Glioma Surgical Aspirate: A Viable Source of Tumor Tissue for Experimental Research

Abstract
:1. Introduction
2. Results and Discussion
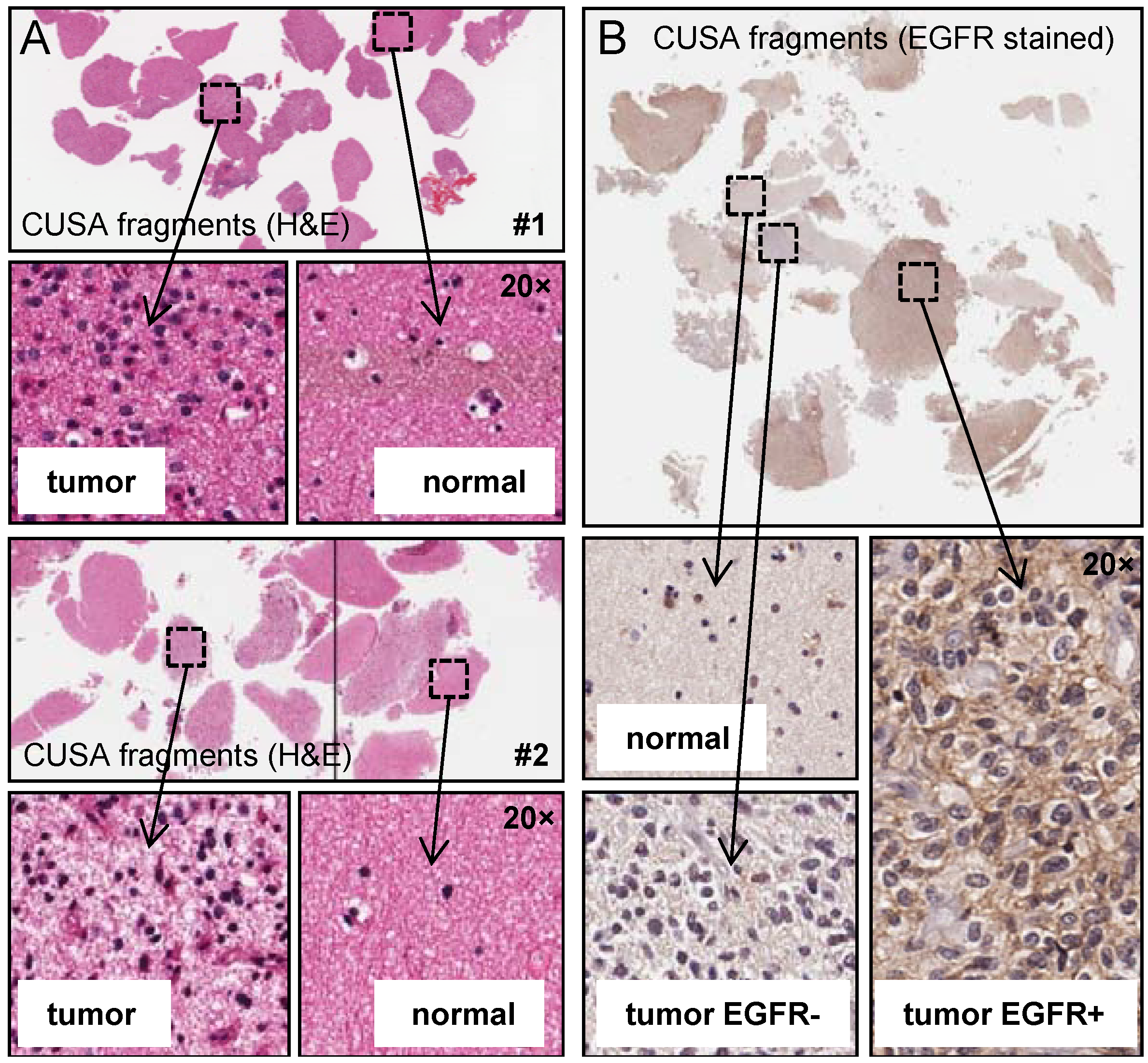
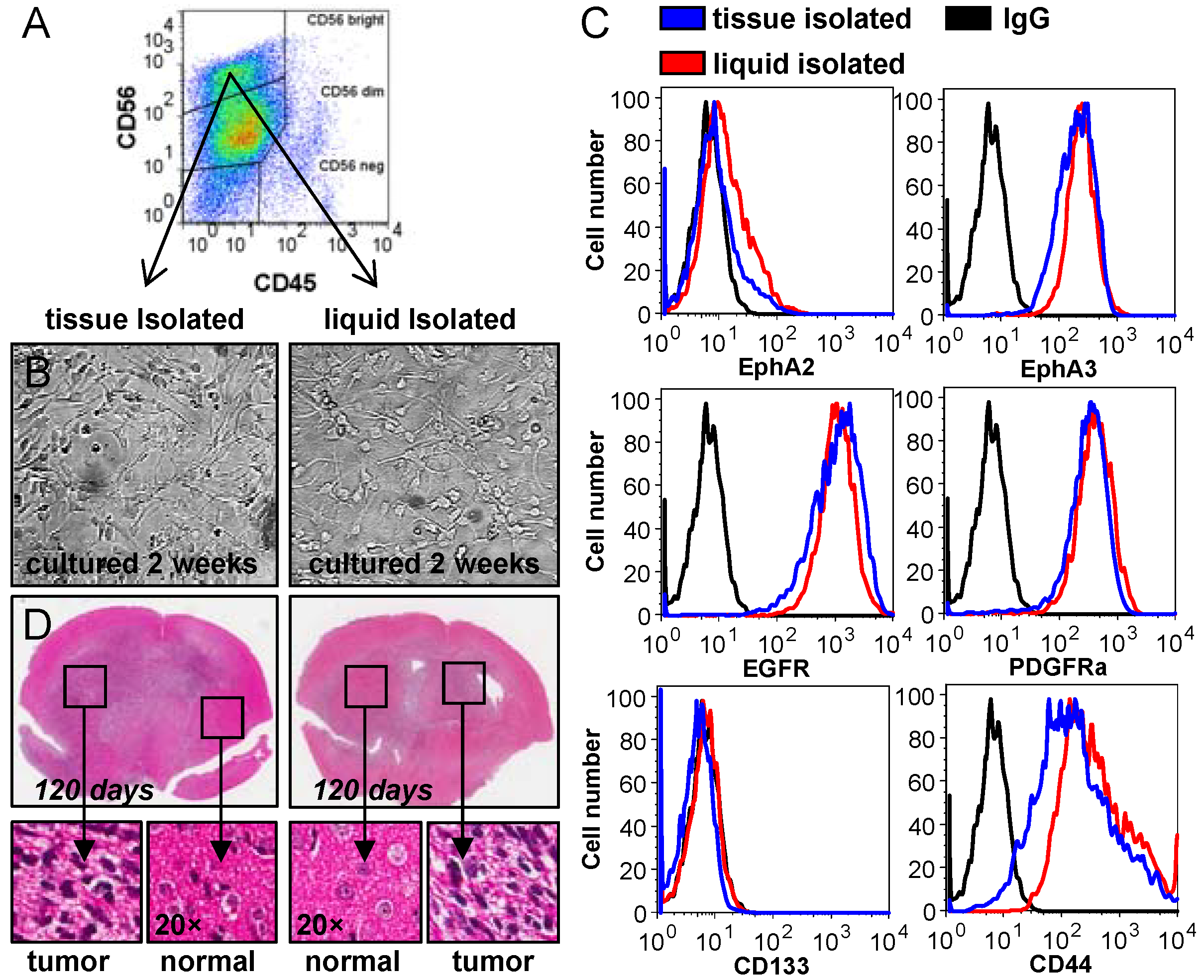
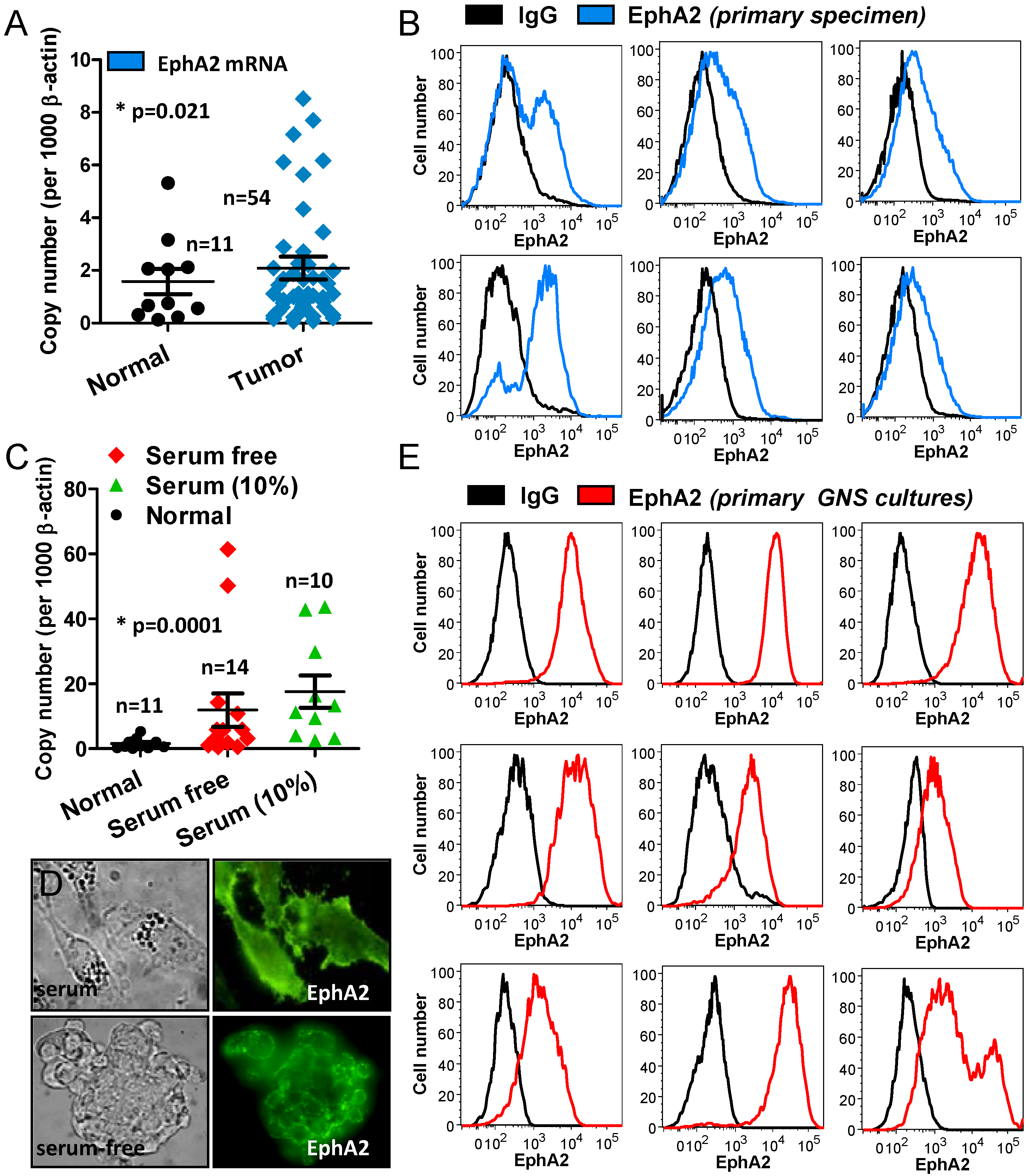
2.1. Analysis of CUSA-Derived Tissue
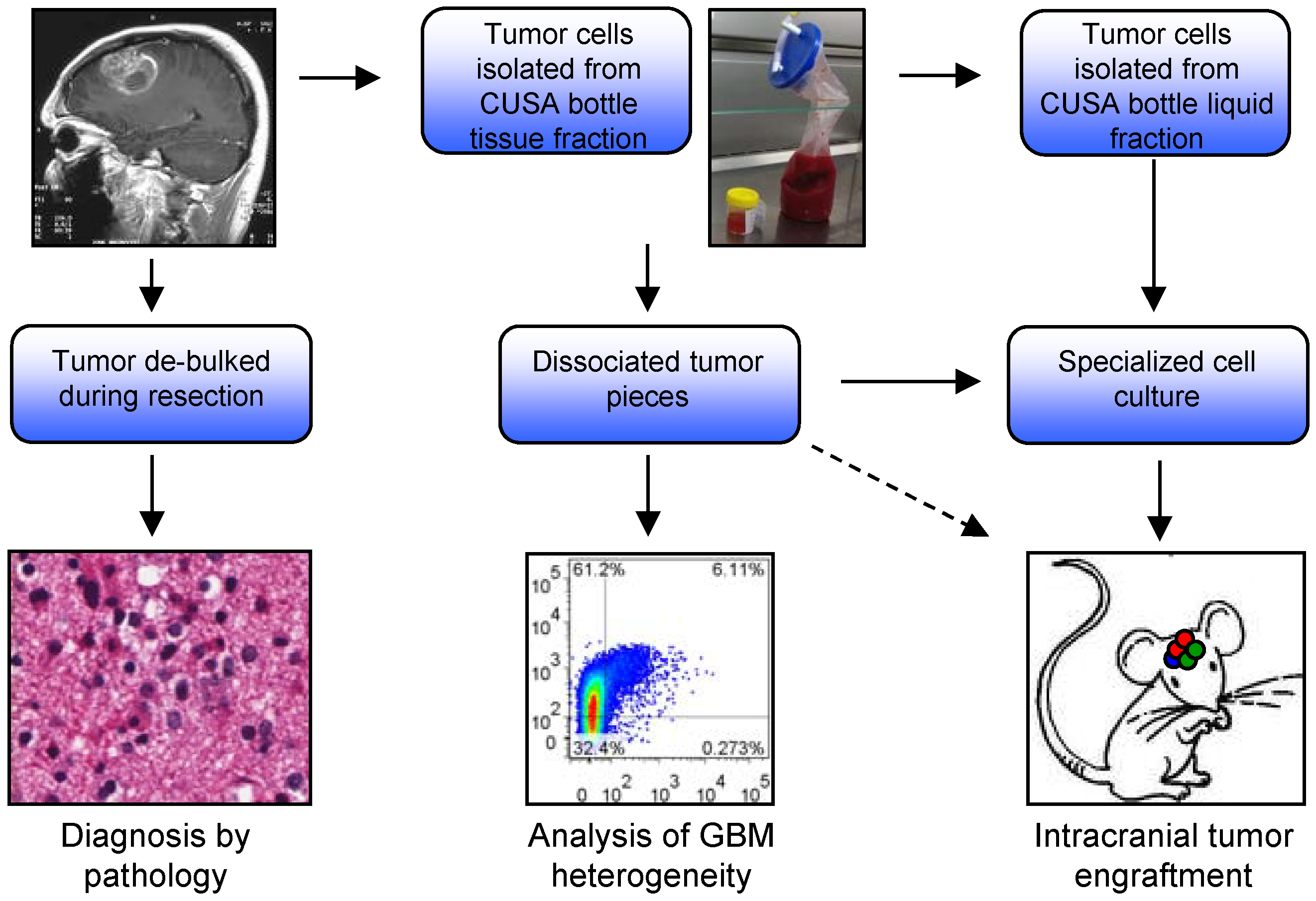

2.2. Analysis of CUSA-Isolated Tumor Cells
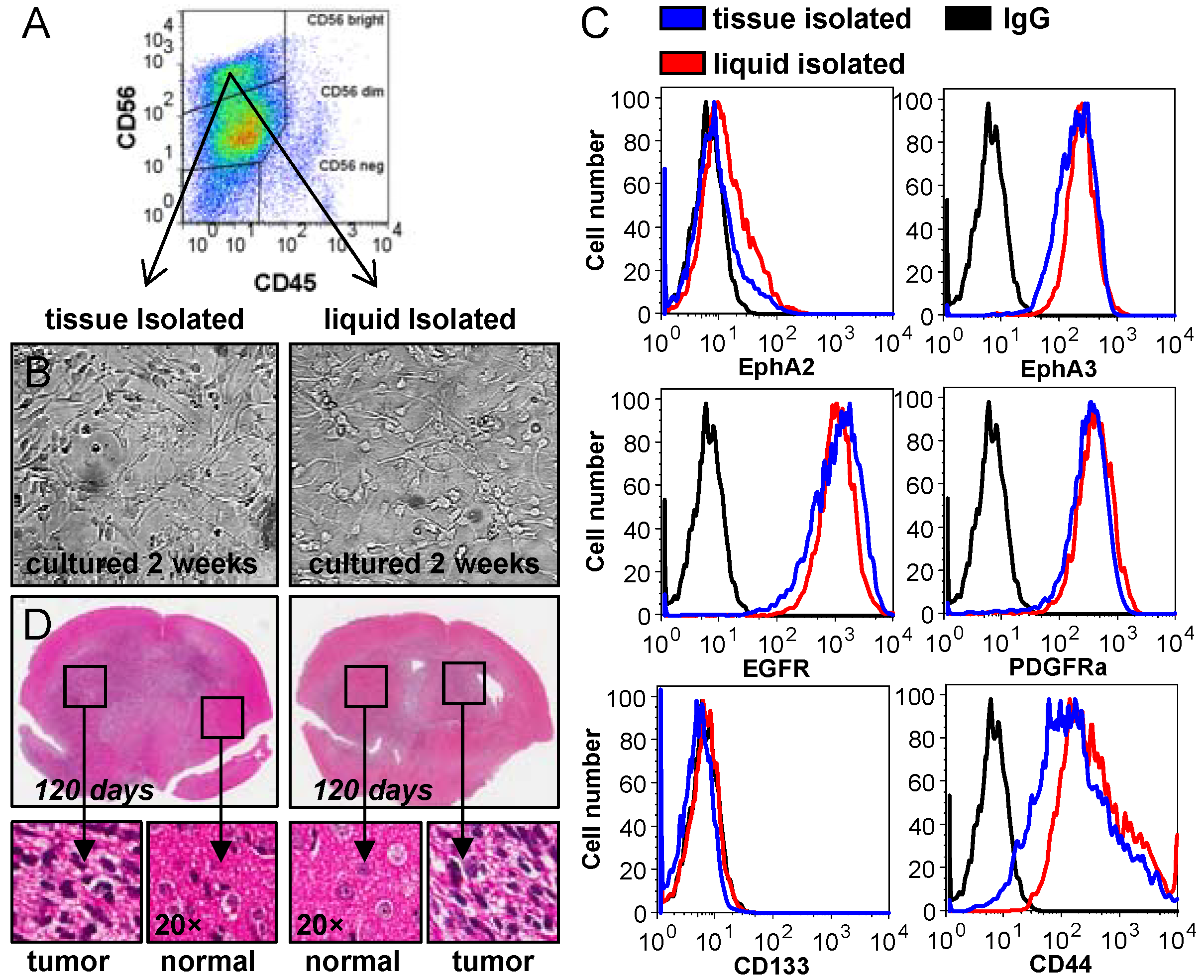

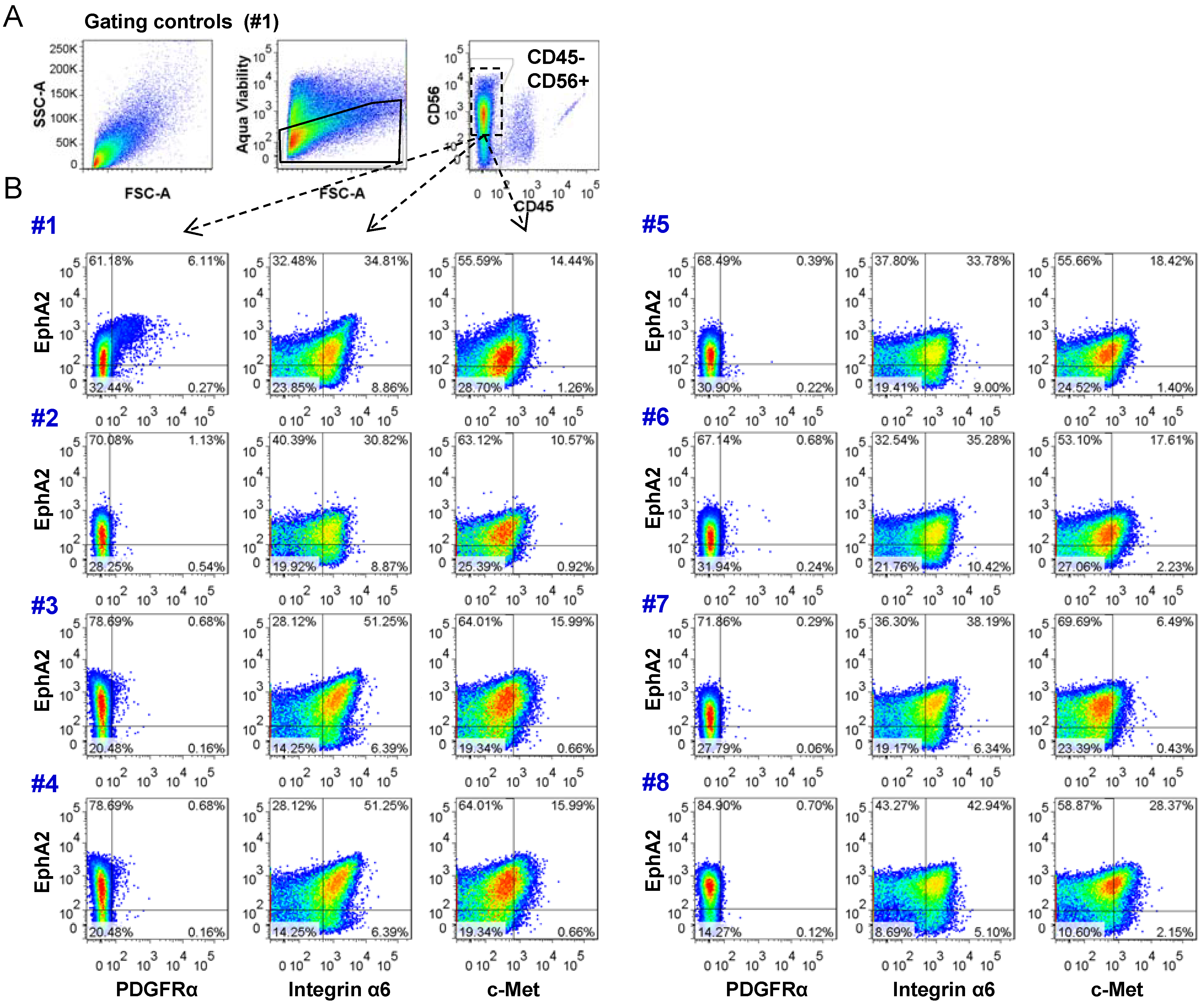
2.3. Analysis of Tumor Heterogeneity Using Multi-Parameter Flow Cytometry
3. Experimental Section
3.1. Ethics Statement
3.2. Patient Specimens

3.3. Cell Culture
3.4. Orthotopic Xenografts
3.5. Relative Quantitation by Real-Time PCR
3.6. Immunofluorescence and IHC
3.7. Flow Cytometric Analysis and Sorting
3.8. Statistical Analysis
4. Conclusions
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Tumor Type | Tumor Location | Age | Sex | Serum Culture | GNS Culture | Tumorigenic in Mice | |
|---|---|---|---|---|---|---|---|---|
| 1 | Normal | Temporal lobe/neocortex | 37 | Male | √ | |||
| 2 | Astrocytoma WHO grade III | Posterior Temporal | 38 | Male | √ | |||
| 3 | Anaplastic ODG WHO grade III | Right frontal | 39 | Male | ||||
| 4 | Diffuse astrocytoma | Left parietal occipital | 19 | Male | √ | √ | ||
| 5 | Diffuse astrocytoma | Left temporal | 38 | Male | ||||
| 6 | GBM | Right frontal | 26 | Male | √ | |||
| 7 | GBM | Left frontal parietal | 32 | Male | √ | √ | ||
| 8 | GBM | Left frontal | 34 | Male | √ | |||
| 9 | GBM | Left frontal parietal | 37 | Male | √ | |||
| 10 | GBM | Right parietal | 37 | Female | √ | √ | ||
| 11 | GBM | Right parietal | 47 | Male | √ | √ | √ | |
| 12 | GBM | Left occipital | 51 | Male | ||||
| 13 | GBM | Left frontal parietal | 51 | Male | √ | √ | √ | |
| 14 | GBM | Right temporal parietal | 51 | Male | √ | |||
| 15 | GBM | Right frontal parietal | 54 | Female | √ | √ | √ | |
| 16 | GBM | Left temporal | 56 | Male | √ | √ | √ | |
| 17 | GBM | Right temporal | 57 | Male | √ | √ | ||
| 18 | GBM | Left temporal | 57 | Female | √ | √ | ||
| 19 | GBM | Right parietal | 58 | Male | √ | |||
| 20 | GBM | Left temporal | 58 | Female | √ | √ | ||
| 21 | GBM | Left frontal | 58 | Female | ||||
| 22 | GBM | Left frontal | 59 | Male | √ | |||
| 23 | GBM | Right temporal | 59 | Male | √ | |||
| 24 | GBM | Right temporal | 61 | Female | √ | √ | ||
| 25 | GBM | Posterior temporal | 61 | Female | √ | |||
| 26 | GBM | Right parietal | 63 | Male | √ | √ | ||
| 27 | GBM | Left temporal | 64 | Female | √ | |||
| 28 | GBM | Left frontal | 65 | Male | √ | |||
| 29 | GBM | - | 65 | Male | √ | |||
| 30 | GBM | Right temporal | 58 | Female | √ | |||
| 31 | GBM | Right temporal | 68 | Male | √ | |||
| 32 | GBM | Right intrinsic | 69 | Male | √ | √ | ||
| 33 | GBM | Left temporal | 69 | Male | ||||
| 34 | GBM | Right frontal | 74 | Male | √ | |||
| 35 | GBM | Parietal | 74 | Female | ||||
| 36 | GBM | Left parietal | 74 | Male | √ | √ | ||
| 37 | GBM | Right frontal | 75 | Male | √ | √ | √ | |
| 38 | GBM | Right frontal | 75 | Female | √ | |||
| 39 | GBM | Left frontal | 84 | Female | √ | √ | ||
| 40 | Oligoastrocytoma WHO grade II | Left fronto-temporal | 46 | Male | √ | |||
| 41 | ODG | Right occipital | 35 | Female | √ | |||
| 42 | ODG | Left frontal | 35 | Male | ||||
| 43 | ODG | Right frontal | 58 | Female | √ | |||
| 44 | Recurrent diffuse glioma WHO grade II | Right frontal parietal | 40 | Female | √ | |||
| 45 | Recurrent GBM | Right parietal | 48 | Male | √ | |||
| 46 | Recurrent GBM | Left parietal | 52 | Male | √ | √ | ||
| 47 | Recurrent GBM | Right parietal | 62 | Male | √ | |||
| 48 | Recurrent GBM | Right parietal | 62 | Male | √ | |||
| Name | Antibody Clone | Origin | Catalogue Number | Fluorochrome |
|---|---|---|---|---|
| CD56 | B159 | BD | 557747 | PE-Cy7 |
| CD45 | 2D1 | BD | 560178 | APC-H7 |
| Aqua Viability Dye | - | Molecular Probes | L34957 | Aqua |
| EphA2 | 1F7 | In-house | N/A | Unconjugated |
| CD140a (PDGFRα) | aR1 | BD | 556002 | PE |
| CD49f (Integrin-α6) | EBioGoH3 | Ebioscience | 46-0495-82 | PerCp-eFluor710 |
| c-Met | LMH85 | In-house | N/A | FITC |
References
- Behin, A.; Hoang-Xuan, K.; Carpentier, A.F.; Delattre, J.Y. Primary brain tumours in adults. Lancet 2003, 361, 323–331. [Google Scholar]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef]
- Finley, J.L.; Silverman, J.F.; Dickens, M.A. Immunocytochemical evaluation of central nervous system tumors obtained by the Cavitron ultrasonic surgical aspirator. Diagn. Cytopathol. 1990, 6, 308–312. [Google Scholar]
- Gomez, G.G.; Kruse, C.A. Isolation and culture of human brain tumor cells. Methods Mol. Med. 2004, 88, 101–109. [Google Scholar]
- Beckner, M.E.; Jane, E.P.; Jankowitz, B.; Agostino, N.R.; Walter, K.A.; Hamilton, R.L.; Pollack, I.F. Tumor cells from ultrasonic aspirations of glioblastomas migrate and form spheres with radial outgrowth. Cancer Lett. 2007, 255, 135–144. [Google Scholar] [CrossRef]
- Oakes, W.J.; Friedman, H.S.; Bigner, S.H.; Bullock, N.H.; Bigner, D.D. Successful laboratory growth and analysis of CUSA-obtained medulloblastoma samples. Technical note. J. Neurosurg. 1990, 72, 821–823. [Google Scholar] [CrossRef]
- Truong, L.N.; Patil, S.; Martin, S.S.; LeBlanc, J.F.; Nanda, A.; Nordberg, M.L.; Beckner, M.E. Rapid detection of high-level oncogene amplifications in ultrasonic surgical aspirations of brain tumors. Diagn Pathol. 2012, 7, 66. [Google Scholar]
- Snuderl, M.; Fazlollahi, L.; Le, L.P.; Nitta, M.; Zhelyazkova, B.H.; Davidson, C.J.; Akhavanfard, S.; Cahill, D.P.; Aldape, K.D.; Betensky, R.A.; et al. Mosaic amplification of multiple receptor tyrosine kinase genes in glioblastoma. Cancer Cell 2011, 20, 810–817. [Google Scholar] [CrossRef]
- Szerlip, N.J.; Pedraza, A.; Chakravarty, D.; Azim, M.; McGuire, J.; Fang, Y.; Ozawa, T.; Holland, E.C.; Huse, J.T.; Jhanwar, S.; et al. Intratumoral heterogeneity of receptor tyrosine kinases EGFR and PDGFRA amplification in glioblastoma defines subpopulations with distinct growth factor response. Proc. Natl. Acad. Sci. USA 2012, 109, 3041–3046. [Google Scholar] [CrossRef]
- Little, S.E.; Popov, S.; Jury, A.; Bax, D.A.; Doey, L.; Al-Sarraj, S.; Jurgensmeier, J.M.; Jones, C. Receptor tyrosine kinase genes amplified in glioblastoma exhibit a mutual exclusivity in variable proportions reflective of individual tumor heterogeneity. Cancer Res. 2012, 72, 1614–1620. [Google Scholar] [CrossRef]
- Pollard, S.M.; Yoshikawa, K.; Clarke, I.D.; Danovi, D.; Stricker, S.; Russell, R.; Bayani, J.; Head, R.; Lee, M.; Bernstein, M.; et al. Glioma stem cell lines expanded in adherent culture have tumor-specific phenotypes and are suitable for chemical and genetic screens. Cell Stem Cell 2009, 4, 568–580. [Google Scholar] [CrossRef]
- Wykosky, J.; Gibo, D.M.; Stanton, C.; Debinski, W. EphA2 as a novel molecular marker and target in glioblastoma multiforme. Mol. Cancer Res. 2005, 3, 541–551. [Google Scholar] [CrossRef]
- Binda, E.; Visioli, A.; Giani, F.; Lamorte, G.; Copetti, M.; Pitter, K.L.; Huse, J.T.; Cajola, L.; Zanetti, N.; Dimeco, F.; et al. The EphA2 receptor drives self-renewal and tumorigenicity in stem-like tumor-propagating cells from human glioblastomas. Cancer Cell 2012, 22, 765–780. [Google Scholar] [CrossRef]
- Day, B.W.; Stringer, B.W.; Al-Ejeh, F.; Ting, M.T.; Wilson, J.; Ensbey, K.S.; Jamieson, P.R.; Bruce, Z.C.; Lim, Y.C.; Offenhauser, C.; et al. EphA3 Maintains Tumorigenicity and is a Therapeutic Target in Glioblastoma Multiforme. Cancer Cell 2013, 23, 238–248. [Google Scholar] [CrossRef]
- Hermanson, M.; Funa, K.; Hartman, M.; Claesson-Welsh, L.; Heldin, C.H.; Westermark, B.; Nister, M. Platelet-derived growth factor and its receptors in human glioma tissue: Expression of messenger RNA and protein suggests the presence of autocrine and paracrine loops. Cancer Res. 1992, 52, 3213–3219. [Google Scholar]
- Lathia, J.D.; Gallagher, J.; Heddleston, J.M.; Wang, J.; Eyler, C.E.; Macswords, J.; Wu, Q.; Vasanji, A.; McLendon, R.E.; Hjelmeland, A.B.; et al. Integrin alpha 6 regulates glioblastoma stem cells. Cell Stem Cell 2010, 6, 421–432. [Google Scholar] [CrossRef]
- Birchmeier, C.; Birchmeier, W.; Gherardi, E.; vande Woude, G.F. Met, metastasis, motility and more. Nat. Rev. Mol. Cell Biol. 2003, 4, 915–925. [Google Scholar] [CrossRef]
- Chen, J.; McKay, R.M.; Parada, L.F. Malignant glioma: Lessons from genomics, mouse models, and stem cells. Cell 2012, 149, 36–47. [Google Scholar] [CrossRef]
- Mesiwala, A.H.; Scampavia, L.D.; Rabinovitch, P.S.; Ruzicka, J.; Rostomily, R.C. On-line flow cytometry for real-time surgical guidance. Neurosurgery 2004, 55, 551–560. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Day, B.W.; Stringer, B.W.; Wilson, J.; Jeffree, R.L.; Jamieson, P.R.; Ensbey, K.S.; Bruce, Z.C.; Inglis, P.; Allan, S.; Winter, C.; et al. Glioma Surgical Aspirate: A Viable Source of Tumor Tissue for Experimental Research. Cancers 2013, 5, 357-371. https://doi.org/10.3390/cancers5020357
Day BW, Stringer BW, Wilson J, Jeffree RL, Jamieson PR, Ensbey KS, Bruce ZC, Inglis P, Allan S, Winter C, et al. Glioma Surgical Aspirate: A Viable Source of Tumor Tissue for Experimental Research. Cancers. 2013; 5(2):357-371. https://doi.org/10.3390/cancers5020357
Chicago/Turabian StyleDay, Bryan W., Brett W. Stringer, John Wilson, Rosalind L. Jeffree, Paul R. Jamieson, Kathleen S. Ensbey, Zara C. Bruce, Po Inglis, Suzanne Allan, Craig Winter, and et al. 2013. "Glioma Surgical Aspirate: A Viable Source of Tumor Tissue for Experimental Research" Cancers 5, no. 2: 357-371. https://doi.org/10.3390/cancers5020357