1,25-Dihydroxyvitamin D3 (1,25(OH)2D3) Signaling Capacity and the Epithelial-Mesenchymal Transition in Non-Small Cell Lung Cancer (NSCLC): Implications for Use of 1,25(OH)2D3 in NSCLC Treatment

Abstract
:1. Introduction
2. Results and Discussion

{kind=link}
{kind=link}
{kind=link}
| VDR Phenotype | VDRhigh | VDRlow | |||||
|---|---|---|---|---|---|---|---|
| Lung Cancer Cell Line | H3122 | H292 | HCC827 | SK-LU-1 | H23 | A427 | A549 |
| Normalized VDR | 1.0 | 0.63 | 0.32 | 0.49 | 0.03 | 0.004 | 0.01 |
| Fold-induction CYP24A1 | 3504 | 9791 | 452 | 27116 | 7.2 | 1.2 | 1.6 |
| % Inhibition by 1,25(OH)2D3 | 70 | 73 * | 82 * | 30 | ND | ND | 4 * |
| EMT phenotype | E | E | M | M | |||
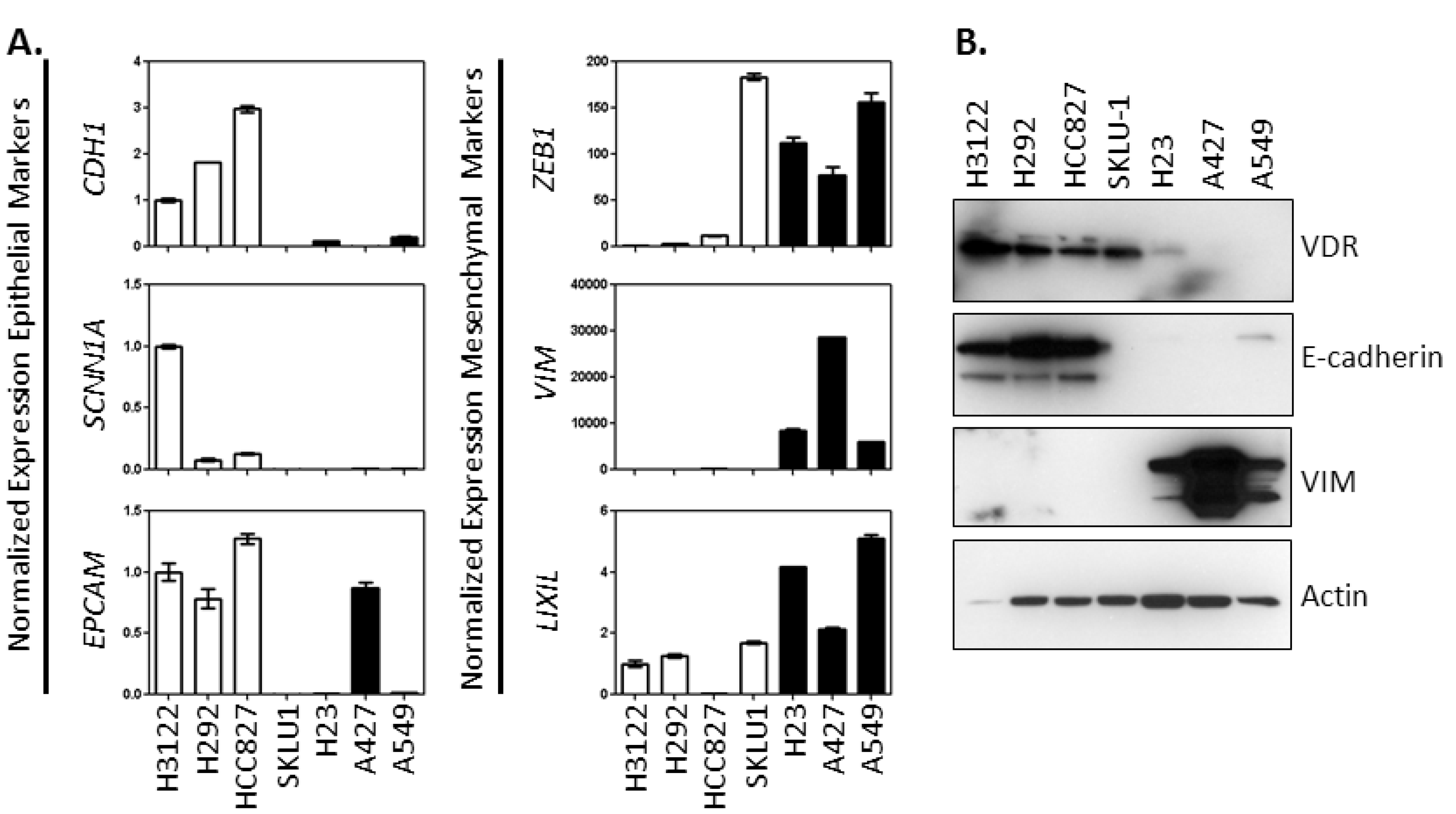
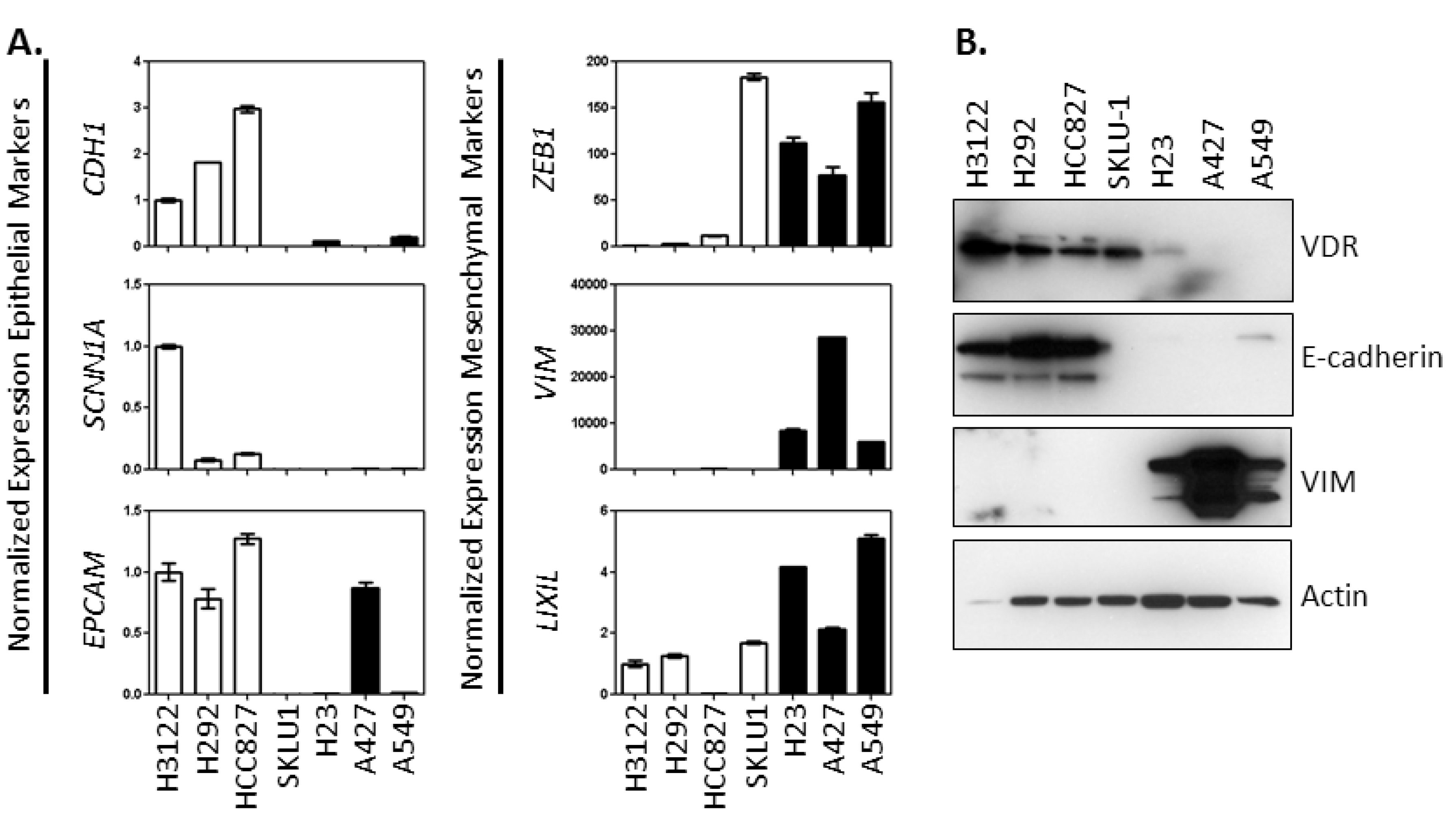
2.1. Characterization of the Association between Vitamin D Signaling Capacity and EMT Phenotype in NSCLC Cells
| Probe ID | Gene ID | Correlation | Probe ID | Gene ID | Correlation |
|---|---|---|---|---|---|
| 212764_at | ZEB1 | −0.39 | 210715_s_at | SPINT2 | 0.52 |
| 210875_s_at | ZEB1 | −0.36 | 219121_s_at | RBM35A | 0.52 |
| 201426_s_at | VIM | 0.01 | 205977_s_at | EPHA1 | 0.52 |
| 208510_s_at | PPARG | 0.03 | 37117_at | PRR5 | 0.54 |
| 201069_at | MMP2 | 0.06 | 205709_s_at | CDS1 | 0.55 |
| 207847_s_at | MUC1 | 0.10 | 220318_at | EPN3 | 0.57 |
| 202686_s_at | AXL | 0.11 | 210058_at | MAPK13 | 0.57 |
| 218792_s_at | BSPRY | 0.12 | 212070_at | GPR56 | 0.58 |
| 211732_x_at | HNMT | 0.24 | 203453_at | SCNN1A | 0.59 |
| 212298_at | NRP1 | 0.25 | 202525_at | PRSS8 | 0.59 |
| 202454_s_at | ERBB3 | 0.25 | 200606_at | DSP | 0.60 |
| 201839_s_at | TACSTD1 | 0.26 | 205980_s_at | PRR5 | 0.60 |
| 204112_s_at | HNMT | 0.27 | 213285_at | TMEM30B | 0.60 |
| 201428_at | CLDN4 | 0.29 | 219476_at | LRRC54 | 0.61 |
| 205847_at | PRSS22 | 0.29 | 218856_at | TNFRSF21 | 0.62 |
| 209488_s_at | RBPMS | 0.30 | 202489_s_at | FXYD3 | 0.62 |
| 35148_at | TJP3 | 0.30 | 203397_s_at | GALNT3 | 0.63 |
| 214702_at | FN1 | 0.32 | 221610_s_at | STAP2 | 0.64 |
| 202005_at | ST14 | 0.34 | 219919_s_at | SSH3 | 0.66 |
| 216905_s_at | ST14 | 0.36 | 203780_at | MPZL2 | 0.67 |
| 202790_at | CLDN7 | 0.38 | 219411_at | ELMO3 | 0.68 |
| 204503_at | EVPL | 0.40 | 218677_at | S100A14 | 0.68 |
| 65517_at | AP1M2 | 0.41 | 203256_at | CDH3 | 0.69 |
| 201506_at | TGFBI | 0.47 | 201650_at | KRT19 | 0.72 |
| 218186_at | RAB25 | 0.48 | 201131_s_at | CDH1 | 0.73 |
| 218261_at | AP1M2 | 0.49 | 204019_s_at | SH3YL1 | 0.74 |
| 211719_x_at | FN1 | 0.50 | 202286_s_at | TACSTD2 | 0.77 |
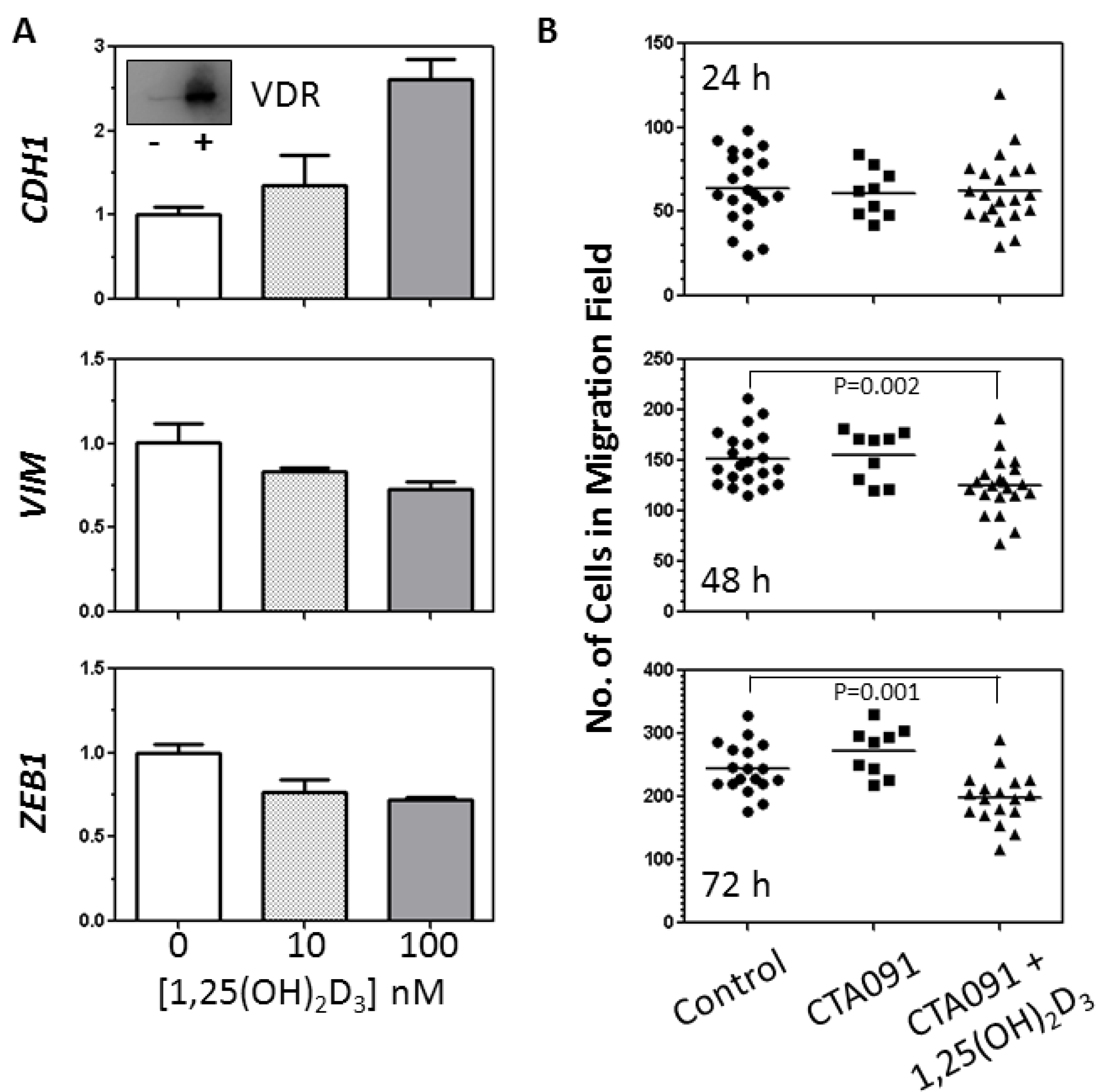
2.2. Analysis of the Effects of 1,25(OH)2D3 on EMT Related Genes and Migration of SK-LU-1 cells
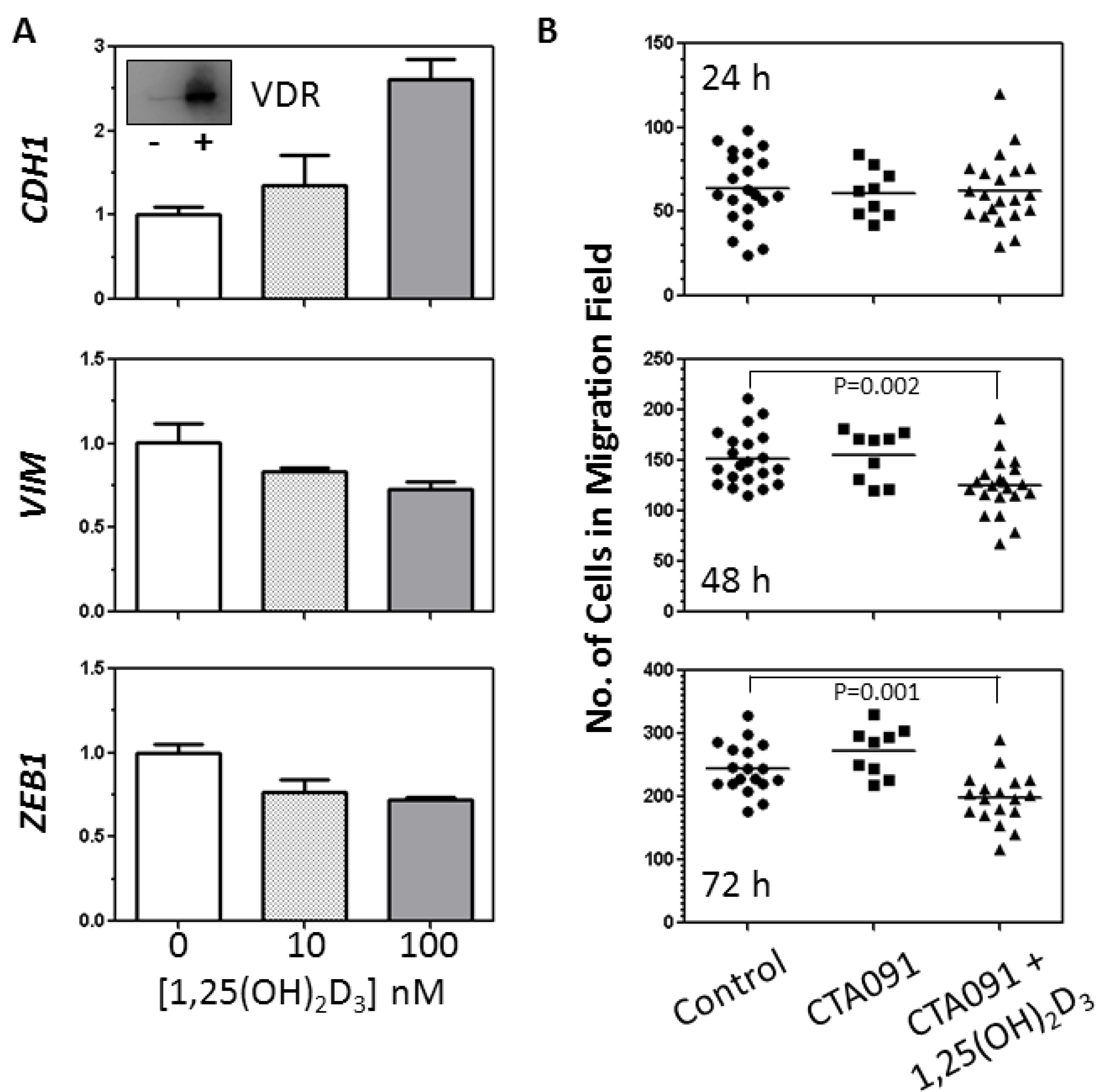
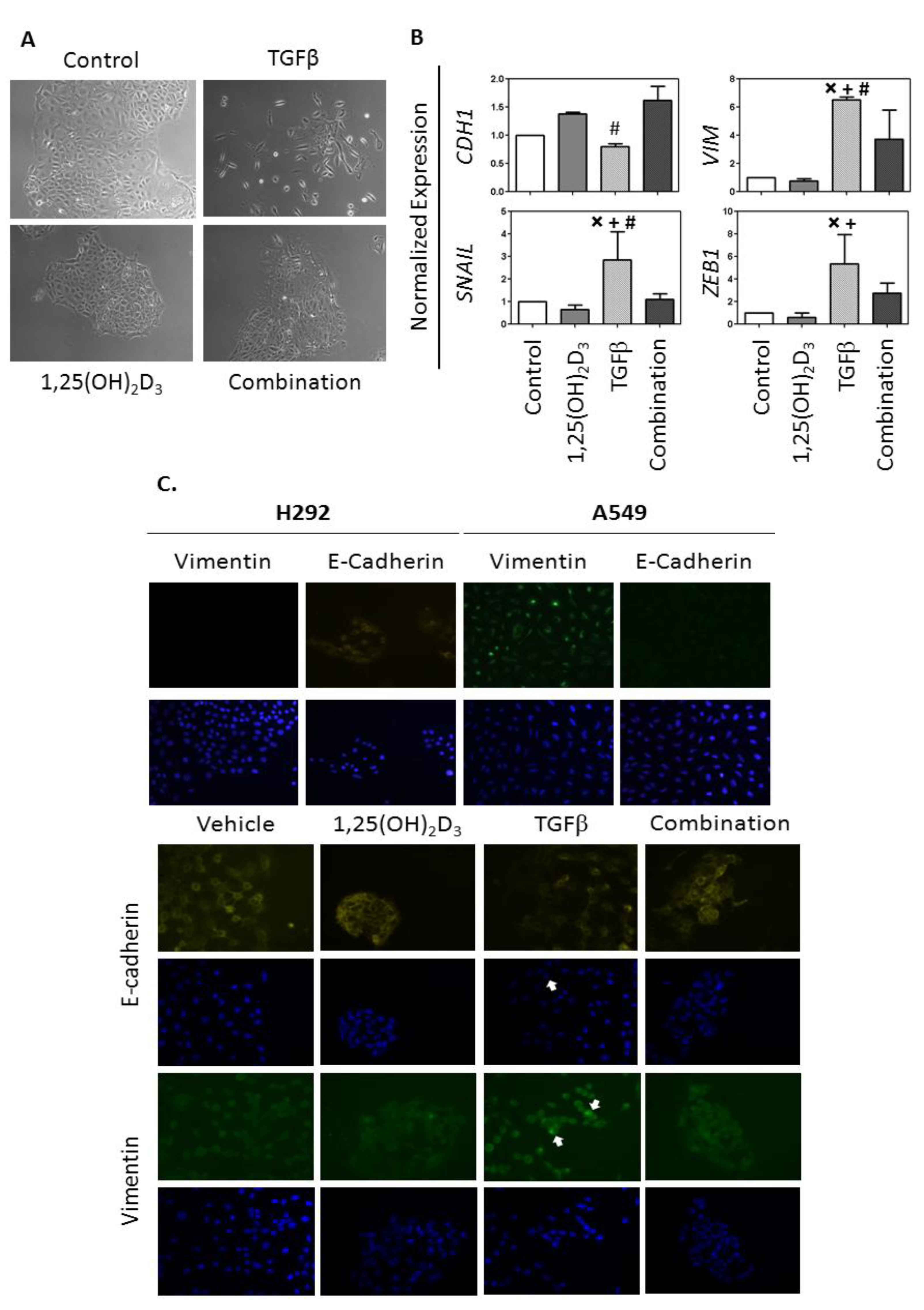
2.3. Analysis of the Effects of 1,25(OH)2D3 on TGFβ Induction of the EMT in VDRhigh NSCLC cells

2.4. Discussion
2.4.1. VDR Expression Is Associated with an Epithelial Phenotype and 1,25(OH)2D3 Sensitivity in NSCLC Cells
2.4.2. 1,25(OH)2D3 Opposes EMT Induction in NSCLC Cells
2.4.3. EMT Signature may Identify NSCLC Patients that Benefit from 1,25(OH)2D3 Treatment
3. Experimental
3.1. Cell Culture
3.2. Reagents and Chemicals
3.3. RNA Isolation
3.4. cDNA Synthesis
3.5. Real-Time PCR
| Gene | primer |
|---|---|
| VIM | F: 5'-TGCCCTTAAAGGAACCAATGAGTC-3' R: 5'-ATTCACGAAGGTGACGAGCCAT-3' |
| ZEB1 | F: 5'-TCCAGCCAAATGGAAATCAGGATG-3' R: 5'-CAGATTCCACACTCATGAGGTCTT-3' |
| SNAIL | F: 5'-TAGCGAGTGGTTCTTCTGCG-3' R: 5'-CTGCTGGAAGGTAAACTCTGGA-3' |
| CDH1 | F: 5'-TGGACCGAGAGAGTTTCCCT-3' R: 5'-ACGACGTTAGCCTCGTTCTC-3' |
| GAPDH | F: 5'-CTCCTCTGACTTCAACAGCG-3' R: 5'-GCCAAATTCGTTGTCATACCAG-3' |
| VDR | F: 5'-ATAAGACCTACGACCCCACCTA-3' R: 5'-GGACGAGTCCATCATGTCTGAA-3' |
| CYP24A1 | F: 5'-GCACAAGAGCCTCAACACCAA-3' R: 5'-AGACTGTTTGCTGTCGTTTCCA-3' |
| SCNN1A | F: 5'-GTCTCCCTCTGTCACGATGGTCA-3' R: 5'-ACCAGTATCGGCTTCGGAACCT-3' |
| EPCAM | F: 5'-GAGCGAGTGAGAACCTACTGG-3' R: 5'-ACGCGTTGTGATCTCCTTCT-3' |
| LIX1L | F: 5'-GCTTTGGGAGTTTCCAGTTTTGCC-3' R: 5'-CCCTGTATTTGGGTTGTCAGCTTC-3' |
3.6. Clonogenic Assay
3.7. Migration Assay
3.8. Preparation of Whole Cell Extracts and Immunoblotting
3.9. Immunofluorescence
3.10. Microarray Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Deeb, K.K.; Trump, D.L.; Johnson, C.S. Vitamin D signalling pathways in cancer: Potential for anticancer therapeutics. Nat. Rev. Cancer 2007, 7, 684–700. [Google Scholar] [CrossRef]
- Schwaller, J.; Koeffler, H.P.; Niklaus, G.; Loetscher, P.; Nagel, S.; Fey, M.F.; Tobler, A. Posttranscriptional stabilization underlies p53-independent induction of p21waf1/cip1/sdi1 in differentiating human leukemic cells. J. Clin. Invest. 1995, 95, 973–979. [Google Scholar] [CrossRef]
- Liu, M.; Lee, M.-H.; Cohen, M.; Bommakanti, M.; Freedman, L.P. Transcriptional activation of the Cdk inhibitor p21 by Vitamin D3 leads to the induced differentiation of the myelomonocytic cell line U937. Genes Dev. 1996, 10, 142–153. [Google Scholar] [CrossRef]
- Verlinden, L.; Verstuyf, A.; Convents, R.; Marcelis, S.; van Camp, M.; Bouillon, R. Action of 1,25(OH)2D3 on the cell cycle genes, cyclin D1, p21, and p27 in MCF-7 cells. Mol. Cell. Endocrinol. 1998, 142, 57–65. [Google Scholar] [CrossRef]
- Hager, G.; Formanek, M.; Gedlicka, C.; Thurnher, D.; Knerer, B.; Kornfehl, J. 1,25(OH)2 Vitamin D3 induces elevated expression of the cell cycle regulating genes p21 and p27 in squamous carcinoma cell lines of the head and neck. Acta Otolaryngol. 2001, 121, 102–109. [Google Scholar]
- Saramaki, A.; Banwell, C.M.; Campbell, M.J.; Carlberg, C. Regulation of the human p21(waf1/cip1) gene promoter via multiple binding sites for p53 and the Vitamin D3 receptor. Nucleic Acids Res. 2006, 34, 543–554. [Google Scholar] [CrossRef]
- Bernardi, R.J.; Johnson, C.S.; Modzelewski, R.A.; Trump, D.L. Antiproliferative effects of 1α,25-dihydroxyvitamin D3 and vitamin D analogs on tumor-derived endothelial cells. Endocrinology 2002, 143, 2508–2514. [Google Scholar] [CrossRef]
- Pendas-Franco, N.; Garcia, J.M.; Pena, C.; Valle, N.; Palmer, H.G.; Heinaniemi, M.; Carlberg, C.; Jimenez, B.; Bonilla, F.; Munoz, A.; et al. Dickkopf-4 is induced by tcf/beta-catenin and upregulated in human colon cancer, promotes tumour cell invasion and angiogenesis and is repressed by 1alpha,25-dihydroxyvitamin D3. Oncogene 2008, 27, 4467–4477. [Google Scholar] [CrossRef]
- Chung, I.; Han, G.; Seshadri, M.; Gillard, B.M.; Yu, W.D.; Foster, B.A.; Trump, D.L.; Johnson, C.S. Role of vitamin D receptor in the antiproliferative effects of calcitriol in tumor-derived endothelial cells and tumor angiogenesis in vivo. Cancer Res. 2009, 69, 967–975. [Google Scholar] [CrossRef]
- Ikeda, N.; Uemura, H.; Ishiguro, H.; Hori, M.; Hosaka, M.; Kyo, S.; Miyamoto, K.; Takeda, E.; Kubota, Y. Combination treatment with 1alpha,25-dihydroxyvitamin D3 and 9-cis-retinoic acid directly inhibits human telomerase reverse transcriptase transcription in prostate cancer cells. Mol. Cancer Ther. 2003, 2, 739–746. [Google Scholar]
- Kasiappan, R.; Shen, Z.; Tse, A.K.; Jinwal, U.; Tang, J.; Lungchukiet, P.; Sun, Y.; Kruk, P.; Nicosia, S.V.; Zhang, X.; et al. 1,25-dihydroxyvitamin D3 suppresses telomerase expression and human cancer growth through microrna-498. J. Biol. Chem. 2012, 287, 41297–41309. [Google Scholar] [CrossRef]
- Pereira, F.; Barbachano, A.; Silva, J.; Bonilla, F.; Campbell, M.J.; Munoz, A.; Larriba, M.J. Kdm6b/jmjd3 histone demethylase is induced by vitamin D and modulates its effects in colon cancer cells. Hum. Mol. Genet. 2011, 20, 4655–4665. [Google Scholar] [CrossRef]
- Pereira, F.; Barbachano, A.; Singh, P.K.; Campbell, M.J.; Munoz, A.; Larriba, M.J. Vitamin D has wide regulatory effects on histone demethylase genes. Cell Cycle 2012, 11, 1081–1089. [Google Scholar] [CrossRef] [Green Version]
- Pervin, S.; Hewison, M.; Braga, M.; Tran, L.; Chun, R.; Karam, A.; Chaudhuri, G.; Norris, K.; Singh, R. Down-regulation of vitamin D receptor in mammospheres: Implications for vitamin D resistance in breast cancer and potential for combination therapy. PLoS One 2013, 8, e53287. [Google Scholar] [CrossRef]
- Ma, Y.; Yu, W.D.; Su, B.; Seshadri, M.; Luo, W.; Trump, D.L.; Johnson, C.S. Regulation of motility, invasion, and metastatic potential of squamous cell carcinoma by 1alpha,25-dihydroxycholecalciferol. Cancer 2013, 119, 563–574. [Google Scholar] [CrossRef]
- De Craene, B.; Berx, G. Regulatory networks defining EMT during cancer initiation and progression. Nat. Rev. Cancer 2013, 13, 97–110. [Google Scholar] [CrossRef]
- Ren, J.; Chen, Y.; Song, H.; Chen, L.; Wang, R. Inhibition of zeb1 reverses EMT and chemoresistance in docetaxel-resistant human lung adenocarcinoma cell line. J. Cell. Biochem. 2013, 114, 1395–1403. [Google Scholar] [CrossRef]
- Gomez-Casal, R.; Bhattacharya, C.; Ganesh, N.; Bailey, L.; Basse, P.; Gibson, M.; Epperly, M.; Levina, V. Non-small cell lung cancer cells survived ionizing radiation treatment display cancer stem cell and epithelial-mesenchymal transition phenotypes. Mol. Cancer 2013, 12. [Google Scholar] [CrossRef]
- Thomson, S.; Buck, E.; Petti, F.; Griffin, G.; Brown, E.; Ramnarine, N.; Iwata, K.K.; Gibson, N.; Haley, J.D. Epithelial to mesenchymal transition is a determinant of sensitivity of non-small-cell lung carcinoma cell lines and xenografts to epidermal growth factor receptor inhibition. Cancer Res. 2005, 65, 9455–9462. [Google Scholar] [CrossRef]
- Witta, S.E.; Gemmill, R.M.; Hirsch, F.R.; Coldren, C.D.; Hedman, K.; Ravdel, L.; Helfrich, B.; Dziadziuszko, R.; Chan, D.C.; Sugita, M.; et al. Restoring E-cadherin expression increases sensitivity to epidermal growth factor receptor inhibitors in lung cancer cell lines. Cancer Res. 2006, 66, 944–950. [Google Scholar] [CrossRef]
- Yauch, R.L.; Januario, T.; Eberhard, D.A.; Cavet, G.; Zhu, W.; Fu, L.; Pham, T.Q.; Soriano, R.; Stinson, J.; Seshagiri, S.; et al. Epithelial versus mesenchymal phenotype determines in vitro sensitivity and predicts clinical activity of erlotinib in lung cancer patients. Clin. Cancer Res. 2005, 11, 8686–8698. [Google Scholar] [CrossRef]
- Zhang, Z.; Lee, J.C.; Lin, L.; Olivas, V.; Au, V.; LaFramboise, T.; Abdel-Rahman, M.; Wang, X.; Levine, A.D.; Rho, J.K.; et al. Activation of the AXL kinase causes resistance to EGFR-targeted therapy in lung cancer. Nat. Genet. 2012, 44, 852–860. [Google Scholar] [CrossRef]
- Byers, L.A.; Diao, L.; Wang, J.; Saintigny, P.; Girard, L.; Peyton, M.; Shen, L.; Fan, Y.; Giri, U.; Tumula, P.K.; et al. An epithelial-mesenchymal transition gene signature predicts resistance to EGFR and PI3K inhibitors and identifies Axl as a therapeutic target for overcoming EGFR inhibitor resistance. Clin. Cancer Res. 2013, 19, 279–290. [Google Scholar] [CrossRef]
- Zhang, Q.; Kanterewicz, B.; Shoemaker, S.; Hu, Q.; Liu, S.; Atwood, K.; Hershberger, P. Differential response to 1alpha,25-dihydroxyvitamin D3 (1alpha,25(OH)2D3) in non-small cell lung cancer cells with distinct oncogene mutations. J. Steroid Biochem. Mol. Biol. 2012, 136, 264–270. [Google Scholar]
- Zhang, Q.; Kanterewicz, B.; Buch, S.; Petkovich, M.; Parise, R.; Beumer, J.; Lin, Y.; Diergaarde, B.; Hershberger, P.A. Cyp24 inhibition preserves 1alpha,25-dihydroxyvitamin D3 anti-proliferative signaling in lung cancer cells. Mol. Cell. Endocrinol. 2012, 355, 153–161. [Google Scholar] [CrossRef]
- Ramirez, A.M.; Wongtrakool, C.; Welch, T.; Steinmeyer, A.; Zugel, U.; Roman, J. Vitamin D inhibition of pro-fibrotic effects of transforming growth factor beta1 in lung fibroblasts and epithelial cells. J. Steroid Biochem. Mol. Biol. 2010, 118, 142–150. [Google Scholar] [CrossRef]
- Parise, R.A.; Egorin, M.J.; Kanterewicz, B.; Taimi, M.; Petkovich, M.; Lew, A.M.; Chuang, S.S.; Nichols, M.; El-Hefnawy, T.; Hershberger, P.A. Cyp24, the enzyme that catabolizes the antiproliferative agent vitamin D, is increased in lung cancer. Int. J. Cancer 2006, 119, 1819–1828. [Google Scholar] [CrossRef]
- Xu, J.; Lamouille, S.; Derynck, R. TGF-beta-induced epithelial to mesenchymal transition. Cell Res. 2009, 19, 156–172. [Google Scholar] [CrossRef]
- Kim, S.H.; Chen, G.; King, A.N.; Jeon, C.K.; Christensen, P.J.; Zhao, L.; Simpson, R.U.; Thomas, D.G.; Giordano, T.J.; Brenner, D.E.; et al. Characterization of vitamin D receptor (VDR) in lung adenocarcinoma. Lung Cancer 2012, 77, 265–271. [Google Scholar] [CrossRef]
- Jeong, Y.; Xie, Y.; Lee, W.; Bookout, A.L.; Girard, L.; Raso, G.; Behrens, C.; Wistuba, I.I.; Gadzar, A.F.; Minna, J.D.; et al. Research resource: Diagnostic and therapeutic potential of nuclear receptor expression in lung cancer. Mol. Endocrinol. 2012, 26, 1443–1454. [Google Scholar] [CrossRef]
- Nguyen, K.S.; Neal, J.W. First-line treatment of EGFR-mutant non-small-cell lung cancer: The role of erlotinib and other tyrosine kinase inhibitors. Biologics 2012, 6, 337–345. [Google Scholar]
- Palmer, H.G.; Larriba, M.J.; Garcia, J.M.; Ordonez-Moran, P.; Pena, C.; Peiro, S.; Puig, I.; Rodriguez, R.; de la Fuente, R.; Bernad, A.; et al. The transcription factor snail represses vitamin D receptor expression and responsiveness in human colon cancer. Nat. Med. 2004, 10, 917–919. [Google Scholar] [CrossRef]
- Zhou, B.B.; Peyton, M.; He, B.; Liu, C.; Girard, L.; Caudler, E.; Lo, Y.; Baribaud, F.; Mikami, I.; Reguart, N.; et al. Targeting adam-mediated ligand cleavage to inhibit HER3 and EGFR pathways in non-small cell lung cancer. Cancer Cell 2006, 10, 39–50. [Google Scholar] [CrossRef]
- Maindonald, J.; Braun, J. Data Analysis and Graphics Using R, 2nd ed.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Creighton, C.J.; Gibbons, D.L.; Kurie, J.M. The role of epithelial-mesenchymal transition programming in invasion and metastasis: A clinical perspective. Cancer Manag. Res. 2013, 5, 187–195. [Google Scholar] [CrossRef]
- Kuo, C.H.; Chen, K.F.; Chou, S.H.; Huang, Y.F.; Wu, C.Y.; Cheng, D.E.; Chen, Y.W.; Yang, C.J.; Hung, J.Y.; Huang, M.S. Lung tumor associated dendritic cell-derived resistin promoted cancer progression by increasing wolf-hirschhorn syndrome candidate 1/twist pathway. Carcinogenesis 2013. [Google Scholar] [CrossRef]
- Zhou, W.; Heist, R.S.; Liu, G.; Asomaning, K.; Neuberg, D.S.; Hollis, B.W.; Wain, J.C.; Lynch, T.J.; Giovannucci, E.; Su, L.; et al. Circulating 25-hydroxyvitamin D levels predict survival in early-stage non-small-cell lung cancer patients. J. Clin. Oncol. 2007, 25, 479–485. [Google Scholar] [CrossRef]
- Zhou, W.; Suk, R.; Liu, G.; Park, S.; Neuberg, D.S.; Wain, J.C.; Lynch, T.J.; Giovannucci, E.; Christiani, D.C. Vitamin D is associated with improved survival in early-stage non-small cell lung cancer patients. Cancer Epidemiol. Biomarkers Prev. 2005, 14, 2303–2309. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Upadhyay, S.K.; Verone, A.; Shoemaker, S.; Qin, M.; Liu, S.; Campbell, M.; Hershberger, P.A. 1,25-Dihydroxyvitamin D3 (1,25(OH)2D3) Signaling Capacity and the Epithelial-Mesenchymal Transition in Non-Small Cell Lung Cancer (NSCLC): Implications for Use of 1,25(OH)2D3 in NSCLC Treatment. Cancers 2013, 5, 1504-1521. https://doi.org/10.3390/cancers5041504
Upadhyay SK, Verone A, Shoemaker S, Qin M, Liu S, Campbell M, Hershberger PA. 1,25-Dihydroxyvitamin D3 (1,25(OH)2D3) Signaling Capacity and the Epithelial-Mesenchymal Transition in Non-Small Cell Lung Cancer (NSCLC): Implications for Use of 1,25(OH)2D3 in NSCLC Treatment. Cancers. 2013; 5(4):1504-1521. https://doi.org/10.3390/cancers5041504
Chicago/Turabian StyleUpadhyay, Santosh Kumar, Alissa Verone, Suzanne Shoemaker, Maochun Qin, Song Liu, Moray Campbell, and Pamela A. Hershberger. 2013. "1,25-Dihydroxyvitamin D3 (1,25(OH)2D3) Signaling Capacity and the Epithelial-Mesenchymal Transition in Non-Small Cell Lung Cancer (NSCLC): Implications for Use of 1,25(OH)2D3 in NSCLC Treatment" Cancers 5, no. 4: 1504-1521. https://doi.org/10.3390/cancers5041504