Epithelial-Mesenchymal Transition Is a Critical Step in Tumorgenesis of Pancreatic Neuroendocrine Tumors

Abstract
:1. Introduction
2. Material and Methods
2.1. Subjects
2.2. Mice
2.3. Genotyping
2.4. Drug Treatment
2.5. Necropsy and Assessment of Islet Cell Tumor Growth
2.6. Immunostaining

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Species | Working dilution | Source |
|---|---|---|---|
| α-Snail | Goat | 1:100 | Santa Cruz, Santa Cruz, CA |
| α -Vimentin | Goat | 1.100 | Santa Cruz, Santa Cruz, CA |
| α -E-cadherin | Rat | 1:200 | Zymed, S.F., CA |
| α -Twist | Goat | 1:250 | Santa Cruz, Santa Cruz, CA |
2.7. RNA Extraction and Real-Time RT-PCR
2.8. Cell Culture
2.9. Protein Analysis
2.10. Cell Adhesion Assay
2.11. Luciferase Reporter Assay
2.12. Statistical Analysis
3. Results
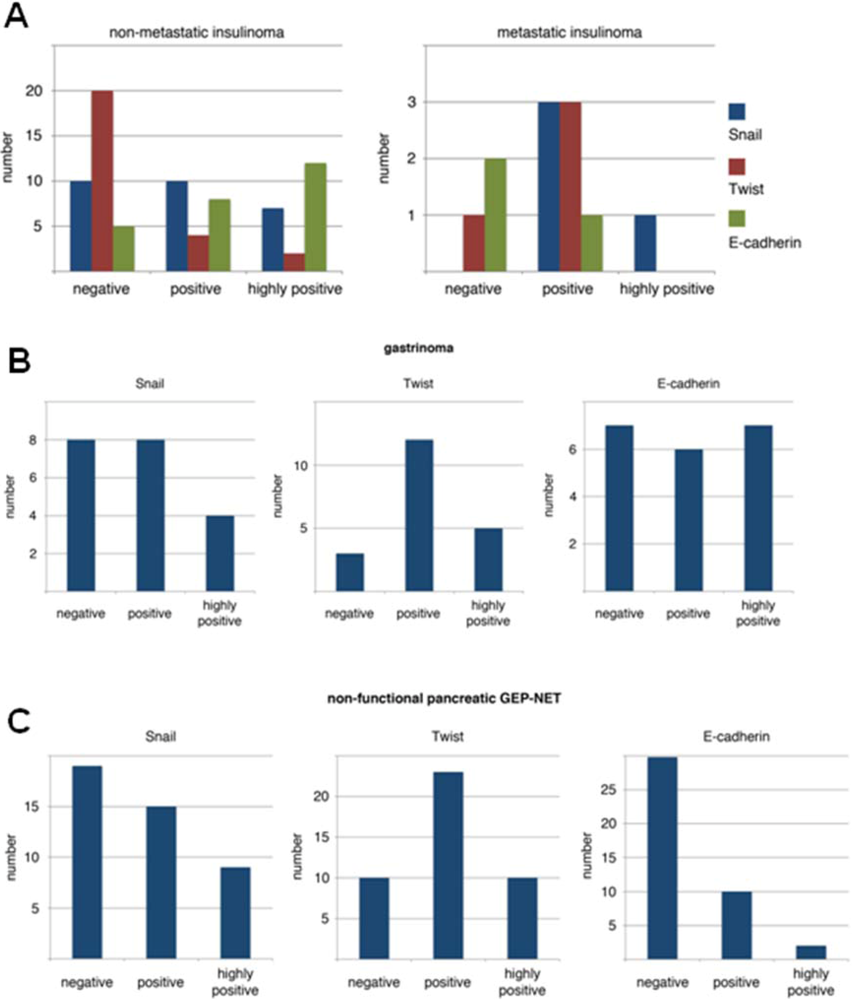
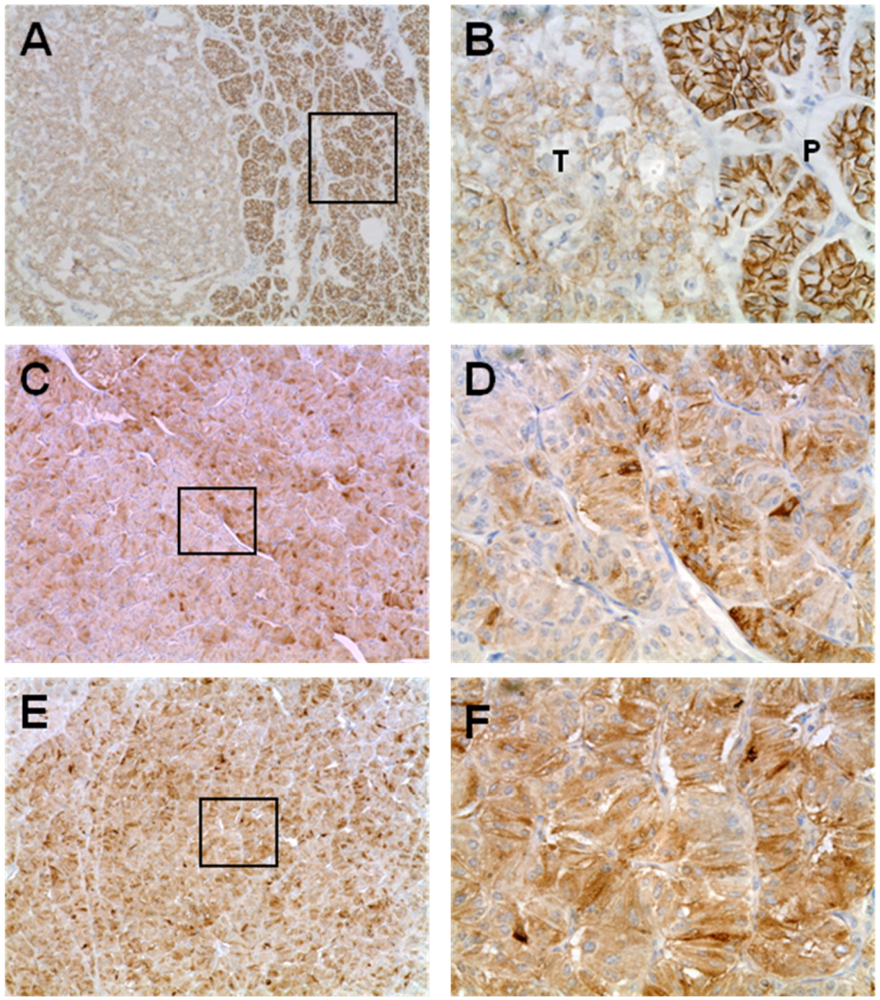
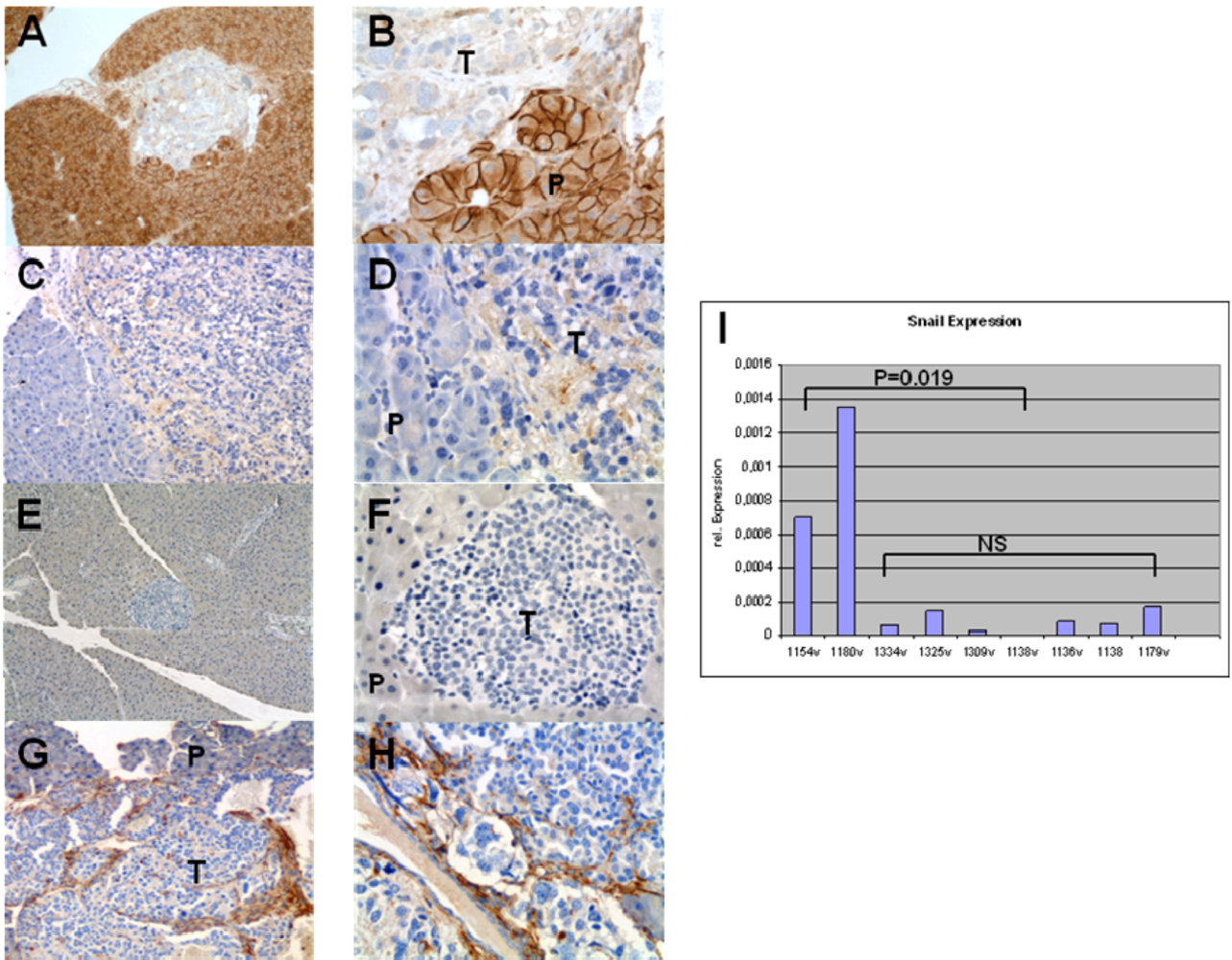
3.1. Expression Pattern of EMT Markers in Human PNETs
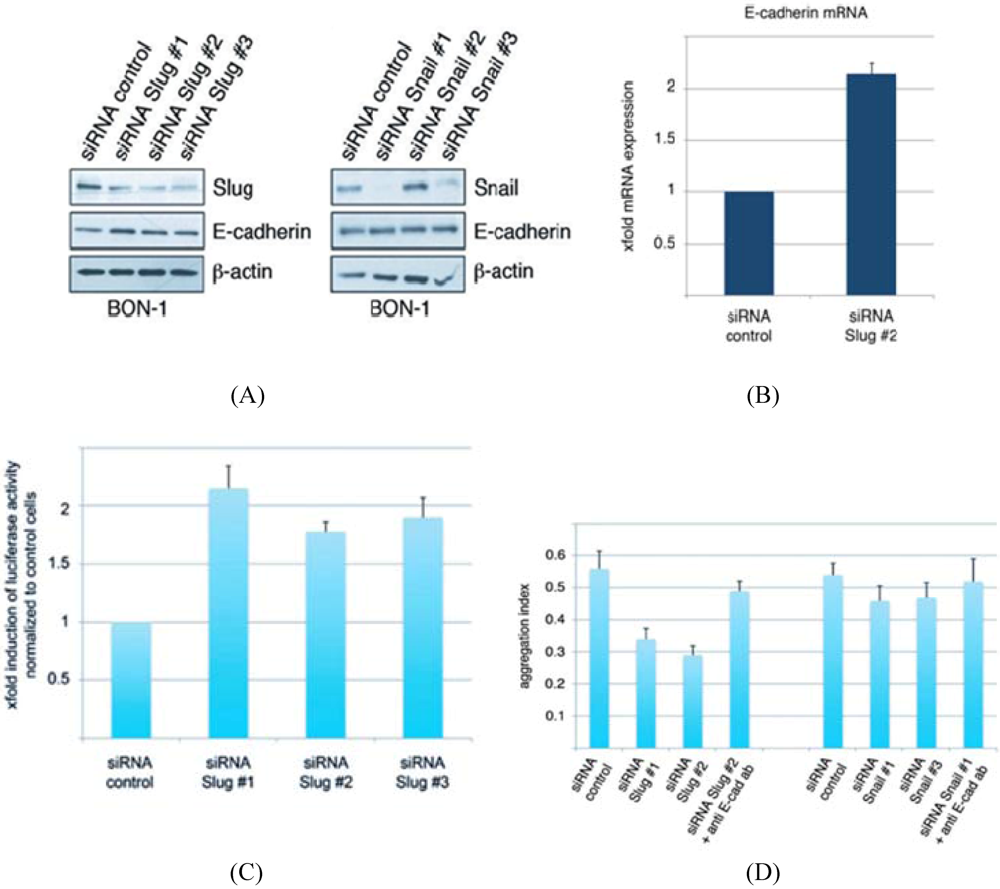
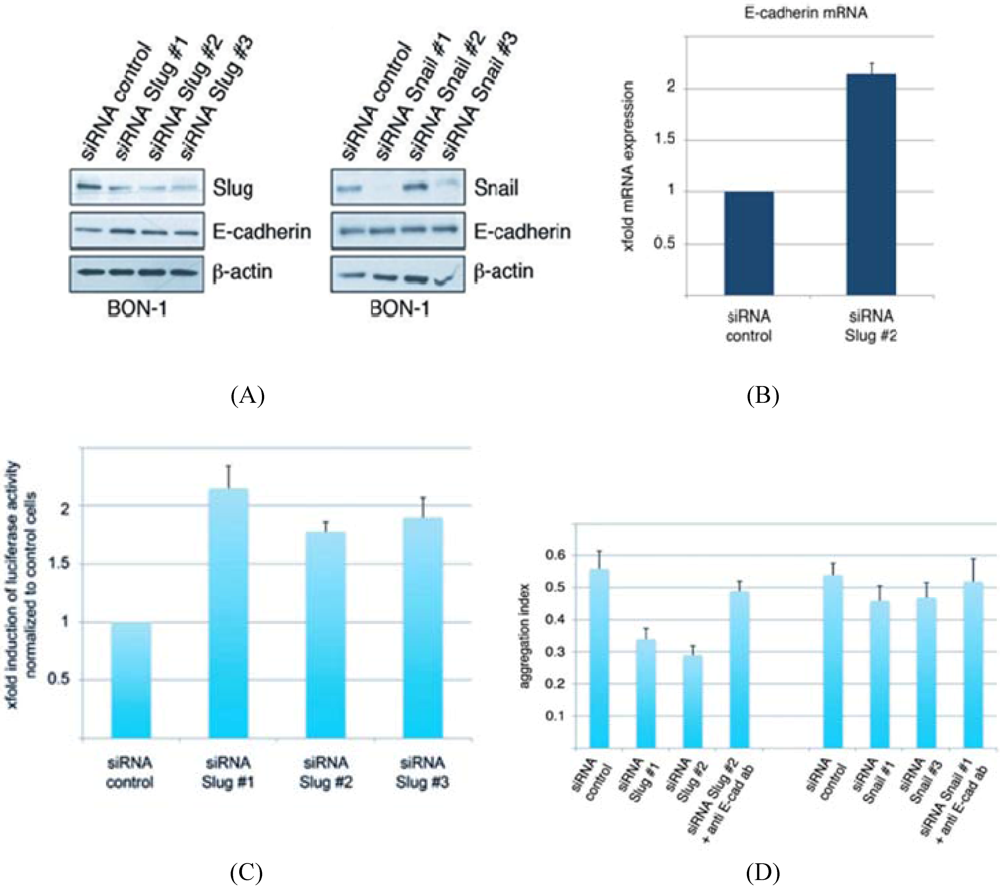
3.2. Conservation of EMT Markers in BON-1 Cells
3.3. Development of Islet Cell Tumors in Rip1Tag2 Mice
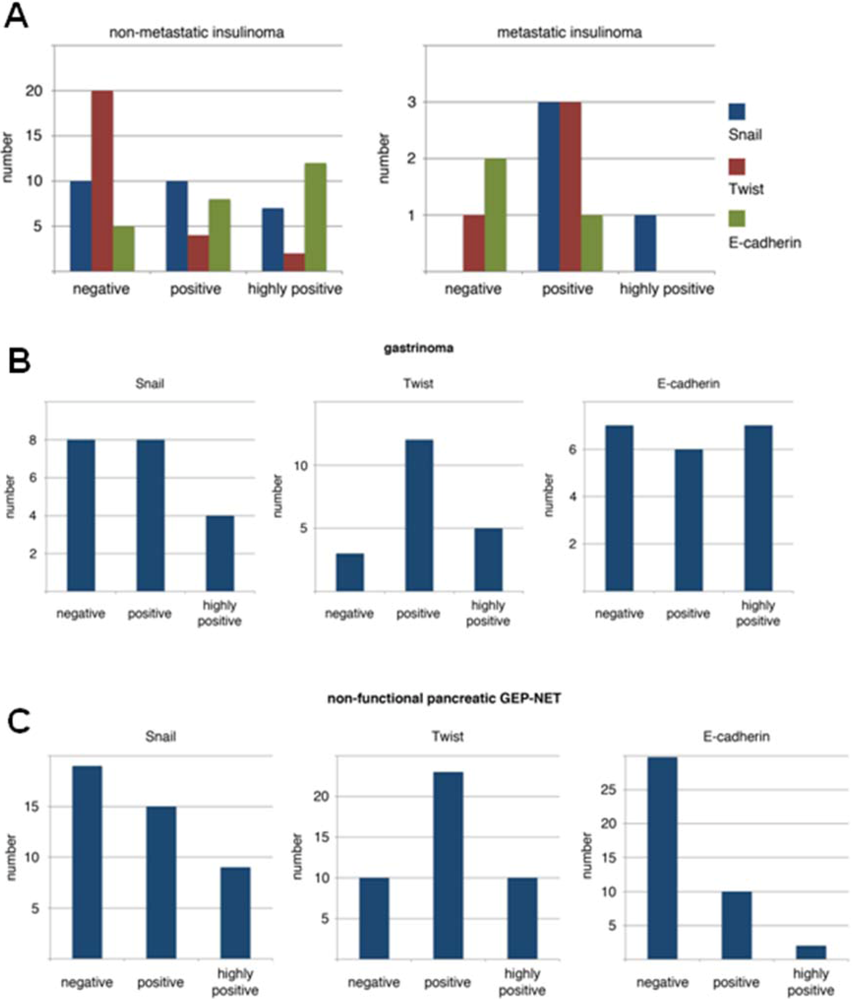
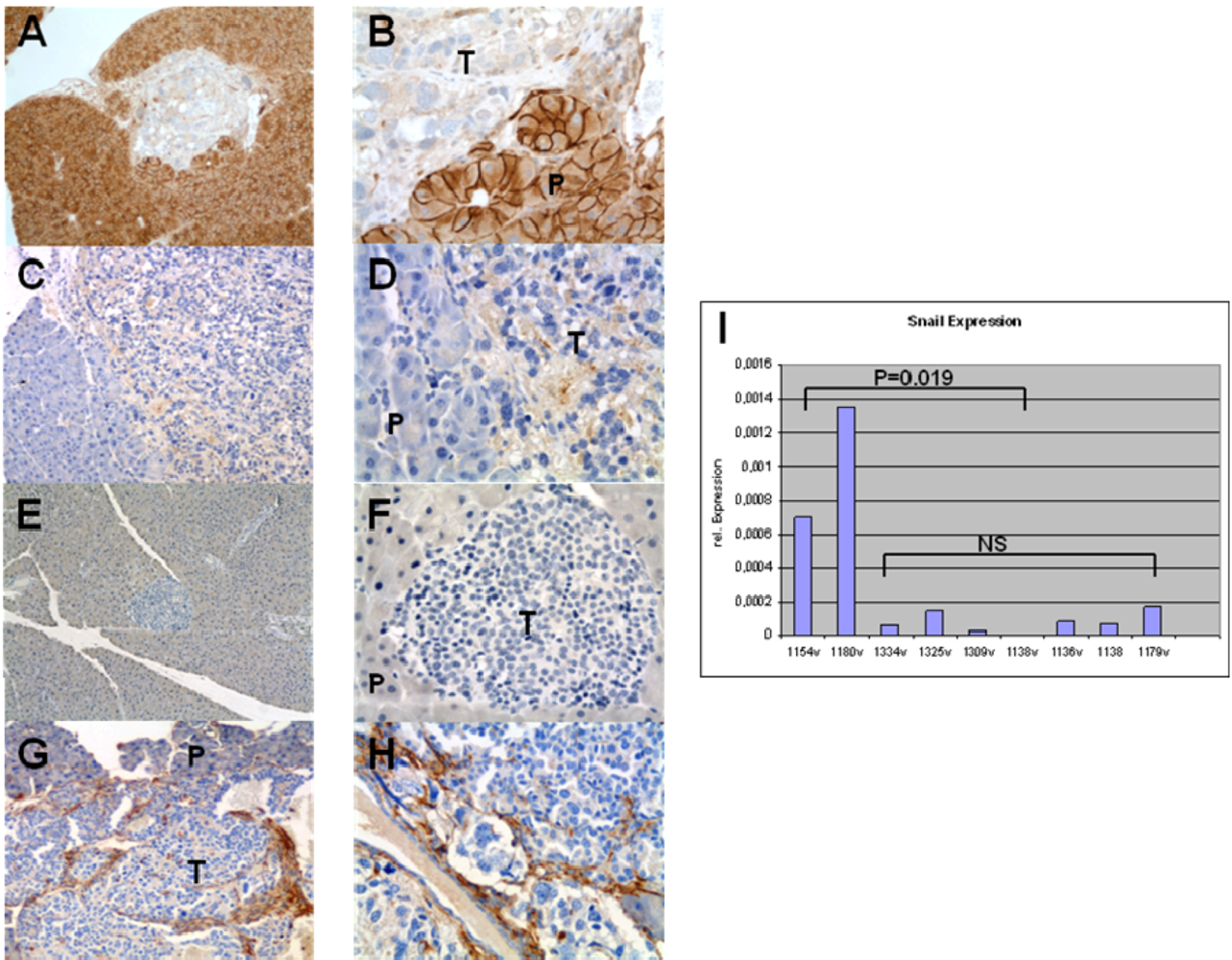
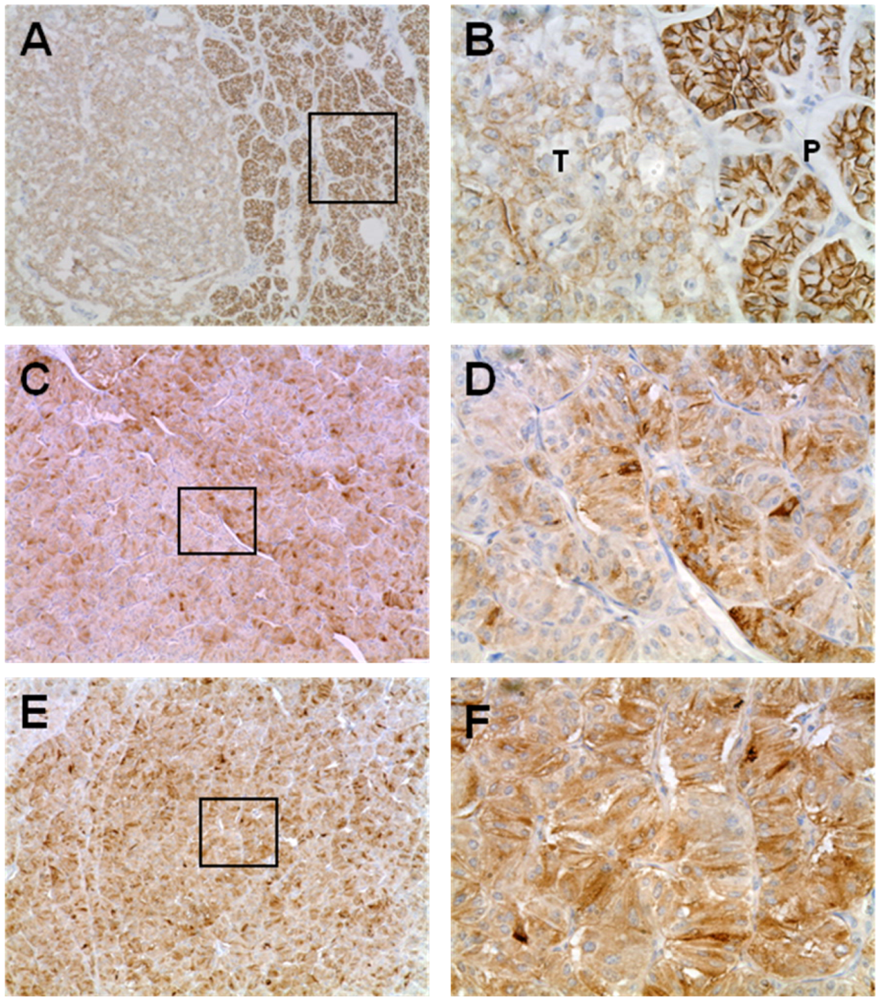
3.4. Expression of EMT Markers in Rip1Tag2 Transgenic Mice
3.5. The Snail Inhibitor PEG Decreases Tumor Growth in Early Stages of Rip1Tag2 Mice
3.6. Snail is Down-Regulated in Islet Cell Tumors by PEG
4. Discussion
Conclusions
Acknowledgements
References
- Öberg, K.; Eriksson, B. Endocrine tumors of the pancreas. Best Pract. Res. Clin. Gastroenterol. 2005, 19, 753–781. [Google Scholar] [CrossRef]
- Metz, D.C.; Jensen, R.T. Gastrointestinal neuroendocrine tumors: Pancreatic endocrine tumors. Gastroenterology 2008, 135, 1469–1492. [Google Scholar] [CrossRef]
- Zhou, B.P.; Hung, M.C. Wnt, hedgehog and snail: Sister pathways that control by GSK-3beta and beta-Trcp in the regulation of metastasis. Cell Cycle 2005, 4, 772–776. [Google Scholar] [CrossRef]
- Thiery, J.P.; Sleeman, J.P. Complex networks orchestrate epithelial-mesenchymal transitions. Nat. Rev. Mol. Cell Biol. 2006, 7, 131–142. [Google Scholar] [CrossRef]
- Christofori, G. New signals from the invasive front. Nature 2006, 44, 444–450. [Google Scholar] [CrossRef]
- Moody, S.E.; Perez, D.; Pan, T.C.; Sarkisian, C.J.; Portocarrero, C.P.; Sterner, C.J.; Notorfrancesco, K.L.; Cardiff, R.D.; Chodosh, L.A. The transcriptional repressor Snail promotes mammary tumor recurrence. Cancer Cell 2005, 8, 197–209. [Google Scholar] [CrossRef]
- Fendrich, V.; Waldmann, J.; Esni, F.; Ramaswamy, A.; Mullendore, M.; Buchholz, M.; Maitra, A.; Feldmann, G. Snail and Sonic Hedgehog activation in neuroendocrine tumors of the ileum. Endocr. Relat. Cancer 2007, 14, 865–874. [Google Scholar] [CrossRef]
- Fendrich, V.; Waldmann, J.; Feldmann, G.; Schlosser, K.; König, A.; Ramaswamy, A.; Bartsch, D.K.; Karakas, E. Unique expression pattern of the EMT markers Snail, Twist and E-cadherin in benign and malignant parathyroid neoplasia. Eur. J. Endocrinol. 2009, 160, 695–703. [Google Scholar]
- Waldmann, J.; Feldmann, G.; Slater, E.P.; Langer, P.; Buchholz, M.; Ramaswamy, A.; Saeger, W.; Rothmund, M.; Fendrich, V. Expression of the zinc finger transcription factor Snail in Adrenocortical Carcinoma is associated with decreased survival. Br. J. Cancer 2008, 99, 1900–1907. [Google Scholar] [CrossRef]
- Waldmann, J.; Slater, E.P.; Langer, P.; Buchholz, M.; Ramaswamy, A.; Walz, M.K.; Schmid, K.W.; Feldmann, G.; Bartsch, D.K.; Fendrich, V. Expression of the transcription factor snail and its target gene twist are associated with malignancy in pheochromocytomas. Ann. Surg. Oncol. 2009, 16, 1997–2005. [Google Scholar] [CrossRef]
- Hanahan, D. Heritable formation of pancreatic beta-cell tumours in transgenic mice expressing recombinant insulin/simian virus 40 oncogene. Nature 1985, 315, 115–122. [Google Scholar] [CrossRef]
- Fendrich, V.; Esni, F.; Garay, M.V.; Feldmann, G.; Habbe, N.; Jensen, J.N.; Dor, Y.; Stoffers, D.; Jensen, J.; Leach, S.D.; Maitra, A. Hedgehog signaling regulates facultative progenitor activity in regenerating exocrine pancreas. Gastroenterology 2008, 135, 621–631. [Google Scholar]
- Parangi, S.; O’Reilly, M.; Christofori, G.; Holmgren, L.; Grosfeld, J.; Folkman, J.; Hanahan, D. Antiangiogenic therapy of transgenic mice impairs de novo tumor growth. Proc. Natl. Acad. Sci. USA 1996, 93, 2002–2007. [Google Scholar]
- Feldmann, G.; Fendrich, V.; McGovern, K.; Bedja, D.; Bisht, S.; Alvarez, H.; Koorstra, J.B.; Habbe, N.; Karikari, C.; Mullendore, M.; et al. An orally bioavailable small-molecule inhibitor of Hedgehog signaling inhibits tumor initiation and metastasis in pancreatic cancer. Mol. Cancer Ther. 2008, 7, 2725–2735. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Vogelmann, R.; Nguyen-Tat, M.D.; Giehl, K.; Adler, G.; Wedlich, D.; Menke, A. TGFbeta-induced downregulation of E-cadherin-based cell-cell adhesion depends on PI3-kinase and PTEN. J. Cell Sci. 2005, 118, 4901–4912. [Google Scholar]
- Wali, R.K.; Kunte, D.P.; Koetsier, J.L.; Bissonnette, M.; Roy, H.K. Polyethylene glycol-mediated colorectal cancer chemoprevention: Roles of epidermal growth factor receptor and Snail. Mol. Cancer Ther. 2008, 7, 3103–3111. [Google Scholar] [CrossRef]
- Gotzmann, J.; Mikula, M.; Eger, A.; Schulte-Hermann, R.; Foisner, R.; Beug, H.; Mikulits, W. Molecular aspects of epithelial cell plasticity: Implications for local tumor invasion and metastasis. Mutat. Res. 2004, 566, 9–20. [Google Scholar] [CrossRef]
- Jiao, W.; Miyazaki, K.; Kitajima, Y. Inverse correlation between E-cadherin and Snail expression in hepatocellular carcinoma cell lines in vitro and in vivo. Br. J. Cancer 2002, 86, 98–101. [Google Scholar] [CrossRef]
- Rosivatz, E.; Becker, K.F.; Kremmer, E.; Schott, C.; Blechschmidt, K.; Hofler, H.; Sarbia, M. Expression and nuclear localization of Snail, an E-cadherin repressor, in adenocarcinomas of the upper gastrointestinal trac. Virchows Arch. 2006, 448, 277–287. [Google Scholar] [CrossRef]
- Roy, H.K.; Smyrk, T.C.; Koetsier, J.; Victor, T.A.; Wali, R.K. The transcriptional repressor SNAIL is overexpressed in human colon cancer. Dig. Dis. Sci. 2005, 50, 42–46. [Google Scholar] [CrossRef]
- Rosenau, J.; Bahr, M.J.; von Wasielewski, R.; Mengel, M.; Schmidt, H.H.; Nashan, B.; Lang, H.; Klempnauer, J.; Manns, M.P.; Boeker, K.H. Ki67, E-cadherin, and p53 as prognostic indicators of longterm outcome after liver transplantation for metastatic neuroendocrine tumour. Transplantation 2002, 73, 386–394. [Google Scholar]
- Fendrich, V.; Langer, P.; Celik, I.; Bartsch, D.K.; Zielke, A.; Ramaswamy, A.; Rothmund, M. An aggressive surgical approach leads to long- term survival in patients with pancreatic endocrine tumors. Ann. Surg. 2006, 244, 845–851. [Google Scholar] [CrossRef]
- Folkman, J.; Watson, K.; Ingber, D.; Hanahan, D. Induction of angiogenesis during the transition from hyperplasia to neoplasia. Nature 1989, 339, 58–61. [Google Scholar]
- Inoue, M.; Hager, J.H.; Ferrara, N.; Gerber, H.P.; Hanahan, D. VEGF-A has a critical, nonredundant role in angiogenic switching and pancreatic beta cell carcinogenesis. Cancer Cell 2002, 1, 193–202. [Google Scholar] [CrossRef]
- Bergmann, F.; Breinig, M.; Höpfner, M.; Rieker, R.J.; Fischer, L.; Köhler, C.; Esposito, I.; Kleeff, J.; Herpel, E.; Ehemann, V.; et al. Expression pattern and functional relevance of epidermal growth factor receptor and cyclooxygenase-2: Novel chemotherapeutic targets in pancreatic endocrine tumors? Am. J. Gastroenterol. 2009, 104, 171–181. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fendrich, V.; Maschuw, K.; Waldmann, J.; Buchholz, M.; Rehm, J.; Gress, T.M.; Bartsch, D.K.; König, A. Epithelial-Mesenchymal Transition Is a Critical Step in Tumorgenesis of Pancreatic Neuroendocrine Tumors. Cancers 2012, 4, 281-294. https://doi.org/10.3390/cancers4010281
Fendrich V, Maschuw K, Waldmann J, Buchholz M, Rehm J, Gress TM, Bartsch DK, König A. Epithelial-Mesenchymal Transition Is a Critical Step in Tumorgenesis of Pancreatic Neuroendocrine Tumors. Cancers. 2012; 4(1):281-294. https://doi.org/10.3390/cancers4010281
Chicago/Turabian StyleFendrich, Volker, Katja Maschuw, Jens Waldmann, Malte Buchholz, Johannes Rehm, Thomas M. Gress, Detlef K. Bartsch, and Alexander König. 2012. "Epithelial-Mesenchymal Transition Is a Critical Step in Tumorgenesis of Pancreatic Neuroendocrine Tumors" Cancers 4, no. 1: 281-294. https://doi.org/10.3390/cancers4010281