
Molecular Epidemiology of Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus in the Ovine Dairy Chain and in Farm-Related Humans

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sample Collection
5.2. Typing of the Bacterial Isolates
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Bergonier, D.; de Crémoux, R.; Rupp, R.; Lagriffoul, G.; Berthelot, X. Mastitis of dairy small ruminants. Vet. Res. 2003, 34, 28, 689–716. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, L.M.; Zilta, M.; De Almeida, P.; De Mendonça, C.L.; Mamizuka, E.M. Novel sequence types (STs) of Staphylococcus aureus isolates causing clinical and subclinical mastitis in flocks of sheep in the northeast of Brazil. J. Dairy Res. 2011, 78, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Mørk, T.; Waage, S.; Tollersrud, T.; Kvitle, B.; Sviland, S. Clinical mastitis in ewes; bacteriology, epidemiology and clinical features. Scand. Acta Vet. 2007, 49, 1–8. [Google Scholar] [CrossRef] [PubMed]
- De Buyser, M.-L.; Dufour, B.; Maire, M.; Lafargé, V.; De, M.-L. Implication of milk and milk products in food-borne diseases in France and in different industrialised countries. Int. J. Food Microbiol. 2001, 67, 1–17. [Google Scholar] [CrossRef]
- Keefe, G. Update on control of Staphylococcus aureus and Streptococcus agalactiae for management of mastitis. Vet. Clin. North Am. Food Anim. Pract. 2012, 28, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Vautor, E.; Abadie, G.; Guibert, J.M.; Huard, C.; Pépin, M. Genotyping of Staphylococcus aureus isolated from various sites on farms with dairy sheep using pulsed-field gel electrophoresis. Vet. Microbiol. 2003, 96, 69–79. [Google Scholar] [CrossRef]
- Concepcion Porrero, M.; Hasman, H.; Vela, A.I.; Fernandez-Garayzabal, J.F.; Dominguez, L.; Aarestrup, F.M. Clonal diversity of Staphylococcus aureus originating from the small ruminants goats and sheep. Vet. Microbiol. 2012, 156, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Guinane, C.M.; Ben Zakour, N.L.; Tormo-Mas, M.A.; Weinert, L.A.; Lowder, B.V.; Cartwright, R.A.; Smyth, D.S.; Smyth, C.J.; Lindsay, J.A.; Gould, K.A.; et al. Evolutionary genomics of Staphylococcus aureus reveals insights into the origin and molecular basis of ruminant host adaptation. Genome Biol. Evol. 2010, 2, 454–466. [Google Scholar] [CrossRef] [PubMed]
- Argudín, M.Á.; Mendoza, M.C.; Rodicio, M.R. Food poisoning and Staphylococcus aureus enterotoxins. Toxins (Basel) 2010, 2, 1751–1773. [Google Scholar] [CrossRef] [PubMed]
- Johler, S.; Sihto, H.-M.; Macori, G.; Stephan, R. Sequence variability in staphylococcal enterotoxin genes seb, sec, and sed. Toxins (Basel) 2016, 8, 169. [Google Scholar] [CrossRef] [PubMed]
- Hodille, E.; Cuerq, C.; Badiou, C.; Bienvenu, F.; Steghens, J.-P.; Cartier, R.; Bes, M.; Tristan, A.; Plesa, A.; Le, V.T.M.; et al. Delta Hemolysin and Phenol-Soluble Modulins, but Not Alpha Hemolysin or Panton-Valentine Leukocidin, Induce Mast Cell Activation. Front. Cell. Infect. Microbiol. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Traversa, A.; Gariano, G.R.; Gallina, S.; Bianchi, D.M.; Orusa, R.; Domenis, L.; Cavallerio, P.; Fossati, L.; Serra, R.; Decastelli, L. Methicillin resistance in Staphylococcus aureus strains isolated from food and wild animal carcasses in Italy. Food Microbiol. 2015, 52, 154–158. [Google Scholar] [CrossRef] [PubMed]
- De Neeling, A.J.; van den Broek, M.J.M.; Spalburg, E.C.; van Santen-Verheuvel, M.G.; Dam-Deisz, W.D. C.; Boshuizen, H.C.; van de Giessen, A.W.; van Duijkeren, E.; Huijsdens, X.W. High prevalence of methicillin resistant Staphylococcus aureus in pigs. Vet. Microbiol. 2007, 3, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Argudín, M.A.; Tenhagen, B.A.; Fetsch, A.; Sachsenröder, J.; Käsbohrer, A.; Schroeter, A.; Hammer, J.A.; Hertwig, S.; Helmuth, R.; Bräunig, J.; et al. Virulence and resistance determinants of German Staphylococcus aureus ST398 isolates from nonhuman sources. Appl. Environ. Microbiol. 2011, 77, 3052–3060. [Google Scholar] [CrossRef] [PubMed]
- Pilla, R.; Piccinini, R.; Gelain, M.E.; Lorenzi, V.; Castiglioni, V.; Scanziani, E.; Anjum, M. Long-term study of MRSA ST1, t127 mastitis in a dairy cow. Vet. Rec. 2012, 170, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Carfora, V.; Caprioli, A.; Marri, N.; Sagrafoli, D.; Boselli, C.; Giacinti, G.; Giangolini, G.; Sorbara, L.; Dottarelli, S.; Battisti, A.; et al. Enterotoxin genes, enterotoxin production, and methicillin resistance in Staphylococcus aureus isolated from milk and dairy products in Central Italy. Int. Dairy J. 2015, 42, 12–15. [Google Scholar] [CrossRef]
- Carfora, V.; Giacinti, G.; Sagrafoli, D.; Marri, N.; Giangolini, G.; Alba, P.; Feltrin, F.; Sorbara, L.; Amoruso, R.; Caprioli, A.; et al. Methicillin-resistant and methicillin-susceptible Staphylococcus aureus in dairy sheep and in-contact humans: An intra-farm study. J. Dairy Sci. 2016, 99, 4251–4258. [Google Scholar] [CrossRef]
- Caruso, M.; Latorre, L.; Santagada, G.; Fraccalvieri, R.; Miccolupo, A.; Sottili, R.; Palazzo, L.; Parisi, A. Methicillin-resistant Staphylococcus aureus (MRSA) in sheep and goat bulk tank milk from Southern Italy. Small Rumin. Res. 2016, 135, 26–31. [Google Scholar] [CrossRef]
- Cortimiglia, C.; Bianchini, V.; Franco, A.; Caprioli, A.; Battisti, A.; Colombo, L.; Stradiotto, K.; Vezzoli, F.; Luini, M. Short communication: Prevalence of Staphylococcus aureus and methicillin-resistant S. aureus in bulk tank milk from dairy goat farms in Northern Italy. J. Dairy Sci. 2015, 98, 2307–2311. [Google Scholar] [CrossRef] [PubMed]
- Alba, P.; Feltrin, F.; Cordaro, G.; Porrero, M.C.; Kraushaar, B.; Argudín, M.A.; Nykäsenoja, S.; Monaco, M.; Stegger, M.; Aarestrup, F.M.; et al. Livestock-associated methicillin resistant and methicillin susceptible Staphylococcus aureus sequence type (CC)1 in European farmed animals: High genetic relatedness of isolates from Italian cattle herds and humans. PLoS One 2015, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.; Hasman, H.; Iurescia, M.; Lorenzetti, R.; Stegger, M.; Pantosti, A.; Feltrin, F.; Ianzano, A.; Porrero, M.C.; Liapi, M.; et al. Molecular characterization of spa-type t127, sequence type 1 methicillin-resistant Staphylococcus aureus from pigs. J. Antimicrob. Chemother. 2011, 66, 1231–1235. [Google Scholar] [CrossRef] [PubMed]
- Boyle-Vavra, S.; Daum, R.S. Community-acquired methicillin-resistant Staphylococcus aureus: The role of Panton–Valentine leukocidin. Lab. Investig. 2007, 87, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Brennan, G.I.; Abbott, Y.; Burns, A.; Leonard, F.; McManus, B.A.; O’Connell, B.; Coleman, D.C.; Shore, A.C. The emergence and spread of multiple livestock-associated clonal complex 398 methicillin-resistant and methicillin-susceptible Staphylococcus aureus strains among animals and humans in the Republic of Ireland, 2010–2014. PLoS One 2016, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wulf, M.W.H.; Markestein, A.; van der Linden, F.T.; Voss, A.; Klaassen, C.; Verduin, C.M. First outbreak of methicillin-resistant Staphylococcus aureus ST398 in a Dutch hospital, June 2007. Euro Surveill. 2008, 13, 1–2. [Google Scholar]
- Juhász-Kaszanyitzky, E.; Jánosi, S.; Somogyi, P.; Dán, A.; Van Der Graaf-van Bloois, L.; Van Duijkeren, E.; Wagenaar, J.A. MRSA transmission between cows and humans. Emerg. Infect. Dis. 2007, 13, 630–632. [Google Scholar] [CrossRef] [PubMed]
- Hanselman, B.A.; Kruth, S.A.; Rousseau, J.; Low, D.E.; Willey, B.M.; McGeer, A.; Weese, J.S. Methicillin resistant Staphylococcus aureuscolonization in veterinary personnel. Emerg. Infect. Dis. 2006, 12, 1933–1938. [Google Scholar] [CrossRef] [PubMed]
- Kluytmans, J.A.J.W. Methicillin-resistant Staphylococcus aureus in food products: Cause for concern or case for complacency? Clin. Microbiol. Infect. 2010, 16, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Basanisi, M.G.; Nobili, G.; La Bella, G.; Russo, R.; Spano, G.; Normanno, G.; La Salandra, G. Molecular characterization of Staphylococcus aureus isolated from sheep and goat cheeses in southern Italy. Small Rumin. Res. 2016, 135, 17–19. [Google Scholar] [CrossRef]
- Spanu, V.; Scarano, C.; Cossu, F.; Pala, C.; Spanu, C.; De Santis, E.P.L. Antibiotic resistance traits and molecular subtyping of Staphylococcus aureus isolated from raw sheep milk cheese. J. Food Sci. 2014, 16, 11–15. [Google Scholar] [CrossRef]
- Hernández, M.; Fernández-Natal, I.; Rodríguez-Lázaro, D. Methicillin-resistant Staphylococcus aureus harboring mecC in livestock in Spain. J. Clin. Microbiol. 2014, 52, 4067–4069. [Google Scholar] [CrossRef]
- Pexara, A.; Solomakos, N.; Sergelidis, D.; Angelidis, A.S.; Govaris, A.; Gr, A.U. Occurrence and antibiotic resistance of enterotoxigenic Staphylococcus aureus in raw ovine and caprine milk in Greece. Dairy Sci. Technol 2016, 96, 345–357. [Google Scholar] [CrossRef]
- Blaiotta, G.; Fusco, V.; von Eiff, C.; Villani, F.; Becker, K. Biotyping of enterotoxigenic Staphylococcus aureus by enterotoxin gene cluster (egc) polymorphism and spa typing analyses. Appl. Environ. Microbiol. 2006, 72, 6117–6123. [Google Scholar] [CrossRef] [PubMed]
- Mello, P.L.; Riboli, D.F.M.; Pinheiro, L.; Martins, L. de A.; Brito, M.A.V.P.; da Cunha, M. de L.R. de S. Detection of enterotoxigenic potential and determination of clonal profile in Staphylococcus aureus and coagulase-negative staphylococci isolated from bovine subclinical mastitis in different Brazilian states. Toxins (Basel). 2016, 8, 1–10. [Google Scholar] [CrossRef]
- Del Giudice, P.; Blanc, V.; De Rougemont, A.; Bes, M.; Lina, G.; Hubiche, T.; Roudière, L.; Vandenesch, F.; Etienne, J. Primary skin abscesses are mainly caused by panton-valentine leukocidin-positive Staphylococcus aureus strains. Dermatology 2009, 219, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Zecconi, A.; Cesaris, L.; Liandris, E.; Daprà, V.; Piccinini, R. Role of several Staphylococcus aureus virulence factors on the inflammatory response in bovine mammary gland. Microb. Pathog. 2006, 40, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Aires-de-Sousa, M. Methicillin-resistant Staphylococcus aureus among animals: Current overview. Clin. Microbiol. Infect. 2016, S1198–743X(16)30557–2. [Google Scholar]
- Esposito, S.; Purrello, S.M.; Bonnet, E.; Novelli, A.; Tripodi, F.; Pascale, R.; Unal, S.; Milkovich, G. Central venous catheter-related biofilm infections: An up-to-date focus on meticillin-resistant Staphylococcus aureus. J. Glob. Antimicrob. Resist. 2013, 1, 71–78. [Google Scholar] [CrossRef]
- Macedo-Viñas, M.; Conly, J.; Francois, P.; Aschbacher, R.; Blanc, D.S.; Coombs, G.; Daikos, G.; Dhawan, B.; Empel, J.; Etienne, J.; et al. Antibiotic susceptibility and molecular epidemiology of Panton-Valentine leukocidin-positive meticillin-resistant Staphylococcus aureus: An international survey. J. Glob. Antimicrob. Resist. 2014, 2, 43–47. [Google Scholar] [CrossRef]
- Stegger, M.; Andersen, P.S.; Kearns, A.; Pichon, B.; Holmes, M.A.; Edwards, G.; Laurent, F.; Teale, C.; Skov, R.; Larsen, A.R. Rapid detection, differentiation and typing of methicillin-resistant Staphylococcus aureus harbouring either mecA or the new mecA homologue mecALGA251. Clin. Microbiol. Infect. 2012, 18, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Cuny, C.; Layer, F.; Werner, G.; Harmsen, D.; Daniels-Haardt, I.; Jurke, A.; Mellmann, A.; Witte, W.; Köck, R. State-wide surveillance of antibiotic resistance patterns and spa-types of methicillin-resistant Staphylococcus aureus from blood cultures in North Rhine-Westphalia, 2011–2013. Clin. Microbiol. Infect. 2015, 21, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Walther, B.; Wieler, L.H.; Vincze, S.; Antão, E.M.; Brandenburg, A.; Stamm, I.; Kopp, P.A.; Kohn, B.; Semmler, T.; Lübke-Becker, A. MRSA variant in companion animals. Emerg. Infect. Dis. 2012, 18, 2017–2020. [Google Scholar] [CrossRef] [PubMed]
- Vincze, S.; Brandenburg, A.G.; Espelage, W.; Stamm, I.; Wieler, L.H.; Kopp, P.A.; Lübke-Becker, A.; Walther, B. Risk factors for MRSA infection in companion animals: Results from a case-control study within Germany. Int. J. Med. Microbiol. 2014, 304, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, W.B.; Melles, D.C.; Alaidan, A.; Al-Ahdal, M.; Boelens, H.A.M.; Snijders, S.V.; Wertheim, H.; Van Duijkeren, E.; Peeters, J.K.; Van Der Spek, P.J.; et al. Host- and tissue-specific pathogenic traits of Staphylococcus aureus. J. Bacteriol. 2005, 187, 4584–4591. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H. Methicillin (Oxacillin)-Resistant Staphylococcus aureus Strains Isolated from Major Food Animals and Their Potential Transmission to Humans. Appl. Environ. Microbiol. 2003, 69, 6489–6494. [Google Scholar] [CrossRef] [PubMed]
- Merz, A.; Stephan, R.; Johler, S. Staphylococcus aureus isolates from goat and sheep milk seem to be closely related and differ from isolates detected from bovine milk. Front. Microbiol. 2016, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Diep, B.A.; Gill, S.R.; Chang, R.F.; Phan, T.H.; Chen, J.H.; Davidson, M.G.; Lin, F.; Lin, J.; Carleton, H.A. Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. Lancet 2006, 367, 731–739. [Google Scholar] [CrossRef]
- Puah, S.M.; Chua, K.H.; Mary Anne Tan, J.A. Virulence factors and antibiotic susceptibility of Staphylococcus aureus isolates in ready-to-eat foods: Detection of S. aureus contamination and a high prevalence of virulence genes. Int. J. Environ. Res. Public Health 2016, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Johler, S.; Giannini, P.; Jermini, M.; Hummerjohann, J.; Baumgartner, A.; Stephan, R. Further evidence for staphylococcal food poisoning outbreaks caused by egc-Encoded enterotoxins. Toxins (Basel). 2015, 7, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Chen, Z.; Liu, C.; Zhang, X.; Lin, X.; Chi, S.; Zhou, T.; Chen, Z.; Chen, X. Prevalence of Staphylococcus aureus carrying Panton-Valentine leukocidin genes among isolates from hospitalised patients in China. Clin. Microbiol. Infect. 2008, 14, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Morandi, S.; Brasca, M.; Andrighetto, C.; Lombardi, A.; Lodi, R. Phenotypic and Genotypic Characterization of Staphylococcus aureus Strains from Italian Dairy Products. Int. J. Microbiol. 2010, 2009. [Google Scholar] [CrossRef]
- Scherrer, D.; Corti, S.; Muehlherr, J.E.; Zweifel, C.; Stephan, R. Phenotypic and genotypic characteristics of Staphylococcus aureus isolates from raw bulk-tank milk samples of goats and sheep. Vet. Microbiol. 2004, 101, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, M.; Wang, G.; Johnson, W.M. Multiplex PCR for detection of genes for Staphylococcus aureus enterotoxins, exfoliative toxins, toxic shock syndrome toxin 1, and methicillin resistance. J. Clin. Microbiol. 2000, 38, 1032–1035. [Google Scholar] [PubMed]
- Kérouanton, A.; Hennekinne, J.A.; Letertre, C.; Petit, L.; Chesneau, O.; Brisabois, A.; De Buyser, M. Characterization of Staphylococcus aureus strains associated with food poisoning outbreaks in France. Int. J. Food Microbiol. 2007, 115, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Devriese, L.A. A simplified system for biotyping Staphylococcus aureus strains isolated from animal species. J. Appl. Bacteriol. 1984, 56, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Hennekinne, J.A.; Kerouanton, A.; Brisabois, A.; De Buyser, M.L. Discrimination of Staphylococcus aureus biotypes by pulsed-field gel electrophoresis of DNA macro-restriction fragments. J. Appl. Microbiol. 2003, 94, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, D.; Claus, H.; Witte, W.; Rothgänger, J.; Claus, H.; Turnwald, D.; Vogel, U. Typing of Methicillin-Resistant Staphylococcus aureus in a University Hospital Setting by Using Novel Software for spa Repeat Determination and Database Management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [PubMed]
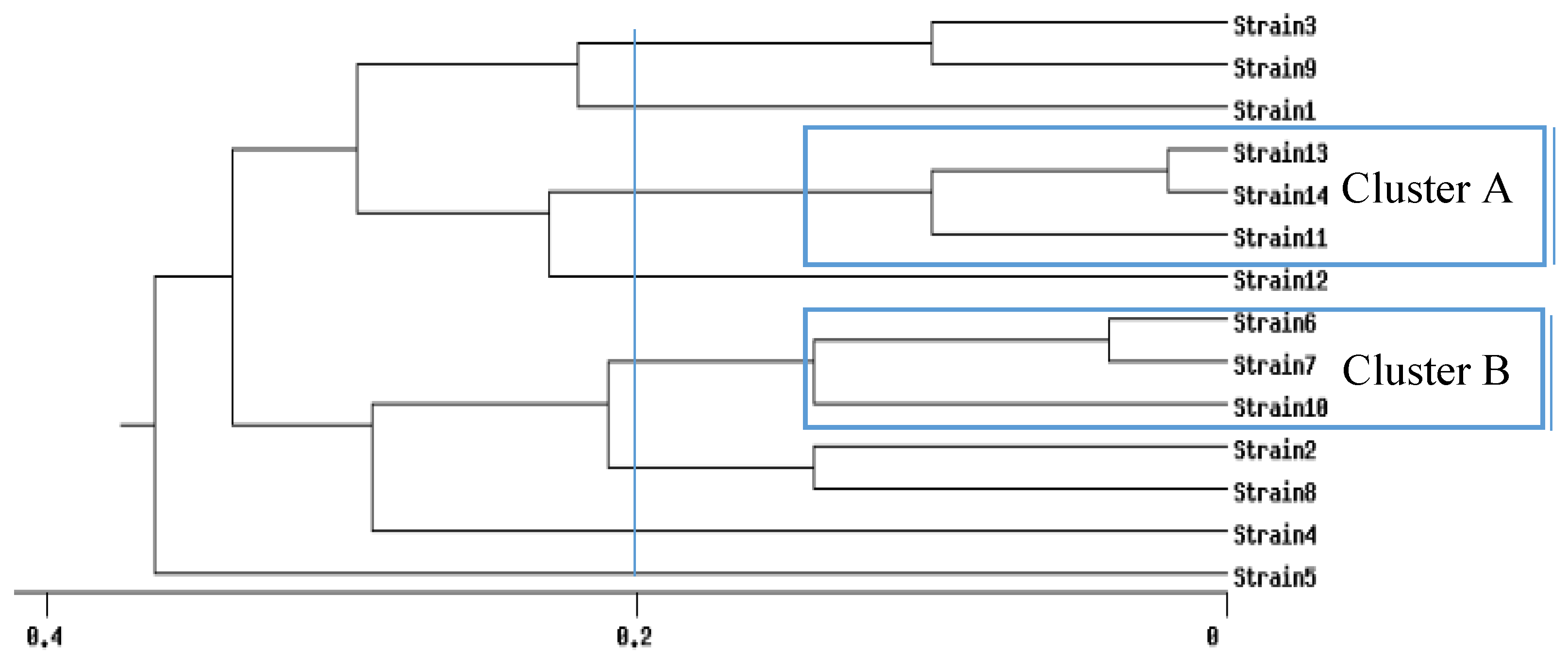

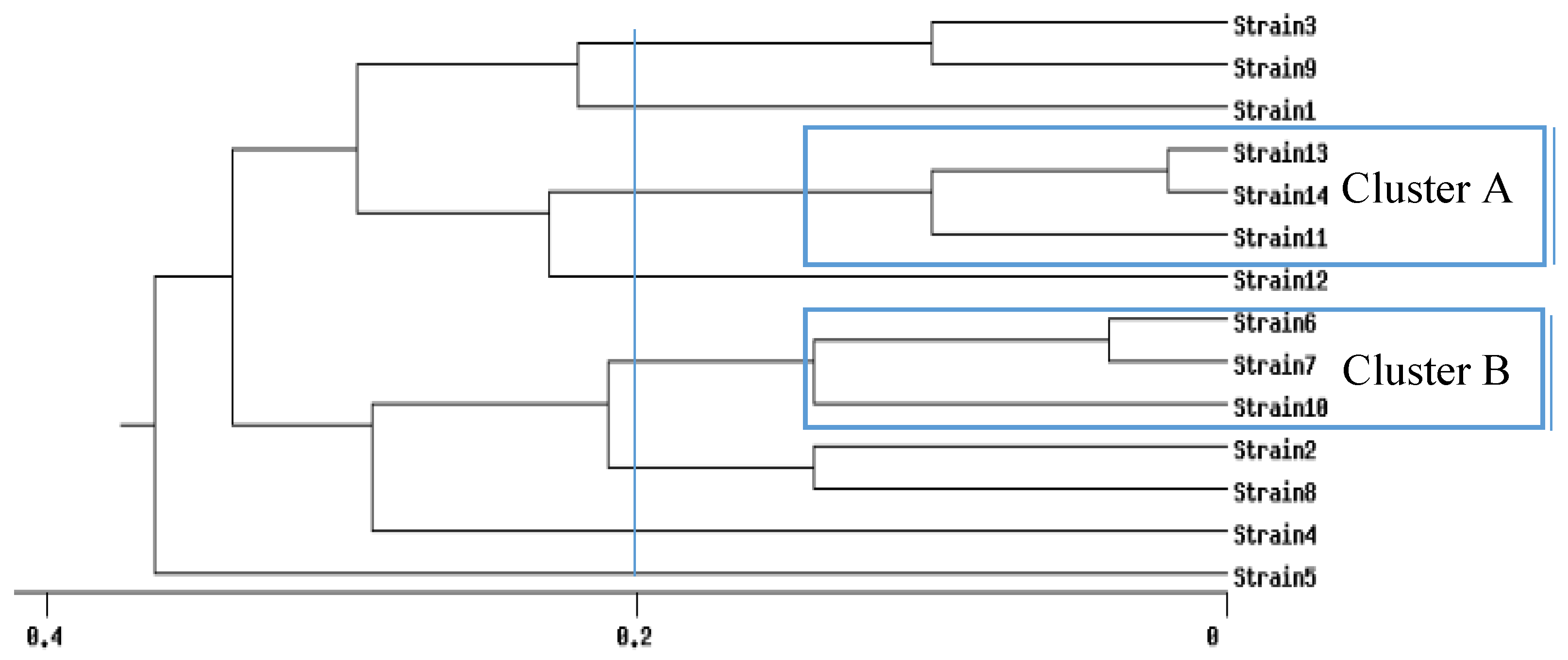{kind=link}
| Staphylococcoal Enterotoxin (SE) Profile | Staphylococcal Enterotoxin Gene(s) | No. of Isolates (%) |
|---|---|---|
| 1 | seh | 17 (33.3) |
| 2 | Negative | 10 (19.7) |
| 3 | sec | 9 (17.6) |
| 4 | seg | 8 (15.7) |
| 5 | seg-sei | 5 (9.8) |
| 6 | sec-seg-sei | 2 (3.9) |
| Farm Number | Farm Workers/Owners Tested | S. aureus | MRSA | |
|---|---|---|---|---|
| 1 | 5 | 2 | 1 | |
| 2 | 2 | 4 | 2 | |
| 3 | 4 | 3 | 1 | |
| 4 | 1 | 3 | 1 | |
| 5 | 1 | 0 | 0 | |
| 6 | 1 | 0 | 0 | |
| Total | 6 | 14 | 12 | 5 |
| ID | pvl | mecA | SEsP | spa-type | SmaI | Biotype | ID | pvl | mecA | SEsP | spa-type | SmaI | Biotype |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | − | + | 4 | t1166 | 6 | NSH4 | 27 | + | + | 2 | t044 | 5 | NSH2 |
| 2 | − | + | 4 | t1773 | 7 | Human | 28 | − | + | 1 | t127 | 6 | NSH3 |
| 3 | − | + | 4 | t127 | 6 | NSH3 | 29 | − | + | 1 | t127 | 6 | NSH6 |
| 4 | − | + | 4 | t127 | 6 | NSH3 | 30 | − | + | 1 | t127 | 7 | Human |
| 5 | − | + | 4 | t127 | 7 | Human | 31 | − | − | 5 | t709 | 2 | Human |
| 6 | − | + | 4 | t127 | 7 | Human | 32 | − | − | 5 | t709 | 2 | Human |
| 7 | − | + | 4 | t2678 | 7 | Human | 33 | − | − | 6 | t050 | 8 | Human |
| 8 | − | − | 2 | t1773 | 12 | NB | 34 | − | + | 1 | t127 | 4 | Human |
| 9 | − | − | 3 | t3583 | 13 | Ovine | 35 | − | − | 6 | t050 | 8 | Human |
| 10 | − | − | 3 | t3583 | 13 | Ovine | 36 | − | + | 1 | t127 | 6 | NSH3 |
| 11 | − | − | 3 | t1773 | 13 | Ovine | 37 | − | + | 1 | t127 | 6 | NSH3 |
| 12 | − | − | 3 | t1166 | 13 | Ovine | 38 | − | + | 1 | t127 | 6 | NSH3 |
| 13 | − | + | 2 | t2678 | 7 | Human | 39 | − | + | 1 | t127 | 6 | NSH3 |
| 14 | − | − | 2 | t127 | 12 | Ovine | 40 | − | + | 1 | t127 | 6 | NSH3 |
| 15 | − | − | 3 | t1773 | 13 | Ovine | 41 | − | + | 1 | t127 | 6 | NSH3 |
| 16 | − | − | 3 | t2678 | 14 | Ovine | 42 | − | + | 1 | t127 | 9 | Human |
| 17 | − | + | 4 | t127 | 6 | NSH3 | 43 | − | − | 2 | t084 | 10 | NSH6 |
| 18 | − | − | 2 | t528 | 1 | Ovine | 44 | − | + | 1 | t127 | 6 | Human |
| 19 | − | − | 2 | t528 | 1 | Ovine | 45 | − | − | 3 | t998 | 11 | Ovine |
| 20 | − | − | 5 | t902 | 2 | Human | 46 | − | − | 3 | t1166 | 11 | Ovine |
| 21 | − | − | 2 | t127 | 3 | NSH2 | 47 | − | − | 3 | t1166 | 14 | Ovine |
| 22 | − | + | 1 | t127 | 4 | NSH3 | 48 | − | + | 1 | t127 | 6 | Human |
| 23 | − | + | 1 | t127 | 4 | NSH3 | 49 | − | − | 5 | t902 | 2 | Human |
| 24 | − | − | 2 | t528 | 3 | NSH3 | 50 | − | + | 1 | t127 | 6 | Human |
| 25 | − | − | 2 | t528 | 3 | NSH2 | 51 | − | − | 5 | t127 | 2 | Human |
| 26 | − | + | 1 | t127 | 4 | NSH3 |
| ID | Farm | Type of Sample | pvl | SEs Gene | Biotyping | spa-type | smaI | |
|---|---|---|---|---|---|---|---|---|
| 45 | 3 | Mastitis udder swab | − | sec | Ovine | t998 | 11 | |
| 46 | 3 | Udder | − | sec | Ovine | t1166 | 11 | |
| 9 | 3 | Individual milk | − | sec | Ovine | t3583 | 13 | |
| 10 | 3 | Individual milk | − | sec | Ovine | t3583 | 13 | |
| 11 | 3 | Mammary pustules | − | sec | Ovine | t1773 | 13 | |
| 12 | 3 | Mammary pustules | − | sec | Ovine | t1166 | 13 | |
| 15 | 3 | Swab skin (sheep) | − | sec | Ovine | t1773 | 13 | |
| 47 | 3 | Swab skin (sheep) | − | sec | Ovine | t1166 | 14 | |
| 16 | 3 | Swab skin (sheep) | − | sec | Ovine | t2678 | 14 | |
| Total | 9 | 1 | sec (100.0%) |
| ID | Farm | Type of Sample | pvl | SEs Gene | Biotyping | spa-type | smaI | |
|---|---|---|---|---|---|---|---|---|
| 1 | 4 | Bulk milk | − | seg | NSH4 | t1166 | 6 | |
| 2 | 3 | Bulk milk | − | seg | Human | t1773 | 7 | |
| 3 | 1 | Bulk milk | − | seg | NSH3 | t127 | 6 | |
| 4 | 1 | Cheese | − | seg | NSH3 | t127 | 6 | |
| 5 | 3 | Individual milk | − | seg | Human | t127 | 7 | |
| 6 | 3 | Individual milk | − | seg | Human | t127 | 7 | |
| 7 | 3 | Individual milk | − | seg | Human | t2678 | 7 | |
| 13 | 3 | Swab skin (ewe) | - | neg | Human | t2678 | 7 | |
| 17 | 4 | Cheese | − | seg | NSH3 | t127 | 6 | |
| 28 | 4 | Individual milk | − | seh | NSH3 | t127 | 6 | |
| 29 | 4 | Individual milk | − | seh | NSH6 | t127 | 6 | |
| 30 | 4 | Swab skin (human) | − | seh | Human | t127 | 7 | |
| 36 | 2 | Bulk milk | − | seh | NSH3 | t127 | 6 | |
| 37 | 1 | Bulk milk | − | seh | NSH3 | t127 | 6 | |
| 38 | 1 | Bulk milk | − | seh | NSH3 | t127 | 6 | |
| 39 | 1 | Bulk milk | − | seh | NSH3 | t127 | 6 | |
| 40 | 1 | Udder half | − | seh | NSH3 | t127 | 6 | |
| 41 | 1 | Environmental swab | − | seh | NSH3 | t127 | 6 | |
| 43 | 1 | Nasal swab | − | neg | NSH6 | t084 | 10 | |
| 44 | 1 | Bulk milk | − | seh | Human | t127 | 6 | |
| 48 | 3 | Nasal swab | − | seh | Human | t127 | 6 | |
| 50 | 3 | Nasal swab | − | seh | Human | t127 | 6 | |
| Total | 22 | 4 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macori, G.; Giacinti, G.; Bellio, A.; Gallina, S.; Bianchi, D.M.; Sagrafoli, D.; Marri, N.; Giangolini, G.; Amatiste, S.; Decastelli, L. Molecular Epidemiology of Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus in the Ovine Dairy Chain and in Farm-Related Humans. Toxins 2017, 9, 161. https://doi.org/10.3390/toxins9050161
Macori G, Giacinti G, Bellio A, Gallina S, Bianchi DM, Sagrafoli D, Marri N, Giangolini G, Amatiste S, Decastelli L. Molecular Epidemiology of Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus in the Ovine Dairy Chain and in Farm-Related Humans. Toxins. 2017; 9(5):161. https://doi.org/10.3390/toxins9050161
Chicago/Turabian StyleMacori, Guerrino, Giuseppina Giacinti, Alberto Bellio, Silvia Gallina, Daniela Manila Bianchi, Daniele Sagrafoli, Nicla Marri, Gilberto Giangolini, Simonetta Amatiste, and Lucia Decastelli. 2017. "Molecular Epidemiology of Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus in the Ovine Dairy Chain and in Farm-Related Humans" Toxins 9, no. 5: 161. https://doi.org/10.3390/toxins9050161