1. Introduction
Uveal melanoma (UM) constitutes about 5% of all melanomas (based on Aronow et al. [
1] and current Surveillance, Epidemiology, and End Results Program (SEER) data (
https://seer.cancer.gov/)), and it is the most prevalent primary intraocular cancer in adults, with an annual incidence of 4–7 per million [
1,
2]. Overall, metastases appear in up to 50% of patients [
3]. There is currently no effective treatment for the metastases, and most patients die within two years (80% in the first year and 92% in the second) [
4]. Both primary and metastatic UM are relatively resistant to systemic chemotherapy and radiotherapy with only a modest efficacy [
5,
6]. Therefore, novel therapeutic approaches are required.
A variety of studies established that tumor cells are more resistant to hypoxia than non-malignant cells and they can remain viable in hypoxic environments [
7]. Moreover, experimental and clinical data have shown that tumor hypoxia is associated with tumor progression to a more malignant phenotype and with the spread of metastases, altogether resulting in poor survival in patients with various solid tumors [
7,
8]. Additionally, tumor hypoxia has been considered as a potential therapeutic problem, both because there is insufficient diffusion of chemotherapy in the hypoxic region and because radiation will not produce its effector—the free oxygen radical—without oxygen [
9]. On the other hand, the on-going development of hypoxic regions in growing tumors provides an opportunity for tumor-selective therapies.
To overcome the problem of these hypoxic treatment-resistant regions, we constructed MuLV-based replication competent retroviral (RCR) vectors [
10] expressing shRNA targeting the hypoxia-response regulating genes.
The adaptive response to hypoxia is orchestrated by a family of transcription factors (TFs), such as the hypoxia-inducible factors-1 (HIF-1) and -2 (HIF-2) [
11], cyclic AMP-response-element (CRE) binding protein (CREB) [
8], and the activator protein-1 (AP-1) [
9].
HIF-1 is a heterodimer that is composed of an oxygen-regulated 1α subunit and a constitutively expressed 1β subunit. Non-hydroxylated active HIF-1 regulates the expression of hundreds of genes that are implicated in events, such as angiogenesis, cell survival/death, metabolism, migration, and metastasis. The genes that are involved in these processes are targeted by HIF-1, which binds to the hypoxia response elements (HRE). In the presence of oxygen, HIF-1 and HIF-2 are subjected to hydroxylation by prolyl-4-hydroxylase domain proteins. Under hypoxic conditions, the rate of hydroxylation declines such that non-hydroxylated proteins accumulate (for review, see Majmundar et al. [
11]).
The HIF-1α and -2α subunits differ in their transactivation domains, which implies that they have unique target genes, although HIF-1α and -2α are highly conserved at the protein level, have similar domain structure, dimerize with HIF-1β, and activate HRE-dependent gene expression [
11]. Indeed, it was shown that HIF-1 and HIF-2 have distinct, tissue specific expression patterns. HIF-1α is ubiquitously expressed, whereas HIF-2α expression is limited to the endothelium, kidneys, lungs, heart, and small intestine. HIF-1α has been shown to have a role in UM tumor progression and metastases formation [
12,
13].
Various mediators activate CREB through the phosphorylation of serine 133 (Ser133), which leads to the activation of CRE mediated transcription. CREB overexpression relative to adjacent normal tissues was found in many solid tumor types, such as non-small lung carcinoma (NSCLC), glioblastoma, mammary carcinoma, hepatocellular carcinoma (HCC), and melanoma [
8,
14,
15]. This could be influenced by the modulation of signaling cascades upstream of CREB as well as the induction of CRE-dependent gene expression downstream of CREB, which leads to increased tumor growth, prevention of cell death, and enhanced metastasis formation and angiogenesis [
8,
12,
13]. Furthermore, the overexpression of CREB in tumors often correlates with disease progression, poor patient survival, and chemotherapy resistance. We have previously demonstrated that the overexpression of a dominant-positive, hypoxia-insensitive CREB300/310 mutant enhanced tumor growth, vascularization, and abrogated hypoxia-induced apoptosis, while the dominant-negative form of CREB (KCREB) or knock-down of CREB abolished mouse hepatocellular carcinoma (HCC) tumor growth [
8,
14].
Similarly, we found that the infection of UM cells (both primary and metastatic cell lines) with a MuLV-based RCR that expresses shRNA targeting CREB (vACE-CREB) [
14] reduces their survival in normoxic conditions by 50% [
16]. The UM cells were found to have an increased apoptotic fraction in hypoxia that did not involve Caspase 3 activation and did not result in increased cell mortality. CREB knockdown increased UM cells’ sensitivity to chemotherapy (decarbazin—DTIC and doxorubicin—DOX), as in the HCC cells, an effect that was stronger in hypoxia [
16].
The advantage of using this vector is that MuLV retroviruses only infect replicating cells, such as tumor cells, which generates a stably integrated provirus in the cells. These cells produce viral particles that express shRNA targeting CREB or HIF-1 that will spread within the tumor [
14]. A point mutation in the target sequence against which we design shRNAs can result in tumor resistance to the treatment. To diminish resistance to the siRNA, we constructed RCR vectors expressing shRNA targeting two sites in each gene. The poly(A) signal is a conserved site in genes and less likely to be mutated. We have previously found that the poly(A) signal and the flanking nucleotides (−10 AAUAAA +10) are unique for each gene in the human genome (unpublished data,
http://bioinfo.ekmd.huji.ac.il/honigman). Targeting siRNA to this region results in a very efficient knockdown. Based on these results, we constructed MuLV vectors expressing shRNA targeting the poly(A) signal, in addition to knocking down CREB via a sequence from exon 8 or targeting the poly(A) signal of HIF-1 in addition to targeting the exon 5 sequence.
In the present work, we aimed to test whether knockdown of either CREB or HIF-1 in UM mouse xenograft models will be as effective at attenuating UM tumor growth in vivo as is the knockdown of these genes at reducing UM cell survival in vitro and in HCC growth in vitro and in vivo. In this work, we demonstrated that both regulators of the cellular response to hypoxia play a pivotal role in UM tumor progression and that CREB may play a more important role than HIF-1 in UM tumor progression.
3. Discussion
Ongoing generation of hypoxic regions is a property of solid tumors. The cellular response to hypoxia is orchestrated by several transcription factors including the HIF family and CREB. In a previous work, we demonstrated in vitro that knockdown of CREB in hypoxia, following infection with vACE-CREB, increased apoptosis by 50% and decreased the expression of CREB-regulated genes such VEGF in two UM cell lines Mel270 and OMM2.5 [
16]. In this work we demonstrate for the first-time inhibition of uveal melanoma growth in a mouse model, following knockdown of either CREB or HIF-1 by a replication competent retrovirus vector. This vector can integrate stably and spread within the infected tumor.
To diminish resistance to the knockdown by shRNA expressed by the armed MuLV vectors we added to each vector another shRNA targeting the poly(A) site of each gene. Thus, each vector expressed two shRNAs from the same RNA molecule targeting two sites in each gene. As demonstrated in
Figure 3 and
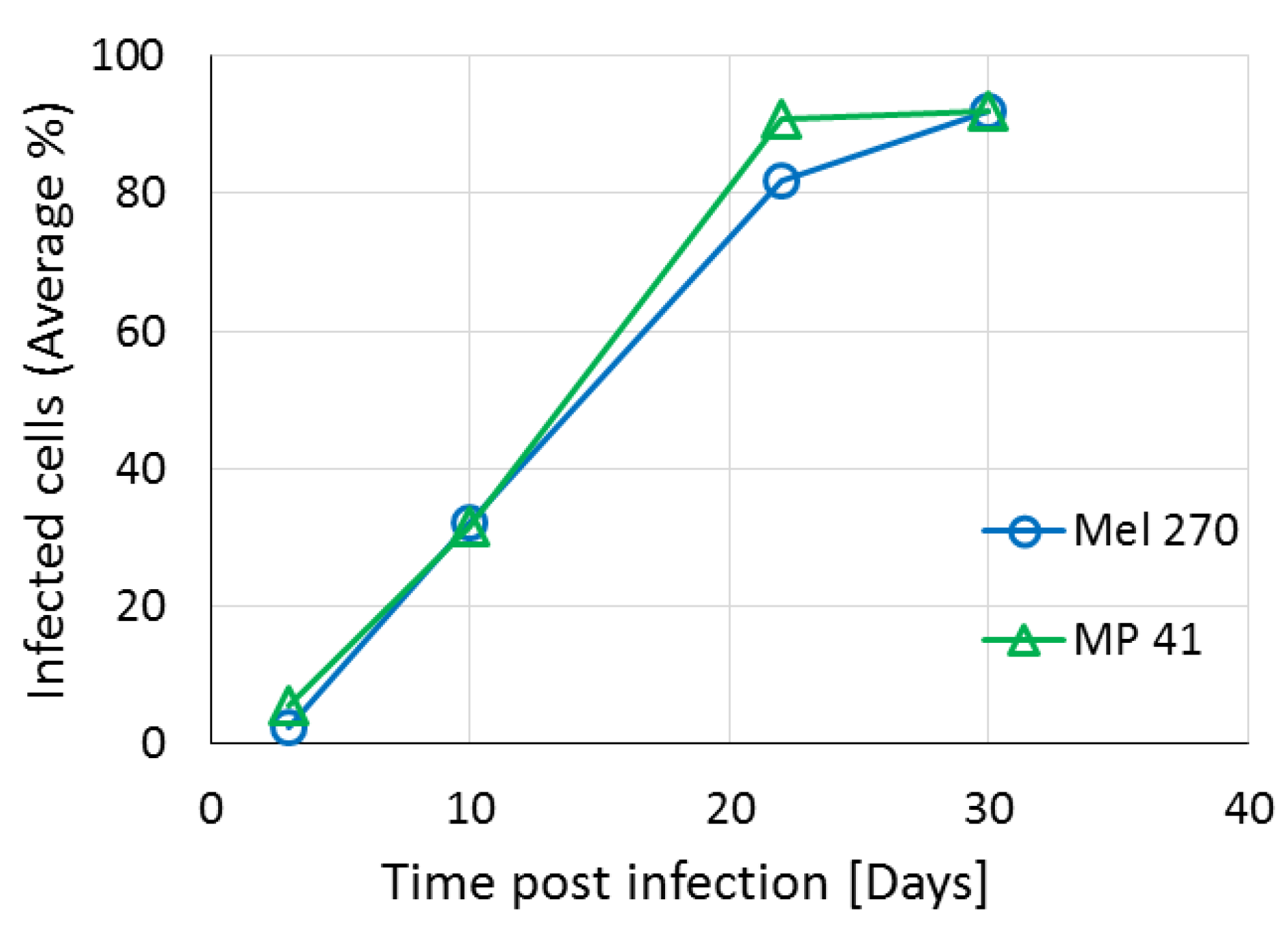
Figure 5, these recombinant vectors efficiently knocked down both CREB and HIF1 in vitro and abolished tumor growth in vivo. There is a debate in the literature regarding the relevance of long-standing cell lines to the primary tumors from patients. To address this concern, we compared in vitro virus infectivity and spread and knockdown of the target genes in three different UM cell lines: Mel270, OMM2.5 and the recently prepared MP41 cell line which has been shown to have the same gene expression profile as the tumor from which it originated from [
20]. In the in vitro experiments, we did not find significant differences in the response of the cells to the infection with the armed viruses.
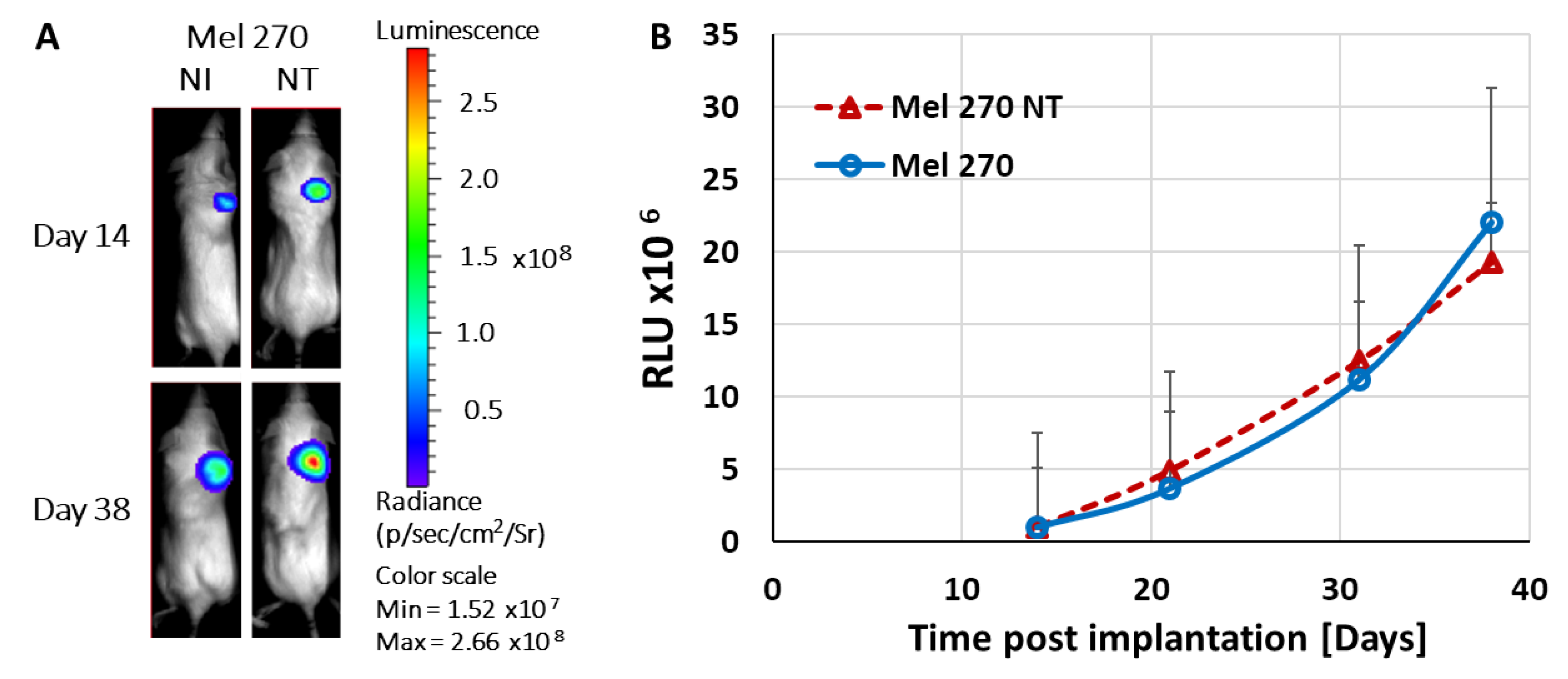
Tumor progression of subcutaneously implanted xenografts stably expressing shRNA targeting either CREB or HIF-1 was diminished as determined by both non-invasive luciferase-based imaging and measurements of the excised tumors (
Figure 5 and
Figure 6). These results suggest that even partial knockdown (about 50% in vitro) of either one of these transcription factors had a major effect on tumor progression. Moreover, we have previously shown in hepatocellular carcinoma that knockdown of either CREB or HIF-1 reduced the expression of VEGF within the tumors which resulted in a decrease in hypoxia-guided neovascularization [
14]. The abrogation of VEGF may be responsible for the stronger effect in UM seen in vivo in the current study compared to the in vitro results in our previous study [
16].
UM is well known for its poor internal vasculature, which led to the discovery of vasculogenic mimicry [
21]. We now know that tumor cells transform into endothelial-like cells and form channels that enable plasma and nutrients to flow throughout the tumors [
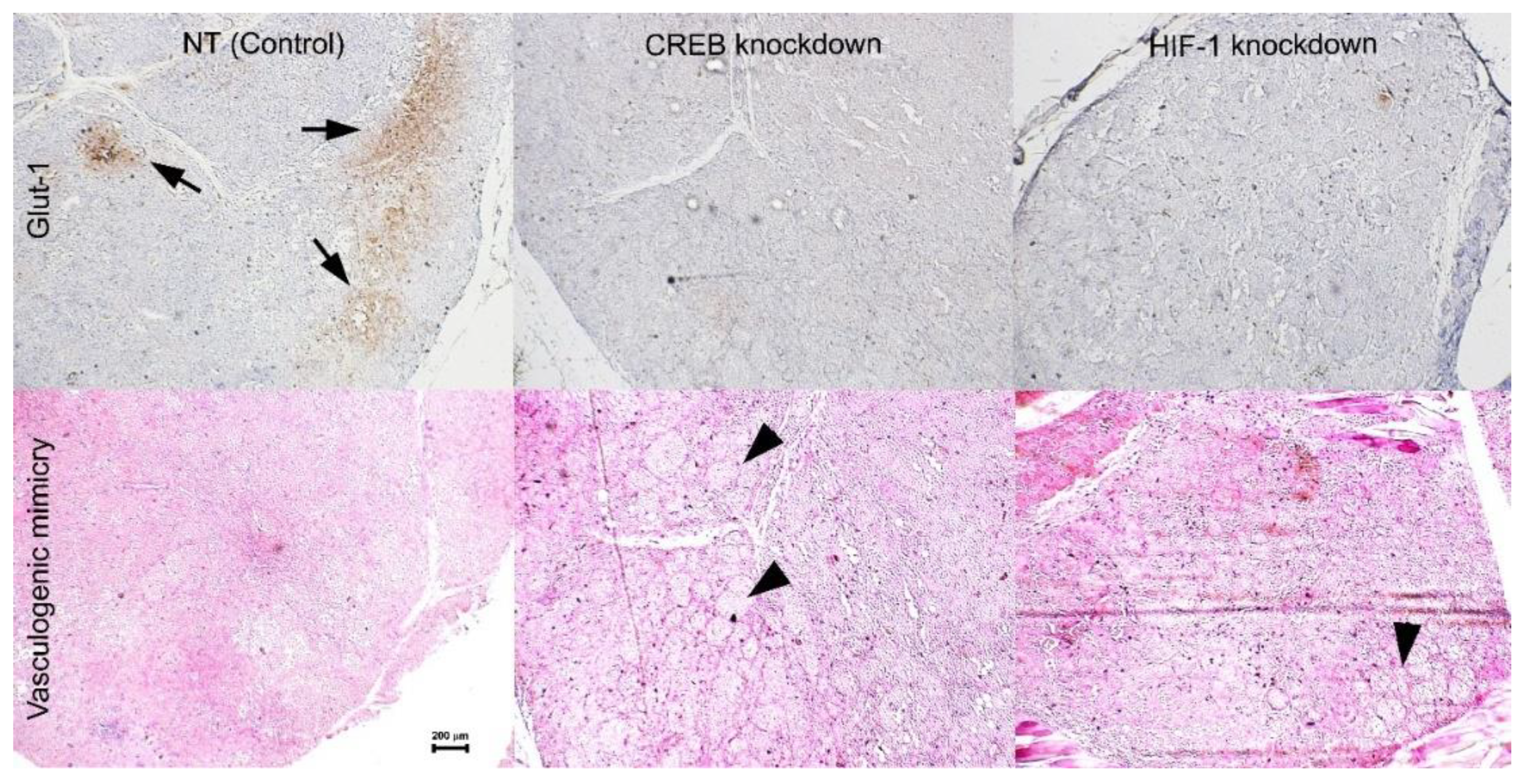
17]. The histopathologic analysis in the current study indicates that formation of vasculogenic mimicry may be inversely correlated with the activity of CREB (
Figure 7).
Tumors infected with the control vector, grew rapidly with the expected formation of hypoxic areas within the tumors where Glut-1 was expressed (
Figure 7). However, when the expression of either CREB or HIF-1 was knocked down, the expression of their downstream gene
glut-1 was abolished. Glut-1 is the major glucose transporter, especially in tumor cells and is important in providing glucose for energy production within these cells [
22]. Activation of Hexokinase II is the initial and rate limiting step in glycolysis. Liu and colleagues found that Hexokinase II is activated by CREB in UM cells. In addition, downregulation of Hexokinase II resulted in reduced glycolysis, increased oxygen consumption and reduced tumor growth [
23]. In vitro, UM cells have sufficient supply of glucose in the growth medium, even in hypoxic conditions, which enables them to survive hypoxia. However, in vivo, glucose is not readily available for the tumor cells. Following the knockdown of either CREB or HIF-1, there was a reduction in Glut-1 expression resulting in limited energy supply preventing UM tumors proliferation in hypoxia. This could be at least part of the reason these tumors hardly grew in vivo (
Figure 5,
Figure 6 and
Figure 7). Taken together with our previous results [
16], it appears that the role played by CREB and HIF-1 in UM is more important in vivo than in vitro.
In cutaneous melanoma there have been publications linking the level of pigmentation to the virulence of the tumor and to HIF-1 activation [
24,
25]. Uveal melanoma is vastly different to cutaneous melanoma in both genetic profile and clinical behavior. Uveal melanoma cell lines do not produce pigment in vitro precluding a proper analysis of the role of melanin. There are no studies correlation the level of pigmentation and HIF-1 in UM cells and their virulence. One study correlated pigmentation with survival of UM patient [
26]. In that study, although there was no difference in prognosis between races, and despite a similar distribution of pigmented vs. non-pigmented tumors clinically, the authors found that in Caucasians more pigmented tumors correlated with a higher metastatic rate. Of note, it is well known that when eyes with uveal melanomas are enucleated, a pigmented surface that would give a higher pigmentation score clinically, can sometimes cover an internal amelanotic tumor. In this work we demonstrate that HIF-1 plays a role in UM tumor progression in vivo.
Our previous results [
16], demonstrated that knockdown of CREB resulted in increased sensitivity to chemotherapeutic agents (doxorubicin and decarbazin). This, together with our findings in the present study, suggest that a combined treatment with the armed viruses and chemotherapy should result in a more potent effect in the reduction of tumor progression.
4. Materials and Methods
4.1. Cell Culture
The human Mel270 cell line [
27] (verified by STR analysis and a kind gift from Prof. Sarah Coupland, Liverpool, UK) and MP-41 [
20] were grown in RPMI 1640 supplemented with 10% fetal bovine serum, 2 mM glutamine, 100 IU/mL penicillin, 100 μg/mL streptomycin (Biological Industries, Kibbutz Beit-Haemek, 25115, Israel) and incubated at 37 °C in a humidified atmosphere with 5% CO
2.
4.2. Plasmids and Viruses
The plasmid pACE-GFP (a kind gift from Prof. Noriyuki Kasahara, Los Angeles, California [
10]) contains a full-length replication-competent amphotropic MuLV provirus with an additional internal ribosome entry site (IRES)-
GFP cassette flanked by BsiWI and NotI restriction enzymes sites. This cassette was replaced by oligonucleotides harboring the H1 promoter driving the transcription of the following shRNA sequences:
5′_GAGAGAGGTCCGTCTAATGTTCAAGAGACATTAGACGGACCTCTCTCTTTTT (pACE-CREB)
5′_GAGAGAGGTCCGTCTAATGTTCAAGAGACATTAGACGGACCTCTCTCTTTTTCAACCTGAAAGACAAAATAAAC (pACE-CREB dbl)
5′_CTAACTGGACACAGTGTGTTTAATATATGAAAACACACTGTGTCCAGTTAGTTTTTT (pACE-HIF-1)
5′_CTAACTGGACACAGTGTGTTTAATATATGAAAACACACTGTGTCCAGTTAGTTTTTTCATCAAATAAACATCTTCTGTG (pACE-HIF-1 dbl)
5′_ACCAAGATGAAGAGCACCAACCTGAACCATTGGTGCTCTTCATCTTGGTTTTTTT (pACE-NT, non-target shRNA)
See
Figure 1 for a schematic presentation of the vectors.
4.3. Virus Preparation
HEK293T cells were transiently transfected with either one of the pACE plasmids, described above and in
Figure 1, using FuGENE HD reagent (Promega-Corp, Madison, WI, USA). The medium containing the virus particles was harvested 48 h later, filtered (MILLEX-HV, PVDF 0.45 µ) and stored at −80 °C.
4.4. Quantitative Real-Time PCR
RNA was extracted from the cells using the NucleoSpin® RNA (Macherey-Nagel, Bethlehem, PA, USA), according to the manufacturer’s instructions. The purified RNA samples were subjected to reverse transcription using GoScript (Promega-Corp, Madison, WI, USA), monitored by quantitative 7900HT real-time PCR apparatus (Applied Biosystems, Foster City, CA, United States) utilizing the GoTaq Real-Time PCR reagents (Promega-Corp, Madison, WI, USA) and the following specific primers:
CREB:
fp-5′_CCCAGCACTTCCTACACAGCCTGC_3′,
rp-5′_ CGAGCTGCTTCCCTGTTCTTCATTAGACG _3′.
HIF-1:
fp-5′_GGGATTAACTCAGTTTGAACTAACTGG_3′,
rp-5′_CCTTTTTCACAAGGCCATTTCTGTGTG_3′.
The results were normalized to the cellular house-keeping gene.
β-actin:
fp-5′_CCTTCCTGGGCATGGAGTCC_3′,
rp-5′_GTGTTGGCGTACAGGTCTTTGC_3′.
4.5. Flow Cytometry (FACS)
Cells infected with vACE-GFP were harvested every seven days, centrifuged (4 °C) at 1750 rpm for 4 min. The supernatant was removed and the cells were re-suspended in PBS +1% FCS, then filtered through a Mesh filter and run on the FACS machine (BD FACScan analyzer, 3A International Business Park, Singapore). Results were analyzed using FCS Express 4 Flow Research Edition (De Novo Software, Glendale, CA, USA, Version 4.07.0020).
4.6. Xenograft Mouse Model
Mel270 cells stably expressing
luc and infected with one of the RCR recombinant viruses (
Figure 1) were harvested and 4 × 10
6 cells in 100 μL were injected subcutaneously (SC) above the foreleg of severe combined immunodeficiency (SCID) mice (C.B-17/ICRHSD-PRKDC-SCID) about three weeks of age (weighing 20–24 gr). To allow descriptive statistics, each group included five mice. Male and female mice were randomly assigned to each treatment group. Each group included a similar distribution of sexes. Tumor growth was monitored weekly following IP injection of D-Luciferin 300 mg/0.1 cc (PBS)/mouse (Promega-Corp, Madison, WI, USA) 10 min. before imaging. This non-invasive luciferase-based method for monitoring tumor growth allows following the same tumor in the same mouse over time and minimizes the number of mice used in the experiment. Mice were anesthetized with isoflurane and bioluminescence was measured with the IVIS In Vivo System (Caliper Life Sciences, Wlatham, MA, USA). No animals were excluded from the analysis, and there was no blinding during the experiments. The IACUC of the Hebrew University of Jerusalem approved the animal protocol used in this study (NIH approval number OPRR-A01-5011 from 29 June 2016).
4.7. Immunohistochemistry
At the end of the experiments, the tumors were excised, photographed, measured, and fixed in 4% formaldehyde for routine processing and embedding, as previously described [
14]. Four-micron thick sections were cut and stained with hematoxylin–eosin (H & E) or with periodic acid Schiff (PAS) without hematoxylin. Paraffin sections were cut at 4 μm for light microscopic immunohistochemistry (IHC). The slides were deparaffinized using xylene and absolute ethanol, rinsed in distilled water, and exposed to H
2O
2 for 5 min. or antigen unmasking solution. The antigen unmasking solution (citrate buffer, Thermo Scientific, Waltham, MA, USA) was heated in a steamer to 105 °C for 10 min. and then cooled to room temperature before use. The sections were rinsed with PBS for 2 min. (Cell Marque, Rocklin, CA, USA) and then blocked with CAS-Block for 5 min. (Invitrogen, Carlsbad, CA, USA, by ThermoFisher Scientific, Waltham, MA, USA). The slides were rinsed with PBS and reacted with primary antibodies targeting CD34 (Cell Marque), and Glut-1 (Cell Marque), followed by rinsing in PBS and reaction with secondary antibodies: MACH-2 rabbit HRP-Polymer (Biocare Medical, Pike Lane Concord, CA, USA) and HRP linked to rabbit anti-FITC, respectively. Sections were rinsed with PBS and incubated with AEC (Cell Marque) for 10 min. Sections were rinsed in distilled water and PBS and counterstained with Mayer’s hematoxylin, and coverslipped with a permanent mounting medium (AquaSlip, American MasterTech, Lodi, CA, USA). The slides were photographed with a Nikon ECLIPSE Ti microscope at ×4 magnification.
4.8. Statistical Analysis
In vitro experiments with at least three biological and three technical repeats for each data point provide sufficient statistical power. For animal experiments, we used five animals per group to allow descriptive statistics on the one hand while minimizing the use of animals to accommodate the ethical restrictions on the other hand.
Statistical analysis was performed with JMP 9.0 (SAS, Cary, NC, USA). Analysis of variance (ANOVA) was used to compare the mRNA levels and luciferase activity in relative light units. Multivariate ANOVA was used to compare the growth rates of the tumors within the mice (
Figure 5 and
Figure 6).
5. Conclusions
Clinical and research studies have shown that uveal melanoma is resistant to hypoxia. We have confirmed UM’s resistance to hypoxia in vitro [
17], and used armed viruses that knock down the expression of the regulators of cellular response to hypoxia, CREB, and HIF-1 to learn whether inhibiting the cells’ response to hypoxia would result in diminished tumor growth. Despite the lack of response in vitro, here we found that knockdown of CREB or HIF-1 in vivo resulted in a markedly decreased tumor growth rate. The resulting tumors were smaller in size and weight, had no vasculature growing into them, and had increased expression of vasculogenic mimicry patterns (more so with the knockdown of CREB).
The expression of the major cancer glucose transporter Glut-1, which is regulated by CREB and HIF-1, was diminished in UM tumors following the knockdown of either CREB or HIF-1. The resultant reduction in glucose uptake together with lower availability of glucose in vivo vs. in vitro probably lead to the curbed growth of the UM cells in vivo. In summary, the results presented here, demonstrating that the knockdown of either CREB or HIF-1 reduced UM tumor growth, may lead to the development of a novel therapeutic approach for metastatic UM, which is currently incurable.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






