Abstract
We analyze a model in which individuals have hereditary reproductive types. The reproductive value of an individual is determined by her reproductive type and the amount of resources she can access. We introduce the possibility of suicide and assume it is also a genetic trait that interacts with the reproductive type of an individual. The main result of the paper is that populations where suicide is possible grow faster than other populations.
1. Introduction
The basic principle of evolution says that behaviour generated by a gene should maximise the gene’s frequency in the population. This principle, in general, implies that individuals try to survive and use as much reproductive resource as possible. For this reason, it is not easy to make sense of suicide or deliberate childlessness from an evolutionary perspective. As a solution, [] proposes a model in which a preference for self-destruction, contingent on the individual’s low reproductive potential compared with that of her kin, is hereditary. If a low fertility individual’s survival decreases the ability of her high fertility genetic kin to pass on their genes, then self-destruction for a net genetic gain should be evolutionarily adaptive. This logic is based on []’s “inclusive fitness” theory.
However, inclusive fitness theory is unlikely to explain the suicide of high-fertility individuals. Moreover, the notion that an individual accepts to die based on a comparison of her reproductive potential relative to that of her kin may well describe the behaviour of semelparous or colonial species1, but it seems less plausible in motivating human suicide. Ultimately, an individual might be able to stop competing for resources with her kin without resorting to suicide. We, therefore, propose a model in which suicidality is a genetic trait that can be expressed for some realisations of the genotype and remains silent for others. In our model, all individuals have the same probability of producing a suicidal offspring who will unconditionally self-destruct regardless of her fertility. We also assume that if an individual commits suicide, her reproductive resources are redistributed uniformly across the population, irrespective of genetic kinship.
Our theory is based on the comparison between the growth rates of two populations: one in which suicide is genetically possible and one in which is it not. Individual selection forces the suicide rate to be proportional to the genetic mutation rate. That is, suicide only occurs as the result of genetic mutation. Our main result is that the population where suicide is possible grows faster than the other one. Due to suicide, this population is able to more effectively eliminate mutations, slowing down the population growth rate and, therefore, to transform reproductive resources into offspring more efficiently. Consequently, our theory predicts that group selection favours those populations where suicide is technologically feasible. We emphasise that the genotype of an individual is primarily determined by individual selection, and the group selection effect is of a second order. Nonetheless, we believe that this second order effect provides a rationale for the suicidal gene to survive the evolutionary process.
The setup of our model is as follows. We first consider a baseline population model without the possibility of suicide. Individuals live for one period and have one of two possible reproductive types, either high or low. An individual’s reproductive type is genetic and imperfectly inherited from her parent. With a small probability, the gene mutates, and the type of an offspring is different from that of her parent. An individual’s reproductive value, which, in our model, is taken to be the number of her offspring, is the product of her reproductive type and the resources she uses to reproduce. In this baseline model, each individual is endowed with one unit of reproductive resources, so an individual’s reproductive value is just her type. In the steady state, the fraction of individuals with the low type is of the order of the probability of mutation.
We then introduce the possibility of suicide into our baseline model. We assume that the choice to commit suicide is also genetically determined. Therefore, a genotype is now a two-dimensional vector. The first coordinate determines the individual’s reproductive type, and the second one describes for which realisation of her reproductive type the individual would commit suicide. For example, an individual whose genotype is is of the high reproductive type and would commit suicide if she were not. Each coordinate of the genotype can independently mutate. Finally, we assume that the choice of suicide is made prior to reproducing and that the resources of those who committed suicide are uniformly distributed among the surviving individuals, irrespective of genetic kinship. Here, both the steady-state fraction of individuals with the low type and that of individuals who commit suicide are proportional to the mutation probability.
The main result of our paper is that the population where suicide is technologically feasible grows faster than the other. Therefore, if both populations were competing for limited resources, the population with the possibility of suicide would crowd out the other. In other words, group selection arguments predict the survival of that population in which suicide is genetically feasible.
The intuition for our result is as follows. In steady-state, whether or not suicide is feasible, most of the individuals have the high reproductive type. In the population with suicide, approximately half of these individuals carry the gene that prescribes self-destruction for the event in which the reproductive type mutates. As a result, half of those offspring who mutate to the low reproductive type commit suicide, and their resources are redistributed among the surviving population. Since that population consists of mostly high-type individuals who are more efficient at transforming resources into offspring, this effect induces a higher growth rate. Of course, suicide also generates the converse, negative effect: some high-type offspring issued from low-type parents also commit suicide. However, this negative effect is shown to have smaller magnitude than the positive effect, due to the low frequency of low-type individuals in the population.
This paper inscribes itself in a literature envisaging that, because suicide prevails at a fairly high rate across most cultures2, it cannot be fully explained as a sociological oddity. Instead, it needs to be made sense of as an essential trait of human behaviour. There is ample evidence suggesting that a predisposition to suicide can be partly inherited, although no one “suicide gene” has so far been identified3.
Psychiatric genetic theory, Darwinian psychiatry and evolutionary genetic theory have proposed three possible answers to the puzzle as to why evolution seems unable to eliminate genes that predispose individuals to suicide or harmful mental disorders, such as schizophrenia or bipolar disorder. The first (ancestral neutrality) purports that these genes were not harmful among our ancestors, the second (balancing selection) that these genes sometimes increased fitness and the third (polygenic mutation-selection balance) that extreme mental disorders reflect the inevitable mutational load on the thousands of genes underlying human behaviour. [] evaluate the merits of these theories as explanations for the persistence of debilitating mental disorders and find that the third is most consistent with data on mental disorder prevalence rates, fitness costs, the likely rarity of genes linked to them and the increased risks of mental disorders with brain trauma, inbreeding and paternal age. Our approach is comparatively naive from the medical point of view.
The negative impact of suicide on social welfare is indisputable4. Economists have attempted to identify socio-economic factors affecting suicide behaviour. [] consider a model in which an individual commits suicide once the discounted utility of her remaining lifetime, as a function of her remaining life expectancy and permanent income, falls below some individual-specific threshold. They find that in the US, suicide rates are generally lower amongst higher-income groups and that the suicide behaviour of older people is more sensitive to changes in permanent income than that of younger ones.
Such a utility-based approach to suicide has since been adopted and further developed, notably5 by [], who incorporate into their analysis the option value from delaying the act of suicide, and [], who study the implications of this for the risk-taking behaviour of individuals over their lifecycle. Our model falls outside this utility-based approach, as it investigates whether suicide as the result of genetic hazard and independent from socio-economic factors is evolutionarily sustainable.
Ref [] develops a signalling model in which a suicide attempt is used to elicit a sympathetic response from a receiver. More serious suicide attempts are more credible, but are more likely to kill the sender. Ref [] incorporates into the utility-based approach the possibility that a failed suicide attempt increases an individual’s utility ex-post.
We refer to literature reviews by [,,,,] for references to empirical work on the relationship between suicide and socio-economic factors, such as income, education, age, employment, marital status or the use of Internet chat rooms.
2. The Model
The baseline model—Time is discrete and denoted by In a population, there is a continuum of individuals. Each individual has one of two possible types, , . Each individual lives for one period, and the number of her offspring is proportional to her type, that is, an individual with type produces offspring, where ρ is a strictly positive constant to be defined later. The type is genetic and is imperfectly inherited by the offspring. To be more specific, the type of an individual is the same as that of her parent with probability . With the remaining probability, ε, a mutation occurs, and the type changes. We shall refer to this type as the reproductive type.
The possibility of suicide—Suppose now that each individual has a two-dimensional type, , where and . The first coordinate is the reproductive type described in the previous paragraph. The second coordinate describes the circumstances under which the individual commits suicide. To be more specific, we assume that if , the individual commits suicide prior to reproducing. Otherwise, the individual produces offspring. If the second coordinate is ∅, the individual does not commit suicide, irrespective of her reproductive type. The evolution of the second coordinate of the type across generations is similar to the first one. It mutates with probability ε, and conditional on this event, it changes to one of the other two types with equal probability. We assume that the probability of mutation is independent across the coordinates. We could have assumed that there are types who always commit suicide. However, such an assumption has no bearing on our results, but would somewhat complicate our analysis.
Reproductive Resources—Each individual receives a unit of endowment at birth. This endowment is the reproductive input and is used to produce offspring by those individuals who do not commit suicide. The endowment of those who commit suicide is uniformly redistributed among the entire surviving population. Hence, the number of offspring produced by an individual with reproductive type χ is , where ρ is the per capita amount of reproductive resources available after the suicide decision is made. Note that in the population where suicide is not possible. In the population where suicide is a possibility, ρ is the inverse of the survival probability, and the survival probability is one minus the probability of suicide.
3. Analysis
We introduce the following pieces of notation. Let and denote the measures of type-α and type-β individuals at time t in the population where there is no suicide. Moreover, let denote the fraction of type-χ in the population at t. Similarly, let denote the measure of individuals with type at t in the population with suicide before the decision regarding suicide is made, and let denote the measure of those individuals at t with reproductive type χ who do not commit suicide, i.e. those individuals that reproduce. In addition, let denote the fraction of individuals with type in the surviving population at t, that is, after the decision regarding suicide is made6, and let . Finally, let denote the per capita resources available at the end of period t, that is:
It can be shown that irrespective of the initial conditions, the populations (with and without suicide) converge to a steady state, where the population grows at a constant rate and the fractions of various types are constant. More precisely, for both
where g denotes the growth factor and is the steady-state fraction of individuals with reproductive type α in the population without suicide. Similarly, in the population with suicide, for all ,
where is the growth factor, is the steady state fraction of individuals with reproductive type α in the surviving population and ρ is the steady-state per capita amount of reproductive resources available in the population with suicide.
The populations with and without suicide are dominated by individuals with reproductive type α. To see this, consider the population without suicide, and note that the dynamics of are described by the following equation:
The reason is that each type-α individual reproduces α offspring, but a fraction, ε, of them mutate and become β types. Therefore, is the measure of type-α individuals at , whose parent is also of type α. Similarly, is the measure of type-α individuals at , whose parent is of type β. Dividing through by , we obtain:
As t goes to infinity, using (1), this equation becomes:
which is essentially the Euler-Lotka equation. Note that , because no individual can reproduce more than α offspring. Therefore, the previous equality implies , which means the fraction of type-β individuals is at most proportional to the mutation rate. A similar argument yields the analogue result for the population with suicide. In addition, it is equally easy to show that the fraction of individuals committing suicide is also proportional to ε.
Next, we show that the ratio of individuals with type to those with type is approximately one in the steady state. This implies that, conditional on her reproductive type mutating from α to β, an offspring commits suicide with a probability of approximately one half.
Lemma 1.
proof. The evolution of and can be described by the following equations:
Let us explain the first equality. Each individual with type produces offspring. The probability that the gene of the offspring does not mutate is . Therefore, the first term corresponds to those individuals of type , whose parents are also of type . An individual with type produces offspring; with probability , the first coordinate of the gene does not mutate, but the second coordinate mutates to β. Therefore, the second term corresponds to those individuals of type , whose parents are of type . There are also individuals with type , whose parents are either of type or of type ; however, they are the result of a double mutation, and these individuals are collected in the last term. The second equation can be explained in a similar manner.
Dividing through the first and second lines of (3) by and , respectively, yields:
Taking t to infinity, by (2), and noting that for large t, the previous equations can be written as:
Note that these equations can only be satisfied simultaneously if:
from which the statement of the lemma follows. ☐
The left panel of Figure 1 below depicts the steady-state measures of genotypes that do not commit suicide. Note that and converge to a common value as ε goes to zero, which is what the previous lemma states. This value is one half, because, as mutations become more rare, the more efficient reproductive type, α, takes over the entire population, and the ratio of individuals carrying reproductive type β goes to zero. The right panel describes ρ, the amount of reproductive resources available per capita in the steady state in the population with suicide. It is easy to show that the probability of suicide is , and hence, .
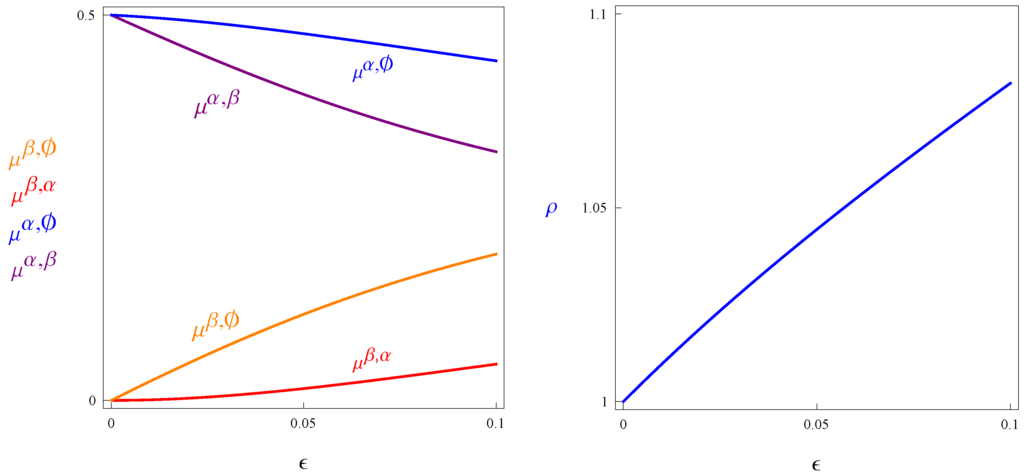
Figure 1.
The left panel describes the steady state fractions of types in the population with suicide. The right panel illustrates ρ, the steady state per capita reproductive resources in the population with suicide. All variables are described as functions of ϵ, letting and .
We now use the previous lemma to compare the fractions of individuals who have reproductive type α and β in both populations. We establish that if the probability of mutation is small enough, type α will be more prevalent in a population if suicide is possible than if it is not.
Proposition 1. There is an , such that if , then .
proof. Consider first the population without the possibility of suicide. The following equation describes the evolution of :
Recall that in the population without suicide, . There are type-β individuals at t, who each produce β offspring, and a fraction ε of these offspring mutate and become type-α. Therefore, is the measure of those type-β individuals at , whose parents are also of type β. Conversely, is the measure of those type-β individuals at , whose parents are of type-α. Divide the previous equation by to get:
Taking t to infinity, by (1), this equation can be written as:
so that, in the steady state:
Now, consider the population with suicide. The evolution of is described by:
The number of type-β offspring whose parent is also of reproductive type β is . Some of these offspring commit suicide, because their second coordinate mutates, which happens with a probability of . Therefore, the number of surviving offspring is . Conversely, the number of type-β offspring whose parent is of reproductive type α is . By Lemma 1, approximately half of them commit suicide, because the second coordinate of their type is β. Therefore, the number of surviving offspring issued from parents of reproductive type α is . Dividing (6) through by yields:
Taking t to infinity, by (2), the previous equation becomes:
Therefore:
If ε goes to zero, then ρ converges to one, and converges to g . Therefore, if ε is small, then the right-hand side of (7) is larger than the right-hand side of (5). The statement of the proposition follows from the observation that the function is increasing. ☐
The figure below shows that in the steady state, the fraction of the population of reproductive type α is larger in the population with suicide than in the population without suicide, illustrating Proposition 1.

Figure 2.
Steady state fractions of individuals of reproductive type α in the population without suicide () and with (), as a function of ϵ and letting and .
Theorem 1. There is an , such that if , then .
proof. The growth factor of the population without the possibility of suicide is:
The growth factor of the population with the possibility of suicide is:
Note that, in the population with suicide, only a fraction of of the population survives and reproduces, each of them uses ρ resources; so, ρ cancels out in the growth equation. By Proposition 1, there is an , such that if , then . Therefore, if , then . ☐
The theorem above established that the population in which suicide is possible grows faster than that population in which it is not, despite the negative impact that suicide, through sheer loss of individuals, has on population growth. This is due to two complementary effects: in the population with suicide, there are more reproductive resources per capita, and there is a higher fraction of α types. If the resources of those individuals who commit suicide were not redistributed among the surviving individuals, the population with suicide would grow slower than the population without. However, the population with suicide grows faster than the population without suicide would if individuals uniformly chosen across the population died, leaving the remaining population with ρ per capita resources.
Figure 3 illustrates the result of Theorem 1. We have seen that both populations are dominated by individuals of type-α. Therefore, if , the growth factors of both populations are α. However, when ε is positive, the share of type-β individuals rises in both populations, resulting in slower growth. In the population with suicide, this negative effect is partially offset by suicide. Indeed, half of those whose reproductive type mutates to β commit suicide (see Lemma 1). As a result, the fraction of individuals with reproductive type α is larger in the population with suicide (see Proposition 1), and the population with suicide grows faster.
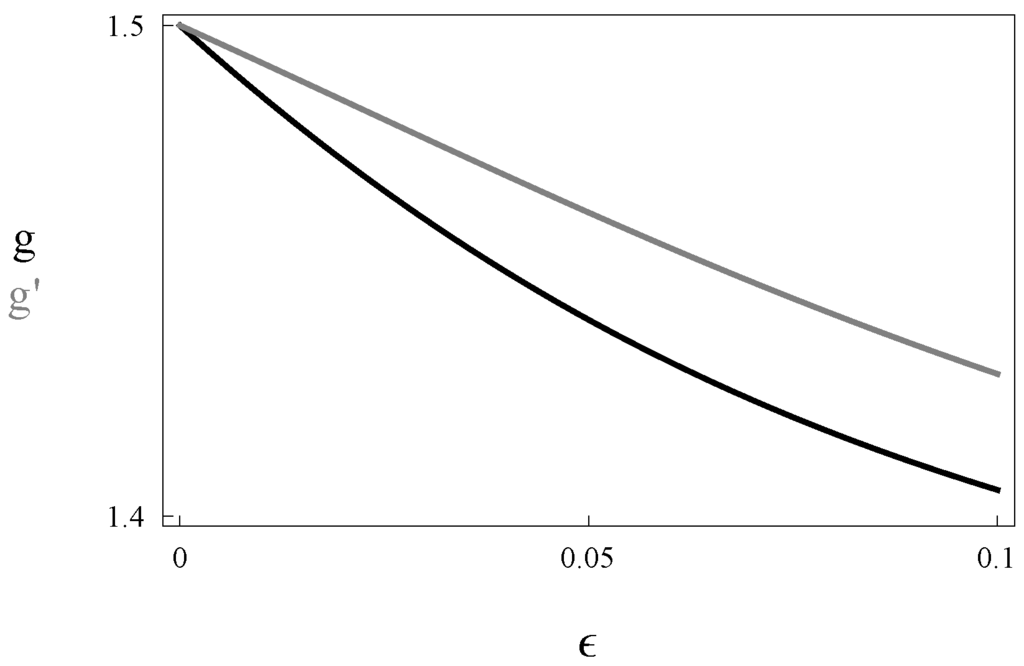
Figure 3.
Steady state growth factor of the population without suicide (g) and with (), as a function of ϵ and letting and .
4. Conclusion
In this paper, we raise the possibility that it may be evolutionarily adaptive for suicide behaviour to be genetically determined. We propose a model in which suicide occurs solely as the result of genetic mutation and is not motivated by an individual’s reproductive potential relative to that of her genetic kin or by socio-economic factors. Rather, offspring with certain genotypes commit suicide independently of their environment, and both high and low fertility individuals have the same ex-ante likelihood of producing suicidal offspring. We show that, in the steady state, a population with suicide behaviour of this type grows faster than a population in which no individual ever commits suicide. The regrettable implication is that it is not evolutionarily inconsistent that suicidality may be a genetically determined trait.
Our goal in this paper was to put forward a qualitative theory and not a quantitatively testable model. We are aware, for instance, that the empirical suicide rate is above that predicted by our model—where it is of the order of the genetic mutation rate. However, it is not hard to modify our model, so that the suicide rate becomes arbitrarily high. A possible way would be to assume that the reproductive type is a multidimensional vector of binary variables. The choice of suicide is then conditional on each coordinate. It is easy to show that, in such a model, the probability of suicide is of the order of , where n is the number of dimensions of the reproductive type and ε is the probability with which each coordinate independently mutates.
Acknowledgments
Balázs Szentes gratefully acknowledges financial support from the European Research Council. Caroline Thomas gratefully acknowledges financial support from Deutsche Bank through the Institute for Advanced Study, Princeton.
Conflict of Interest
The authors declare no conflict of interest.
References
- De Catanzaro, D. A mathematical model of evolutionary pressures regulating self-preservation and self-destruction. Suicide Life Threat. Behav. 1986, 16, 166–181. [Google Scholar] [PubMed]
- Hamilton, W.D. The genetical evolution of social behaviour. I. J. Theor. Biol. 1964, 7, 1–16. [Google Scholar] [CrossRef]
- Värnik, P. Suicide in the world. Int. J. Environ. Res. Public Health 2012, 9, 760–771. [Google Scholar] [CrossRef] [PubMed]
- Brent, D.A.; Mann, J.J. Family genetic studies, suicide, and suicidal behavior. Am. J. Med. Genet. Part C: Semin. Med. Genet. 2005, 133, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Bondy, B.; Buettner, A.; Zill, P. Genetics of suicide. Mol. Psychiatry 2006, 11, 336–351. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.C.; Miller, G. Resolving the paradox of common, harmful, heritable mental disorders: Which evolutionary genetic models work best? Behav. Brain Sci. 2006, 29, 385–404. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.; Revicki, D.; Halpern, M.; Hatziandreu, E. The cost of suicide and suicide attempts in the United States. Clin. Neuropharmacol. 1995, 18, S25. [Google Scholar] [CrossRef]
- Hamermesh, D.S.; Soss, N.M. An economic theory of suicide. J. Polit. Econ. 1974, 82, 83–98. [Google Scholar] [CrossRef]
- Dixit, A.K.; Pindyck, R.S. Investment under Uncertainty; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Cutler, D.M.; Glaeser, E.L.; Norberg, K.E. Explaining the Rise in Youth Suicide. In Risky Behavior among Youths: An Economic Analysis; University of Chicago Press: Chicago, IL, USA, 2001; pp. 219–270. [Google Scholar]
- Becker, G.S.; Posner, R.A. Suicide: An economic approach. University of Chicago, 2004; (unpublished work). [Google Scholar]
- Rosenthal, R.W. Suicide attempts and signalling games. Math. Soc. Sci. 1993, 26, 25–33. [Google Scholar] [CrossRef]
- Marcotte, D.E. The economics of suicide, revisited. South. Econ. J. 2003, 69, 628–643. [Google Scholar] [CrossRef]
- Platt, S. Unemployment and suicidal behaviour: A review of the literature. Soc. Sci. Med. 1984, 19, 93–115. [Google Scholar] [CrossRef]
- Platt, S.; Hawton, K. Suicidal Behaviour and the Labour Market. In The International Handbook of Suicide and Attempted Suicide; Hawton, K, Van Heeringen, K, Eds.; John Wiley & Sons Ltd.: Chichester, 2000; pp. 309–384. [Google Scholar]
- Stack, S. Suicide: A 15-year review of the sociological literature. Part I: Cultural and economic factors. Suicide Life Threat. Behav. 2000, 30, 145–162. [Google Scholar] [PubMed]
- Stack, S. Suicide: A 15-year review of the sociological literature. Part II: Modernization and social integration perspectives. Suicide Life-Threat. Behav. 2000, 30, 163–176. [Google Scholar] [PubMed]
- Chen, J.; Choi, Y.J.; Mori, K.; Sawada, Y.; Sugano, S. Socio-economic studies on suicide: A survey. J. Econ. Surv. 2012, 26, 271–306. [Google Scholar] [CrossRef]
- 1.For a number of semelparous species (such as the Pacific salmon or the octopus), the decomposing bodies of adults assist in providing nutrients to the developing eggs in the very first stages of development. At the approach of a predator, a colonial rodent emitting alarm calls for the benefit of the colony makes itself vulnerable by attracting attention to itself. Additionally, in several species of termites and ants, individuals can make themselves explode (autothysis) to spray an assailant with a toxic substance.
- 2.Worldwide, an average of of deaths are caused by suicide, and the figure lies between and everywhere, except Africa () and the Middle East (), which are the regions with the highest shares of deaths due to war and conflict; see [] and the references therein.
- 3.See [] for a review of family studies of suicidal behaviour and [] for recent developments in the genetics of suicide.
- 4.Notwithstanding the unquantifiable emotional cost incurred by friends and family members, it has been estimated that in the US, in 1994, completed suicides cost billion in terms of lost earnings alone. Failed suicide attempts requiring hospitalisation or long-term disability support cost an additional billion. (See [].)
- 5.See, also, the brief discussion in [] pp. 24-25
- 6.For example, .
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).