
Experimental Assays to Assess the Efficacy of Vinegar and Other Topical First-Aid Approaches on Cubozoan (Alatina alata) Tentacle Firing and Venom Toxicity

 , ,
, ,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Model Validation
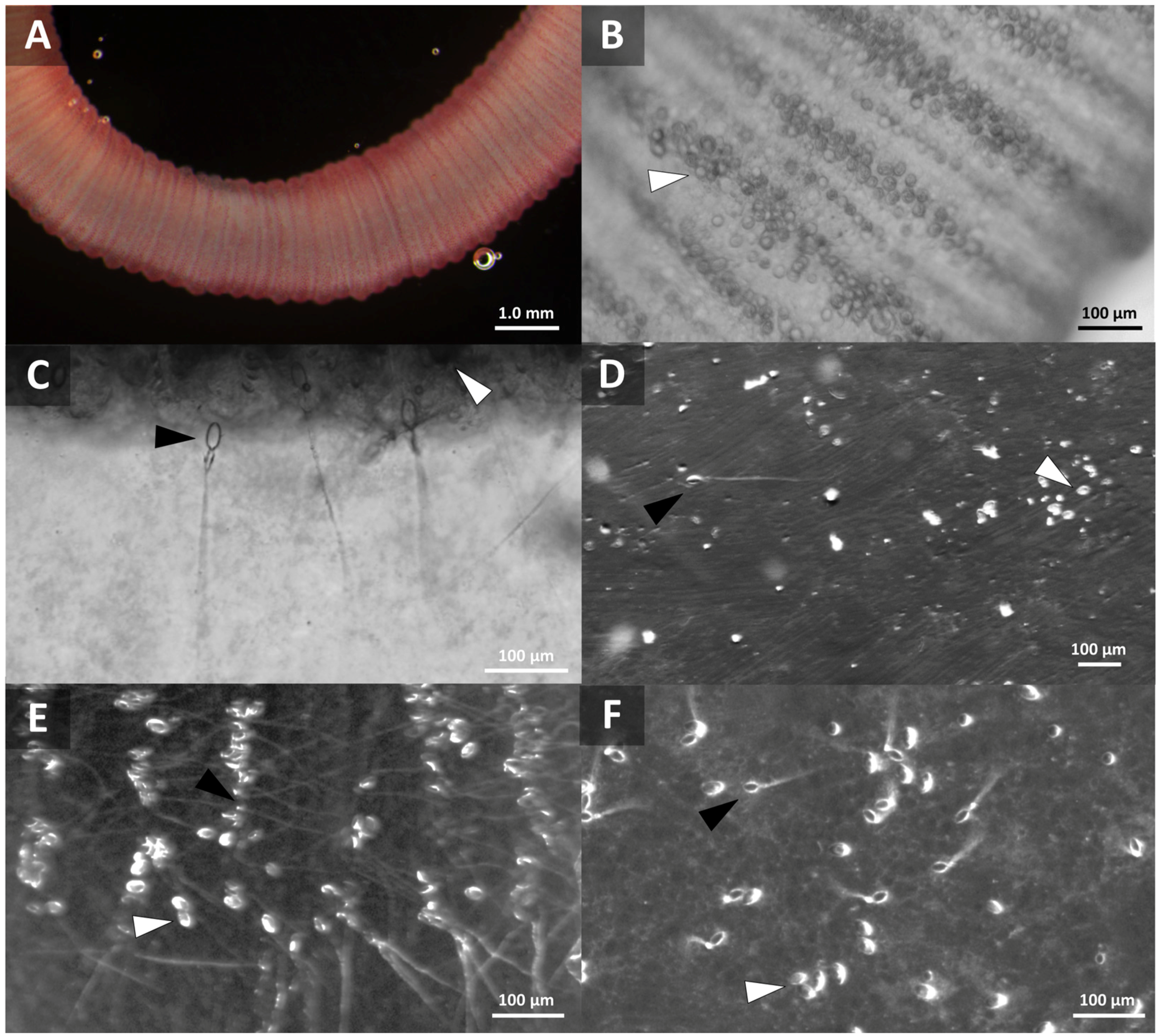
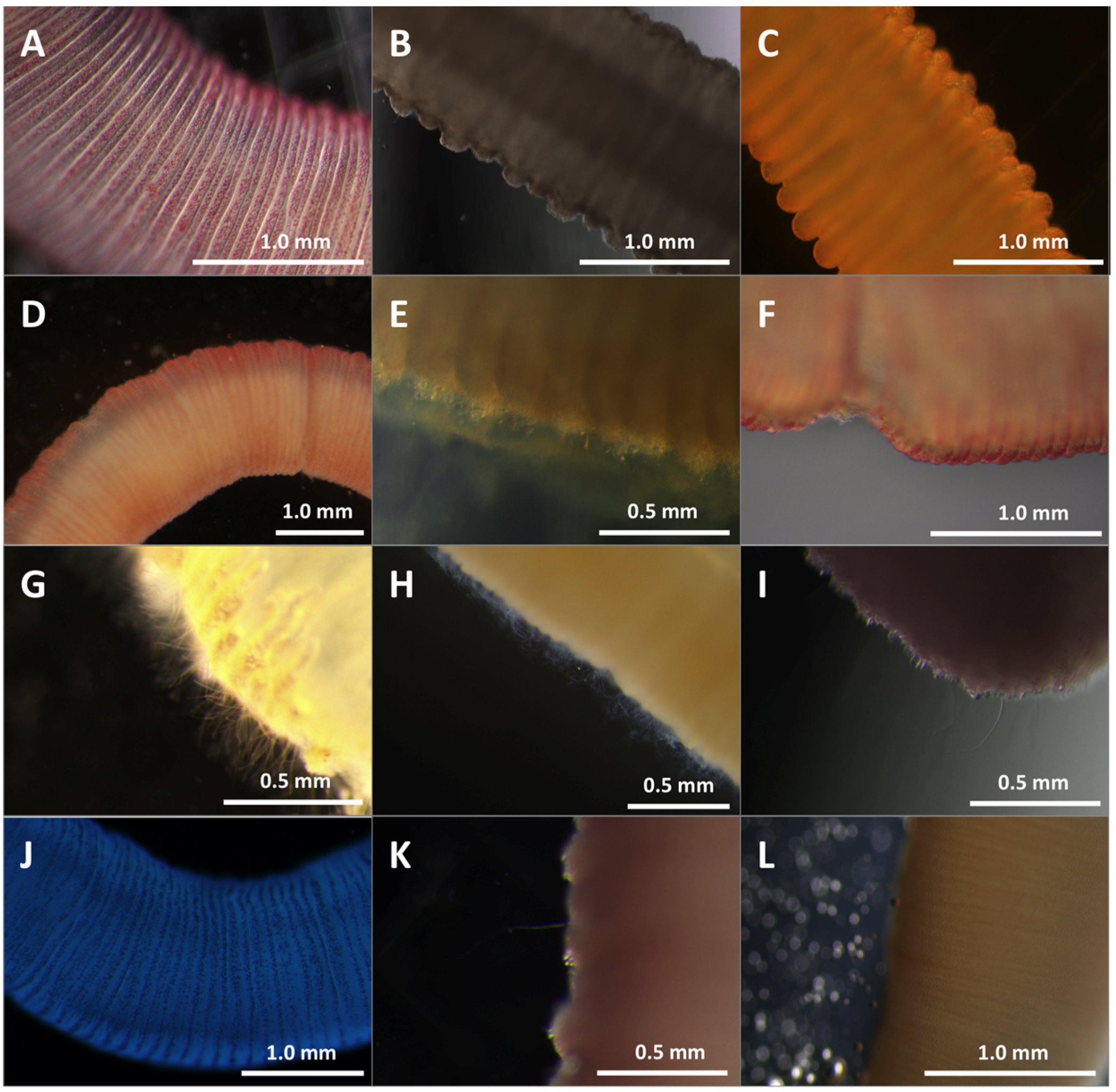
2.1.1. Live Tentacles Spontaneously Sting and Imaging
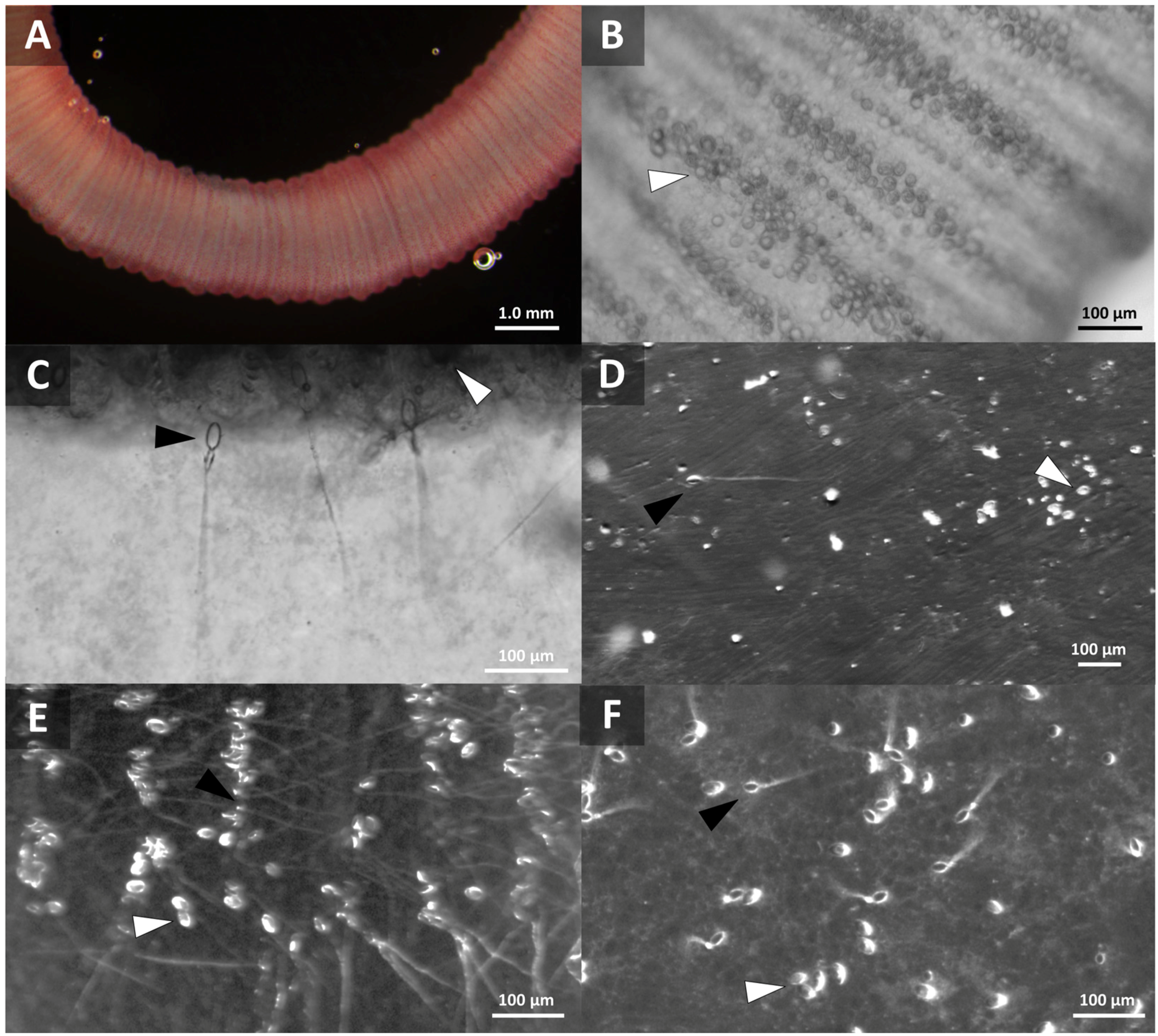
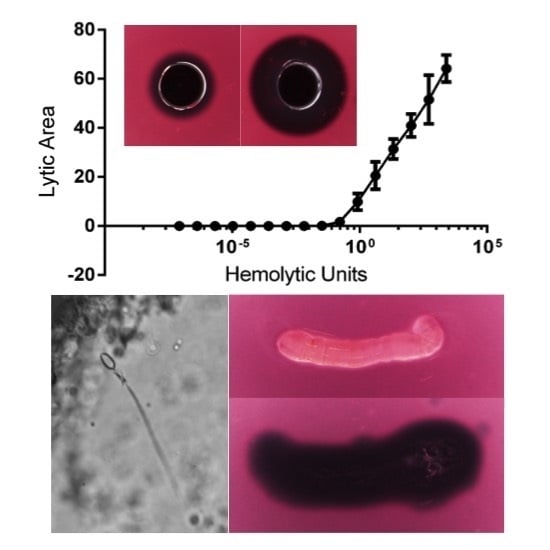
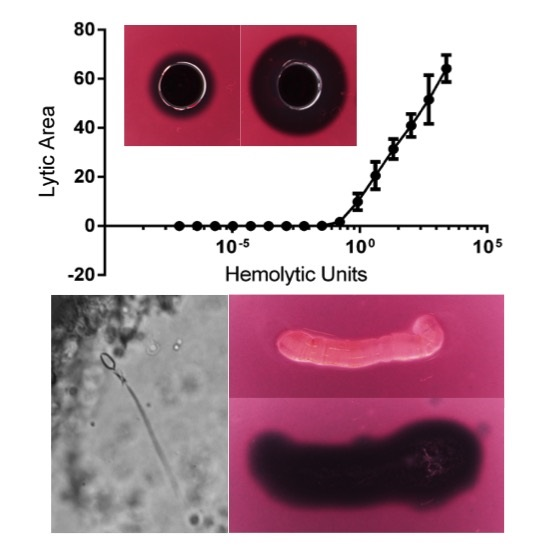
2.1.2. Quantification of Cnidae Discharge

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sting Substrate | Tentacle Cnidae/mm2 | Cnidae Adherent (Total per mm2) | Cnidae Discharged (Total per mm2) | Percent of Tentacle Cnidae Discharged |
|---|---|---|---|---|
| Human Skin | 1055–1355 | 2.6–25.3 | 0.4–4.5 | 0.03%–0.43% |
| 1200 ± 68 | 9.3 ± 1.8 | 1.5 ± 1.0 | 0.21% ± 0.10% | |
| TBAA | 804–1499 | 4.5–42.2 | 1.0–6.3 | 0.07%–0.46% |
| 1211 ± 97 | 14.0 ± 4.2 | 3.3 ± 0.7 | 0.24% ± 0.05% |
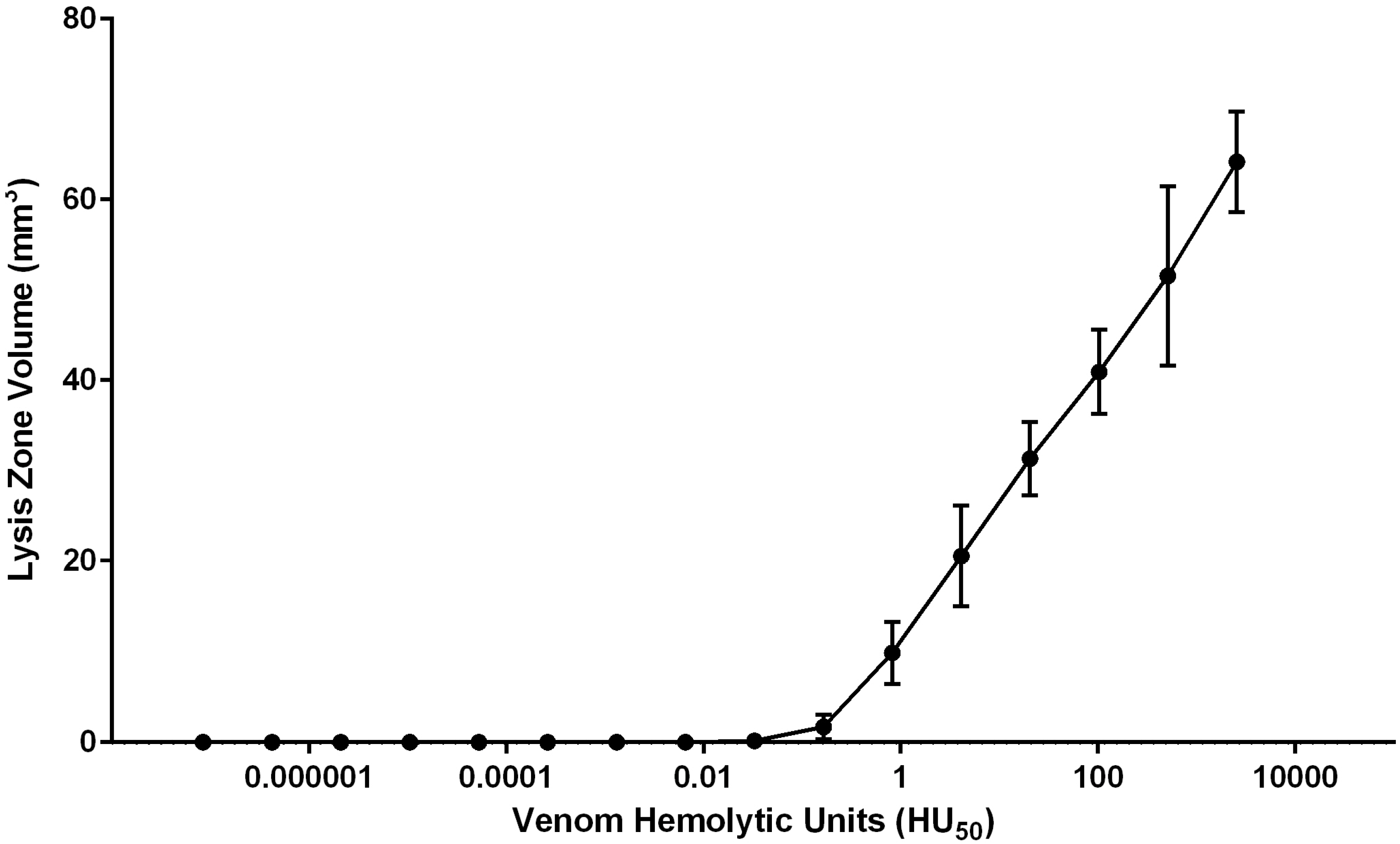
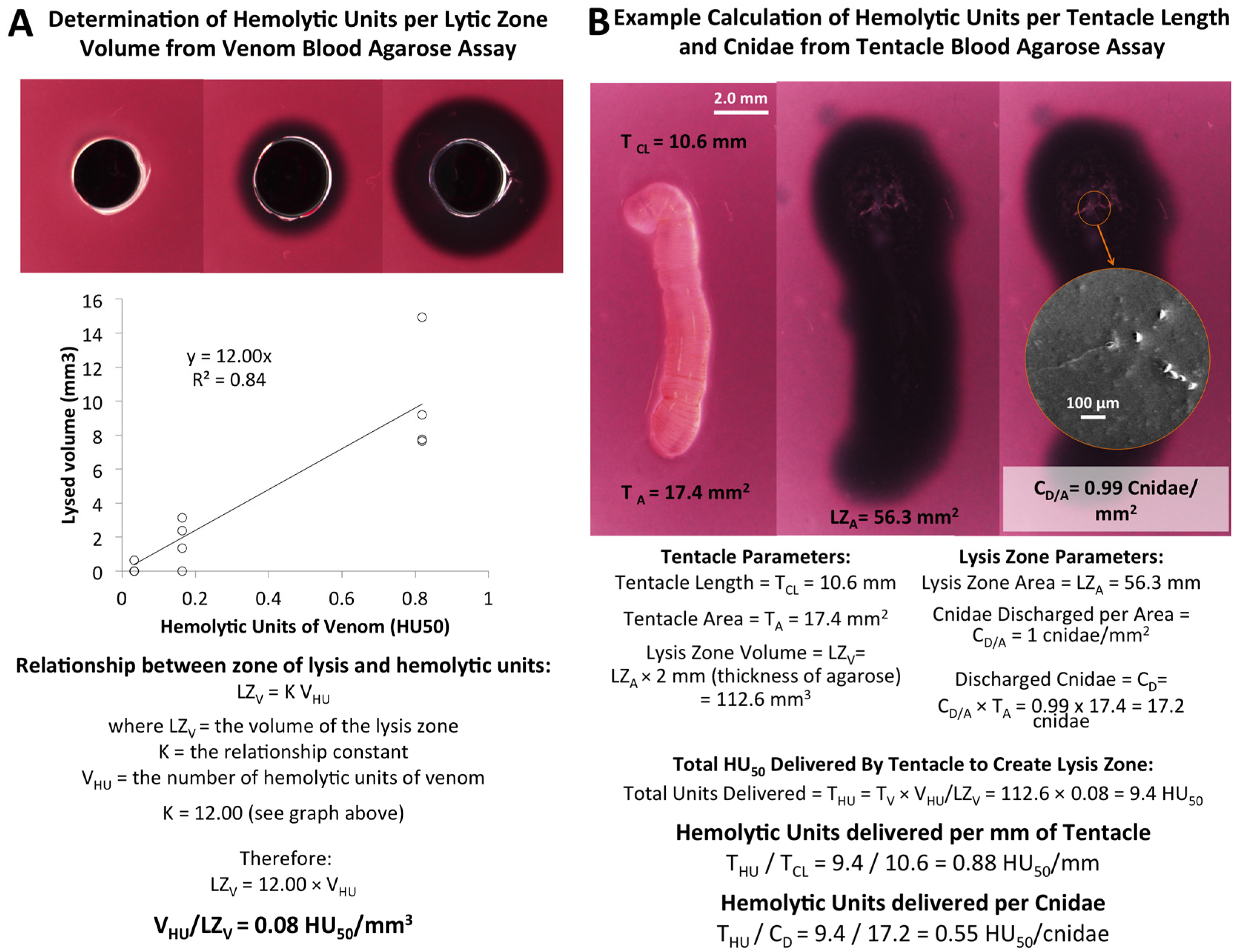
2.1.3. Quantification of Envenomation Triggered Hemolytic Activity in Blood Agarose
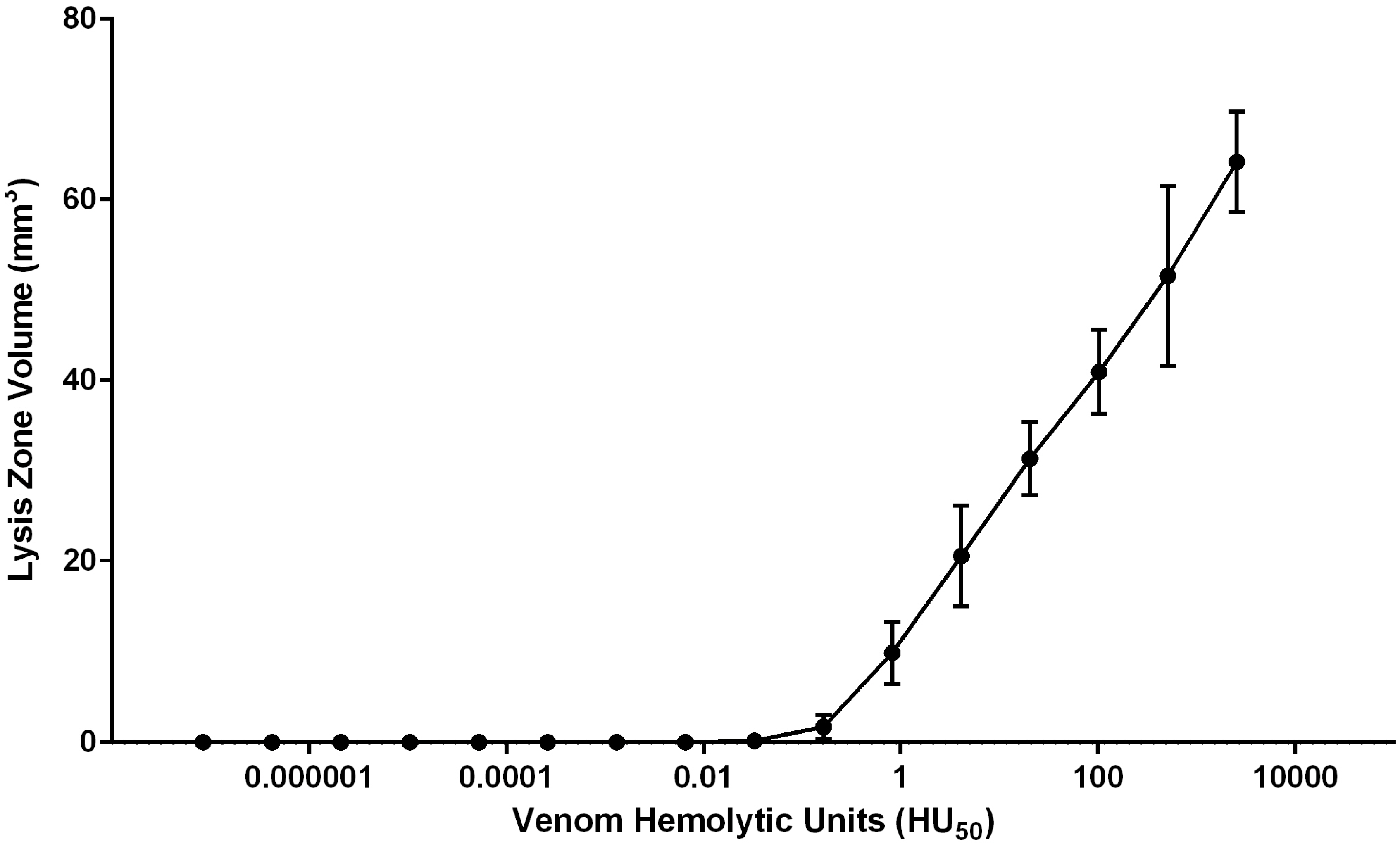
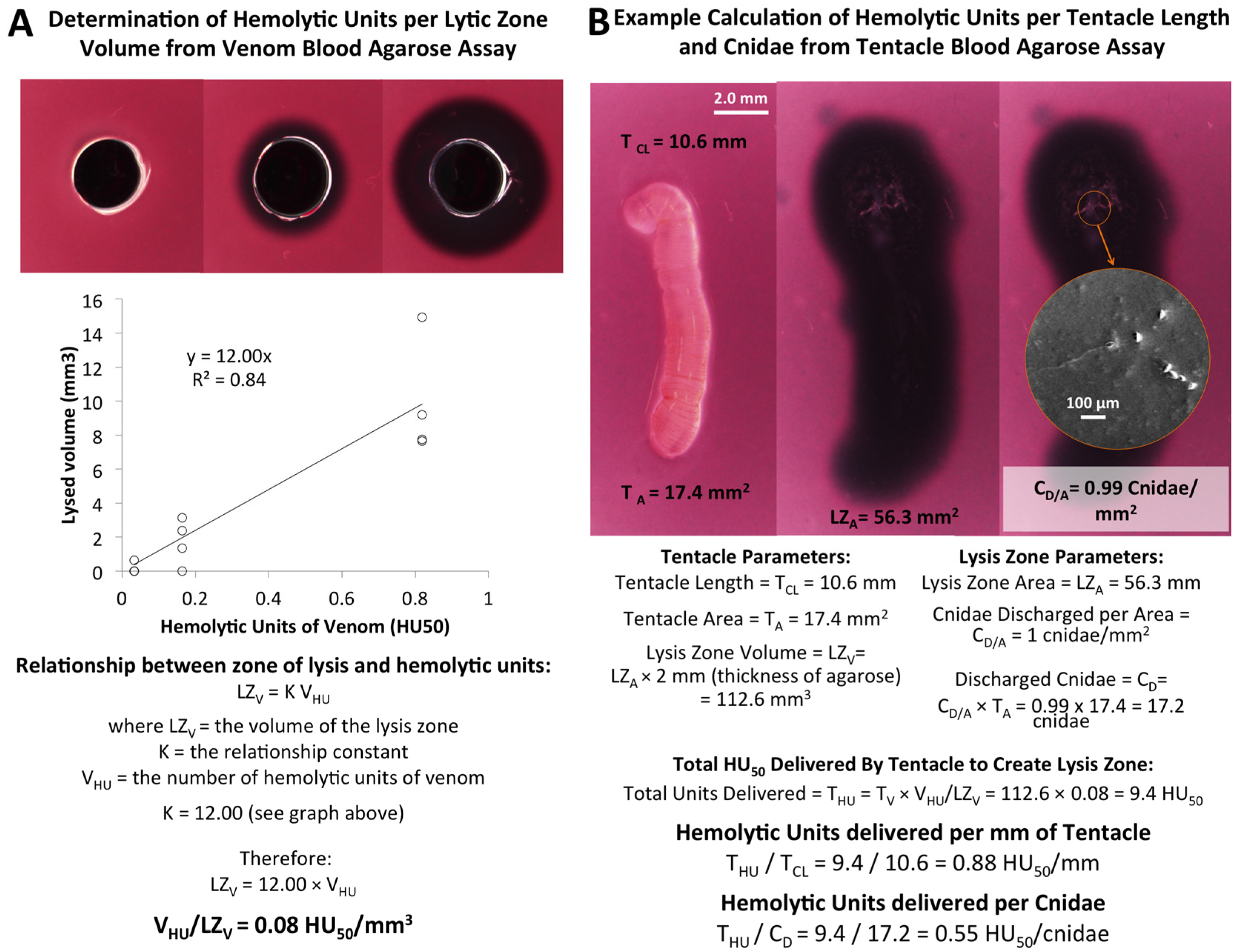
| Tentacle Length (TCL) | Volume of Lysis (LZV) | Cnidae Discharged per mm2 (CD/A) | Hemolytic Units per mm Tentacle | Hemolytic Units per Cnidae |
|---|---|---|---|---|
| 6.45–10.61 mm | 65.7–118.6 mm3 | 1.0–6.3 cnidae/mm2 | 0.7–1.4 HU50/mm | 0.1–0.5 HU50/cnidae |
| 8.55 ± 0.61 mm | 99.2 ± 6.2 mm3 | 3.3 ± 0.7 cnidae/mm2 | 1.0 ± 0.08 HU50/mm | 0.3 ± 0.06 HU50/cnidae |
2.1.4. Rigorous Controls
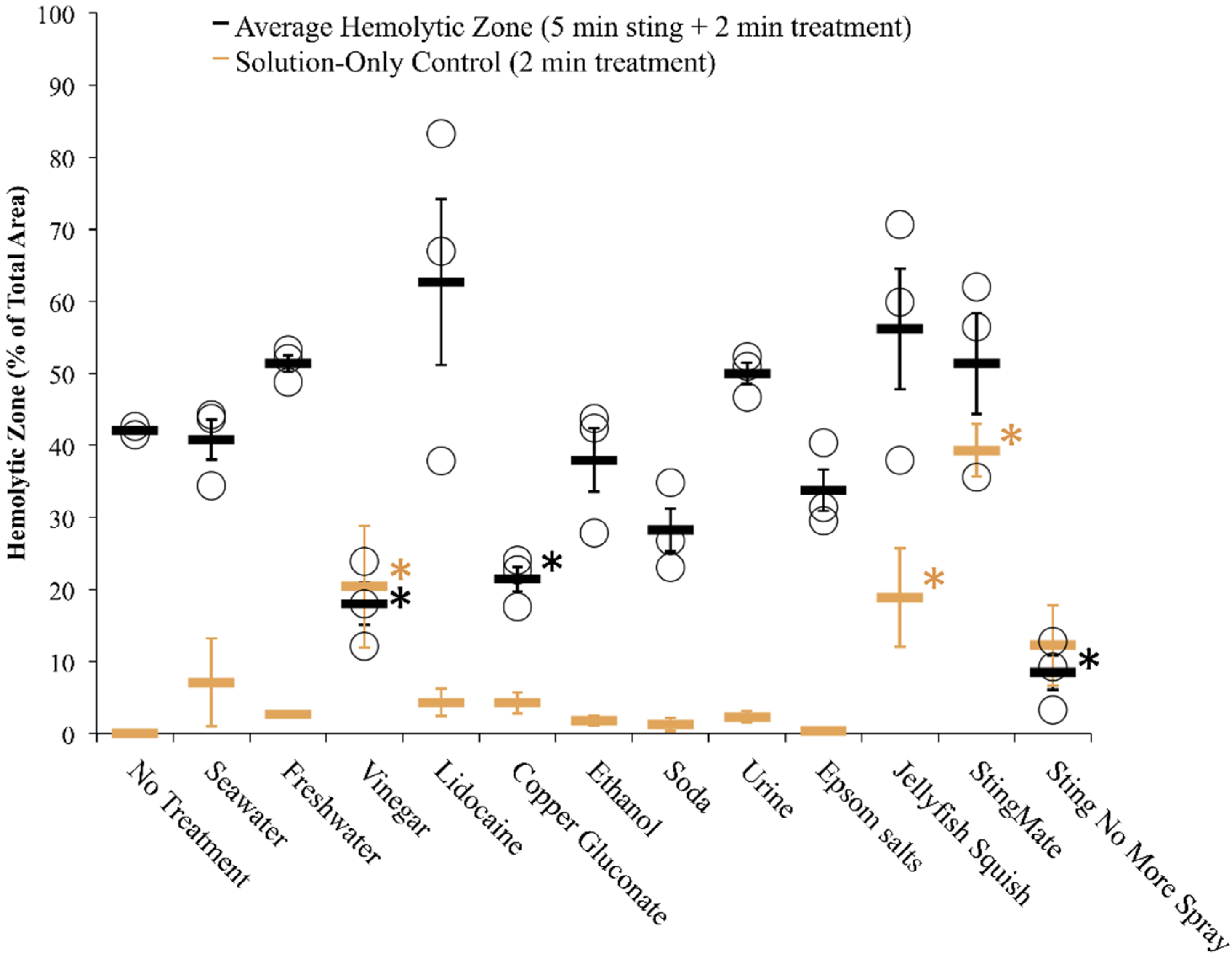
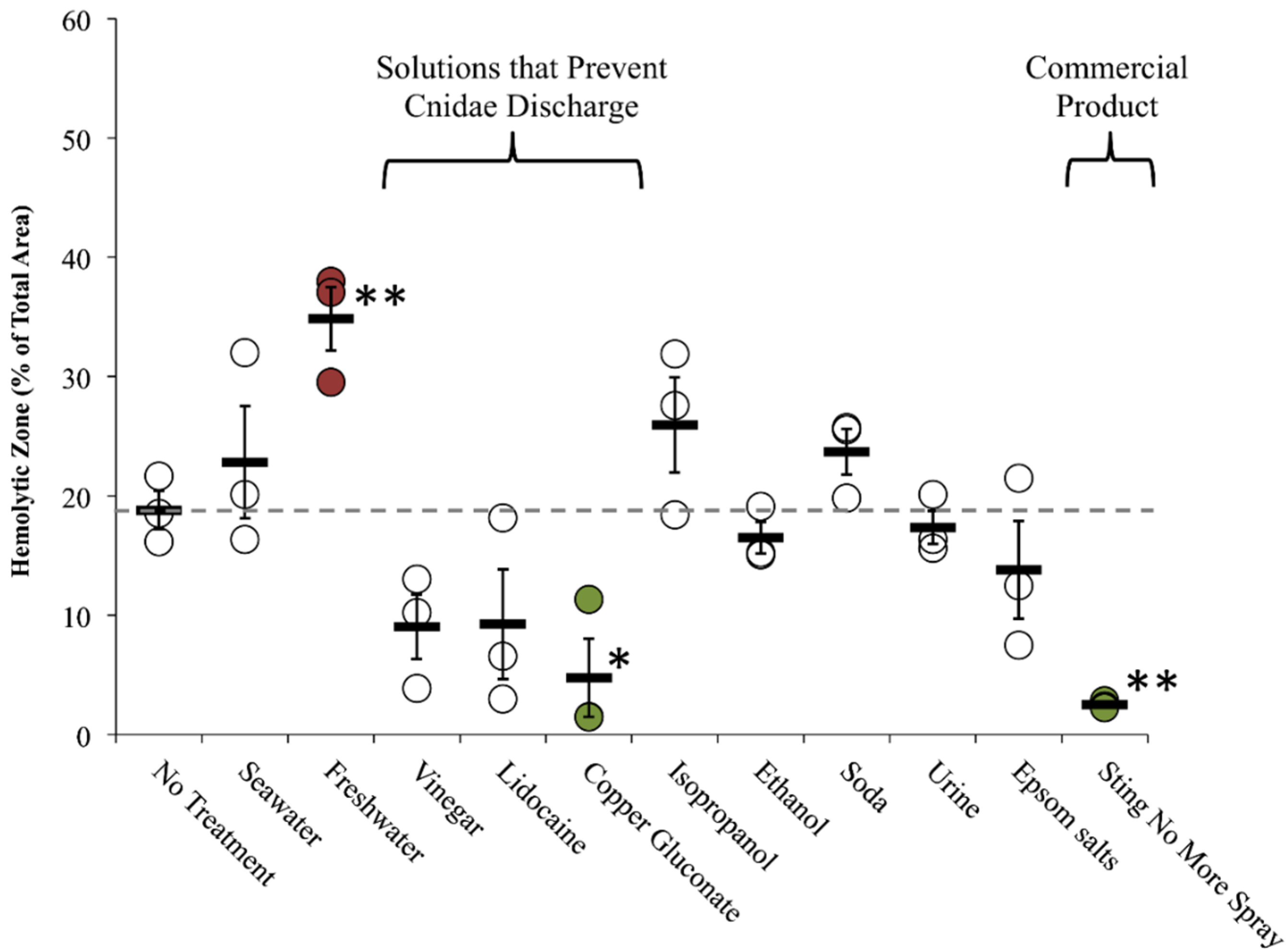
2.2. Evaluation of First Aid Approaches
2.2.1. Tentacle Solution Assay (TSA)

| Test Solution | Solution-Induced Discharge | Isopropanol-Stimulated Discharge | Pressure-Stimulated Discharge |
|---|---|---|---|
| Seawater | 1.8% ± 1.8% | 42.4% ± 9.3% § | 47.2% ± 4.7% § |
| Vinegar | 0.3% ± 0.3% | 2.1% ± 1.2% † | 0.0% ± 0.0% * |
| Lidocaine | 0.2% ± 0.2% | 70.5% ± 5.3% § | 31.8% ± 6.6% § |
| Copper Gluconate | 0.0% ± 0.0% | 13.2% ± 3.7% † | 40.4% ± 7.7% § |
| Isopropanol | 52.1% ± 13.2% * | - | 82.3% ± 2.0% * |
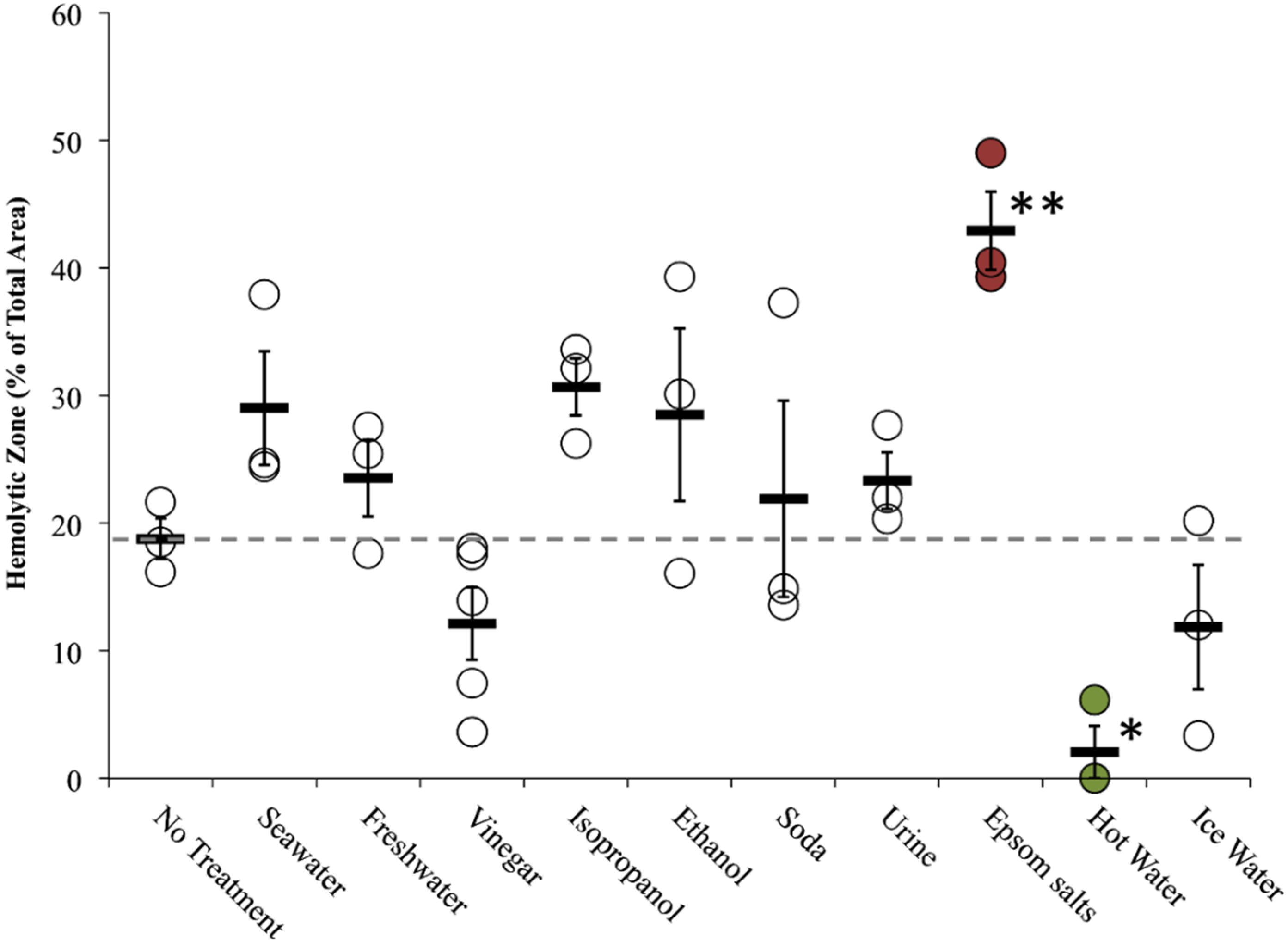
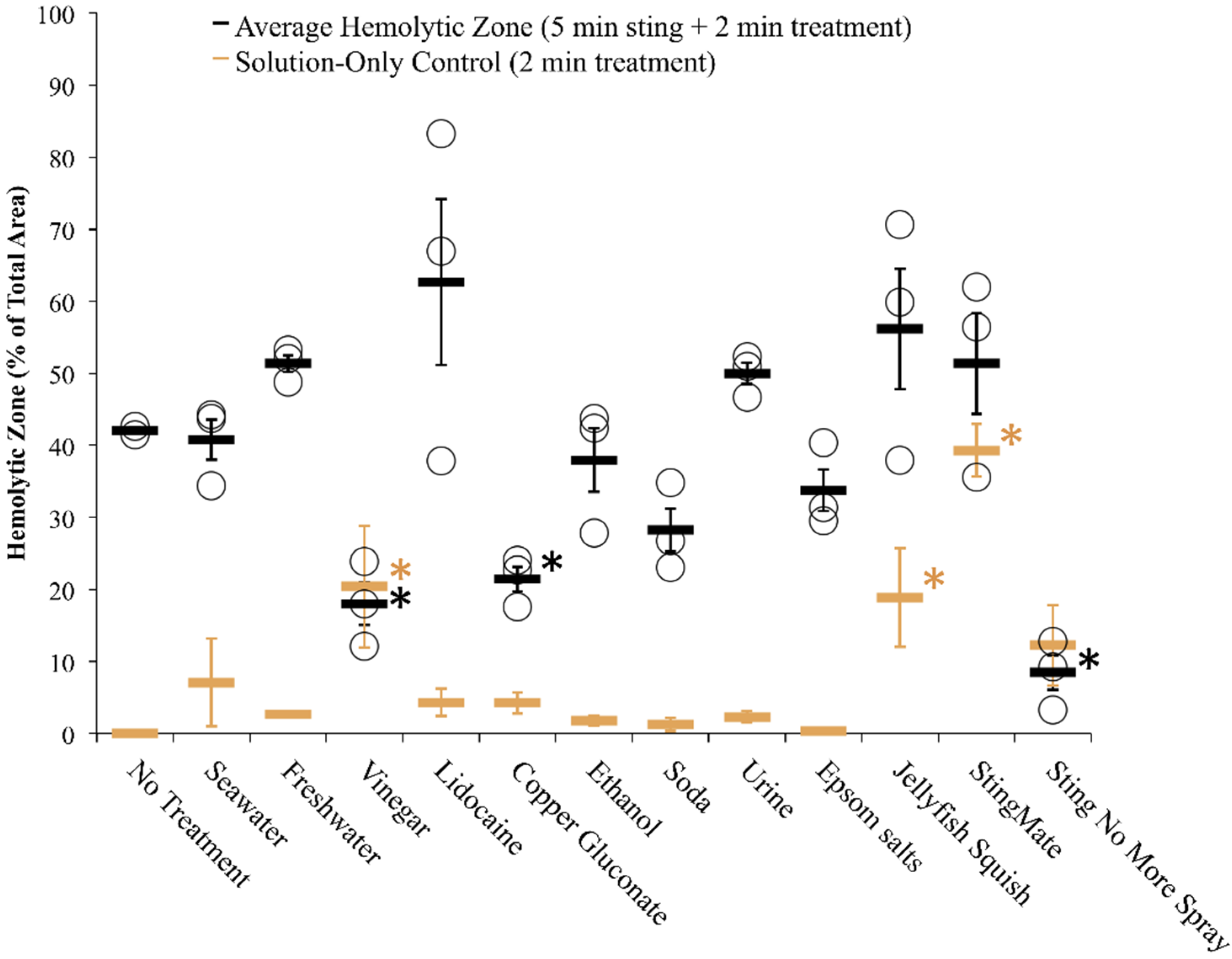
2.2.2. Tentacle Skin Blood Agarose Assay (TSBAA)
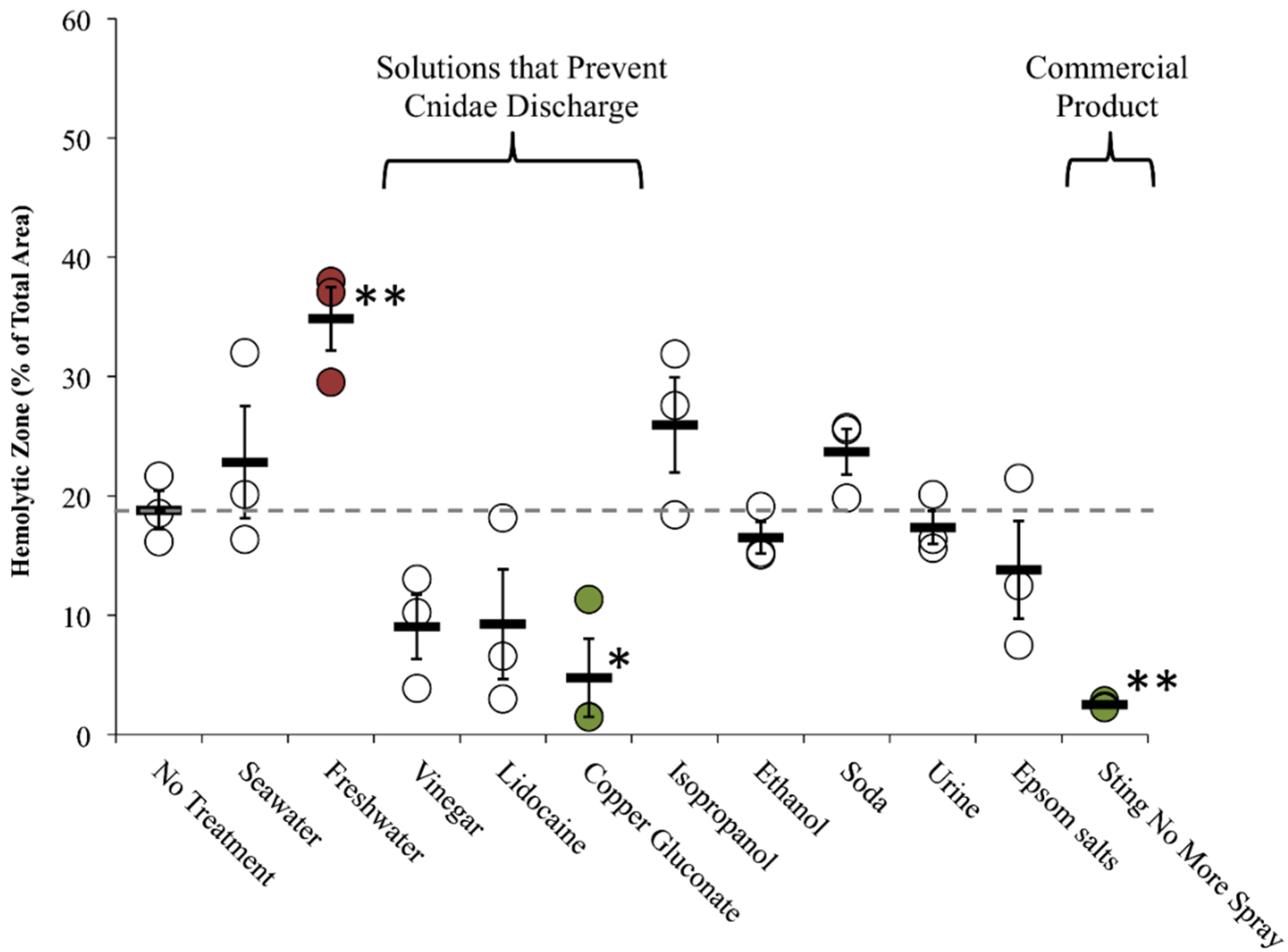
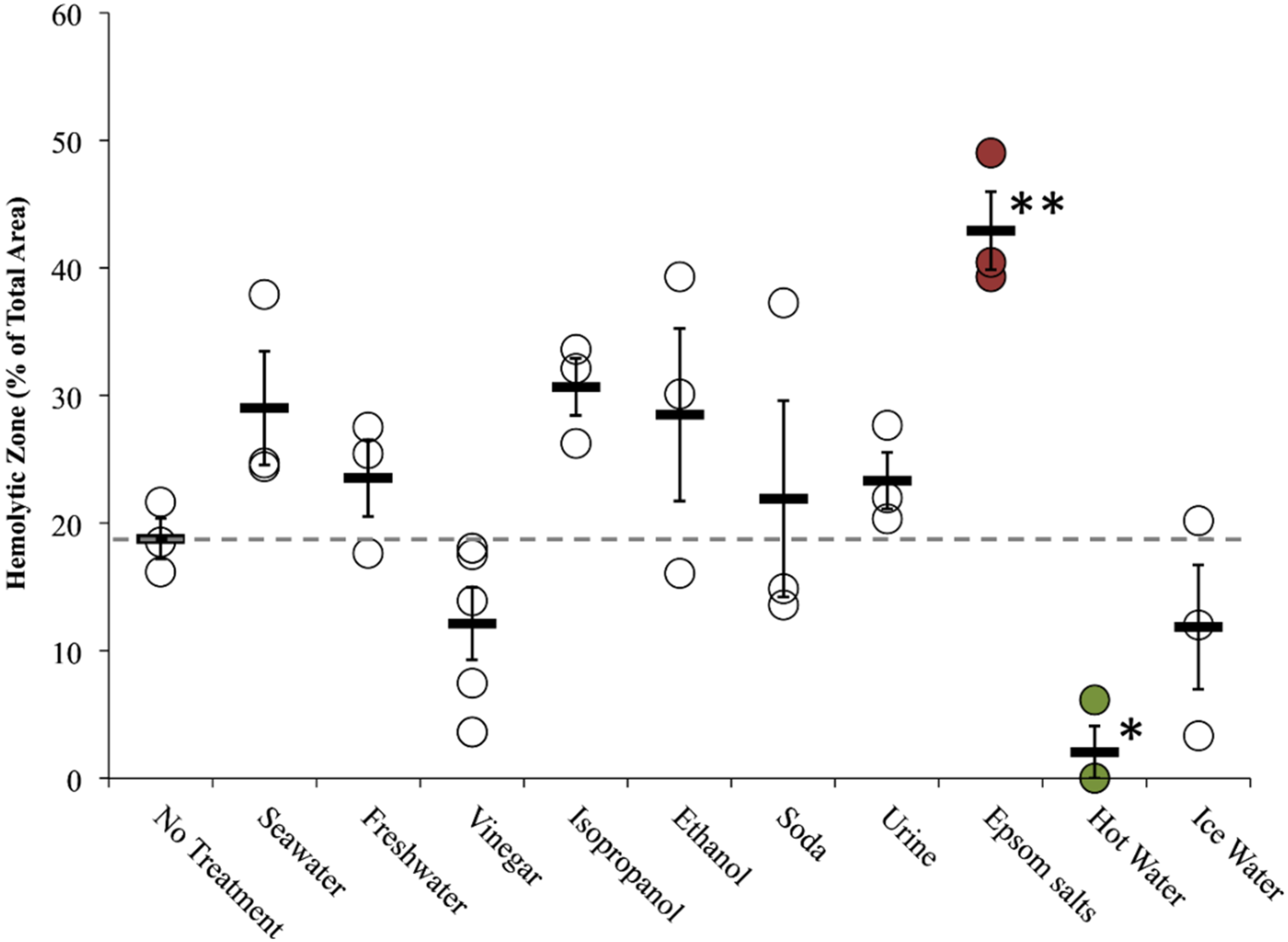
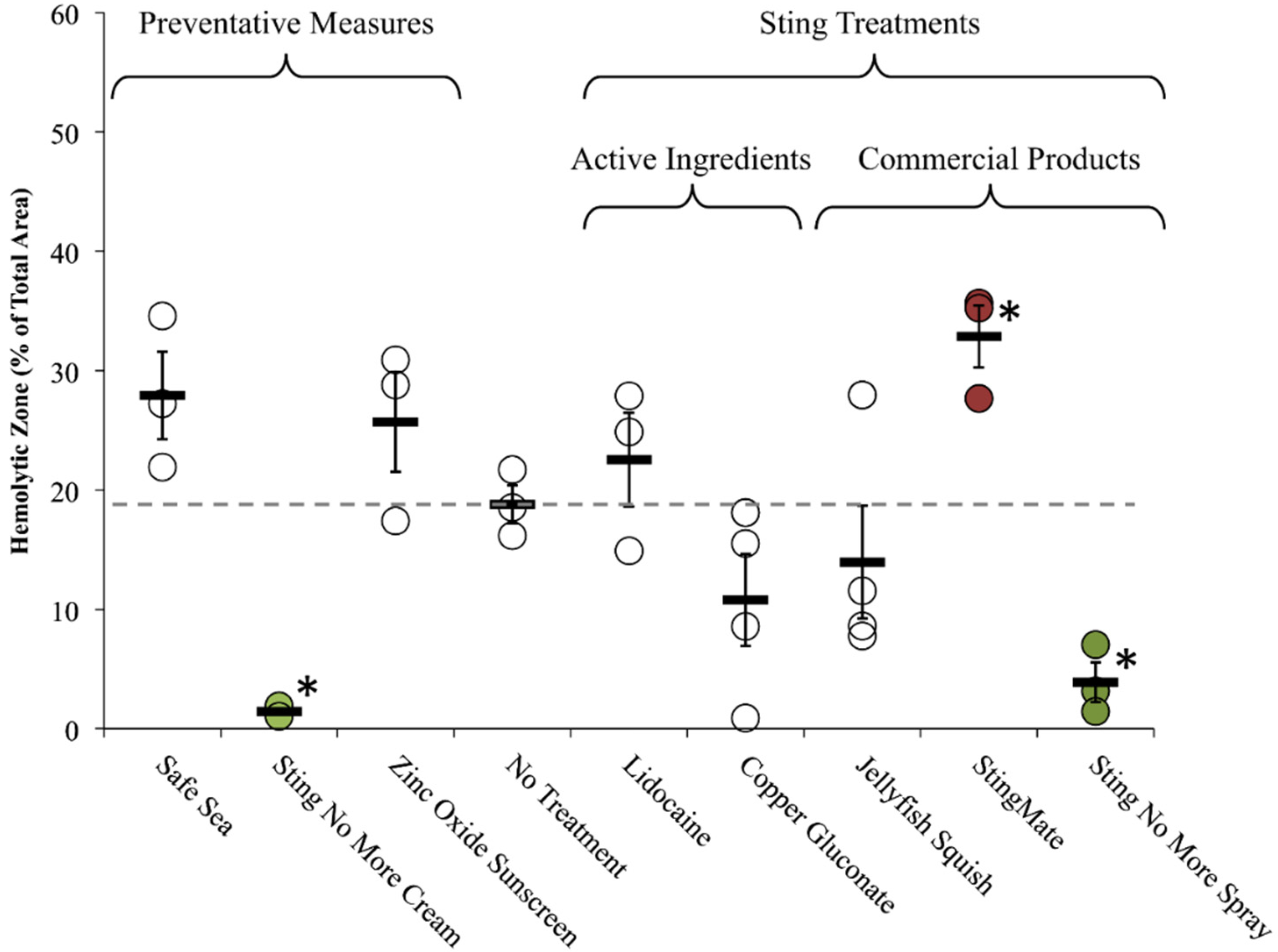

2.2.3. Informed First-Aid Treatments
| First Aid | Summary of Published Cnidarian Sting Literature | Formulation in this Study | TSA (% Discharge) | TSBAA (Lysis % Area) | ||||
|---|---|---|---|---|---|---|---|---|
| Nematocyst Discharge | Pain | Rinse Solution | Post-Sting Treatment | |||||
| Freshwater | C | + [28] | C | 0 [28], 0 [33] | Double distilled water (ddH2O) | C 29.6 ± 7.6 | C 34.9 ± 2.7 † | C 23.5 ± 3.0 |
| S | + [28] | S | 0 [28], 0 [33] | |||||
| H | 0 [36], 0 [40] | H | N.D. | |||||
| Seawater | C | 0 [28], 0 [29], 0 [30], 0 [31] | C | – [28], 0 [33] | Autoclaved Instant Ocean | C 1.8 ± 1.8 | C 22.8 ± 1.6 | C 29.0 ± 4.5 |
| S | 0 [28], 0 [33], 0 [34] | S | – [28], 0 [33] | |||||
| H | 0 [33], 0 [39] | H | 0 [38] | |||||
| Vinegar (Weak Acetic Acid) | C | – [28] (no test for inhibition), – [29], – [30], – [31], – [32] | C | 0 [28], – [31], + [33] (5%), 0/– [42] | Distilled white vinegar | C 0.3 ± 0.3 | C 9.0 ± 2.7 | C 12.1 ± 2.8 |
| S | + [28], 0 [33], – [34], + [35], + [41] | S | + [28], + [33] (5%) | |||||
| H | + [33], – [35], + [36], 0/+ [37], –39, 0/+ [40] | H | – [38], – [39] | |||||
| Lidocaine | C | 0 [28] (Xylocaine®; no test for inhibition) | C | – [28] (see note), – [33] | 4% lidocaine-HCl in 110 mM saline | C 0.2 ± 0.2 | C 9.2 ± 4.6 | C 22.5 ± 6.8 |
| S | – [33], – [34] (1%) | S | – [28] (see note), – [33] | |||||
| H | – [33] | H | N.D. | |||||
| Urine | C | + [29] | C | N.D. | Human, fresh | C 56.1 ± 15.6 δ | C 17.4 ± 1.4 | C 23.3 ± 2.2 |
| S | 0 [33] (10% urea) | S | N.D. | |||||
| H | 0 [33] (10% urea), 0/+ [40] | H | N.D. | |||||
| Carbonated Cola (Soda) | C | – [31] | C | – [31] | Regular Coca-Cola | C 7.3 ± 1.4 | C 23.7 ± 1.9 | C 21.9 ± 7.7 |
| S | N.D. | S | N.D. | |||||
| H | N.D. | H | N.D. | |||||
| Ethanol | C | + [28], + [29] (20%–100%) | C | + [28], + [33] | 70% in ddH2O | C 23.2 ± 5.6 | C 16.5 ± 1.3 | C 28.5 ± 6.8 |
| S | + [28], + [33], – [34], – [41] | S | + [28], + [33] | |||||
| H | + [33], + [36], +[39], + [40] (Methylated Spirits) | H | + [38] (Methylated Spirits) | |||||
| Epsom salts | N.D. | N.D. | N.D. | N.D. | Saturated in ddH2O | C 18.9 ± 7.8 | C 13.8 ± 4.1 | C 42.9 ± 3.1 † |
| Isopropanol | C | + [28] | C | + [28] | Household (70%) | C 81.2 ± 3.7 δ | C 25.9 ± 4.0 | C 30.7 ± 2.5 * |
| S | + [28] | |||||||
| Heat | N.D. | N.D. | C | – [42], – [43], – [44], – [45] | 42 °C–45 °C water pack | not performed | not performed | C 2.1 ± 2.0 |
| S | N.D. | |||||||
| H | N.D. | |||||||
| Cold | N.D. | N.D. | C | 0/– [42], 0 [43], 0 [44], 0 [45] | Ice water pack | not performed | not performed | C 11.9 ± 4.9 |
| S | N.D. | |||||||
| H | – [46] | |||||||
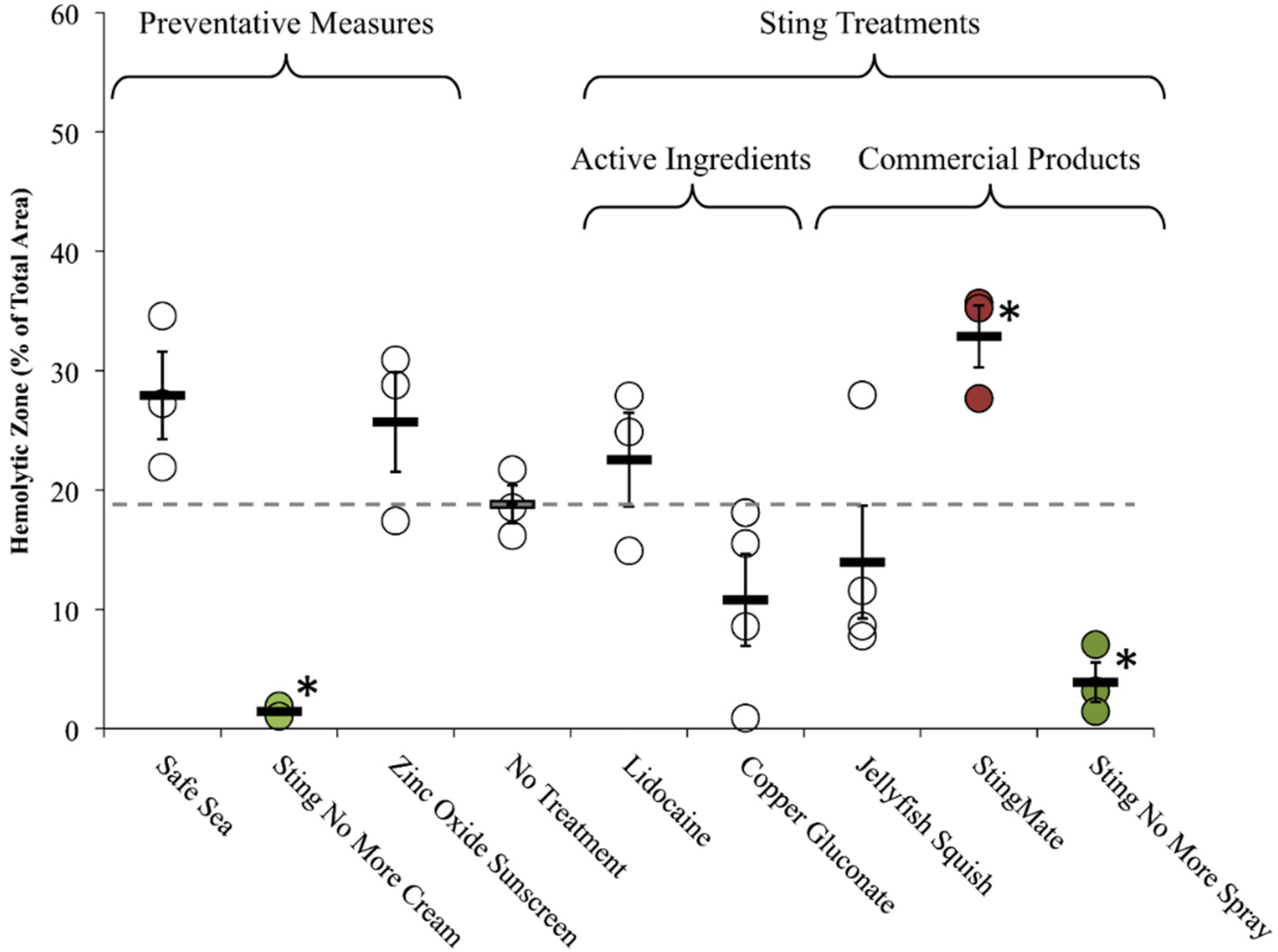
| Name | Manufacturer | Active Ingredients | Claims | Lab or Clinical Investigations | TSBAA (Lysis % Area ± SEM) |
|---|---|---|---|---|---|
| Jellyfish Squish® | Coastal Solutions, Inc | 4% Lidocaine HCl | Prevents skin lesions; much or complete relief of pain within minutes [49] | Studies mentioned on website (no peer-reviewed references) | Post-Sting Treatment: C 14.0 ± 4.7 |
| StingMate® | First Aid Mates, LLC | 5% acetic acid, menthol | Pain relief within minutes; reduction in skin pathology [50] | Studies mentioned on website (no peer-reviewed references) | Post-Sting Treatment: C 32.9 ± 2.6 † |
| Safe Sea® | Nidaria Technology Ltd | Octinoxate 7.5%, Octixalate 5%, Zinc Oxide 5%, Titanium Dioxide 2% | Prevents the stimulation of jellyfish tentacles after contact with human skin [51] | Relative risk reduction of 82% when compared with normal sunscreen (only 13 stings total) ([52,53,54], See Note A) | Preventative: C 27.9 ± 6.3 |
| Sting No More™ (spray) | Alatalab Solutions, LLC | 30 mM copper gluconate, 350 mM urea, 30 mM magnesium sulfate, 3% acetic acid | Relieves jellyfish stings and fire ant bites [55] | This study | Rinse Solution: C 2.5 ± 0.2 * |
| Post-Sting Treatment: C 3.9 ± 1.7 * | |||||
| Sting No More™ (cream) | Alatalab Solutions, LLC | 50 mM copper gluconate, 180 mM magnesium sulfate, 10% urea, in a pharmaceutical-grade, non-greasy dermal base | Relieves jellyfish stings and fire ant bites [55] | This study | Preventative: C 1.4 ± 0.3 * |
3. Experimental Section
3.1. Tentacle Solution Assay (TSA) to Evaluate Nematocyst Discharge
3.2. Ex Vivo Assays to Evaluate Hemolytic Activity
3.2.1. Venom Blood Agarose Assay (VBAA)
3.2.2. Tentacle Blood Agarose Assay (TBAA)
3.2.3. Tentacle Skin Blood Agarose Assay (TSBAA)
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brotz, L.; Cheung, W.W.; Kleisner, K.; Pakhomov, E.; Pauly, D. Increasing jellyfish populations: Trends in large marine ecosystems. Hydrobiologia 2012, 690, 3–20. [Google Scholar] [CrossRef]
- Cnidaria Envenomation Treatment & Management. Available online: http://emedicine.medscape.com/article/769538-treatment (accessed on 25 August 2015).
- Jellyfish Stings Treatment. Available online: http://www.emedicinehealth.com/jellyfish_stings/page4_em.htm#jellyfish_stings_treatment (accessed on 25 August 2015).
- Jellyfish Stings. Available online: http://www.mayoclinic.org/diseases-conditions/jellyfish-stings/basics/treatment/con-20034045 (accessed on 25 August 2015).
- Australian Resuscitation Council Guideline 9.4.5 Envenomation: Jellyfish Stings July 2010. Available online: http://resus.org.au/?wpfb_dl=41 (accessed on 13 June 2015).
- Jellyfish and other sea creature stings–Treatment. Available online: http://www.nhs.uk/Conditions/Stings-marine-creatures/Pages/Treatment.aspx (accessed on 25 August 2015).
- Markenson, D.; Ferguson, J.D.; Chameides, L.; Cassan, P.; Chung, K.-L.; Epstein, J.; Gonzales, L.; Herrington, R.A.; Pellegrino, J.L.; Ratcliff, N.; et al. Part 17: First aid: American Heart Association and American Red Cross guidelines. Circulation 2010, 122, S934–S946. [Google Scholar] [CrossRef] [PubMed]
- How to Treat Jellyfish Stings. Available online: http://www.wikihow.com/Treat-Jellyfish-Stings (accessed on 25 August 2015).
- Groundbreaking Study: A Sting in the Tail for Venom Research. Available online: http://www-public.jcu.edu.au/news/JCU_135736 (accessed on 25 August 2015).
- Vinegar on Jellyfish Sting Can be Deadly: Study. Available online: http://www.sbs.com.au/news/article/2014/04/08/vinegar-jellyfish-sting-can-be-deadly-study (accessed on 25 August 2015).
- Vinegar May Kill Rather than Cure Victims of Box Jellyfish Stings: Study. Available online: http://www.frenchtribune.com/teneur/1422205-vinegar-may-kill-rather-cure-victims-box-jellyfish-stings-study (accessed on 25 August 2015).
- What to Do if You Are Stung by a Jellyfish - and What You Really SHOULDN’T Do. Available online: http://www.mirror.co.uk/lifestyle/health/what-you-stung-jellyfish---6013059 (accessed on 25 August 2015).
- Ward, N.; Darracq, M.; Tomaszewski, C.; Clark, R. Evidence-based treatment of jellyfish stings in North America and Hawaii. Ann. Emerg. Med. 2012, 60, 399–414. [Google Scholar] [CrossRef] [PubMed]
- Fenner, P. Jellyfish stings—First aid and early medical treatments revisited. Ann. ACTM 2006, 7, 3–7. [Google Scholar]
- International Shark Attack File. Available online: https://www.flmnh.ufl.edu/fish/sharks/isaf/isaf.htm (accessed on 31 March 2015).
- Tibballs, J. Australian venomous jellyfish, envenomation syndromes, toxins and therapy. Toxicon 2006, 48, 830–859. [Google Scholar] [CrossRef] [PubMed]
- Tibballs, J.; Yanagihara, A.A.; Turner, H.; Winkel, K. Immunological and toxicological responses to jellyfish stings. Inflamm. Allergy Drug Targets 2011, 10, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Cegolon, L.; Heymann, W.; Lange, J.; Mastrangelo, G. Jellyfish stings and their management: A review. Marine Drugs 2013, 11, 523–550. [Google Scholar] [CrossRef] [PubMed]
- Yanagihara, A.A.; Shohet, R. Cubozoan venom-induced cardiovascular collapse is caused by hyperkalemia and prevented by zinc gluconate in mice. PLoS ONE 2012, 7, e51368. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Ratnapala, L.; Cooke, I.; Yanagihara, A.A. Partial purification and characterization of a hemolysin (CAH1) from Hawaiian box jellyfish (Carybdea alata) venom. Toxicon 2001, 39, 981–90. [Google Scholar] [CrossRef]
- Brinkman, D.; Burnell, J. Partial purification of cytolytic venom proteins from the box jellyfish, Chironex fleckeri. Toxicon 2008, 51, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Barnes, J.H. Cause and effect of Irukandji stingings. Med. J. Aust. 1964, 1, 897–904. [Google Scholar] [PubMed]
- Yanagihara, A.A. Compositions and Multi-Step Methods of Using the Same for the Treatment of Jellyfish Stings. U.S. Patent WO2012112230A2, 18 February 2012. [Google Scholar]
- Yanagihara, A.A. Methods and Compositions for Treating and/or Inhibiting Toxins Using Copper-Containing Compounds. U.S. Patent WO2000064472A1, 9 June 2015. [Google Scholar]
- Anderson, P.A.V.; McKay, C.M. The electrophysiology of cnidocytes. J. Exp. Biol. 1987, 133, 215–230. [Google Scholar]
- Jouiaei, M.; Casewell, N.R.; Yanagihara, A.A.; Nouwens, A.; Cribb, B.W.; Whitehead, D.; Jackson, T.N.W.; Ali, S.A.; Wagstaff, S.C.; Koludarov, I.; et al. Firing the sting: Chemically induced discharge of cnidae reveals novel proteins and peptides from box jellyfish (Chironex fleckeri) venom. Toxins 2015, 7, 936–950. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.L.; Carrette, T.; Cullen, P.; Mulcahy, R.F.; Little, M.; Seymour, J. Pressure immobilisation bandages in first aid treatment of jellyfish envenomation: Current recommendations reconsidered. Med. J. Aust. 2000, 173, 650–652. [Google Scholar] [PubMed]
- Pyo, M.J.; Lee, H.; Bae, S.K.; Heo, Y.; Choudhary, I.; Yoon, W.D.; Kang, C.; Kim, E. Modulation of jellyfish nematocyst discharges and management of human skin stings in Nemopilema nomurai and Carybdea mora. Toxicon 2016, 109, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Hartwick, R.; Callanan, V.; Williamson, J. Disarming the box jellyfish: Nematocyst inhibition in Chironex fleckeri. Med. J. Aust. 1980, 1, 5–20. [Google Scholar]
- Fenner, P.J.; Williamson, J. Experiments with the nematocysts of Carybdea rastoni (“Jimble”). Med. J. Aust. 1987, 147, 258–259. [Google Scholar] [PubMed]
- Currie, B.; Ho, S.; Alderslade, P. Box-jellyfish, Coca-Cola and old wine. Med. J. Aust. 1993, 158, 868. [Google Scholar] [PubMed]
- Fenner, P.J.; Fitzpatrick, P.F.; Hartwick, R.J.; Skinner, R. “Morbakka”, another cubozoan. Med. J. Aust. 1985, 143, 550–551, 554–555. [Google Scholar] [PubMed]
- Birsa, L.; Verity, P.; Lee, R. Evaluation of the effects of various chemicals on discharge of and pain caused by jellyfish nematocysts. Comp. Biochem. Physiol. C 2010, 151, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Morabito, R.; Marino, A.; Dossena, S.; La Spada, G. Nematocyst discharge in Pelagia noctiluca (Cnidaria, Scyphozoa) oral arms can be affected by lidocaine, ethanol, ammonia and acetic acid. Toxicon 2014, 83, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Burnett, J.; Rubinstein, H.; Calton, G. First aid for jellyfish envenomation. South Med. J. 1983, 76, 870–872. [Google Scholar] [CrossRef] [PubMed]
- Exton, D.R. Treatment of Physalia physalis envenomation. Med. J. Aust. 1988, 149, 54. [Google Scholar] [PubMed]
- Fenner, P.J.; Williamson, J.A.; Burnett, J.W.; Rifkin, J. First aid treatment of jellyfish stings in Australia: Response to a newly differentiated species. Med. J. Aust. 1993, 158, 498–501. [Google Scholar] [PubMed]
- Turner, B.; Sullivan, P.; Pennefather, J. Disarming the bluebottle: Treatment of Physalia envenomation. Med. J. Aust. 1980, 2, 394–395. [Google Scholar] [PubMed]
- Mianzan, H.W.; Fenner, P.J.; Cornelius, P.F.S.; Ramirez, F.C. Vinegar as a disarming agent to prevent further discharge of the nematocysts of the stinging hydrozoan Olindias sambaquiensis. Cutis 2001, 68, 45–48. [Google Scholar] [PubMed]
- Rifkin, J.; Fenner, P.J.; Williamson, J.A. First aid treatment from the sting from the hydroid Lytocarpus philippinus: The structure of, and in vitro discharge experiments with its nematocysts. J. Wilderness Med. 1993, 4, 252–260. [Google Scholar] [CrossRef]
- Fenner, P.J.; Fitzpatrick, P.F. Experiments with the nematocysts of Cyanea capillata. Med. J. Aust. 1986, 145, 174. [Google Scholar] [PubMed]
- Taylor, J.G. Treatment of jellyfish stings. Med. J. Aust. 2007, 186, 43. [Google Scholar] [PubMed]
- Thomas, C.S.; Scott, S.A.; Galanis, D.J.; Goto, R.S. Box jellyfish (Carybdea alata) in Waikiki: Their influx cycle plus the analgesic effect of hot and cold packs on their stings to swimmers at the beach: A randomized, placebo-controlled, clinical trial. Hawaii Med. J. 2001, 60, 100–107. [Google Scholar] [PubMed]
- Yoshimoto, C.M.; Yanagihara, A.A. Cnidarian (coelenterate) envenomations in Hawaii improve following heat application. Trans. R. Soc. Trop. Med. Hygeine 2002, 96, 300–303. [Google Scholar] [CrossRef]
- Nomura, J.T.; Sato, R.L.; Ahern, R.M.; Snow, J.L.; Kuwaye, T.T.; Yamamoto, L.G. A randomized paired comparison trial of cutaneous treatments for acute jellyfish (Carybdea alata) stings. Am. J. Emerg. Med. 2002, 20, 624–626. [Google Scholar] [CrossRef] [PubMed]
- Exton, D.R.; Fenner, P.J.; Williamson, J.A. Cold packs: Effective topical analgesia in the treatment of painful stings by Physalia and other jellyfish. Med. J. Aust. 1988, 151, 625–626. [Google Scholar]
- Welfare, P.; Little, M.; Pereira, P.; Seymour, J. An in vitro examination of the effect of vinegar on discharged nematocysts of Chironex fleckeri. Diving Hyperb. Med. 2014, 44, 30–34. [Google Scholar] [PubMed]
- O’Reilly, G.M.; Isbister, G.K.; Lawrie, P.M.; Treston, G.T.; Currie, B.J. Prospective study of jellyfish stings from tropical Australia, including the major box jellyfish Chironex fleckeri. Med. J. Aust. 2001, 175, 652–655. [Google Scholar] [PubMed]
- Jellyfish Squish. Available online: http://www.coastalsolutionsinc.com/products/jellyfish_squish_/ (accessed on 13 June 2015).
- StingMate. Available online: http://stingmate.com (accessed on 13 June 2015).
- SafeSea. Available online: http://www.safesea.eu (accessed on 13 June 2015).
- Boulware, D.R. A randomized, controlled field trial for the prevention of jellyfish stings with a topical sting inhibitor. J. Travel Med. 2006, 13, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Burnett, J.W.; Purcell, J.E.; Learn, D.B.; Meyers, T. A protocol to investigate the blockade of jellyfish nematocysts by topical agents. Contact Dermat. 1999, 40, 55–56. [Google Scholar] [CrossRef]
- Burnett, J.W. Lack of efficacy of a combination sunblock and “jellyfish sting inhibitor” topical preparation against Physalia sting. Dermatitis 2005, 16, 151. [Google Scholar] [CrossRef] [PubMed]
- Sting No More. Available online: http://stingnomore.com (accessed on 20 August 2015).
- Cook, W.A.; Hiles, M.C.; Kozma, T.G.; Patel, U.H. Graft Prosthesis Materials. U.S. Patent 6206931, 27 March 2001. [Google Scholar]
- Aickin, M.; Gensler, H. Adjusting for multiple testing when reporting research results: The Bonferroni vs Holm methods. Am. J. Public Health 1996, 86, 726–728. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yanagihara, A.A.; Wilcox, C.; King, R.; Hurwitz, K.; Castelfranco, A.M. Experimental Assays to Assess the Efficacy of Vinegar and Other Topical First-Aid Approaches on Cubozoan (Alatina alata) Tentacle Firing and Venom Toxicity. Toxins 2016, 8, 19. https://doi.org/10.3390/toxins8010019
Yanagihara AA, Wilcox C, King R, Hurwitz K, Castelfranco AM. Experimental Assays to Assess the Efficacy of Vinegar and Other Topical First-Aid Approaches on Cubozoan (Alatina alata) Tentacle Firing and Venom Toxicity. Toxins. 2016; 8(1):19. https://doi.org/10.3390/toxins8010019
Chicago/Turabian StyleYanagihara, Angel A., Christie Wilcox, Rebecca King, Kikiana Hurwitz, and Ann M. Castelfranco. 2016. "Experimental Assays to Assess the Efficacy of Vinegar and Other Topical First-Aid Approaches on Cubozoan (Alatina alata) Tentacle Firing and Venom Toxicity" Toxins 8, no. 1: 19. https://doi.org/10.3390/toxins8010019
APA StyleYanagihara, A. A., Wilcox, C., King, R., Hurwitz, K., & Castelfranco, A. M. (2016). Experimental Assays to Assess the Efficacy of Vinegar and Other Topical First-Aid Approaches on Cubozoan (Alatina alata) Tentacle Firing and Venom Toxicity. Toxins, 8(1), 19. https://doi.org/10.3390/toxins8010019