Genomic Instability: The Driving Force behind Refractory/Relapsing Hodgkin’s Lymphoma


Abstract
:1. Introduction

2. Nuclear Architecture and Function
2.1. Chromosome Territories
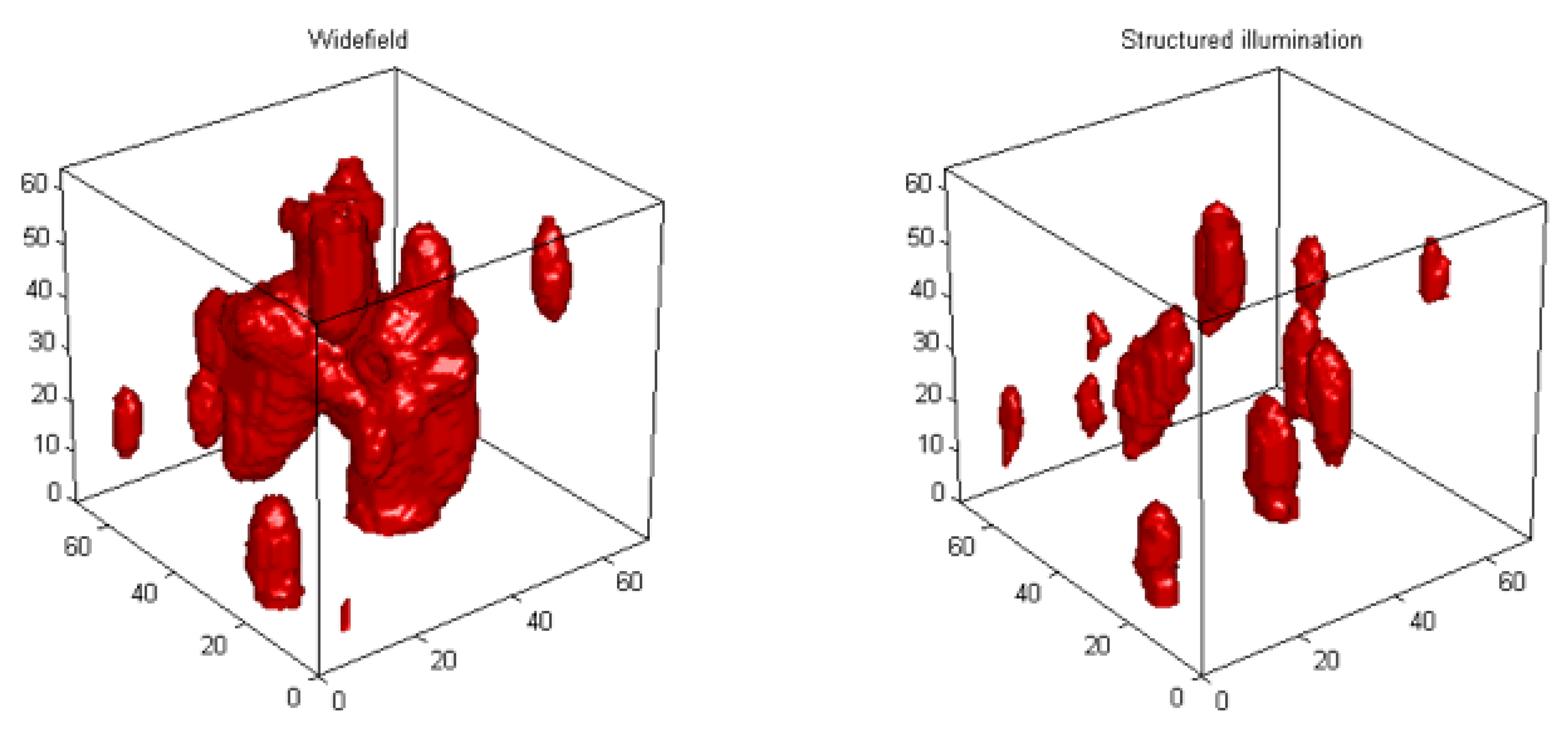
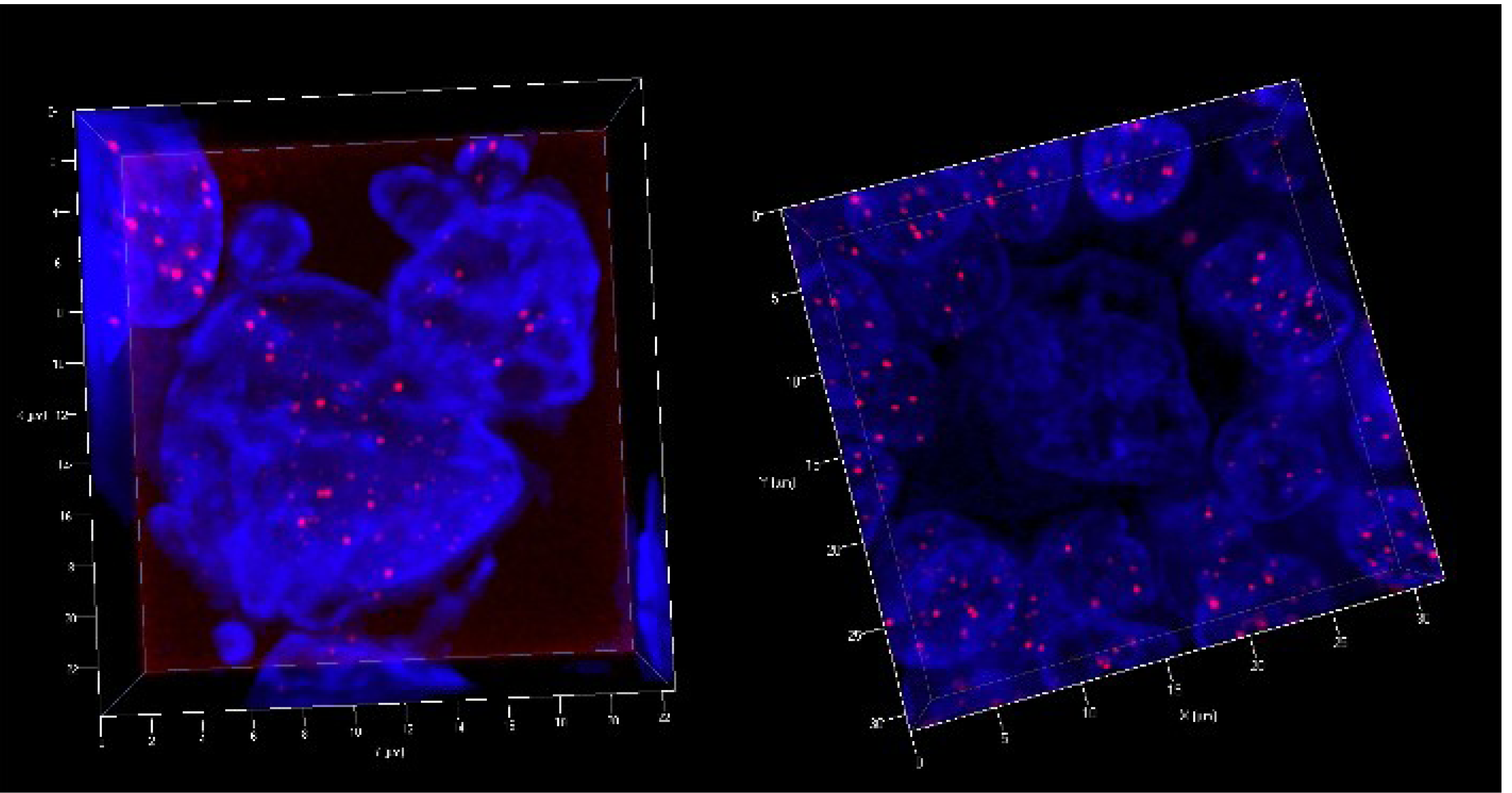
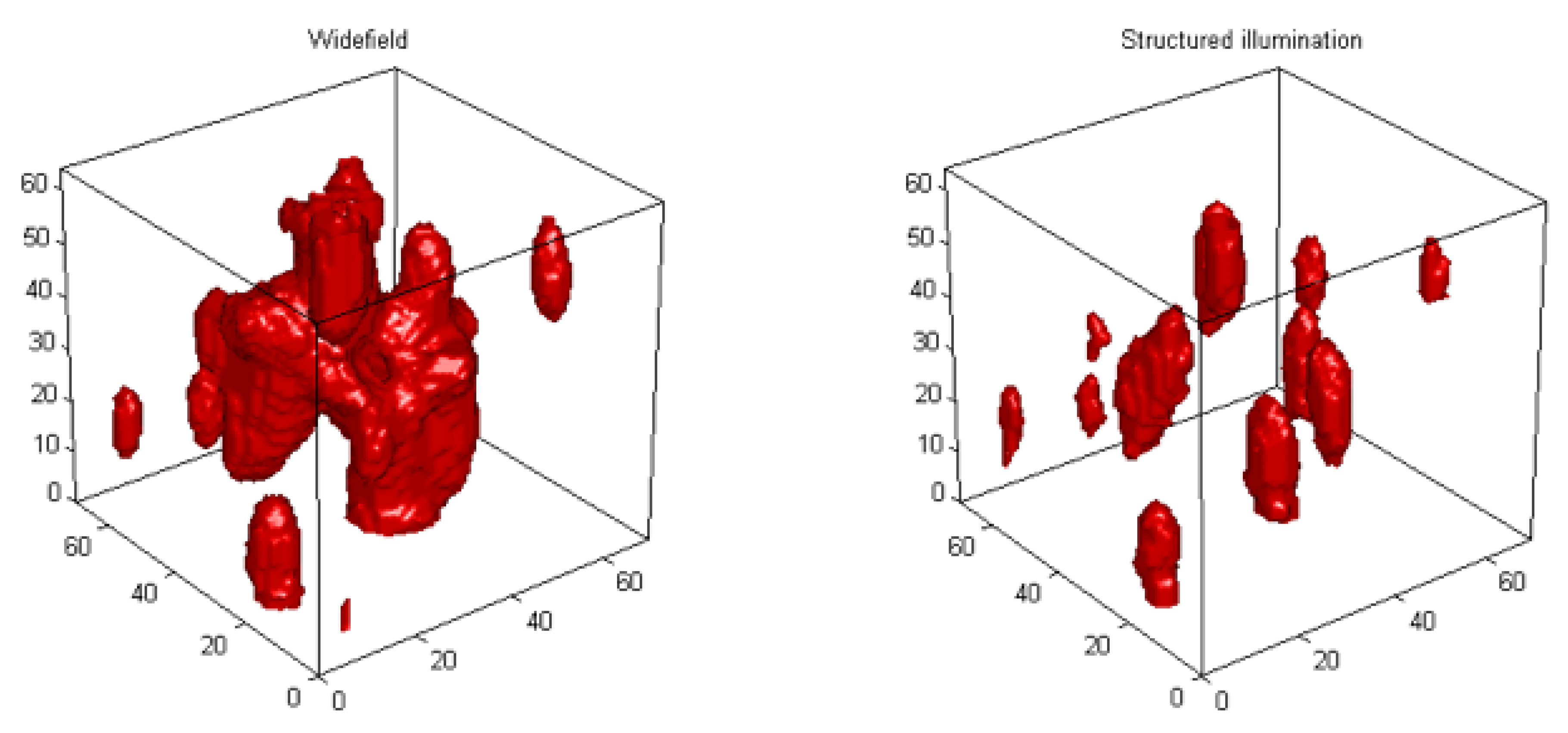
3. 3D Telomere Dynamics in Translational Oncology



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Sex | Age | Type | LMP1 | Stage | Rapid | 0–5,000 u |
|---|---|---|---|---|---|---|---|
| very small telomeres (%) | |||||||
| 1 | F | 57 | MC | Pos | IIIA | 4 ABVD | 39.3 |
| 2 | M | 24 | MC | Pos | IIIB | 1 ABVD | 37.3 |
| 3 | M | 42 | MC | Neg | IIA | 2 ABVD | 27.8 |
| 4 | M | 19 | SN | Neg | IVA | 2 ABVD | 35.9 |
| 5 | F | 28 | SN | Pos | IIA | 4 ABVD | 32.7 |
| 6 | M | 27 | SN | Neg | IIIs | 4 ABVD | 46.4 |
| 7 | M | 34 | SN | Neg | IIIB | 4 ABVD | 16.7 |
| 12 | M | 71 | MC | Pos | IIIB | 4 ABVD | 34.3 |
| 16 | F | 73 | NS | Neg | IIIB | 4 ABVD | 78.6 |
| 11 | M | 17 | NS | Neg | IIB bulky | 4 DBVE-PC+IFRT | 49.8 |
| Case | Sex | Age | Type | LMP1 | Stage | 1st Line | 0–5,000 u |
|---|---|---|---|---|---|---|---|
| very small telomeres (%) | |||||||
| 17a | F | 77 | NS | Pos | IIIB | 4 AVBD | 62.3 progressive |
| 19a | M | 61 | NS | Neg | IIIB | 6 ABVD | 68.2 early relapse |
| 20a | M | 38 | NS | Neg | IVB | 6 ABVD | 77.9 late relapse |
| 21a | M | 21 | NS | Neg | IIIB | 4 AVBD | 48.1 progressive |
| 22a | M | 49 | NS | Neg | IIIB | 6 AVBD | 71.4 late relapse |
| 23a | F | 14 | NS | Neg | IVA | 4 DVBE-PC + IFRT | 77.6 early relapse |
| 25a | M | 48 | NS | Neg | IVB | 6 ABVD | 52.9 early relapse |
| 26a | M | 28 | NS | Neg | IIB | 4 ABVD | 54.0 early relapse |
| 10a | F | 35 | NS | Neg | IIIsB | 2 CVPP-A0 | 74.1 late relapse |
| 40a | F | 44 | NS | Pos | IVB | 2 ABVD | 69.9 progressive |
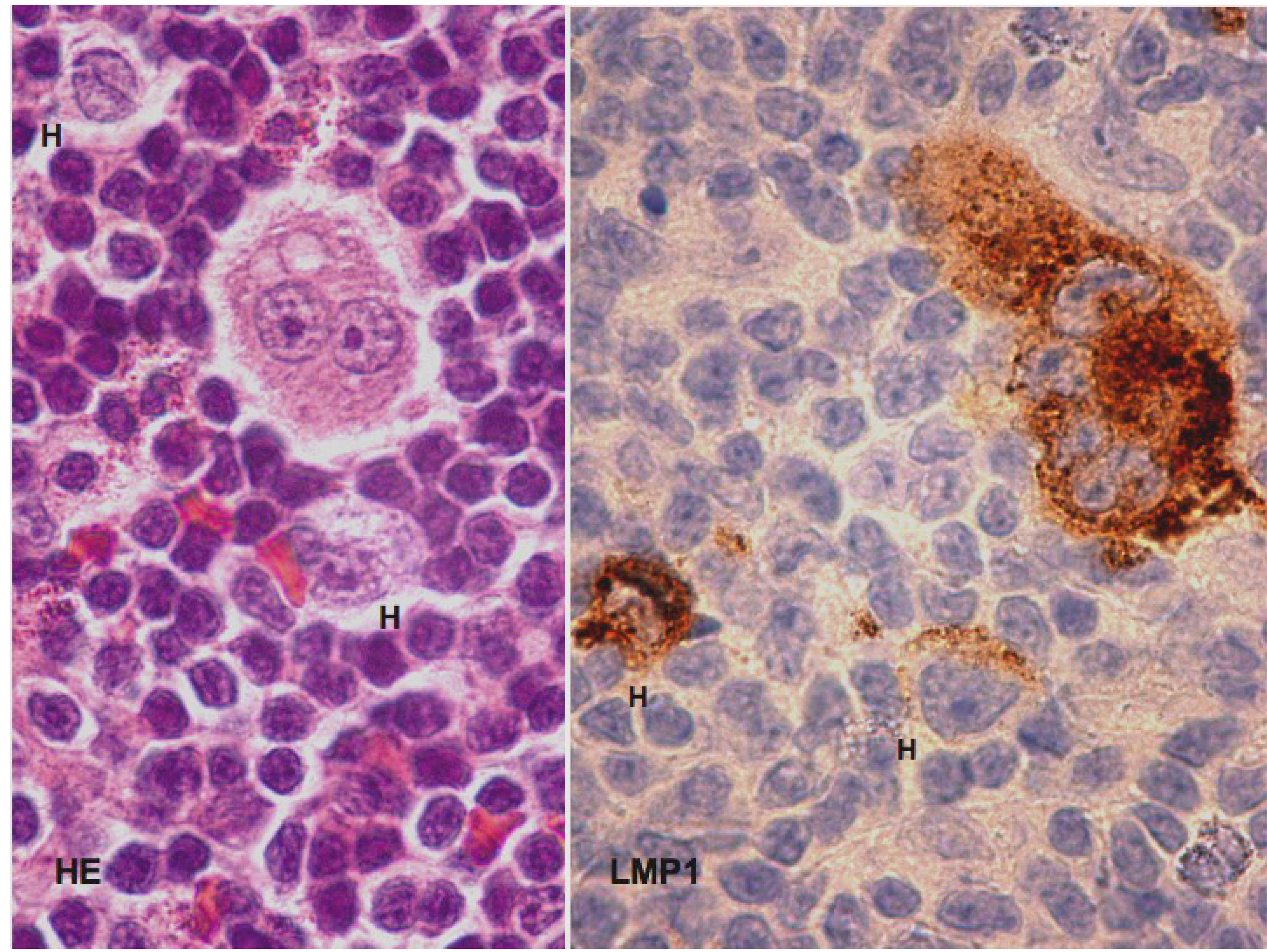
4. EBV Is probably Not an Innocent Bystander
5. The Telomere-Shelterin Complex: Achilles Tendon of Relapsing/Refractory HL
6. Conclusions
References
- Küppers, R.; Engert, A.; Hansmann, M.L. Hodgkin lymphoma. J. Clin. Invest. 2012, 122, 3439–3447. [Google Scholar] [CrossRef]
- Küppers, R. New insights in the biology of Hodgkin lymphoma. Hematology Am. Soc. Hematol. Educ. Program 2012, 2012, 328–334. [Google Scholar]
- Steidl, C.; Connors, J.M.; Gascoyne, R.D. Molecular pathogenesis of Hodgkin’s lymphoma: Increasing evidence of the importance of the microenvironment. J. Clin. Oncol. 2011, 29, 1812–1826. [Google Scholar] [CrossRef]
- Farrell, K.; Jarrett, R.F. The molecular pathogenesis of Hodgkin lymphoma. Histopathology 2011, 58, 15–25. [Google Scholar] [CrossRef]
- Schwarzer, R.; Jundt, F. Notch and NF-κB signaling pathways in the biology of classical Hodgkin lymphoma. Curr. Mol. Med. 2011, 11, 236–245. [Google Scholar] [CrossRef]
- Knecht, H.; Brüderlein, S.; Mai, S.; Möller, P.; Sawan, B. 3D structural and functional characterization of the transition from Hodgkin to Reed-Sternberg cells. Ann. Anat. 2010, 192, 302–308. [Google Scholar] [CrossRef]
- Gadji, M.; Vallente, R.; Klewes, L.; Righolt, C.; Wark, L.; Kongruttanachok, N.; Knecht, H.; Mai, S. Nuclear remodeling as a mechanism for genomic instability in cancer. Adv. Cancer Res. 2011, 112, 77–126. [Google Scholar] [CrossRef]
- Cremer, T.; Cremer, M.; Dietzel, S.; Müller, S.; Solovei, I.; Fakan, S. Chromosome territories-a functional nuclear landscape. Curr. Opin. Cell Biol. 2006, 18, 307–316. [Google Scholar] [CrossRef]
- Lanctôt, C.; Cheutin, T.; Cremer, M.; Cavalli, G.; Cremer, T. Dynamic genome architecture in the nuclear space: regulation of gene expression in three dimensions. Nat. Rev. Genet. 2007, 8, 104–115. [Google Scholar] [CrossRef]
- Solovei, I.; Kreysing, M.; Lanctôt, C.; Kösem, S.; Peichl, L.; Cremer, T.; Guck, J.; Joffe, B. Nuclear architecture of rod photoreceptor cells adapts to vision in mammalian evolution. Cell 2009, 137, 356–368. [Google Scholar] [CrossRef]
- Londoño-Vallejo, J.A.; Wellinger, R.J. Telomeres and telomerase dance to the rhythm of the cell cycle. Trends Biochem. Sci. 2012, 37, 391–399. [Google Scholar] [CrossRef]
- Chuang, T.C.; Moshir, S.; Garini, Y.; Chuang, A.Y.; Young, I.T.; Vermolen, B.; van den Doel, R.; Mougey, V.; Perrin, M.; Braun, M.; et al. The three-dimensional organization of telomeres in the nucleus of mammalian cells. BMC Biol. 2004, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Vermolen, B.J.; Garini, Y.; Mai, S.; Mougey, V.; Fest, T.; Chuang, T.C.; Chuang, A.Y.; Wark, L.; Young, I.T. Characterizing the three-dimensional organization of telomeres. Cytometry A 2005, 67, 144–150. [Google Scholar]
- Mai, S.; Garini, Y. The significance of telomeric aggregates in the interphase nuclei of tumor cells. J. Cell. Biochem. 2006, 97, 904–915. [Google Scholar]
- Louis, S.F.; Vermolen, B.J.; Garini, Y.; Young, I.T.; Guffei, A.; Lichtensztejn, Z.; Kuttler, F.; Chuang, T.C.; Moshir, S.; Mougey, V.; et al. c-Myc induces chromosomal rearrangements through telomere and chromosome remodeling in the interphase nucleus. Proc. Natl. Acad. Sci. USA 2005, 102, 9613–9618. [Google Scholar] [CrossRef]
- Lacoste, S.; Wiechec, E.; dos Santos Silva, A.G.; Guffei, A.; Williams, G.; Lowbeer, M.; Benedek, K.; Henriksson, M.; Klein, G.; Mai, S. Chromosomal rearrangements after ex vivo Epstein-Barr virus (EBV) infection of human B cells. Oncogene 2010, 29, 503–515. [Google Scholar] [CrossRef]
- Schermelleh, L.; Carlton, P.M.; Haase, S.; Shao, L.; Winoto, L.; Kner, P.; Burke, B.; Cardoso, M.C.; Agard, D.A.; Gustafsson, M.G.; et al. Subdiffraction multicolor imaging of the nuclear periphery with 3D structured illumination microscopy. Science 2008, 320, 1332–1336. [Google Scholar] [CrossRef]
- Xu, L.; Blackburn, E.H. Human cancer cells harbour T-stumps, a distinct class of extremely short telomeres. Mol. Cell 2007, 28, 315–327. [Google Scholar] [CrossRef]
- Knecht, H.; Sawan, B.; Lichtensztejn, D.; Lemieux, B.; Wellinger, R.; Mai, S. The 3D nuclear organization of telomeres marks the transition from Hodgkin to Reed-Sternberg cells. Leukemia 2009, 23, 565–573. [Google Scholar] [CrossRef]
- Knecht, H.; Brüderlein, S.; Wegener, S.; Lichtensztejn, D.; Lichtensztejn, Z.; Möller, P.; Mai, S. 3D nuclear organization of telomeres in the Hodgkin cell lines U-HO1 and U-HO1-PTPN1: PTPN1 expression prevents the formation of very short telomeres including “t-stumps”. BMC Cell Biol. 2010, 11, 99. [Google Scholar] [CrossRef]
- Guffei, A.; Sarkar, R.; Klewes, L.; Righolt, C.; Knecht, H.; Mai, S. Dynamic chromosomal rearrangements in Hodgkin’s lymphoma are due to ongoing three-dimensional nuclear remodeling and breakage-bridge-fusion cycles. Haematologica 2010, 95, 2038–2046. [Google Scholar] [CrossRef]
- Knecht, H.; Sawan, B.; Lichtensztejn, Z.; Lichtensztejn, D.; Mai, S. 3D Telomere FISH defines LMP1 expressing Reed-Sternberg Cells as End-Stage Cells with Telomere-poor “Ghost” Nuclei and very short Telomeres. Lab. Invest. 2010, 90, 611–619. [Google Scholar] [CrossRef]
- Knecht, H.; Kongruttanachok, N.; Sawan, B.; Brossard, J.; Prévost, P.; Turcotte, E.; Lichtensztejn, Z.; Lichtensztejn, D.; Mai, S. 3D telomere signatures of Hodgkin- and Reed-Sternberg cells at diagnosis identify patients with poor response to conventional chemotherapy. Transl. Oncol. 2012, 5, 269–277. [Google Scholar]
- Lajoie, V.; Lemieux, B.; Sawan, B.; Lichtensztejn, D.; Lichtensztejn, Z.; Rowe, M.; Wellinger, R.; Mai, S.; Knecht, H. The Epstein-Barr virus (EBV) encoded oncoprotein LMP1 mediates down regulation of shelterin proteins, formation of telomere aggregates and multi-nuclearity. ASH Annual Meeting Abstracts 2010, 116, 3625. [Google Scholar]
- Kamranvar, S.A.; Masucci, M.G. The Epstein-Barr virus nuclear antigen-1 promotes telomere dysfunction via induction of oxidative stress. Leukemia 2011, 25, 1017–1025. [Google Scholar] [CrossRef]
- Deng, Z.; Wang, Z.; Lieberman, P.M. Telomeres and viruses: Common themes of genome maintenance. Front. Oncol. 2012, 2, 201. [Google Scholar]
- Faumont, N.; Chanut, A.; Benard, A.; Cogne, N.; Delsol, G.; Feuillard, J.; Meggetto, F. Comparative analysis of oncogenic properties and nuclear factor-kappaB activity of latent membrane protein 1 natural variants from Hodgkin’s lymphoma’s Reed-Sternberg cells and normal B-lymphocytes. Haematologica 2009, 94, 355–363. [Google Scholar] [CrossRef]
- Zuercher, E.; Butticaz, C.; Wyniger, J.; Martinez, R.; Battegay, M.; Boffi El Amari, E.; Dang, T.; Egger, J.F.; Fehr, J.; Mueller-Garamvögyi, E.; et al. Genetic diversity of EBV-encoded LMP1 in the Swiss HIV Cohort Study and implication for NF-κB activation. PLoS One 2012, 7, e32168. [Google Scholar]
- Tiacci, E.; Döring, C.; Brune, V.; van Noesel, C.J.; Klapper, W.; Mechtersheimer, G.; Falini, B.; Küppers, R.; Hansmann, M.L. Aalyzing primary Hodgkin and Reed-Sternberg cells to capture the molecular and cellular pathogenesis of classical Hodgkin lymphoma. Blood 2012, 120, 4609–4620. [Google Scholar]
- Krem, M.M.; Luo, P.; Ing, B.I.; Horwitz, M.S. The kelch protein KLHDC8B guards against mitotic errors, centrosomal amplification, and chromosomal instability. J. Biol. Chem. 2012, 287, 39083–39093. [Google Scholar]
- Szczepanowski, M.; Masqué-Soler, N.; Oschlies, I.; Schmidt, W.; Lück, A.; Klapper, W. Composite lymphoma of nodular lymphocyte-predominant and classical Hodgkin lymphoma-Epstein-Barr virus association suggests divergent pathogenesis despite clonal relatedness. Hum. Pathol. 2013, in press. [Google Scholar]
- Younes, A.; Bartlett, N.L.; Leonard, J.P.; Kennedy, D.A.; Lynch, C.M.; Sievers, E.L.; Forero-Torres, A. Brentuximab vedotin (SGN-35) for relapsed CD30-positive lymphomas. N. Engl. J. Med. 2010, 363, 1812–1821. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Knecht, H.; Righolt, C.; Mai, S. Genomic Instability: The Driving Force behind Refractory/Relapsing Hodgkin’s Lymphoma. Cancers 2013, 5, 714-725. https://doi.org/10.3390/cancers5020714
Knecht H, Righolt C, Mai S. Genomic Instability: The Driving Force behind Refractory/Relapsing Hodgkin’s Lymphoma. Cancers. 2013; 5(2):714-725. https://doi.org/10.3390/cancers5020714
Chicago/Turabian StyleKnecht, Hans, Christiaan Righolt, and Sabine Mai. 2013. "Genomic Instability: The Driving Force behind Refractory/Relapsing Hodgkin’s Lymphoma" Cancers 5, no. 2: 714-725. https://doi.org/10.3390/cancers5020714