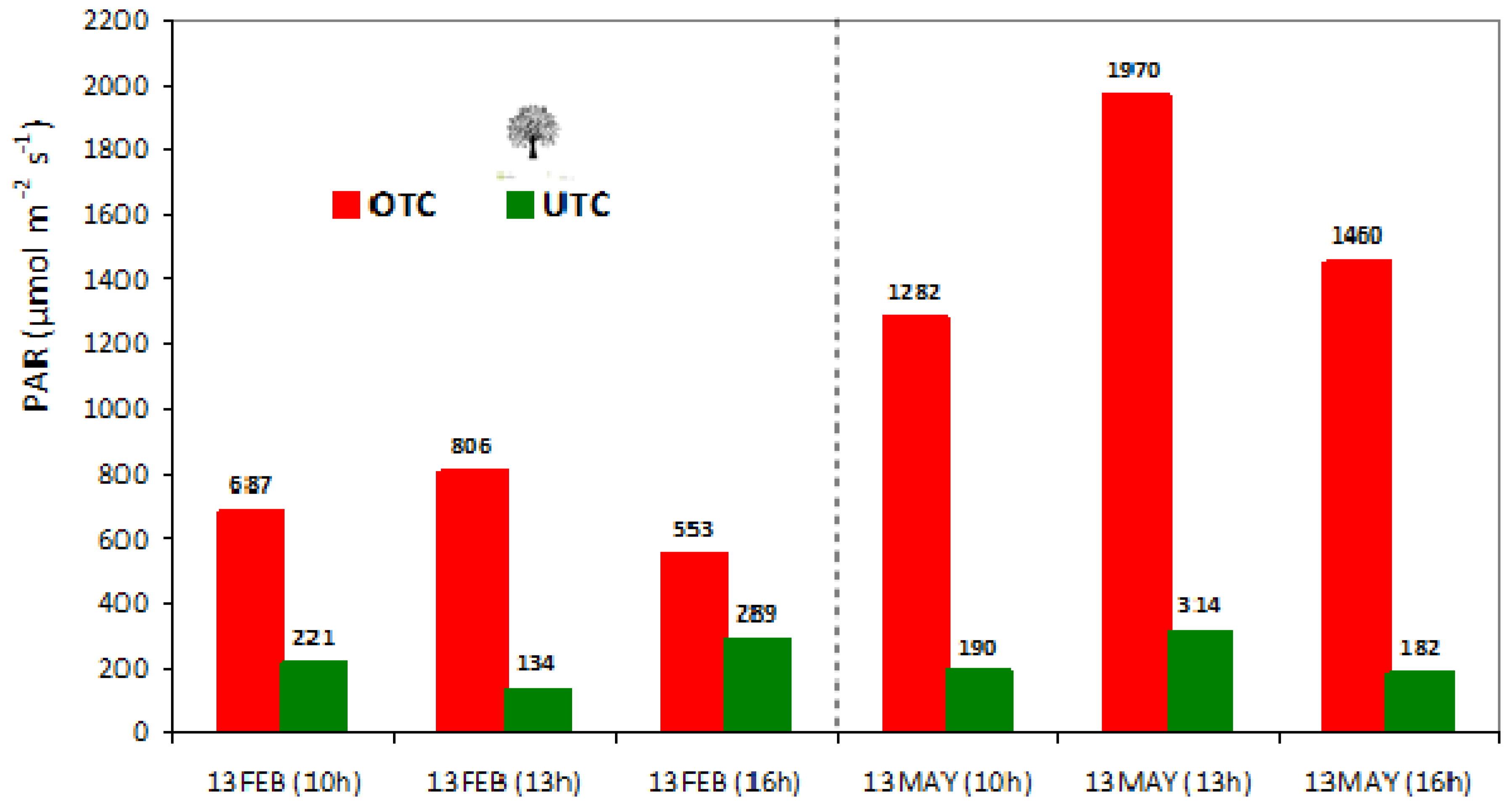
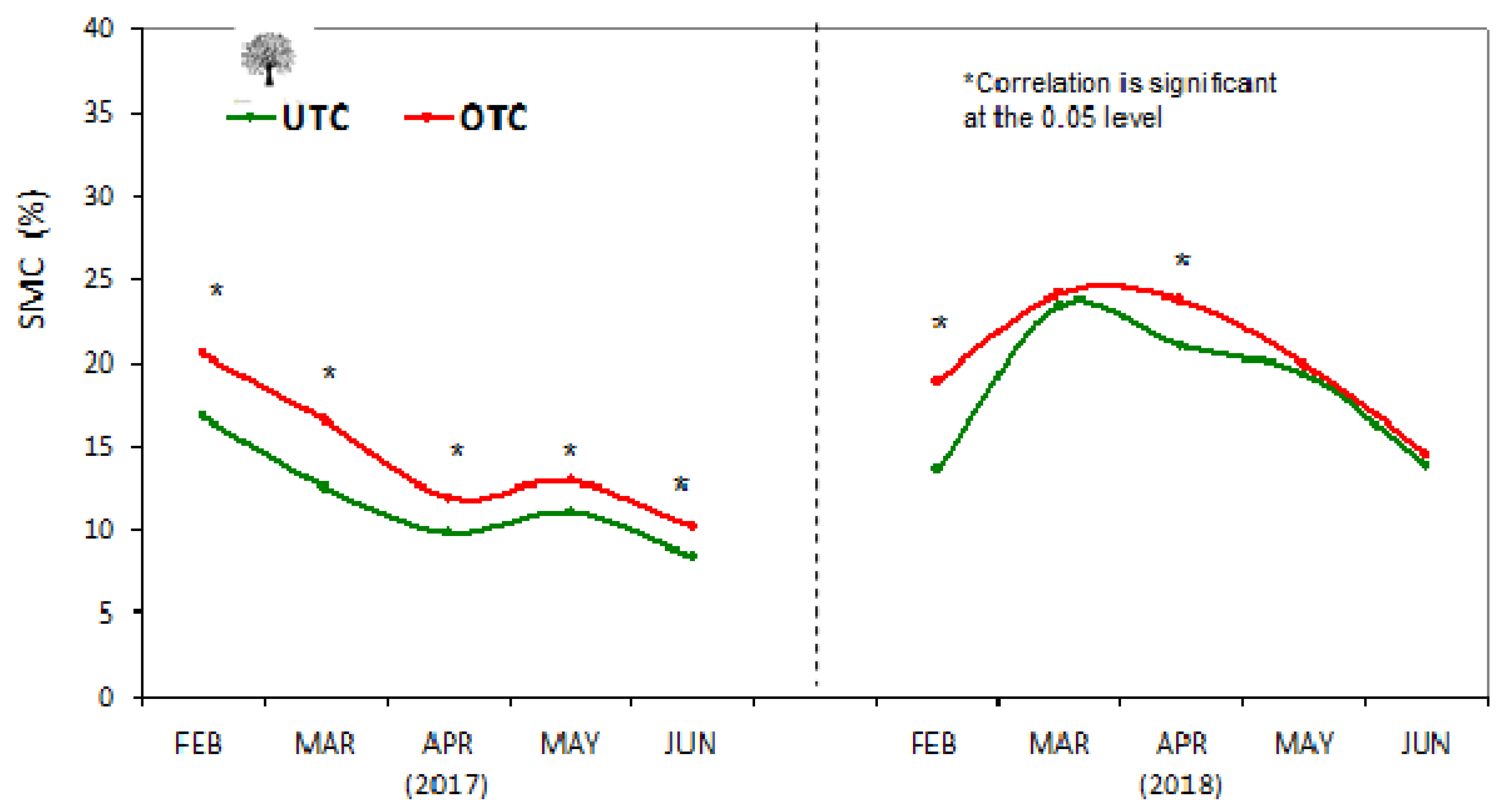
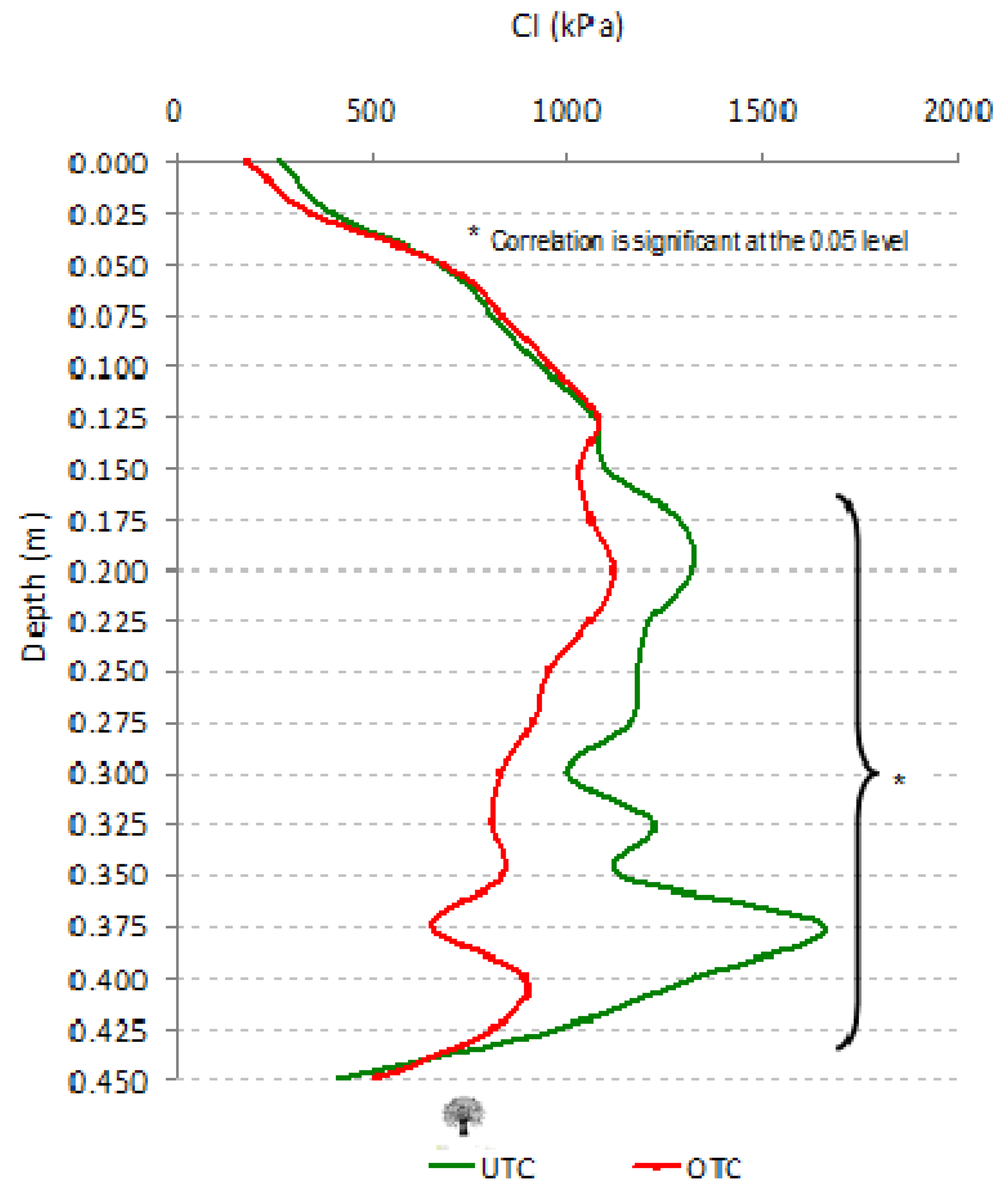
3.6. Effect of Tree Canopy in Photosynthetically Active Radiation (PAR), Surface Temperature (Tir), Soil Moisture Content (SMC), and Cone Index (CI)
3.7. Relationship between Soil and Pasture Characteristics and Parameters Measured by Proximal Sensors
4. Discussion
4.1. Soil Spatial and Temporal Variability
4.2. Spatial and Temporal Variability of Pasture Productivity and Quality
4.3. Spatial and Temporal Variability of Pasture Floristic Composition
4.4. Technologies for Monitoring Soil and Pasture Variability
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carvalho, M.; Goss, M.J.; Teixeira, D. Manganese toxicity in Portuguese Cambisols derived from granitic rocks: Causes, limitations of soil analyses and possible solutions. Revista de Ciências Agrárias 2015, 38, 518–527. [Google Scholar] [CrossRef]
- David, T.S.; Pinto, C.A.; Nadezhdina, N.; Kurz-Besson, C.; Henriques, M.O.; Quilhó, T.; Cermak, J.; Chaves, M.M.; Pereira, J.S.; David, J.S. Root functioning, tree water use and hydraulic redistribution in Quercus suber trees: A modeling approach based on root sap flow. Forest. Ecol. Manag. 2013, 307, 136–146. [Google Scholar] [CrossRef] [Green Version]
- Puche, N.; Senapati, N.; Flechard, C.R.; Klumpp, K.; Kirschbaum, M.U.F.; Chabbi, A. Modeling carbon and water fluxes of managed grasslands: Comparing flux variability and net carbon budgets between grazed and mowed systems. Agronomy 2019, 9, 183. [Google Scholar] [CrossRef] [Green Version]
- Franzluebbers, A.J.; Starks, P.J.; Steiner, J.L. Conservation of soil organic carbon and nitrogen fractions in a Tallgrass prairie in Oklahoma. Agronomy 2019, 9, 204. [Google Scholar] [CrossRef] [Green Version]
- Sales-Baptista, E.; d’Abreu, M.C.; Oliveira, M.I. Overgrazing in the Montado? The need for monitoring grazing pressure at paddock scale. Agrofor. Syst. 2016, 90, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Efe Serrano, J. Pastures in Alentejo: Technical Basis for Characterization, Grazing and Improvement; Universidade de Évora—ICAM, Ed.; Gráfica Eborense: Évora, Portugal, 2006; pp. 165–178. [Google Scholar]
- Schellberg, J.; Hill, M.J.; Gerhards, R.; Rothmund, M.; Braun, M. Precision agriculture on grassland: Applications, perspectives and constraints. Eur. J. Agron. 2008, 29, 59–71. [Google Scholar] [CrossRef]
- McCormick, S.; Jordan, C.; Bailey, J. Within and between-field spatial variation in soil phosphorus in permanent grassland. Precis. Agric. 2009, 10, 262–276. [Google Scholar] [CrossRef]
- Sales-Baptista, E.; Oliveira, I.F.; Santos, M.B.; Castro, J.A.; Pereira, A.; Rafael, J.; Serrano, J. Tecnologia GNSS de baixo custo na monitorizacão de ovinos em pastoreio. Revista de Ciências Agrárias 2016, 39, 251–260. (In Portuguese) [Google Scholar] [CrossRef]
- Serrano, J.; Sales-Baptista, E.; Shahidian, S.; Marques da Silva, J.; Ferraz de Oliveira, I.; Lopes de Castro, J.; Pereira, A.; Cancela d’Abreu, M.; Carvalho, M. Proximal sensors for monitoring seasonal changes of feeding sites selected by grazing ewes. Agroforest. Syst. 2018. [Google Scholar] [CrossRef]
- Marcos, G.M.; Obrador, J.J.; Garcia, E.; Cubera, E.; Montero, M.J.; Pulido, F.; Dupraz, C. Driving competitive and facilitative interactions in oak dehesas through management practices. Agrofor. Syst. 2007, 70, 25–40. [Google Scholar] [CrossRef]
- Benavides, R.; Douglas, G.B.; Osoro, K. Silvopastoralism in New Zealand: Review of effects of evergreen and deciduous trees on pasture dynamics. Agrofor. Syst. 2009, 76, 327–350. [Google Scholar] [CrossRef]
- Seddaiu, G.; Porcua, G.; Luigi, L.; Roggero, P.P.; Agnelli, A.; Cortic, G. Soil organic matter content and composition as influenced by soil management in a semi-arid Mediterranean agro-silvo-pastoral system. Agric. Ecosys. Environ. 2013, 167, 1–11. [Google Scholar] [CrossRef]
- Jackson, T.J.; Chen, D.; Cosh, M.; Li, F.; Anderson, M.; Walthall, C.; Doriaswamy, P.; Hunt, E.R. Vegetation water content mapping using Landsat data derived normalized difference water index for corn and soybeans. Remote Sens. Environ. 2004, 92, 475–482. [Google Scholar] [CrossRef]
- Guevara-Escobar, A.; Kemp, P.D.; Mackay, A.D.; Hodgson, J. Pasture production and composition under poplar in a hill environment in New Zealand. Agrofor. Syst. 2007, 3, 199–213. [Google Scholar] [CrossRef]
- Diacono, M.; Persiani, A.; Fiore, A.; Montemurro, F.; Canali, S. Agro-ecology for potential adaptation of horticultural systems to climate change: Agronomic and energetic performance evaluation. Agronomy 2017, 7, 35. [Google Scholar] [CrossRef] [Green Version]
- Serrano, J.; Shahidian, S.; Marques da Silva, J. Monitoring seasonal pasture quality degradation in the Mediterranean montado ecosystem: Proximal versus remote sensing. Water 2018, 10, 1422. [Google Scholar] [CrossRef] [Green Version]
- Serrano, J.; Shahidian, S.; Marques da Silva, J.; Sales-Baptista, E.; Ferraz de Oliveira, I.; Lopes de Castro, J.; Pereira, A.; Cancela d’Abreu, M.; Machado, E.; Carvalho, M. Tree influence on soil and pasture: Contribution of proximal sensing to pasture productivity and quality estimation in montado ecosystems. Int. J. Remote Sens. 2018, 39, 4801–4829. [Google Scholar] [CrossRef]
- Nawar, S.; Corstanje, R.; Halcro, G.; Mulla, D.; Mouazen, A.M. Delineation of soil management zones for variable-rate fertilization: A review. Adv. Agron. 2017, 143, 175–245. [Google Scholar]
- Serrano, J.; Shahidian, S.; Marques da Silva, J.; Paixão, L.; Calado, J.; Carvalho, M. Integration of soil electrical conductivity and indices obtained through satellite imagery for differential management of pasture fertilization. AgriEngineering 2019, 1, 567–585. [Google Scholar] [CrossRef] [Green Version]
- Corwin, D.L.; Lesch, S.M. Application of soil electrical conductivity to precision agriculture: Theory, principles, and guidelines. Agron. J. 2003, 95, 455–471. [Google Scholar] [CrossRef]
- Sudduth, K.A.; Kitchen, N.R.; Wiebold, W.J.; Batchelor, W.D.; Bollero, G.A.; Bullock, D.G. Relating apparent electrical conductivity to soil properties across the north-central USA. Comput. Electron. Agric. 2005, 46, 263–283. [Google Scholar] [CrossRef]
- Peralta, N.R.; Costa, J.L.; Balzarini, M.; Franco, M.C.; Córdoba, M.; Bullock, D. Delineation of management zones to improve nitrogen management of wheat. Comput. Electron. Agric. 2015, 110, 103–113. [Google Scholar] [CrossRef]
- Serrano, J.; Shahidian, S.; Marques da Silva, J. Calibration of GrassMaster II to estimate green and dry matter yield in Mediterranean pastures: Effect of pasture moisture content. Crop. Pasture Sci. 2016, 67, 780–791. [Google Scholar] [CrossRef]
- Louhaichi, M.; Hassan, S.; Clifton, K.; Johnson, D.E. A reliable and non-destructive method for estimating forage shrub cover and biomass in arid environments using digital vegetation charting technique. Agrofor. Syst. 2018, 92, 1341–1352. [Google Scholar] [CrossRef] [Green Version]
- Serrano, J.; Shahidian, S.; Marques da Silva, J. Evaluation of normalized difference water index as a tool for monitoring pasture seasonal and inter-annual variability in a Mediterranean agro-silvo-pastoral system. Water 2019, 11, 62. [Google Scholar] [CrossRef] [Green Version]
- AEMET. Iberian Climate Atlas; Agencia Estatal de Meteorología (España) and Instituto de Meteorología (Portugal): Madrid, Spain, 2011. [Google Scholar]
- FAO. World Reference Base for Soil Resources; Food and Agriculture Organization of the United Nations, World Soil Resources Reports N 103; FAO: Rome, Italy, 2006. [Google Scholar]
- Egner, H.; Riehm, H.; Domingo, W.R. Utersuchungeniiber die chemische Bodenanalyse als Grudlagefir die Beurteilung des Nahrstof-zunstandes der Boden. II. K. Lantbrhogsk. Annlr 1960, 20, 199–216. (In German) [Google Scholar]
- AOAC. Official Method of Analysis of AOAC International, 18th ed.; AOAC International: Arlington, AT, USA, 2005. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie, 3rd ed.; Grundzüge der Vegetationskunde; Springer: Vienna, Austria; New York, NY, USA, 1964. [Google Scholar]
- Lozano-García, B.; Parras-Alcántara, L. Land use and management effects on carbon and nitrogen in Mediterranean Cambisols. Agric. Ecosyst. Environ. 2013, 179, 208–214. [Google Scholar] [CrossRef]
- Lal, R. Restoring Soil Quality to Mitigate Soil Degradation. Sustainability 2015, 7, 5875–5895. [Google Scholar] [CrossRef] [Green Version]
- Yadav, R.P.; Sharma, P.; Arya, S.L.; Panwar, P. Acacia nilotica-based silvipastoral systems for resource conservation and improved productivity from degraded lands of the Lower Himalayas. Agrofor. Syst. 2014, 88, 851–853. [Google Scholar] [CrossRef]
- Lorenz, K.; Lal, R. Soil organic carbon sequestration in agroforestry systems. A review. Agron. Sustain. Dev. 2014, 34, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Bernardi, A.C.C.; Bettiol, G.M.; Ferreira, R.P.; Santos, K.E.L.; Rabello, L.M.; Inamasu, R.Y. Spatial variability of soil properties and yield of a grazed alfalfa pasture in Brazil. Precis. Agric. 2016, 17, 737–752. [Google Scholar] [CrossRef] [Green Version]
- Somarriba, E. Pasture growth and floristic composition under the shade of Guava (Psidium guajava L.) trees in Costa Rica. Agrofor. Syst. 1988, 6, 153–162. [Google Scholar] [CrossRef]
- Gómez-Rey, M.X.; Garcês, A.; Madeira, M. Soil organic-C accumulation and N availability under improved pastures established in Mediterranean Oak Woodlands. Soil Use Manag. 2012, 28, 497–507. [Google Scholar] [CrossRef]
- Ferraz de Oliveira, M.I.; Lamy, E.; Bugalho, M.N.; Vaz, M.; Pinheiro, C.; Cancela d’Abreu, M.; Capela e Silva, F.; Sales-Baptista, E. Assessing foraging strategies of herbivores in Mediterranean oak woodlands: A review of key issues and selected methodologies. Agrofor. Syst. 2013, 87, 1421–1437. [Google Scholar] [CrossRef]
- Hussain, Z.; Kemp, P.D.; Horne, D.J. Pasture production under densely planted young willow and poplar in a silvopastoral system. Agrofor. Syst. 2009, 76, 351–362. [Google Scholar] [CrossRef]
- Devkota, N.R.; Kemp, P.D.; Hodgson, J.; Valentine, I. Relationship between tree canopy height and the production of pasture species in a silvopastoral system based on alder trees. Agrofor. Syst. 2009, 76, 363–374. [Google Scholar] [CrossRef]
- Pérez-Ramos, I.M.; Zavala, M.A.; Maranon, T.; Díaz-Villa, M.D.; Valladares, F. Dynamics of understory herbaceous plant diversity following shrub clearing of cork oak forests: A five-year study. For. Ecol. Manag. 2008, 255, 3242–3253. [Google Scholar] [CrossRef]
- Castrignanò, A.; Buttafuoco, G.; Quarto, R.; Vitti, C.; Langella, G.; Terribile, F.; Venezia, A. A combined approach of sensor data fusion and multivariate geostatistics for delineation of homogeneous zones in an agricultural field. Sensors 2017, 17, 2794. [Google Scholar] [CrossRef]
- Moral, F.; Terrón, J.; Da Silva, J.M. Delineation of management zones using mobile measurements of soil apparent electrical conductivity and multivariate geostatistical techniques. Soil Tillage Res. 2010, 106, 335–343. [Google Scholar] [CrossRef]
- Serrano, J.; Shahidian, S.; Marques da Silva, J. Monitoring pasture variability: Optical OptRx® crop sensor versus Grassmaster II capacitance probe. Environ. Monit. Assess. 2016, 188, 1–17. [Google Scholar] [CrossRef]
- Padilla, F.M.; Souza, R.; Peña-Fleitas, M.T.; Grasso, R.; Gallardo, M.; Thompson, R.B. Infuence of time of day on measurement with chlorophyll meters and canopy refectance sensors of diferent crop N status. Precis. Agric. 2019, 20, 1087–1106. [Google Scholar] [CrossRef] [Green Version]
- Gebremedhin, A.; Badenhorst, P.E.; Wang, J.; Spangenberg, G.C.; Smith, K.F. Prospects for measurement of dry matter yield in forage breeding programs using sensor technologies. Agronomy 2019, 9, 65. [Google Scholar] [CrossRef] [Green Version]
- Chai, X.; Zhang, T.; Shao, Y.; Gong, H.; Liu, L.; Xie, K. Modeling and mapping soil moisture of plateau pasture using RADARSAT-2 imagery. Remote Sens. 2015, 7, 1279–1299. [Google Scholar] [CrossRef] [Green Version]
- Albayrak, S. Use of reflectance measurements for the detection of N, P, K, ADF and NDF contents in Sainfoin pasture. Sensors 2008, 8, 7275–7286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pullanagari, R.; Yule, I.; Tuohy, M.; Hedley, M.; Dynes, R.; King, W. Proximal sensing of the seasonal variability of pasture nutritive value using multispectral radiometry. Grass Forage Sci. 2013, 68, 110–119. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date and | Global | UTC | OTC | Significance | ||
|---|---|---|---|---|---|---|
| Soil Parameters | Mean ± SD | CV (%) | Range | Mean ± SD | Mean ± SD | (Probability) |
| October 2015 | ||||||
| Coarse sand (%) | 48.4 ± 2.6 | 5.3 | 44.0–54.8 | 49.0 ± 2.4 | 47.8 ± 2.6 | ns |
| Fine sand (%) | 32.2 ± 2.0 | 6.3 | 29.4–36.7 | 31.8 ± 1.5 | 32.6 ± 2.4 | ns |
| Silt (%) | 10.1 ± 1.7 | 16.4 | 6.7–13.0 | 9.8 ± 1.5 | 10.3 ± 1.8 | ns |
| Clay (%) | 9.3 ± 1.4 | 14.9 | 6.7–11.9 | 9.4 ± 1.0 | 9.3 ± 1.7 | ns |
| OM (%) | 2.0 ± 0.8 | 40.2 | 1.0–3.5 | 2.7 ± 0.5 | 1.3 ± 0.2 | 0.0000 |
| pH | 5.4 ± 0.3 | 6.0 | 4.9–6.2 | 5.4 ± 0.4 | 5.3 ± 0.2 | ns |
| P2O5 (mg kg−1) | 92.9 ± 62.4 | 67.1 | 26.0–343.0 | 117.7 ± 77.0 | 68.2 ± 29.5 | 0.0571 |
| K2O (mg kg−1) | 269.9 ± 135.9 | 50.3 | 94.0–540.0 | 359.3 ± 112.8 | 180.5 ± 91.9 | 0.0012 |
| Mg (mg kg−1) | 95.6 ± 43.7 | 45.7 | 20.0–180.0 | 115.0 ± 38.8 | 76.3 ± 40.9 | 0.0503 |
| Mn (mg kg−1) | 76.4 ± 44.9 | 58.7 | 14.0–175.0 | 100.0 ± 45.7 | 52.8 ± 30.1 | 0.0131 |
| CEC (cmol kg−1) | 7.3 ± 1.2 | 16.3 | 4.9–9.4 | 8.0 ± 0.8 | 6.6 ± 1.1 | 0.0058 |
| March 2018 | ||||||
| OM (%) | 2.4 ± 0.8 | 33.3 | 1.1–4.0 | 3.1 ± 0.5 | 1.7 ± 0.4 | 0.0000 |
| pH | 5.6 ± 0.3 | 4.5 | 5.2–6.1 | 5.6 ± 0.3 | 5.6 ± 0.2 | ns |
| P2O5 (mg kg−1) | 70.3 ± 27.2 | 38.8 | 17.0–132.0 | 88.6 ± 20.7 | 51.9 ± 19.7 | 0.0002 |
| K2O (mg kg−1) | 165.8 ± 66.0 | 39.8 | 78.0–304.0 | 210.3 ± 63.1 | 121.3 ± 28.5 | 0.0002 |
| Mg (mg kg−1) | 92.5 ± 24.3 | 26.2 | 55.0–150.0 | 99.6 ± 27.0 | 85.4 ± 19.8 | 0.0441 |
| Mn (mg kg−1) | 32.9 ± 14.2 | 43.2 | 11.6–64.8 | 39.3 ± 15.9 | 26.5 ± 8.9 | 0.0380 |
| Pasture Parameter | Global | UTC | OTC | Significance | ||
|---|---|---|---|---|---|---|
| Biomass (kg ha−1) | Mean ± SD | CV (%) | Range | Mean ± SD | Mean ± SD | (Probability) |
| March 2016 | 11426 ± 6328 | 55.4 | 1920–29700 | 8747 ± 4210 | 14106 ± 7095 | 0.0307 |
| April 2016 | 16903 ± 8264 | 48.9 | 6170–37530 | 12403 ± 3910 | 21403 ± 9128 | 0.0002 |
| May 2016 | 24148 ± 13584 | 56.3 | 7010–52290 | 15148 ± 5856 | 33149 ± 13221 | 0.0000 |
| June 2016 | 11003 ± 6483 | 58.9 | 1440–25920 | 6017 ± 3122 | 15990 ± 4888 | 0.0000 |
| February 2017 | 6588 ± 3161 | 48.0 | 800–11900 | 6875 ± 2500 | 6300 ± 3802 | ns |
| March 2017 | 11050 ± 5375 | 48.6 | 3200–25000 | 10808 ± 4876 | 11292 ± 6041 | ns |
| April 2017 | 12771 ± 7106 | 55.6 | 5100–36700 | 10975 ± 5227 | 14567 ± 8440 | ns |
| May 2017 | 5242 ± 2151 | 41.0 | 1600–9800 | 4717 ± 2080 | 5767 ± 2178 | 0.0765 |
| June 2017 | 1604 ± 744 | 46.4 | 400–3900 | 1483 ± 737 | 1725 ± 763 | ns |
| February 2018 | 4487 ± 2953 | 65.8 | 0–11680 | 5000 ± 2733 | 3973 ± 3193 | ns |
| March 2018 | 7427 ± 4519 | 60.8 | 1440–18400 | 7267 ± 3601 | 7587 ± 5448 | ns |
| April 2018 | 17520 ± 9215 | 52.6 | 5760–46080 | 18160 ± 7067 | 16880 ± 11256 | ns |
| May 2018 | 11996 ± 3983 | 33.2 | 4500–21000 | 12158 ± 3504 | 11833 ± 4566 | ns |
| March 2016 | 13763 ± 5779 | 42.0 | 6800–26200 | 9875 ± 1794 | 17650 ± 5800 | 0.0011 |
| Pasture Parameter | Global | UTC | OTC | Significance | ||
|---|---|---|---|---|---|---|
| CP (%) | Mean ± SD | CV (%) | Range | Mean ± SD | Mean ± SD | (Probability) |
| March 2016 | 13.8 ± 2.8 | 20.6 | 8.7–19.8 | 14.8 ± 3.5 | 12.8 ± 1.6 | 0.0300 |
| April 2016 | 11.9 ± 3.5 | 29.2 | 6.5–22.2 | 13.4 ± 4.2 | 10.5 ± 1.8 | 0.0375 |
| May 2016 | 10.3 ± 3.0 | 29.5 | 5.1–17.5 | 12.0 ± 2.9 | 8.6 ± 2.2 | 0.0009 |
| June 2016 | 7.6 ± 2.5 | 32.6 | 4.1–14.5 | 8.5 ± 3.1 | 6.7 ± 1.3 | ns |
| February 2017 | 16.9 ± 4.9 | 28.7 | 8.6–28.1 | 19.7 ± 4.5 | 14.2 ± 3.5 | 0.0083 |
| March 2017 | 15.4 ± 3.3 | 21.7 | 10.2–21.3 | 16.0 ± 3.5 | 14.8 ± 3.2 | ns |
| April 2017 | 9.2 ± 2.6 | 28.2 | 5.6–16.1 | 11.0 ± 2.2 | 7.4 ± 1.3 | 0.0005 |
| May 2017 | 9.1 ± 2.0 | 22.3 | 5.5–13.2 | 10.6 ± 1.6 | 7.7 ± 1.2 | 0.0006 |
| June 2017 | 6.7 ± 1.7 | 24.9 | 4.4–9.1 | 8.0 ± 1.1 | 5.4 ± 0.8 | 0.0000 |
| February 2018 | 18.7 ± 4.9 | 26.1 | 8.7–25.3 | 21.3 ± 3.8 | 15.9 ± 4.5 | 0.0025 |
| March 2018 | 18.3 ± 4.6 | 25.2 | 8.3–27.0 | 21.7 ± 3.4 | 15.0 ± 3.0 | 0.0001 |
| April 2018 | 13.2 ± 3.7 | 28.3 | 8.3–25.5 | 15.5 ± 3.6 | 10.8 ± 2.0 | 0.0008 |
| May 2018 | 15.2 ± 3.2 | 21.0 | 10.2–24.1 | 16.4 ± 2.8 | 14.0 ± 3.2 | 0.0621 |
| June 2018 | 13.8 ± 2.8 | 20.6 | 8.7–19.8 | 14.8 ± 3.5 | 12.8 ± 1.6 | 0.0300 |
| NDF (%) | ||||||
| March 2016 | 38.1 ± 5.3 | 13.8 | 29.5–50.2 | 40.0 ± 5.8 | 36.2 ± 4.1 | 0.0987 |
| April 2016 | 52.9 ± 5.9 | 11.1 | 43.1–64.3 | 55.4 ± 5.3 | 50.5 ± 5.5 | 0.0084 |
| May 2016 | 64.0 ± 4.4 | 6.8 | 53.8–74.1 | 63.6 ± 5.6 | 64.5 ± 2.9 | ns |
| June 2016 | 66.8 ± 6.1 | 9.1 | 55.3–78.3 | 68.5 ± 6.8 | 65.1 ± 5.0 | ns |
| February 2017 | 49.9 ± 10.8 | 21.7 | 35.0–77.5 | 48.5 ± 8.6 | 51.2 ± 12.9 | ns |
| March 2017 | 50.5 ± 6.4 | 12.6 | 36.9–64.6 | 51.5 ± 4.7 | 49.6 ± 7.8 | ns |
| April 2017 | 60.6 ± 4.2 | 7.0 | 53.1–70.2 | 60.8 ± 3.4 | 60.4 ± 5.1 | 0.0729 |
| May 2017 | 70.7 ± 3.5 | 5.0 | 65.6–79.6 | 68.8 ± 1.9 | 72.7 ± 3.7 | 0.0007 |
| June 2017 | 76.0 ± 3.7 | 4.8 | 68.4–83.9 | 74.0 ± 3.5 | 78.1 ± 2.6 | 0.0057 |
| February 2018 | 34.3 ± 11.9 | 34.6 | 18.6–58.9 | 35.0 ± 8.3 | 33.5 ± 15.3 | ns |
| March 2018 | 36.0 ± 10.4 | 28.7 | 17.4–52.6 | 38.3 ± 9.8 | 33.8 ± 10.8 | ns |
| April 2018 | 40.3 ± 7.1 | 17.7 | 31.3–52.6 | 42.9 ± 7.1 | 37.7 ± 6.5 | 0.0227 |
| May 2018 | 46.8 ± 7.1 | 15.2 | 33.0–60.3 | 49.0 ± 6.9 | 44.7 ± 7.0 | 0.0325 |
| June 2018 | 60.2 ± 3.4 | 5.7 | 51.7–66.4 | 61.1 ± 2.0 | 59.2 ± 4.3 | ns |
| Pasture Parameter | Global | UTC | OTC | Significance | ||
|---|---|---|---|---|---|---|
| PMC (%) | Mean ± SD | CV (%) | Range | Mean ± SD | Mean ± SD | (Probability) |
| March 2016 | 85.2 ± 4.2 | 4.9 | 74.6–91.3 | 84.7 ± 5.4 | 85.8 ± 2.6 | ns |
| April 2016 | 85.5 ± 2.4 | 2.8 | 78.8–89.7 | 85.0 ± 3.1 | 85.9 ± 1.3 | ns |
| May 2016 | 84.6 ± 5.2 | 6.1 | 76.1–93.2 | 81.1 ± 3.8 | 88.0 ± 4.0 | 0.0003 |
| June 2016 | 59.8 ± 6.4 | 10.7 | 45.5–72.4 | 59.1 ± 6.1 | 60.5 ± 6.9 | ns |
| February 2017 | 74.8 ± 11.0 | 14.7 | 42.1–88.1 | 78.7 ± 6.1 | 71.0 ± 13.6 | ns |
| March 2017 | 79.4 ± 5.9 | 7.5 | 62.5–86.9 | 79.0 ± 7.0 | 79.7 ± 5.0 | ns |
| April 2017 | 64.3 ± 6.2 | 9.7 | 54.5–77.9 | 62.5 ± 6.3 | 66.2 ± 5.8 | ns |
| May 2017 | 45.8 ± 9.5 | 20.8 | 22.5–60.2 | 43.8 ± 10.5 | 47.9 ± 8.4 | ns |
| June 2017 | 14.5 ± 4.7 | 32.2 | 6.3–26.3 | 14.3 ± 4.7 | 14.7 ± 4.8 | ns |
| February 2018 | 77.5 ± 7.8 | 10.1 | 55.6–86.1 | 78.6 ± 6.9 | 76.2 ± 8.9 | ns |
| March 2018 | 82.4 ± 5.2 | 6.3 | 66.7–88.9 | 83.9 ± 3.1 | 80.9 ± 6.5 | ns |
| April 2018 | 84.6 ± 2.4 | 2.9 | 79.6–88.6 | 85.6 ± 2.2 | 83.6 ± 2.3 | 0.0234 |
| May 2018 | 82.7 ± 2.8 | 3.3 | 73.3–87.1 | 83.6 ± 2.0 | 81.7 ± 3.1 | 0.0583 |
| June 2018 | 68.5 ± 5.7 | 8.3 | 54.2–77.8 | 66.5 ± 6.9 | 70.6 ± 3.1 | 0.0996 |
| PQDI | ||||||
| March 2016 | 2.9 ± 0.6 | 21.4 | 1.5–4.6 | 2.9 ± 0.8 | 2.8 ± 0.4 | ns |
| April 2016 | 4.8 ± 1.5 | 31.7 | 2.3–8.8 | 4.5 ± 1.5 | 5.1 ± 1.5 | ns |
| May 2016 | 6.9 ± 2.5 | 36.8 | 3.1–12.6 | 5.6 ± 1.6 | 8.1 ± 2.7 | 0.0071 |
| June 2016 | 9.6 ± 3.0 | 31.0 | 4.2–16.8 | 9.0 ± 3.3 | 10.1 ± 2.6 | ns |
| February 2017 | 3.3 ± 1.7 | 52.2 | 1.4–9.0 | 2.6 ± 0.9 | 4.0 ± 2.1 | 0.0355 |
| March 2017 | 3.5 ± 1.0 | 29.1 | 2.3–6.4 | 3.4 ± 0.8 | 3.5 ± 1.2 | ns |
| April 2017 | 7.1 ± 2.1 | 29.4 | 3.9–12.5 | 5.7 ± 1.1 | 8.5 ± 1.9 | 0.0050 |
| May 2017 | 8.2 ± 2.2 | 27.2 | 5.2–14.4 | 6.7 ± 1.2 | 9.7 ± 2.0 | 0.0002 |
| June 2017 | 12.2 ± 3.6 | 29.2 | 7.5–18.8 | 9.5 ± 2.0 | 14.9 ± 2.5 | 0.0001 |
| February 2018 | 2.1 ± 1.4 | 66.4 | 0.9–6.3 | 1.7 ± 0.6 | 2.5 ± 1.9 | 0.0109 |
| March 2018 | 2.1 ± 1.1 | 51.8 | 1.0–6.1 | 1.8 ± 0.4 | 2.5 ± 1.4 | 0.0518 |
| April 2018 | 3.2 ± 0.8 | 25.9 | 1.9–5.5 | 2.8 ± 0.6 | 3.6 ± 0.9 | 0.0137 |
| May 2018 | 3.2 ± 0.9 | 27.7 | 1.5–5.9 | 3.0 ± 0.5 | 3.4 ± 1.1 | ns |
| June 2018 | 6.0 ± 1.4 | 22.9 | 3.8–8.2 | 5.4 ± 1.2 | 6.7 ± 1.3 | 0.082 |
| Botanical Species | Spring 2016 | Spring 2018 | ||
|---|---|---|---|---|
| (Mean Cover, %) | UTC | OTC | UTC | OTC |
| Anagalis arvensis | 1.0 | 0.6 | 0.0 | 0.0 |
| Arum italicum | 0.0 | 0.0 | 1.1 | 0.0 |
| Astragalus pelecinus | 0.0 | 0.3 | 0.0 | 0.0 |
| Bromus diandrus | 1.7 | 0.4 | 42.8 | 19.4 |
| Calendula arvensis | 0.1 | 0.1 | 0.0 | 0.0 |
| Cerastium glomeratum | 0.1 | 1.8 | 4.6 | 0.0 |
| Chamaemelum fuscatum | 0.0 | 0.0 | 0.4 | 8.1 |
| Chamaemelum mixtum | 6.9 | 17.0 | 0.0 | 0.0 |
| Crepis capillaris | 0.0 | 0.0 | 0.0 | 2.4 |
| Diplotaxis catholica | 0.6 | 6.3 | 0.5 | 15.5 |
| Echium sp. | 0.0 | 0.1 | 0.0 | 0.0 |
| Erodium malacoides | 0.0 | 0.0 | 5.4 | 0.0 |
| Erodium moschatum | 40.2 | 15.6 | 36.2 | 37.9 |
| Geranium molle | 1.0 | 1.4 | 0.0 | 0.0 |
| Gramínea sp. | 12.9 | 1.6 | 0.0 | 0.0 |
| Gynandriris sisyrinchium | 1.4 | 0.0 | 0.0 | 0.2 |
| Hordeum murinum | 0.0 | 0.0 | 5.1 | 0.0 |
| Leontodon taraxacoides | 6.2 | 12.2 | 0.0 | 2.4 |
| Ornithopus sp. | 0.0 | 0.3 | 0.0 | 0.0 |
| Plantago coronopus | 1.8 | 8.4 | 0.0 | 0.0 |
| Plantago lagopus | 0.0 | 0.0 | 0.0 | 1.4 |
| Plantago lanceolata | 0.4 | 1.7 | 0.0 | 0.0 |
| Poa annua | 1.2 | 1.1 | 0.0 | 0.0 |
| Raphanus raphanistrum | 0.0 | 1.0 | 0.0 | 0.6 |
| Rumex bucephalophorus | 0.2 | 5.9 | 0.0 | 2.5 |
| Rumex conglomeratus | 0.6 | 0.0 | 0.0 | 0.0 |
| Senecio vulgaris | 0.0 | 0.4 | 1.7 | 4.5 |
| Sherardia arvensis | 0.1 | 0.1 | 0.0 | 0.5 |
| Silene sp. | 0.1 | 0.0 | 0.0 | 0.0 |
| Sonchus oleraceus | 2.7 | 2.1 | 0.0 | 0.0 |
| Spergula arvensis | 0.5 | 1.7 | 0.0 | 2.7 |
| Stachys arvensis | 0.0 | 0.3 | 0.0 | 0.0 |
| Stellaria media | 3.5 | 1.0 | 0.0 | 0.8 |
| Trifolium glomeratum | 2.0 | 0.1 | 0.0 | 0.0 |
| Trifolium incarnatum | 0.0 | 1.0 | 0.0 | 0.0 |
| Trifolium repens | 1.4 | 4.2 | 0.0 | 0.0 |
| Trifolium resupinatum | 0.5 | 9.6 | 0.0 | 0.0 |
| Tripholium subterraneum | 0.0 | 0.0 | 0.0 | 1.1 |
| Urtica sp. | 1.9 | 0.1 | 0.0 | 0.0 |
| Urtica urens | 0.0 | 0.0 | 2.2 | 0.0 |
| Vulpia sp. | 11.1 | 3.3 | 0.0 | 0.0 |
| Total | 100.0 | 100.0 | 100.0 | 100.0 |
| Pasture Parameter | Global | UTC | OTC | Significance | ||
|---|---|---|---|---|---|---|
| NDVI | Mean ± SD | CV (%) | Range | Mean ± SD | Mean ± SD | (Probability) |
| March 2016 | 0.780 ± 0.063 | 8.1 | 0.625–0.849 | 0.764 ± 0.079 | 0.796 ± 0.039 | 0.0790 |
| April 2016 | 0.732 ± 0.065 | 8.9 | 0.621–0.843 | 0.741 ± 0.078 | 0.723 ± 0.050 | ns |
| May 2016 | 0.644 ± 0.063 | 9.8 | 0.524–0.757 | 0.667 ± 0.051 | 0.622 ± 0.069 | ns |
| June 2016 | 0.345 ± 0.073 | 21.1 | 0.204–0.506 | 0.361 ± 0.070 | 0.329 ± 0.075 | ns |
| February 2017 | 0.779 ± 0.092 | 11.8 | 0.558–0.885 | 0.825 ± 0.046 | 0.733 ± 0.104 | 0.0090 |
| March 2017 | 0.807 ± 0.061 | 7.6 | 0.642–0.880 | 0.803 ± 0.077 | 0.811 ± 0.043 | ns |
| April 2017 | 0.583 ± 0.100 | 17.1 | 0.397–0.844 | 0.609 ± 0.126 | 0.556 ± 0.057 | 0.0433 |
| May 2017 | 0.342 ± 0.078 | 22.8 | 0.248–0.505 | 0.369 ± 0.078 | 0.315 ± 0.071 | 0.0181 |
| June 2017 | 0.197 ± 0.025 | 12.5 | 0.160–0.242 | 0.216 ± 0.018 | 0.177 ± 0.010 | 0.0000 |
| February 2018 | 0.802 ± 0.084 | 10.5 | 0.493–0.898 | 0.829 ± 0.041 | 0.774 ± 0.107 | 0.0464 |
| March 2018 | 0.804 ± 0.087 | 10.8 | 0.474–0.877 | 0.835 ± 0.029 | 0.773 ± 0.113 | 0.0719 |
| April 2018 | 0.796 ± 0.056 | 7.0 | 0.605–0.864 | 0.822 ± 0.032 | 0.771 ± 0.064 | 0.0297 |
| May 2018 | 0.781 ± 0.044 | 5.7 | 0.678–0.824 | 0.805 ± 0.020 | 0.757 ± 0.049 | 0.0152 |
| June 2018 | 0.548 ± 0.100 | 18.3 | 0.422–0.772 | 0.579 ± 0.121 | 0.517 ± 0.065 | ns |
| CMR | ||||||
| March 2016 | 6992 ± 1879 | 26.9 | 3453–10731 | 5944 ± 1367 | 8039 ± 1765 | 0.0026 |
| April 2016 | 8564 ± 2101 | 24.5 | 4760–12153 | 6994 ± 1199 | 10134 ± 1553 | 0.0000 |
| May 2016 | 9497 ± 2160 | 28.8 | 3922–12994 | 5818 ± 1124 | 11176 ± 1529 | 0.0000 |
| June 2016 | 5735 ± 1953 | 34.1 | 3400–10861 | 4322 ± 568 | 7148 ± 1817 | 0.0000 |
| February 2017 | 4489 ± 631 | 14.1 | 3446–5618 | 4605 ± 633 | 4372 ± 634 | ns |
| March 2017 | 7082 ± 1761 | 24.9 | 3850–9888 | 6782 ± 1863 | 7381 ± 1679 | ns |
| April 2017 | 4941 ± 905 | 18.3 | 3829–7583 | 4919 ± 821 | 4965 ± 1035 | ns |
| May 2017 | 3804 ± 545 | 14.3 | 3125–5389 | 3528 ± 239 | 4056 ± 630 | ns |
| June 2017 | - | - | - | - | - | - |
| February 2018 | 4688 ± 929 | 19.8 | 3435–6564 | 5035 ± 970 | 4311 ± 750 | 0.0320 |
| March 2018 | 5535 ± 1077 | 19.5 | 3535–7093 | 5977 ± 802 | 5093 ± 1166 | 0.0162 |
| April 2018 | 6295 ± 1049 | 16.7 | 4604–8253 | 6605 ± 764 | 5985 ± 1227 | ns |
| May 2018 | 7356 ± 1085 | 14.7 | 5883–9321 | 7087 ± 1008 | 7626 ± 1134 | ns |
| June 2018 | 6498 ± 1249 | 19.2 | 4024–8531 | 5731 ± 1088 | 7266 ± 890 | 0.0015 |
| Soil and Pasture Characteristics (as Dependent Variables) | Equation | Parameters Measured by Proximal Sensors (as Independent Variables, x) | R2 |
|---|---|---|---|
| Clay (%) | Clay = 0.309x + 5.7418 | 0.5487 * | |
| CEC (cmol kg−1) | CEC = 0.2237x + 4.6912 | (i) ECa (mS m−1) | 0.3908 * |
| SMC (%) | SMC = 0.3018x + 6.0362 | 0.4802 * | |
| Biomass (kg ha−1) | Biomass = 5 × 10−5x2 + 2.3518x − 5205.7 | (ii) CMR | 0.7505 ** |
| PMC (%) | PMC = −3 × 10−6x2 + 0.0411x − 76.598 | 0.6049 * | |
| Biomass (kg ha−1) | Biomass = −114008x2 + 131904x − 22068 | 0.4703 * | |
| PMC (%) | PMC = −198.04x2 + 300.23x − 32.397 | 0.9134 ** | |
| CP (%) | CP = 34.748x2 − 21.137x + 10.586 | (iii) NDVI | 0.8186 ** |
| NDF (%) | NDF = −90.48x2 + 39.537x + 68.62 | 0.8421 ** | |
| PQDI | PQDI = −14.296x + 14.49 | 0.9282 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serrano, J.; Shahidian, S.; Marques da Silva, J.; Paixão, L.; Carreira, E.; Pereira, A.; Carvalho, M. Climate Changes Challenges to the Management of Mediterranean Montado Ecosystem: Perspectives for Use of Precision Agriculture Technologies. Agronomy 2020, 10, 218. https://doi.org/10.3390/agronomy10020218
Serrano J, Shahidian S, Marques da Silva J, Paixão L, Carreira E, Pereira A, Carvalho M. Climate Changes Challenges to the Management of Mediterranean Montado Ecosystem: Perspectives for Use of Precision Agriculture Technologies. Agronomy. 2020; 10(2):218. https://doi.org/10.3390/agronomy10020218
Chicago/Turabian StyleSerrano, João, Shakib Shahidian, José Marques da Silva, Luís Paixão, Emanuel Carreira, Alfredo Pereira, and Mário Carvalho. 2020. "Climate Changes Challenges to the Management of Mediterranean Montado Ecosystem: Perspectives for Use of Precision Agriculture Technologies" Agronomy 10, no. 2: 218. https://doi.org/10.3390/agronomy10020218