
Are We Underestimating Benthic Cyanotoxins? Extensive Sampling Results from Spain


Abstract
:1. Introduction
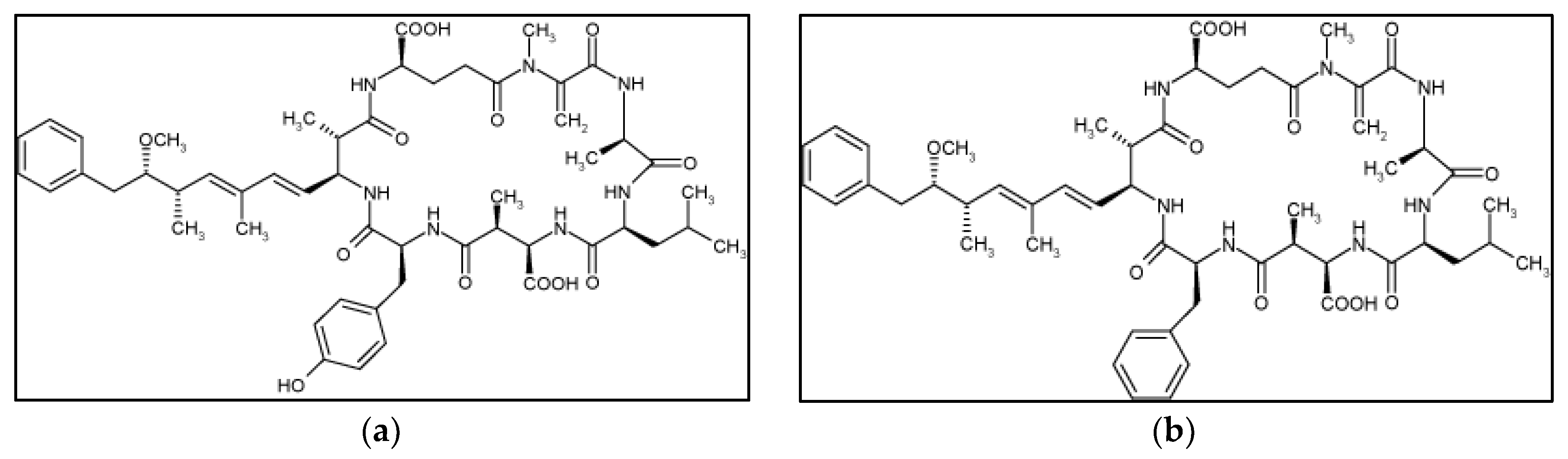
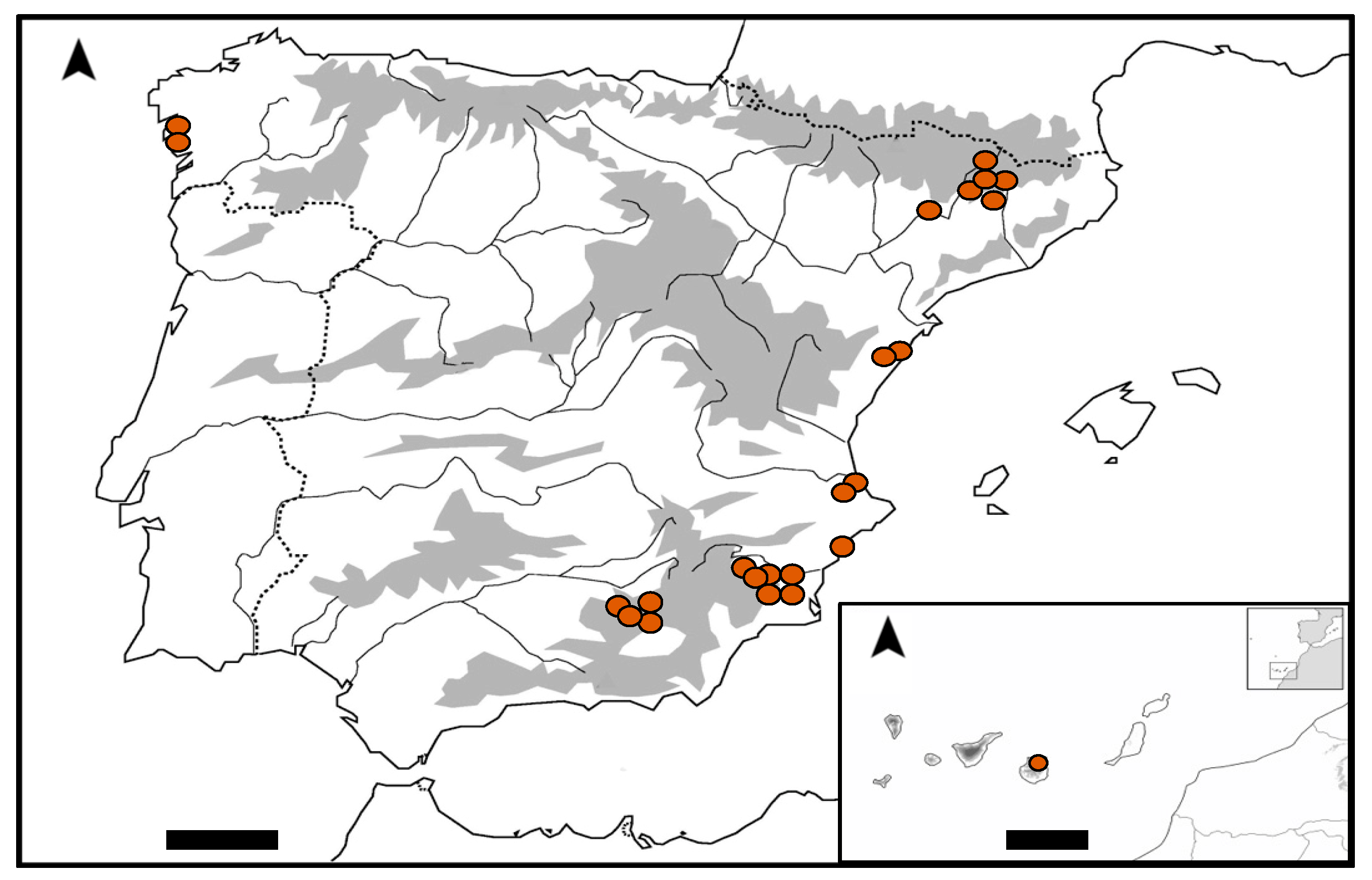
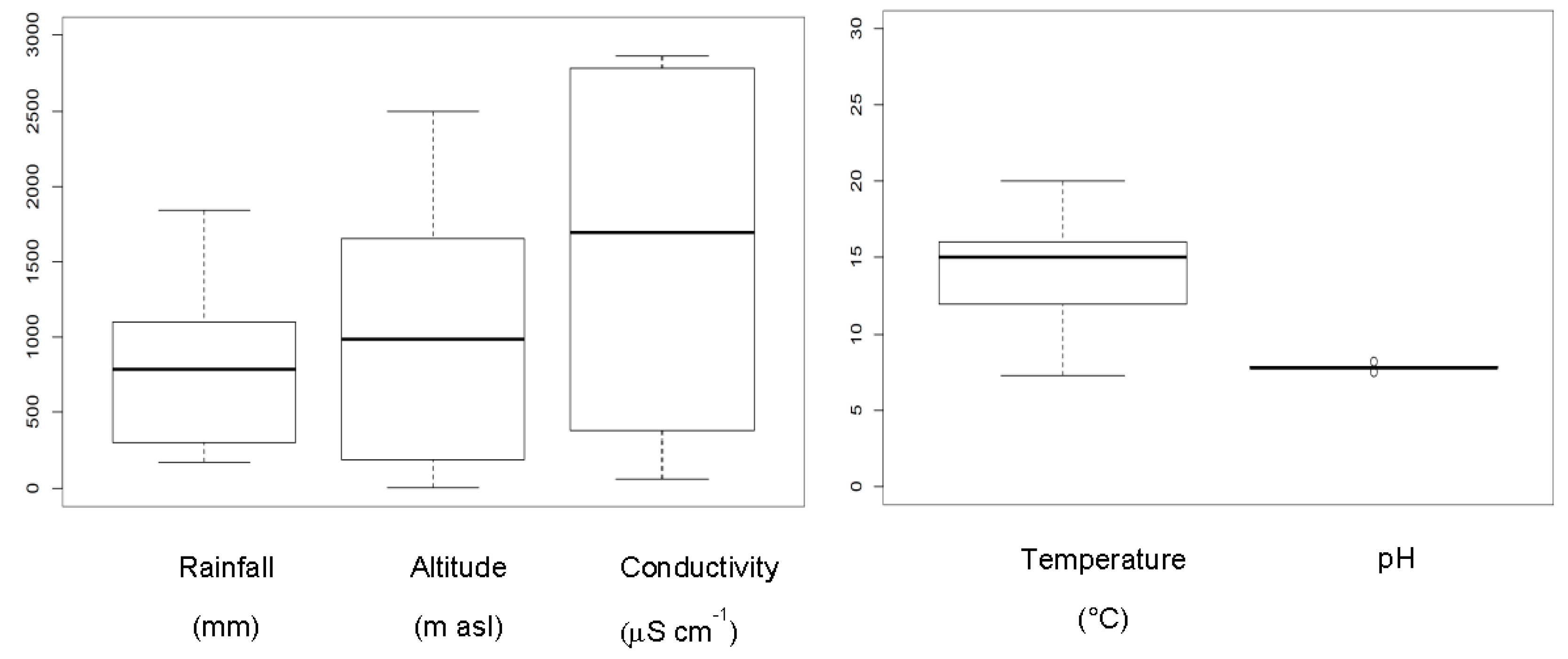
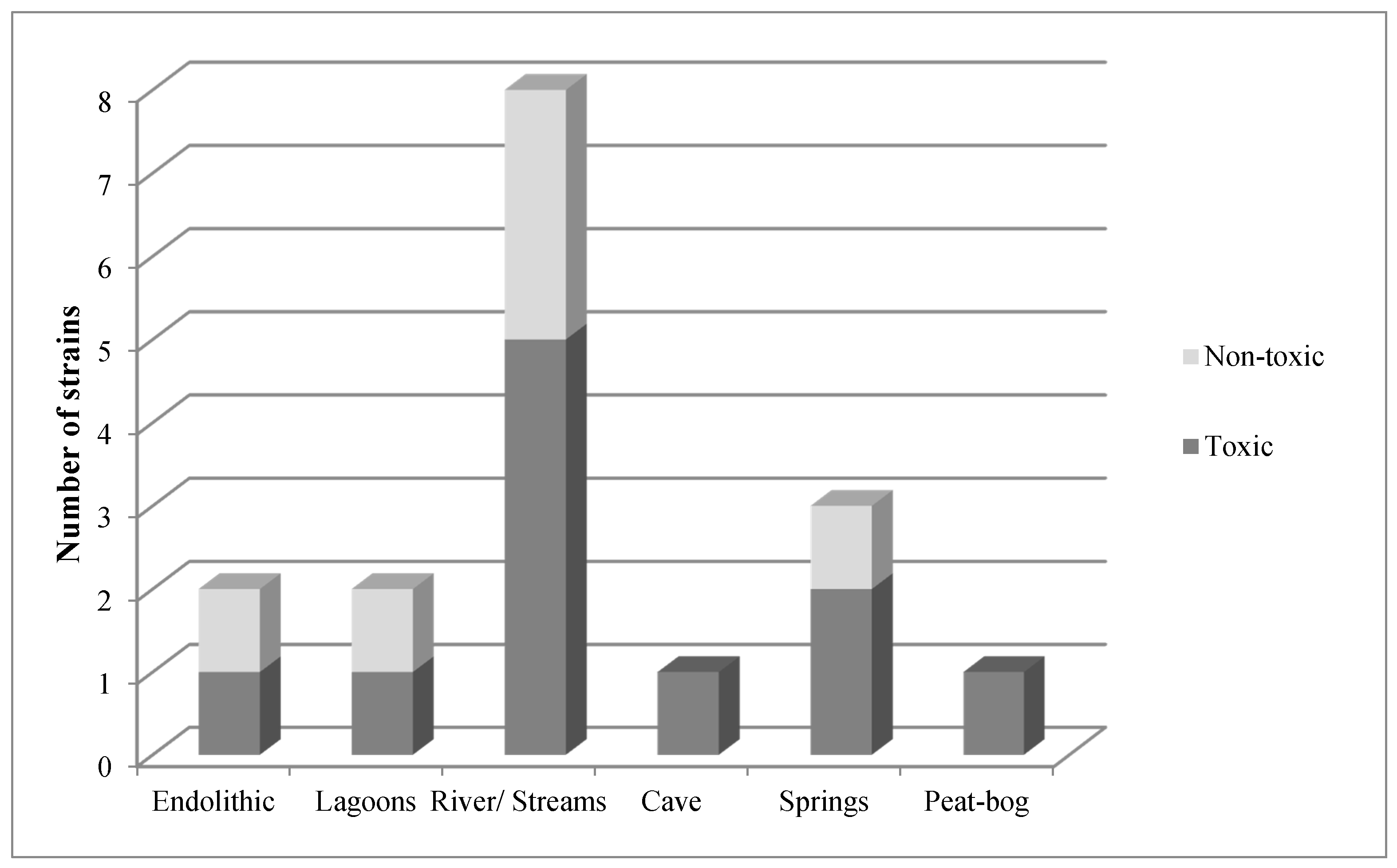
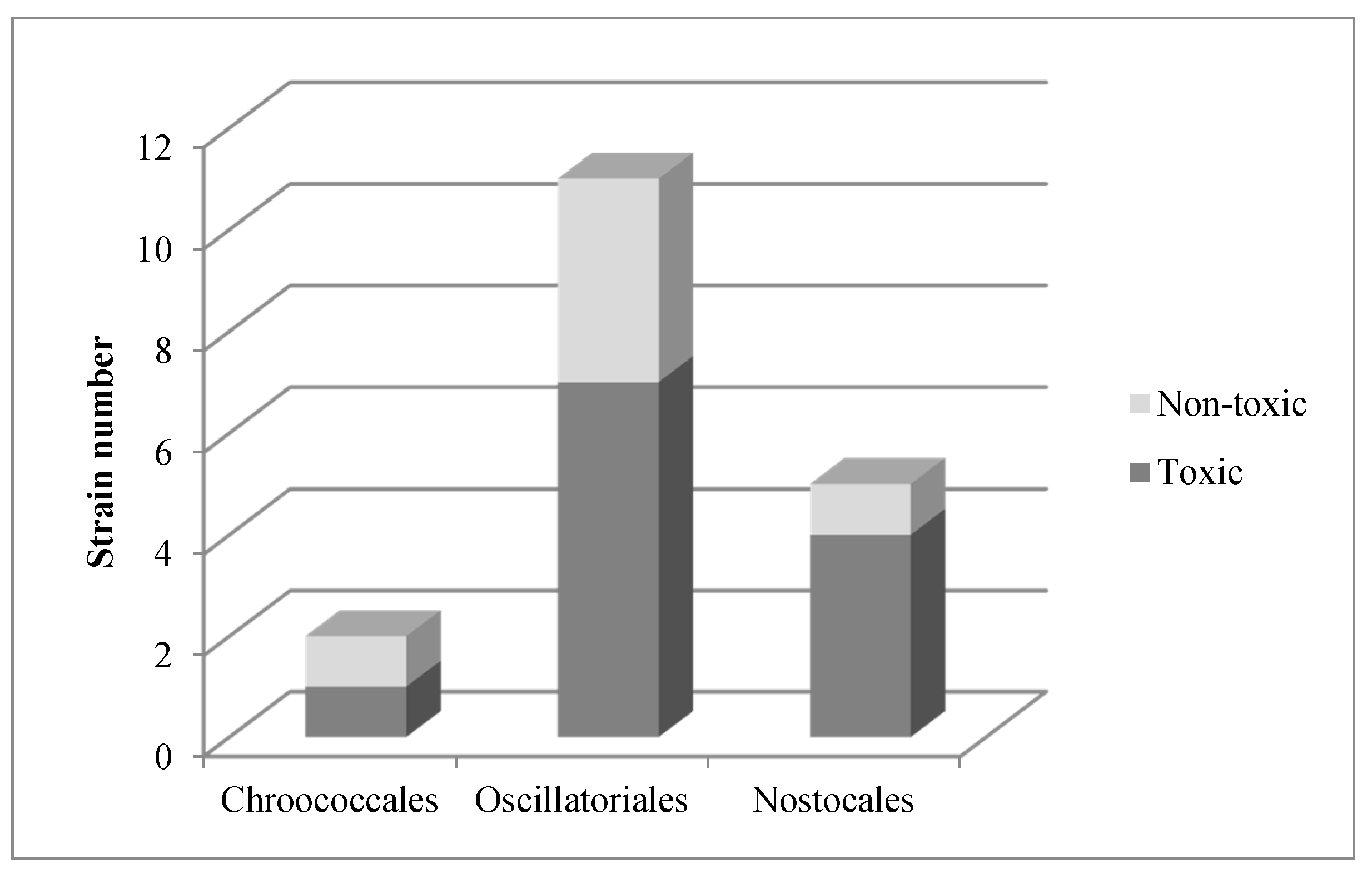
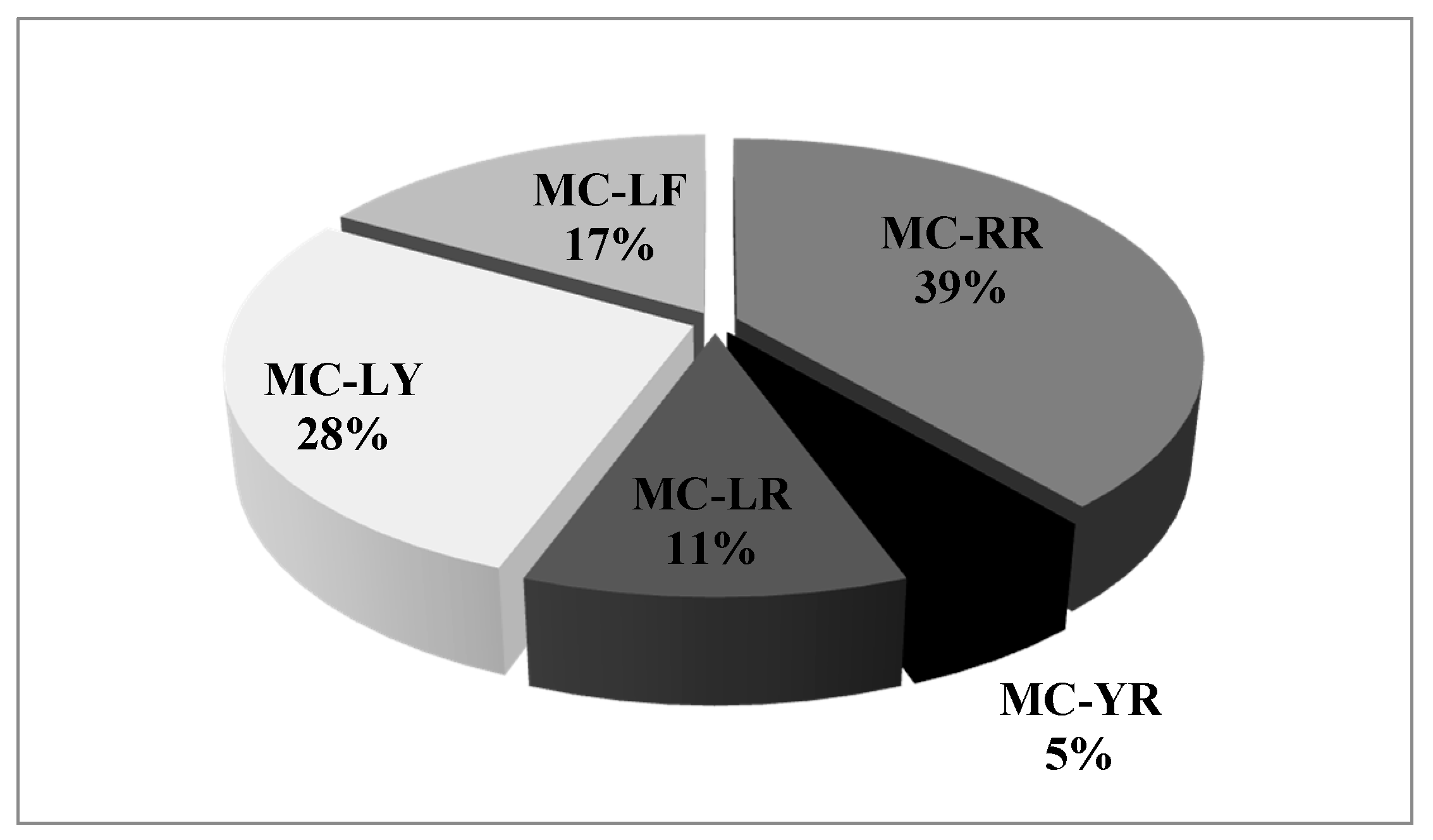
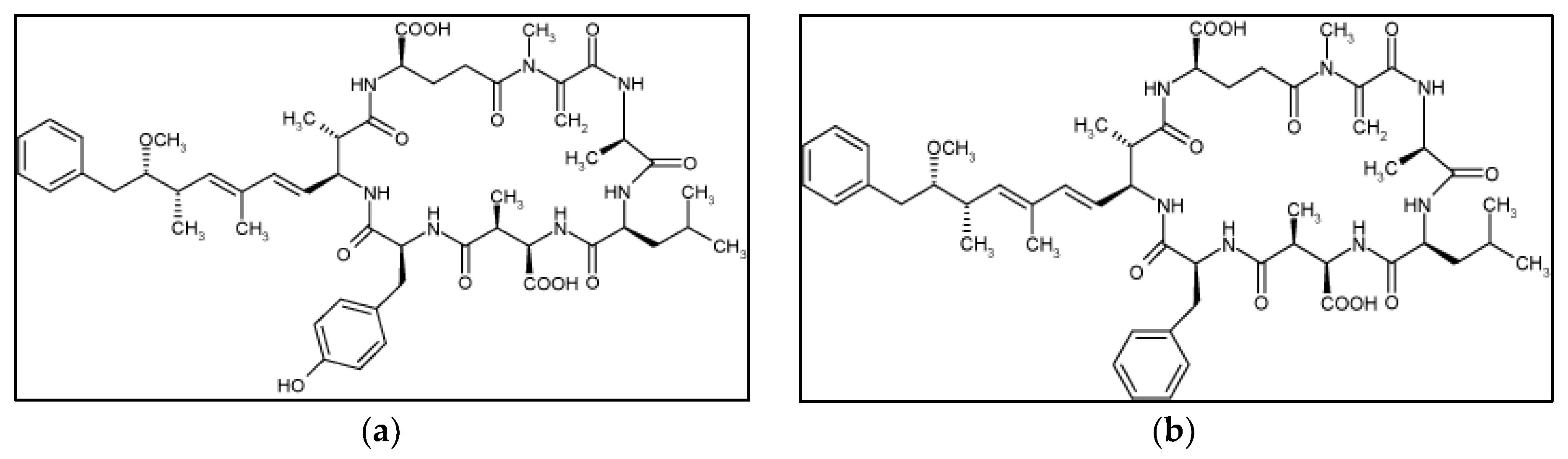
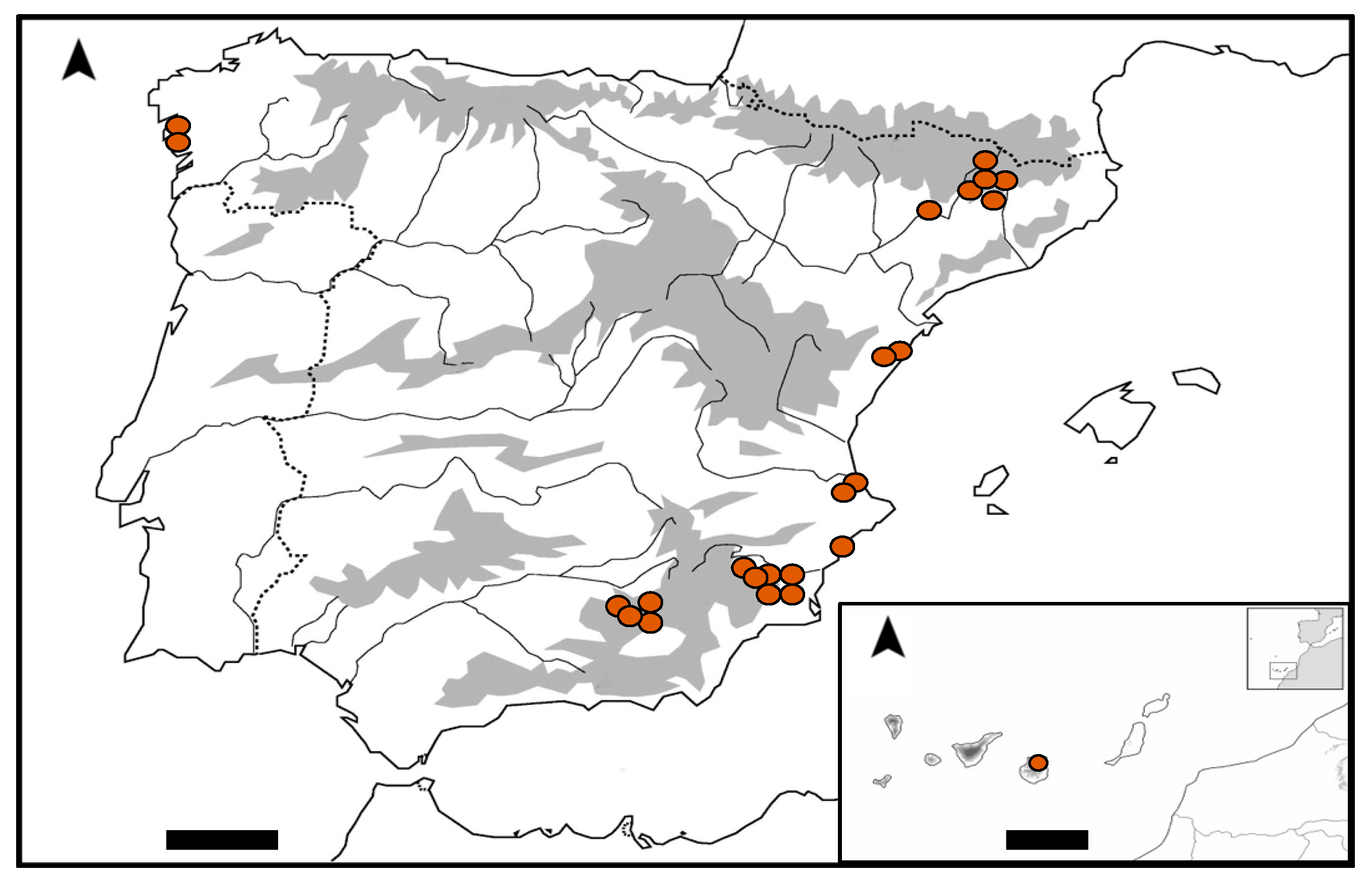
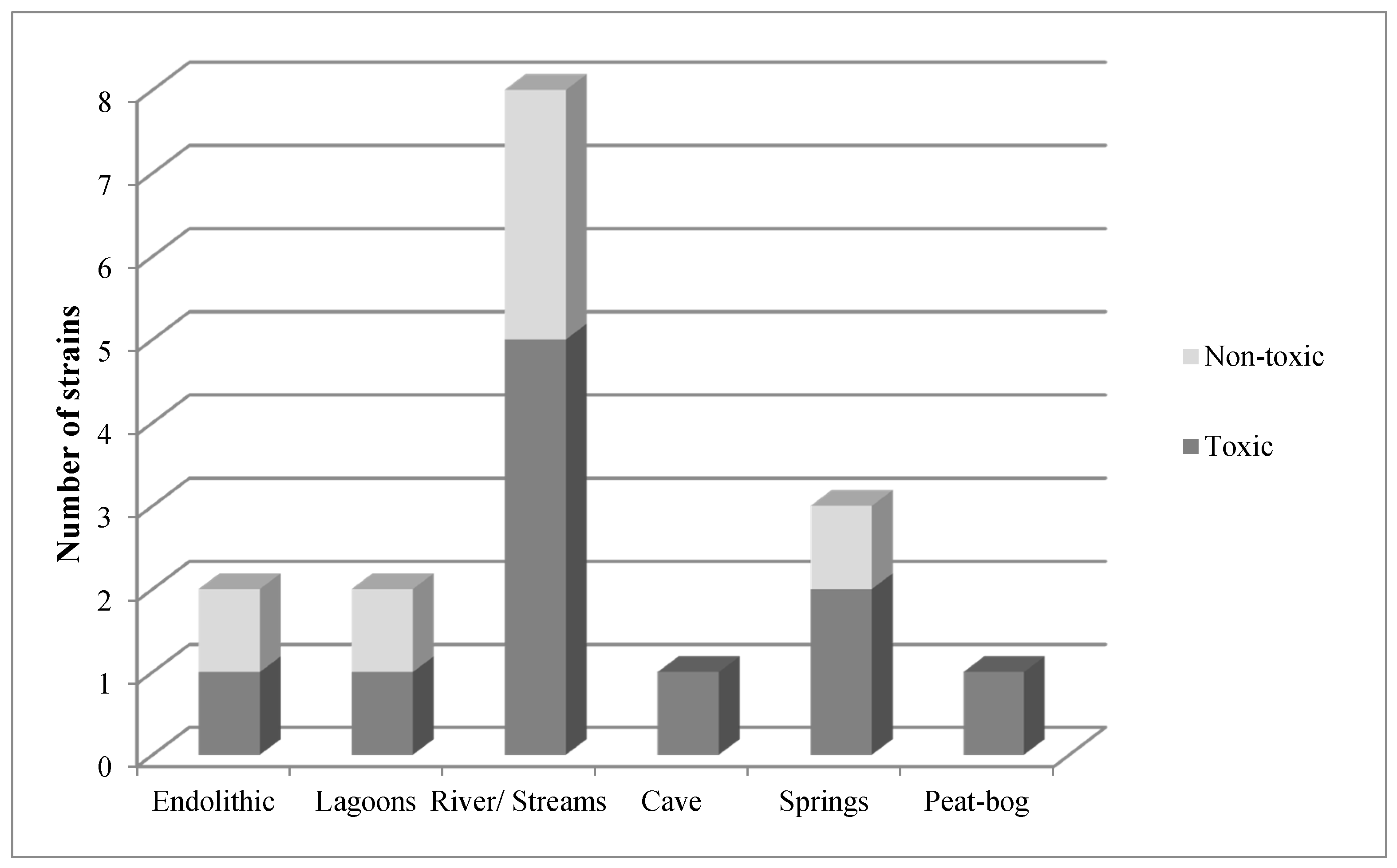
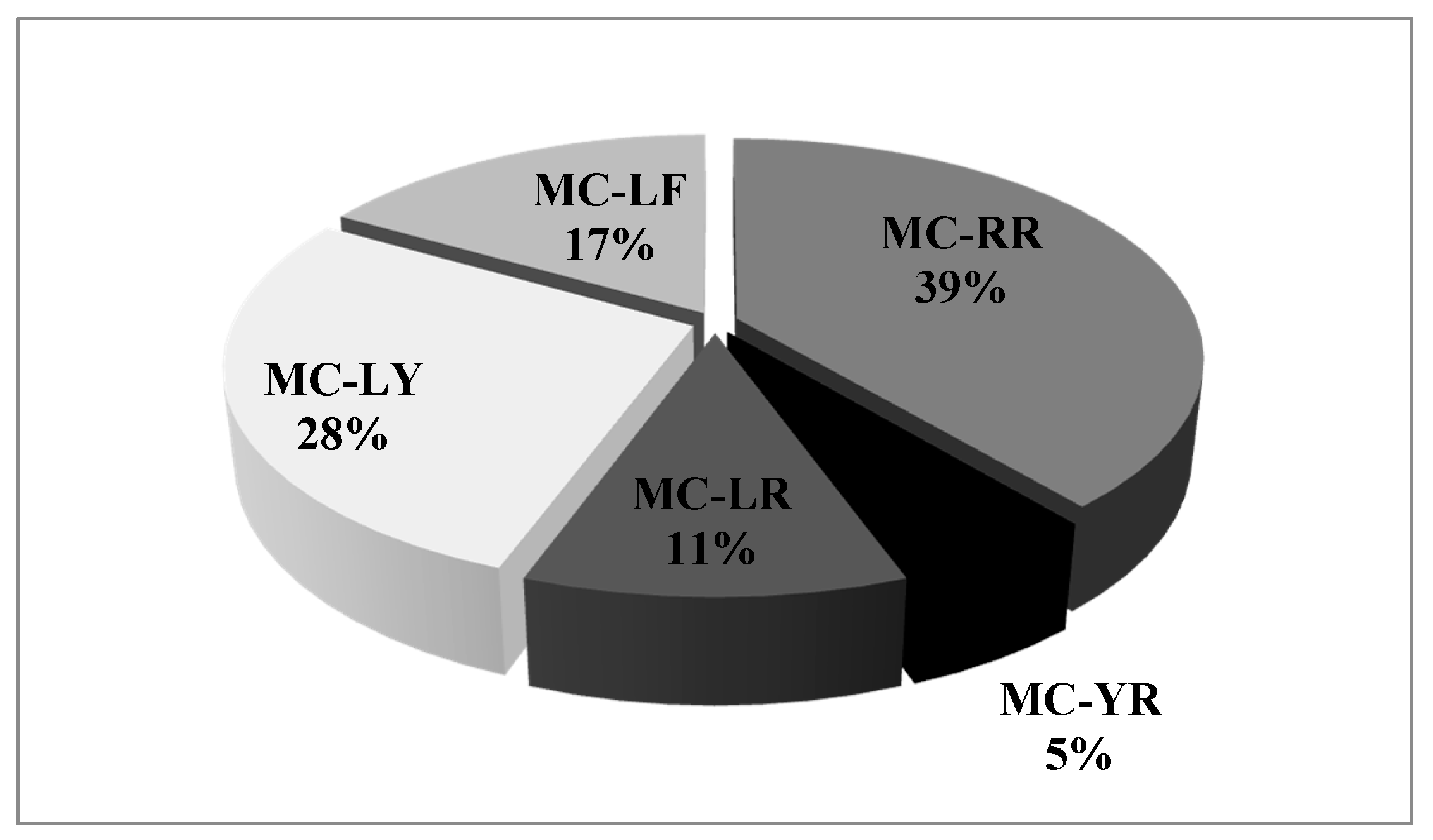
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Area
4.2. Culturing
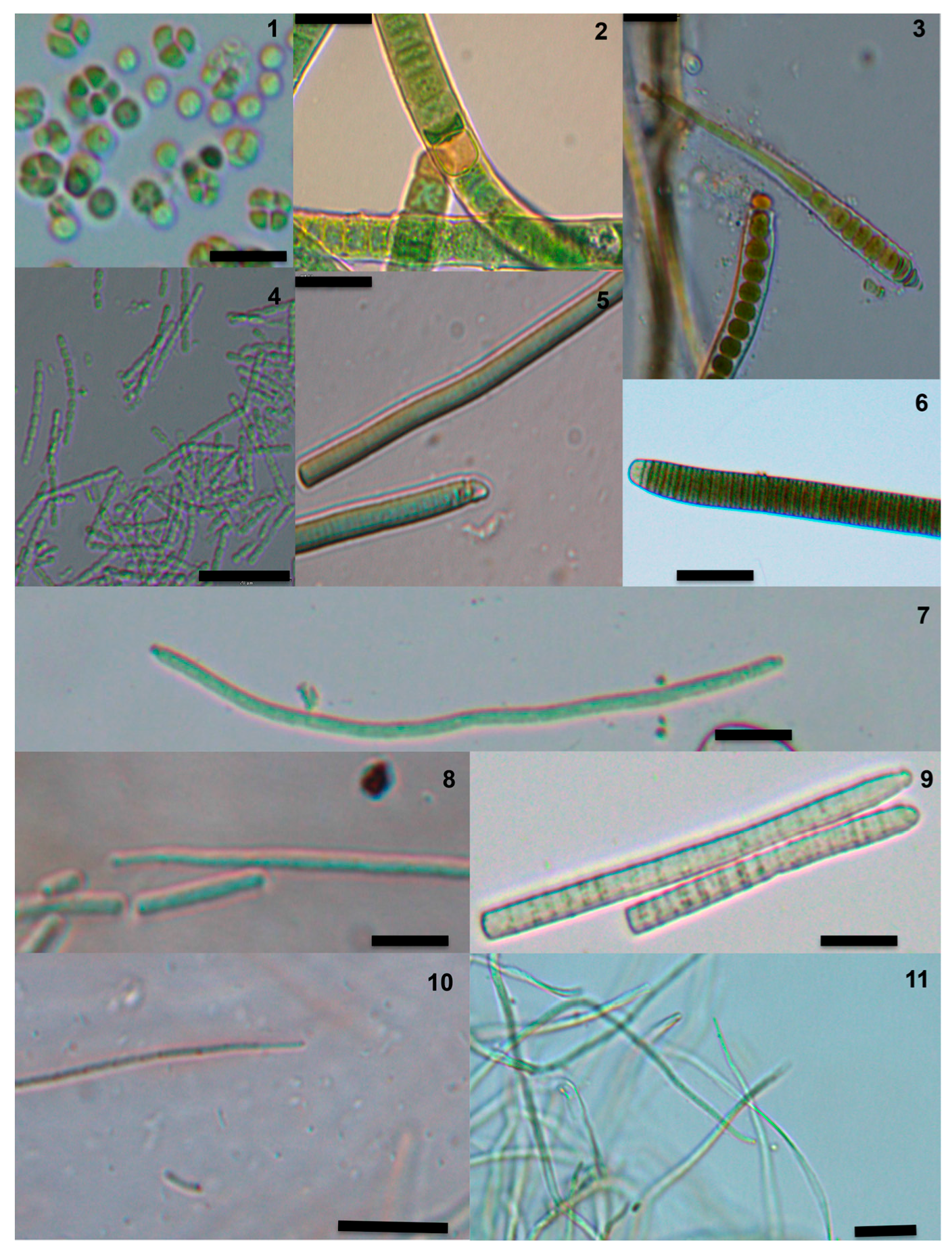
4.3. Taxonomic Identification
4.4. Microcystin Extraction and Quantification
4.5. Chemicals
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Mez, K.; Beattie, K.; Codd, G.A.; Hanselmann, K.; Hauser, B.; Naegeli, H.; Preisig, H. Identification of a microcystin in benthic cyanobacteria linked to cattle deaths on alpine pastures in Switzerland. Eur. J. Phycol. 1997, 32, 111–117. [Google Scholar] [CrossRef]
- Aboal, M.; Puig, M.A. Intracellular and dissolved microcystin in reservoirs of the river Segura basin, Murcia, SE Spain. Toxicon 2005, 45, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Aboal, M.; Puig, M.A.; Asencio, A.D. Production of microcystins in calcareous Mediterranean streams: The Alharabe River, Segura River basin in south-east Spain. J. Appl. Phycol. 2005, 17, 231–243. [Google Scholar] [CrossRef]
- Izaguirre, G.; Jungblut, A.D.; Neilan, B.A. Benthic cyanobacteria (Oscillatoriaceae) that produce microcystin-LR, isolated from four reservoirs in southern California. Water Res. 2007, 41, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, Z.A.; El-Sharouny, H.M.; Ali, W.S.M. Microcystin production in benthic mats of cyanobacteria in the Nile River and irrigation canals, Egypt. Toxicon 2006, 47, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Oudra, B.; Dadi-El Andaloussi, M.; Vasconcelos, V.M. Identification and quantification of microcystins from Nostoc muscorum bloom occurring in Oukaïmeden River (High-Atlas mountains of Marrakech, Morocco). Environ. Monit. Asess. 2009, 149, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Douma, M.; Loudiki, M.; Oudra, B.; Mouhri, K.; Ouahid, Y.; del Campo, F. Taxonomic diversity and toxicological assessment of Cyanobacteria in Moroccan inland waters. Revue des Sciences de l’eau 2017, 223, 435–449. [Google Scholar] [CrossRef]
- Wood, S.A.; Heath, M.W.; Holland, P.T.; Munday, R.; McGregor, G.B.; Ryan, K.G. Identification of a benthic microcystin-producing filamentous cyanobacterium (Oscillatoriales) associated with a dog poisoning in New Zealand. Toxicon 2010, 55, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Aboal, M. Benthic microcystin and climatic change. In Stress Biology of Cyanobacteria. Molecular Mechanisms to Cellular Responses; Srivastava, A.K., Rai, A.N., Neilan, B., Eds.; CRC Press: Boca Raton, FL, USA, 2013; Chapter 17; pp. 321–340. [Google Scholar]
- Fetscher, A.E.; Howard, M.D.A.; Stancheva, R.; Kudela, R.M.; Stein, E.D.; Sutula, M.A.; Busse, L.B.; Sheath, R.G. Wadeable streams as widespread sources of benthic cyanotoxins in California, USA. Harmful Algae 2015, 49, 105–116. [Google Scholar] [CrossRef]
- Oksanen, I.; Jokela, J.; Fewer, D.P.; Wahlsten, M.; Rikkinen, J.; Sivonen, K. Discovery of rare and highly toxic microcystins from lichen-associated cyanobacterium Nostoc sp. strain IO-102-I. Appl. Environ. Microbiol. 2004, 70, 5756–5763. [Google Scholar] [CrossRef] [PubMed]
- Kaasalainen, U.; Fewer, D.P.; Jokela, J.; Wahlsten, M.; Sivonen, K.; Rikkinen, J. Cyanobacteria produce a high variety of hepatotoxic peptides in lichen symbiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 5886–5891. [Google Scholar] [CrossRef] [PubMed]
- Kurmayer, R. The toxic cyanobacterium Nostoc sp. strain 152 produces highest amounts of microcystin and nostophycin under stress conditions. J. Phycol. 2010, 47, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, H.; Yamaguchi, Y.; Sakaki, S.; Watarai, K.; Tanaka, N.; Hori, M.; Seki, H.; Tsuchida, M.; Yamada, A.; Nishimori, T.; et al. Safety evaluation of Nostoc flagelliforme (Nostocales, Cyanophyceae) as a potential food. Food Chem. Toxicol. 1998, 36, 1073–1077. [Google Scholar] [CrossRef]
- Van Apeldoorn, M.E.; van Egmond, H.P.; Speijers, G.J.A.; Bakker, G.J.I. Toxins of cyanobacteria. Mol. Nutr. Food Res. 2007, 51, 7–60. [Google Scholar] [CrossRef] [PubMed]
- Borges, H.L.F.; Branco, L.H.Z.; Martins, M.D.; Lima, C.S.; Barbosa, P.T.; Lira, G.A.S.T.; Bittencourt-Oliveira, M.C.; Molica, R.J.R. Cyanotoxin production and phylogeny of benthic cyanobacterial strains isolated from the northeast of Brazil. Harmful Algae 2015, 43, 46–57. [Google Scholar] [CrossRef]
- Omidi, A.; Esterhuizen-Londt, M.; Pflugmacher, S. Still challenging the ecological function of the cyanobacterial toxin microcystin- What we know so far. Toxin Rev. 2017. [Google Scholar] [CrossRef]
- Ward, C.J.; Beattie, K.A.; Lee, E.Y.C.; Codd, G.A. Colorimetric protein phosphatase inhibition assay of laboratory strains and natural blooms of cyanobacteria: Comparisons with high-performance liquid chromatographic analysis of microcystins. FEMS Microbiol. Lett. 1997, 153, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y. Serine/Threonine phosphatases: Mechanism through structure. Cell 2009, 139, 468–484. [Google Scholar] [CrossRef] [PubMed]
- Valdor, R.; Aboal, M. Effects of living cyanobacteria, cyanobacterial extracts and pure microcystins on growth and ultrastructure of microalgae and bacteria. Toxicon 2006, 49, 769–779. [Google Scholar] [CrossRef] [PubMed]
- García-Espín, L.; Cantoral, E.A.; Asencio, A.D.; Aboal, M. Microcystins and cyanophyte extracts inhibit or promote the photosynthesis of fluvial algae. Ecological and management implications. Ecotoxicology 2017, 26, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Vesterkvist, P.S.M.; Misiorek, J.O.; Spoof, L.E.M.; Toivola, D.M.; Meriluoto, J.A.O. Comparative cellular toxicity of hydrophilic and hydrophobic microcystins on Caco-2 cells. Toxins 2012, 4, 1008–1023. [Google Scholar] [CrossRef] [PubMed]
- Liarte, S.; Ubero-Pascal, N.; García-Ayala, A.; Puig, M.A. Histological effects and localization of dissolved microcystins LR and LW in the mayfly Ecdyonurus angelieri Thomas (Insecta, Ephemeroptera). Toxicon 2014, 92, 31–35. [Google Scholar] [CrossRef] [PubMed]
- El Ghazali, I.; Saqrane, S.; Carvalho, A.P.; Ouahid, Y.; del Campo, F.; Oudra, B.; Vasconcelos, V. Effect of different microcystin profiles on toxin bioaccumulation in common carp (Cyprinus carpio) larvae via Artemia nauplii. Ecotox. Environ. Saf. 2010, 73, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Trinchet, I.; Djediat, C.; Huet, H.; Puiseux Dao, S.; Edery, M. Pathological modifications following sub-chronic exposure of medaka fish (Oryzias latipes) to microcystin-LR. Reprod. Toxicol. 2011, 32, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Drobac, D.; Tokodi, N.; Lujic, J.; Matinovic, Z.; Subakov-Simic, G.; Dulic, T.; Vazic, T.; Nybom, S.; Meriluoto, J.; Codd, G.A.; et al. Cyanobacteria and cyanotoxins in fishponds and their effects on fish tissue. Harmful Algae 2016, 55, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E.J.; Lürling, M. Occurrence of the microcystins MC-LW and MC-LF in Dutch surface waters and their contribution to total microcystin toxicity. Mar. Drugs 2013, 11, 2643–2654. [Google Scholar] [CrossRef] [PubMed]
- Paldavicienè, A.; Mazur-Marzec, H.; Razinkovas, A. Toxic cyanobacteria blooms in the Lithuanian part of the Curonian Lagoon. Oceanologia 2009, 51, 203–216. [Google Scholar] [CrossRef]
- Bittencourt-Oliveira, M.C.; Oliveira, M.C.; Pinto, E. Diversity of microcystin-producing genotypes in Brazilian strains of Microcystis (cyanobacteria). Braz. J. Biol. 2011, 71, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Hindle, R.; Zhang, X.; Kinniburgh, D. Identification of Unknown Microcystins in Alberta Lake Water; Agilent Technologies: Santa Clara, CA, USA, 2014. [Google Scholar]
- Beversdorf, L.J.; Chaston, S.D.; Miller, T.R.; McMahon, K.D. Microcystin mcyA and mcyE gene abundances are not appropriate indicators of microcystin concentrations in lakes. PLoS ONE 2015, 10, e0125353. [Google Scholar] [CrossRef] [PubMed]
- Edwards, C.; Beattie, K.A.; Scrimgeour, C.M.; Codd, G.A. Identification of anatoxin-A in benthic cyanobacteria (blue-green algae) and in associated dog poisonings at Loch Insh, Scotland. Toxicon 1992, 30, 1165–1175. [Google Scholar] [CrossRef]
- Gugger, M.; Lenoir, S.; Berger, C.; Ledreux, A.; Druaf, J.C.; Humbert, J.F.; Guette, C.; Bernard, C. First report in a river in France of the benthic cyanobacterium Phormidium favosum producing anatoxin-a associated with dog neurotoxicosis. Toxicon 2005, 45, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, D.; Moreno, E.; Paniagua, T.; de Hoyos, C.; Wormer, L.; Sanchís, D.; Cirés, S.; Martín-del-Pozo, D.; Codd, G.A.; Quesada, A. Anatoxin-a occurrence and potential cyanobacterial anatoxin-a producers in Spanish reservoirs. J. Phycol. 2007, 43, 1120–1125. [Google Scholar] [CrossRef]
- Park, H.D.; Watanabe, M.F.; Harda, K.; Nagal, H.; Suzuki, M.; Watanabe, M.; Hayashi, H. Hepatotoxin (microcystin) and neurotoxin (anatoxin-a) contained in natural blooms and strains of cyanobacteria from Japanese freshwaters. Nat. Toxins 1993, 1, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Chrapusta, E.; Wegrzyn, M.; Zabagio, K.; Kaminski, A.; Adamski, M.; Wietrzyk, P.; Bialczyk, J. Microcystins and anatoxin-a in Arctic biocrust cyanobacterial communities. Toxicon 2015, 101, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Bartram, J. (Eds.) Toxic Cyanobacteria in Water. A Guide to Their Public Health Consequences, Monitoring and Management; E & FN Spon: London, UK, 2001; pp. 15–40. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Codd, G.A. Cyanotoxins. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 651–670. [Google Scholar]
- Rantala, A.; Fewer, D.P.; Hisbergues, M.; Rouhiainen, L.; Vaitomaa, J.; Börner, T. Phylogenetic evidence for the early evolution of microcystin synthesis. Proc. Natl. Acad. Sci. USA 2004, 101, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Botana, L.M.; Louzao, C.; Vilariño, N. (Eds.) Climate Change and Marine and Freshwater Toxins; De Greyter: Berlin, Germany, 2015. [Google Scholar]
- Codd, G.A.; Bell, S.G.; Kaya, K.; Ward, C.J.; Beattie, K.A.; Metcalf, J.S. Cyanobacterial toxins, exposure routes and human health. Eur. J. Phycol. 1999, 34, 405–415. [Google Scholar] [CrossRef]
- Boopathi, T.; Ki, J.-S. Impact of environmental factors on the regulation of cyanotoxin production. Toxins 2014, 6, 1951–1978. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Van Oosterhout, E.; Faassen, E.J.; Lürling, M.; Helmsing, N.R.; van de Waal, D.B. Elevated pCO2 causes a shift towards more toxic microcystin variants in nitrogen-limited Microcystis aeruginosa. FEMS Microb. Ecol. 2016, 92. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Hoeger, S.J.; Stemmer, K.; Feurstein, D.J.; Knobeloch, D.; Nussler, A.; Dietrich, D.R. The role of organic transporting polypeptides (OATPs/SLCOs) in the toxicity of different congeners in vitro: A comparison of primary human hepatocytes and OATP-transfected HEK 293 cells. Toxicol. Appl. Pharmacol. 2010, 245, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Vichi, S.; Lavarini, P.; Funari, E.; Scardala, S.; Testai, E. Contamination by Microcystis and microcystins of blue-green algae food supplements (BGAS) on the Italian market and possible risk for the exposed population. Food Chem. Toxicol. 2012, 50, 4493–4499. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Rodriguez, R.; Jin, Z.; Harvie, J.; Cabecinha, A. Evaluation of three field test kits to detect microcystins from public health perspective. Harmful Algae 2015, 42, 34–42. [Google Scholar] [CrossRef]
- Moore, C.E.; Juan, J.; Lin, Y.; Gaskill, C.L.; Puschner, B. Comparison of protein phosphatase inhibition assay with LC-MS/MS for diagnosis of microcystin toxicosis in veterinary cases. Mar. Drugs 2016, 14, 54. [Google Scholar] [CrossRef] [PubMed]
- Meriluoto, J.; Codd, G.A. (Eds.) Toxic Cyanobacterial Monitoring and Cyanotoxin Analysis; Åbo Akademis Förlag—Åbo Akademi University Press: Åbo, Finland, 2005. [Google Scholar]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Kaufononga, S.A.F.; Cary, S.C.; Hamilton, D.P. High levels of structural diversity observed in microcystins from Microcystis CAWBG11 and characterization of six new microcystin congeners. Mar. Drugs 2014, 12, 5372–5395. [Google Scholar] [CrossRef] [PubMed]
- Quiblier, C.; Wood, S.; Echenique-Subiabre, I.; Heath, M.; Villeneuve, A.; Humbert, J.F. A review of current knowledge on toxic benthic freshwater cyanobacteria—Ecology, toxin production and risk management. Water Res. 2013, 47, 5464–5479. [Google Scholar] [CrossRef]
- Wood, S.A.; Maier, M.Y.; Puddick, J.; Pochon, X.; Zaiko, A.; Dietrich, D.R.; Hamilton, D.P. Trophic state and geographic gradients influence planktonic cyanobacterial diversity and distribution in New Zealand lakes. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [PubMed]
- Barco, M.; Lawton, L.A.; Rivera, J.; Caixach, J. Optimization of intracellular microcystin extraction for the subsequent analysis by high-performance liquid chromatography. J. Chromatogr. A 2005, 1074, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota-1. Teil: Chroococcales. In Süsswasserflora von Mitteleuropa 19/1; Ettl, H., Gärtner, G., Heynig, H., Mollenhauer, D., Eds.; Elsevier/Spektrum: Heidelberg, Germany, 1998; p. 548. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota-2. Teil: Oscillatoriales. In Süsswasserflora von Mitteleuropa 19/2; Büdel, B., Krienitz, L., Gärtner, G., Schagerl, M., Eds.; Elsevier/Spektrum: Heidelberg, Germany, 2007; p. 759. [Google Scholar]
- Komárek, J. Cyanoprokaryota-3. Heterocystous genera. In Süsswasserflora von Mitteleuropa 19/3; Büdel, B., Krienitz, L., Gärtner, G., Schagerl, M., Eds.; Elsevier/Spektrum: Heidelberg, Germany, 2013; p. 1087. [Google Scholar]
- Gjolme, N.; Utkilen, H. The extraction and the stability of microcystin-RR in different solvents. Phycologia 1996, 35, 80–82. [Google Scholar] [CrossRef]
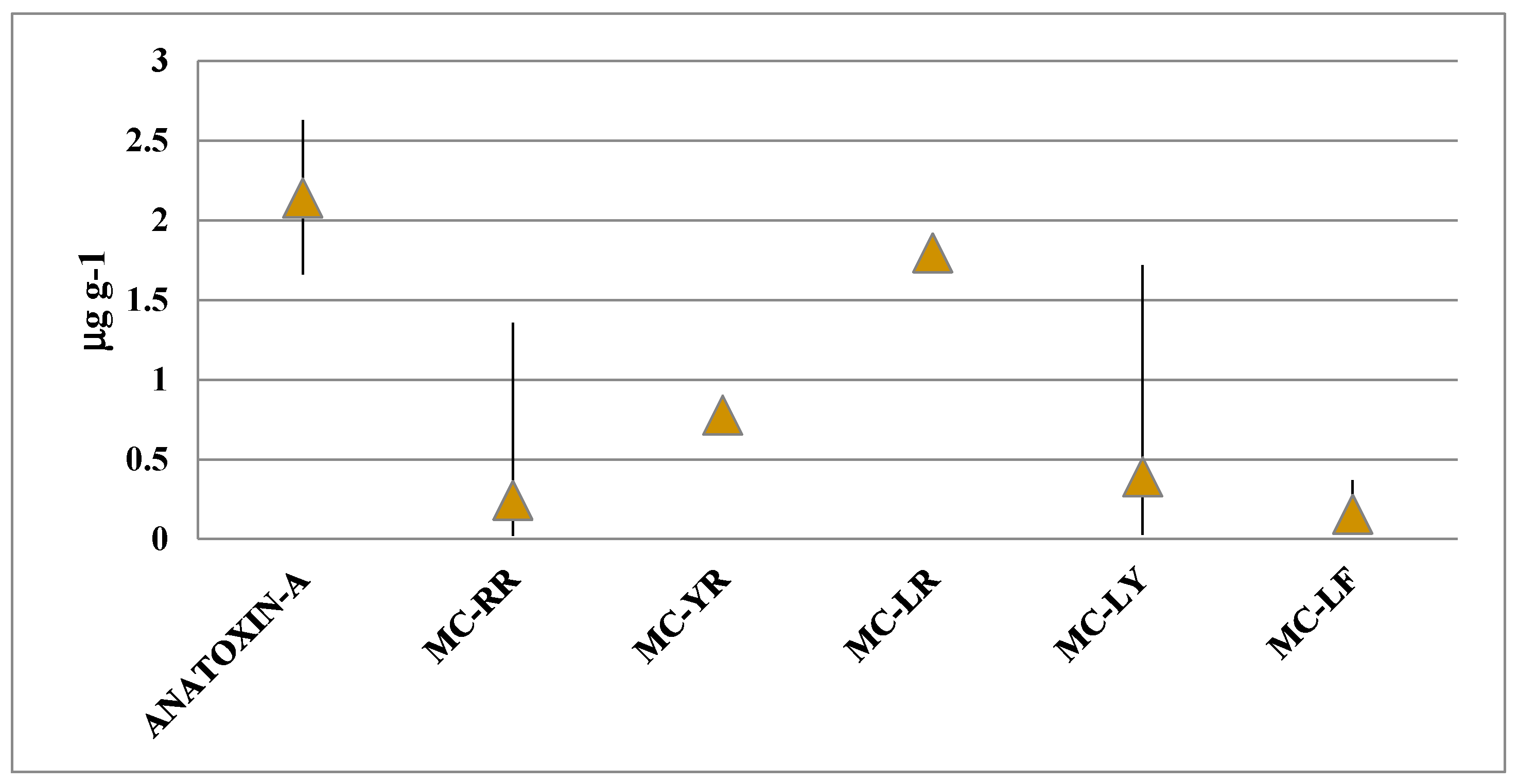

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Localities | Habitat | Lithology | Altitude(m) | Coordinates | Rainfall(mm) | Mean Temperature (°C) |
|---|---|---|---|---|---|---|
| Prado Redondo, Sierra Nevada, Granada | Creek | Granitic | 2100 | 37°05′23.0″ N 3°24′56.8″ W | 1322 | 7.8 |
| San Juan, Sierra Nevada, Granada | Creek | Granitic | 2500 | 37°05′16.7″ N 3°22′18.5″ W | 1322 | 7.8 |
| Poqueira, Sierra Nevada, Granada | Creek | Granitic | 1540 | 36°59′26.8″ N 3°21′00.2″ W | 935 | 11.6 |
| Vall de Mulleres, Vielha, Lleida | Peat bog | Granitic | 1609 | 42°37′40.6″ N 0°45′34.9″ E | 1843 | 8.5 |
| Riu Escrita, Lleida | Stream | Granitic | 1700 | 42°34′38″ N 0°56′52″ E | 1100 | 5.0 |
| Fonts Lac S. Maurici, Lleida | Springs | Granitic | 1910 | 42°32′28.9″ N 9°01′21.6″ W | 1100 | 5.0 |
| Riu Llebreta, Lleida | Stream | Granitic | 1999 | 42°34′38″ N 0°56′52″ E | 1100 | 5.0 |
| Riu Ter, Villalonga, Girona | River | Granitic | 1067 | 42°19′59″ N 2°18′47″ E | 933 | 9.8 |
| Lagoa Carregal, Corrubedo, A Coruña | Lagoon | Granitic | 5 | 42°33′002″ N 9°02′00″ W | 933 | 9.8 |
| Lagoa Vixán, Corrubedo, A Coruña | Lagoon | Granitic | 5 | 42°33′002″N 9°02′00″W | 1014 | 14.8 |
| Guayadeque, Gran Canaria | Stream | Volcanic | 1273 | 27°56′00.7″ N 15°28′57.7″ W | 175 | 22.5 |
| Palacio Guevara, Lorca, Murcia | Building | Marble | 353 | 37°40′29.82″ N 1°41′51.54″ W | 232 | 17.6 |
| Río Alhárabe, Moratalla, Murcia | Stream | Calcareous | 900 | 38°12′50″ N 1°57′46″ W | 522 | 15.7 |
| Azud de Ojós, Ojós, Murcia | Reservoir | Calcareous | 132 | 38°8′ 52″ N 1°20′32″ W | 306 | 17.4 |
| Cueva de los Grajos, Cieza, Murcia | Cave | Calcareous | 250 | 38°14′21″ N 1°25′08″ W | 307 | 17.2 |
| Río Chícamo, Abanilla, Murcia | Stream | Calcareous | 290 | 38º24′97″ N 1º00′18″ W | <200 | 10.0 |
| Marjal Almenara, Almenara, Castellón | Freshwater Spring | Calcareous | 26 | 39°44′54.1″ N 0°11′17.3″ W | 467 | 17.5 |
| Marjal de Pego-Oliva, Valencia | Saline Spring | Calcareous | 5 | 38° 52′08.2″ N 0°02′57.92″ W | 637 | 17.8 |
| Río Amadorio, Vilajoyosa, Alicante | Stream | Calcareous | 32 | 38°30′19″ N 0°13′58″ W | 300 | 18.0 |
| Río Algar, Callosa, Alicante | Stream | Calcareous | 247 | 38°39′33.67″ N 0°5′45.58″ W | 519 | 16.9 |
| Taxa | Order | Habitat | Locality |
|---|---|---|---|
| Hyella balani Lehman | Chroococcales | Euendolithic, spring | Marjal Oliva-Pego, Valencia |
| Pseudocapsa dubia Ercegovic | Chroococcales | Chasmoendolithic | Palacio Guevara, Lorca, Murcia |
| Gloeotrichia natans (Hedwig) Rabenhorst ex Bornet et Flahault | Nostocales | Epiphytic, lagoon | Lagoa Vixán, Corrubedo, A Coruña |
| Nostochopsis lobata Wood ex Bornet & Flahault | Nostocales | Epiphytic, lagoon | Lagoa Vixán, Corrubedo, A Coruña |
| Rivularia biasolettiana (Meneghini ex Bornet & Flahault | Nostocales | Epilithic, stream | Río Alhárabe, Moratalla, Murcia Río Chícamo, Abanilla, Murcia |
| Scytonema drilosiphon Elenkin & V. I. Polyansky | Nostocales | Epilithic, cave | Cueva Grajos, Cieza, Murcia |
| Geitlerinema splendidum (Greville ex Gomont) Anagnostidis | Oscillatoriales | Epilithic, spring | Ullal Almenara, Castellón |
| Homoeothrix juliana (Bornet & Flahault ex Gomont) Kirchner | Oscillatoriales | Epilithic, stream | Río Amadorio, Alicante |
| Leptolyngbya subtilis (West) Anagnostidis | Oscillatoriales | Epilithic, spring | Ullal Almenara, Castellón |
| Leptolygnbya truncata (Lemmermann) Anagnostidis & Komárek | Oscillatoriales | Epilithic, stream | Río Amadorio, Alicante |
| Oscillatoria margaritifera Kützing ex Gomont | Oscillatoriales | Epilithic, spring | Ullal Almenara, Castellón |
| Oscillatoria sancta Kützing ex Gomont | Oscillatoriales | Epilithic, stream | Río Ter, Vilallonga, Lérida |
| Geitlerinema carotinum (Geitler) Anagnostidis | Oscillatoriales | Epipelic, reservoir | Azud Ojós, Ojós, Murcia |
| Phormidium autumnale (Agardh) Trevisan ex Gomont | Oscillatoriales | Epilithic, stream | Río Alhárabe, Moratalla, Murcia |
| Pseudanabaena frigida (Fritsch) Anagnostidis | Oscillatoriales | Epipelic, peat bog | Vall de Molleres, Viella, Lérida |
| Schizothrix rivularianum Voronichin | Oscillatoriales | Epipelic, stream | Río Alhárabe, Moratalla, Murcia |
| Phormidium uncinatum Gomont ex Gomont | Oscillatoriales | Epipelic, stream | Barranco Guayadeque, Gran Canaria |
| Taxa | Locality | Microcystins | Anatoxins |
|---|---|---|---|
| Gloeotrichia natans | Corrubedo, Galicia | MC-LF, MC-RR | nd |
| Geitlerinema carotinum | Ojós, Murcia | MC-LY | ANT-a |
| Geitlerinema splendidum | Almenara, Valencia | MC-LF, MC-RR | ANT-a |
| Nostoc cf. commune | Sierra Nevada, Granada | MC-LF, MC-LY | nd |
| Oscillatoria margaritifera | Almenara, Valencia | MC-LF, MC-LY, MC-RR | nd |
| Phormidium autumnale | Moratalla, Murcia | MC-LR | nd |
| Phormidium uncinatum | Guayadeque, Gran Canaria | MC-LF, MC-LY | ANT-a |
| Phormidium sp. | Sierra Nevada, Granada | MC-LF, MC-LY | nd |
| Pseudoanabaena frigida | Vall de Mulleres, Lérida | MC-RR | nd |
| Pseudocapsa dubia | Palacio de Guevara, Murcia | MC-RR, MC-YR | nd |
| Rivularia biasolettiana | Río Chícamo, Murcia | MC-LR, MC-LY, MC-RR | nd |
| Schizothrix rivularianum | Río Alhárabe, Murcia | MC-LY, MC-RR | nd |
| Scytonema drilosiphon | Cueva de los Grajos, Murcia | MC-LY | nd |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cantoral Uriza, E.A.; Asencio, A.D.; Aboal, M. Are We Underestimating Benthic Cyanotoxins? Extensive Sampling Results from Spain. Toxins 2017, 9, 385. https://doi.org/10.3390/toxins9120385
Cantoral Uriza EA, Asencio AD, Aboal M. Are We Underestimating Benthic Cyanotoxins? Extensive Sampling Results from Spain. Toxins. 2017; 9(12):385. https://doi.org/10.3390/toxins9120385
Chicago/Turabian StyleCantoral Uriza, Enrique A., Antonia D. Asencio, and Marina Aboal. 2017. "Are We Underestimating Benthic Cyanotoxins? Extensive Sampling Results from Spain" Toxins 9, no. 12: 385. https://doi.org/10.3390/toxins9120385
APA StyleCantoral Uriza, E. A., Asencio, A. D., & Aboal, M. (2017). Are We Underestimating Benthic Cyanotoxins? Extensive Sampling Results from Spain. Toxins, 9(12), 385. https://doi.org/10.3390/toxins9120385