Sub-Emetic Toxicity of Bacillus cereus Toxin Cereulide on Cultured Human Enterocyte-Like Caco-2 Cells


 ,
,
Abstract
:1. Introduction
2. Results and Discussion
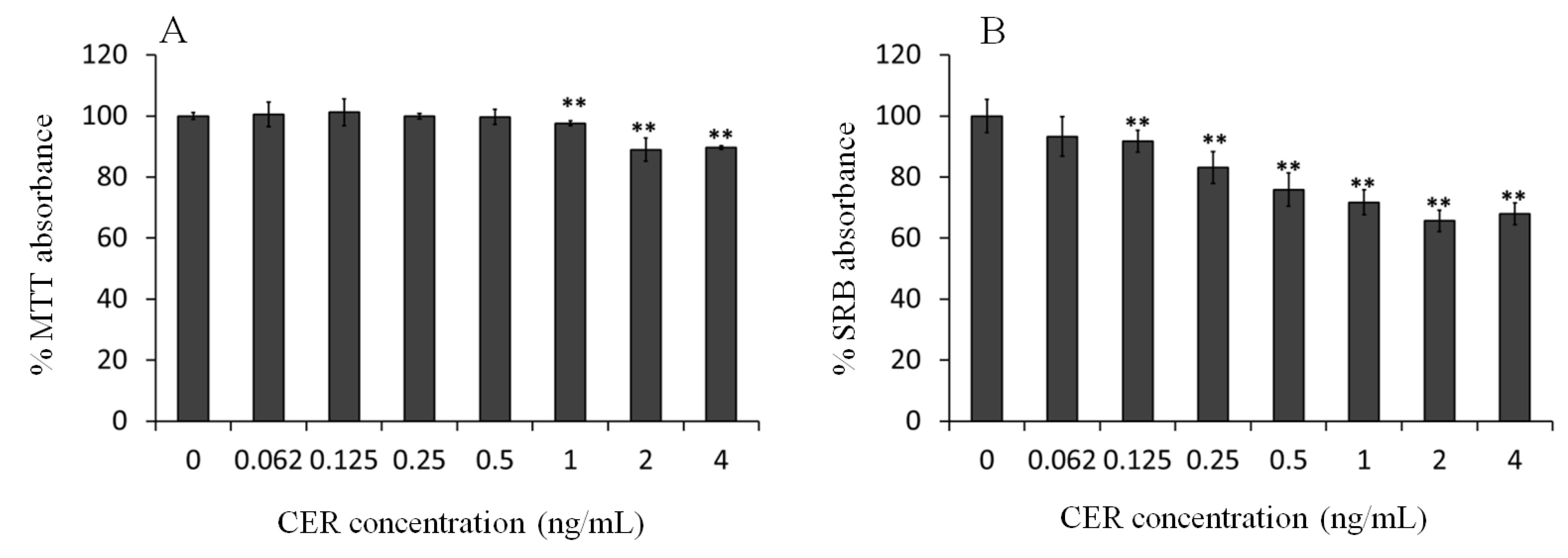
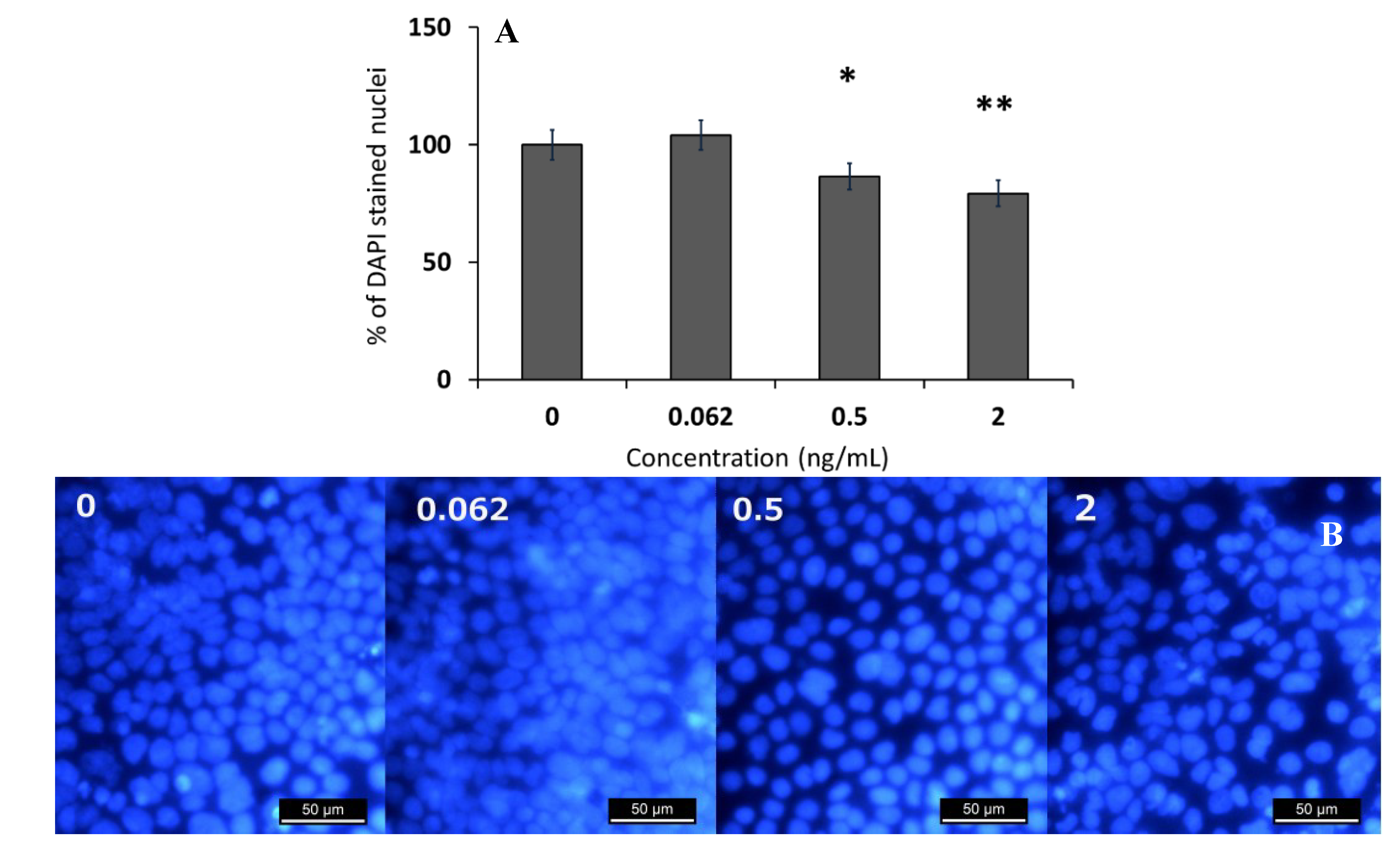
2.1. Effect of Sub-Emetic Cereulide Concentrations on the Toxicity, Protein Content and Cell Morphology of Caco-2 Cells
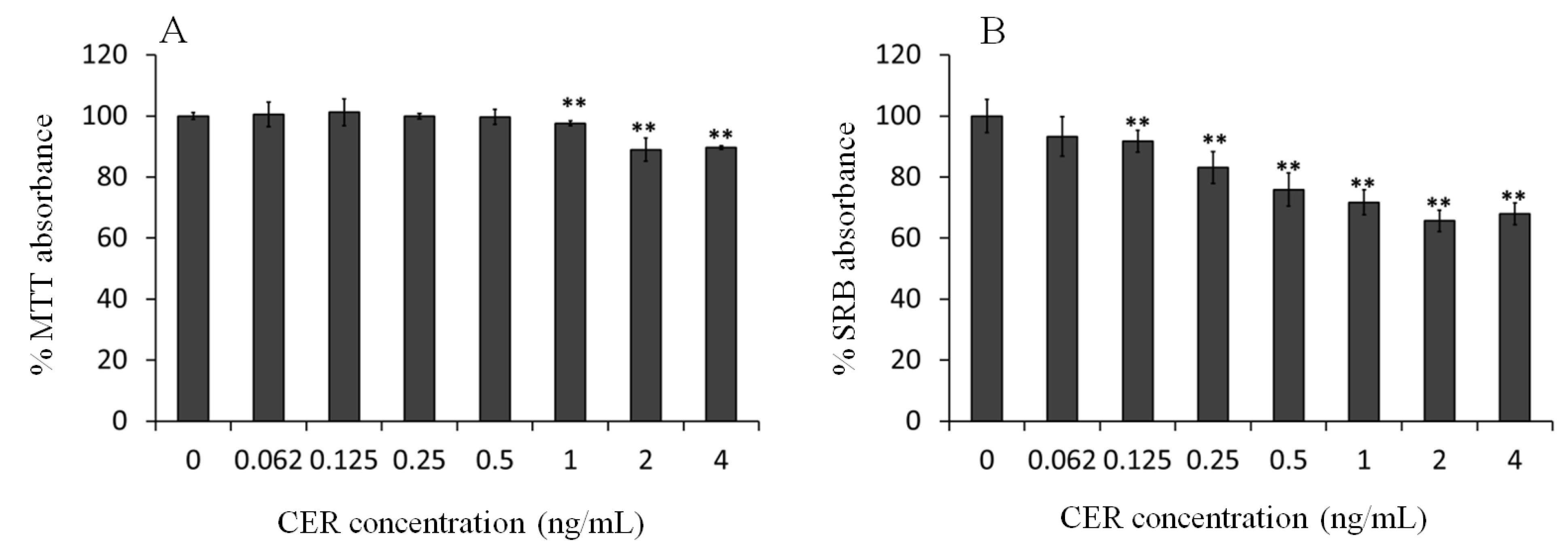
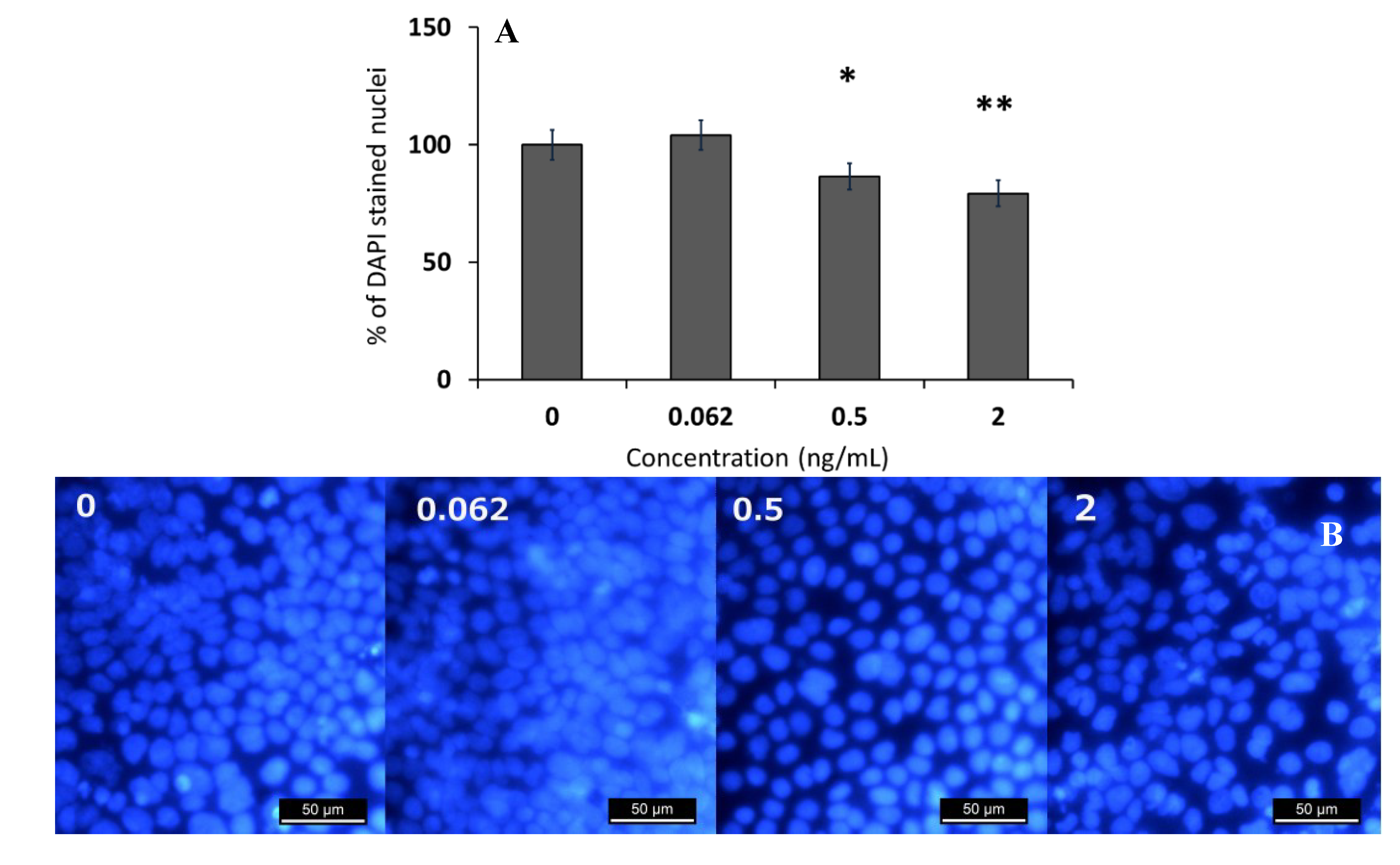
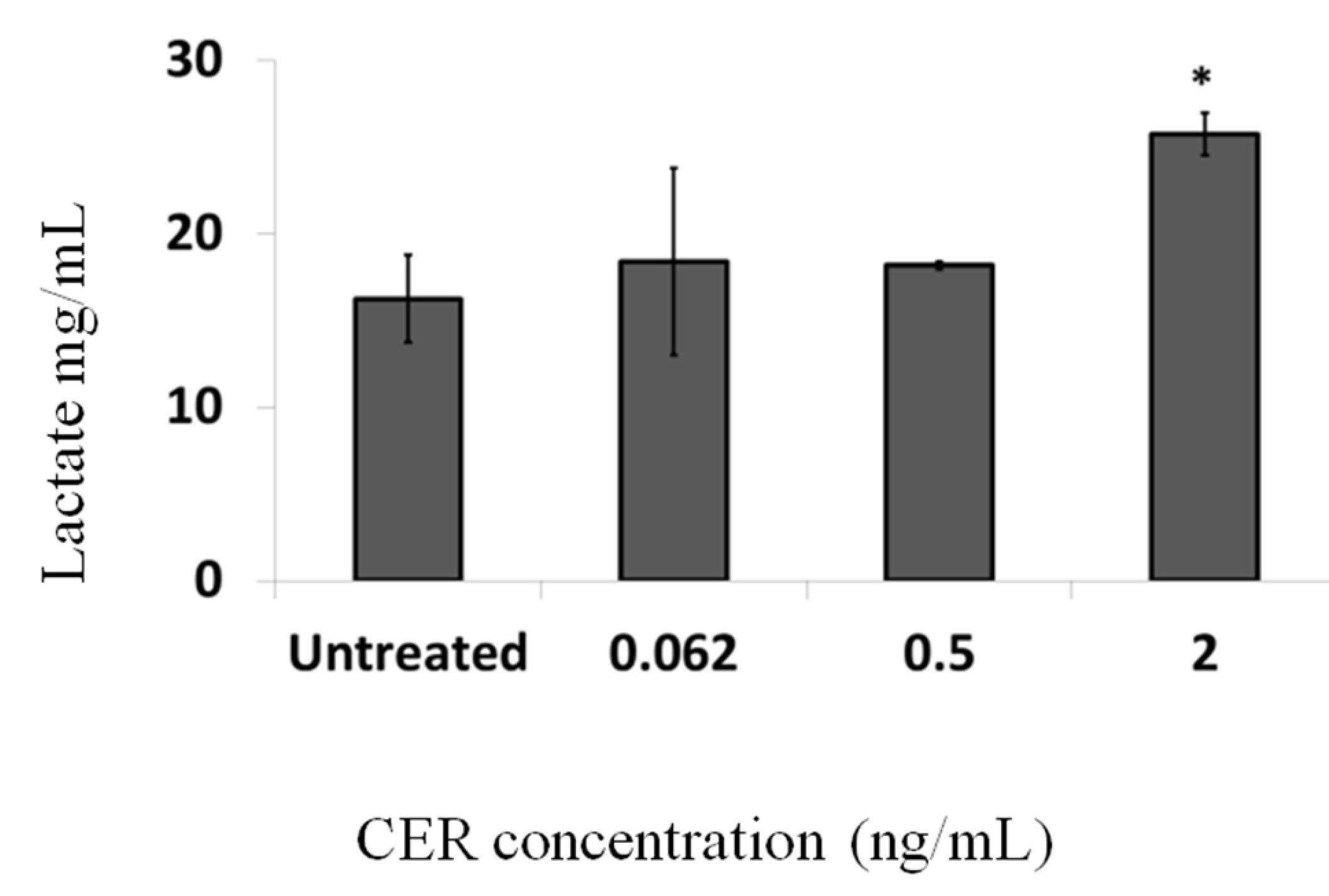
2.2. Effect of Sub-Emetic Cereulide Concentrations on Acidification and Lactate Formation by Caco-2 Cells
2.3. Comparison of the Mitochondrial Effects of Sub-Emetic Cereulide Concentrations between the Caco-2 MTT Assay and the Boar Sperm Test
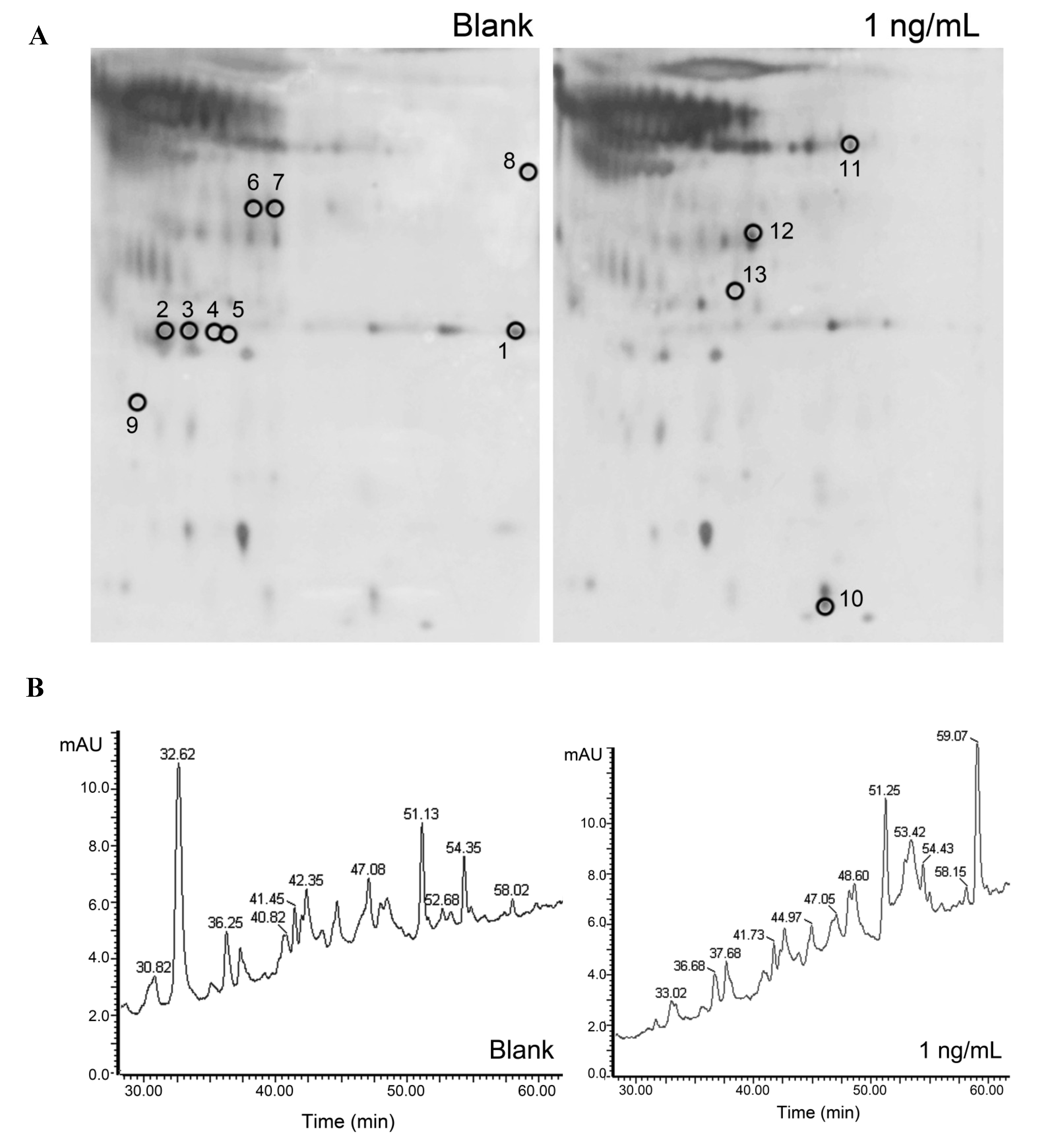
2.4. Differences in Protein Expression between Treated and Untreated Caco-2 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. a | Fraction | Protein | gi number b | MW | pI c | Coverage % | MASCOT score | Expression | Function |
|---|---|---|---|---|---|---|---|---|---|
| 1 | M/C | Triosephosphate isomerase 1 (TPI1) | 17389815 | 26,625 | 6.5 | 38 | 145 * | - | Gluconeogenesis, glycolysis, pentose shunt |
| 2 | M | Apolipoprotein A-I (ApoA-I) | 90108664 | 28,062 | 5.3 | 48 | 197 * | - | Lipid metabolism, major component of HDL, clear cholesterol from tissues, positive effect on cardiovascular diseases |
| 3 | M | Apolipoprotein A-I (ApoA-I) | 90108664 | 28,062 | 5.3 | 56 | 234 * | - | Lipid metabolism, major component of HDL, clear cholesterol from tissues, positive effect on cardiovascular diseases |
| 4 | M | Proapolipoprotein | 178775 | 28,944 | 5.5 | 39 | 133 * | - | Lipid metabolism, major component of HDL, clear cholesterol from tissues, positive effect on cardiovascular diseases |
| 5 | M | Apolipoprotein A-I preproprotein | 4557321 | 30,745 | 5.6 | 37 | 169 * | - | Lipid metabolism, major component of HDL, clear cholesterol from tissues, positive effect on cardiovascular diseases |
| 6 | M | Immunoglobulin-like and fibronectin type III (IGFN) | 257196151 | 383,568 | 5.7 | 32 | 71 * | - | Cell adhesion (wound healing), differentiation, migration, cytokine and tyrosine kinase receptors |
| 7 | M | Domain-containing protein 1 | 193783098 | 349,013 | 5.6 | 30 | 101 * | - | unknown |
| 8 | M | Phosphoglycerate kinase 1 (PGK1) | 48145549 | 44,574 | 8.3 | 25 | 121 * | - | Conversion pyruvate to lactate, cellular response to extracellular stimulus |
| 9 | M | Nucleolar protein 5A (56kDa with KKE/D repeat), isoform CRA_b | 119630990 | 47,898 | 9.4 | 20 | 74 * | - | Ribosomal action |
| 10 | M | Proprotein convertase subtilisin/kexin type 9 (Pcsk9) | 149243243 | 74,239 | 6.1 | 41 | 218 * | + | Cholesterol homeostasis, LDL decrease => hypercholesterolemia |
| 11 | M | Cathepsin D preproprotein | 4503143 | 44,524 | 6.1 | 28 | 118 * | + | Protease, ECM remodeling, CVD, apoptosis, immune response, tumor biomarker |
| 12 | M/C | Lactate dehydrogenase (isoform CRA_a) (LDH) | 49259209/ | 36,516 | 5.9 | 25 | 110 * | + | Conversion of lactate to pyruvate, marker for cell death (marker of membrane integrity) |
| 13 | M | Deoxyribonuclease I splicing isomer 1 (DNASE1) | 109809705 | 31,386 | 4.6 | 28 | 115 * | + | Waste-management endonuclease, DNA fragmentation, apoptosis |
| 14 | C | Tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein (YWHA) | 23114 | 28,083 | 4.8 | 8 | 98 ** | - | Signal transduction, trafficking, apoptosis, stress response, and malignant transformation |
| 15 | C | Malate dehydrogenase (MDH), cytoplasmic isoform 3 | 1255604 | 36,426 | 6.9 | 6 | 83 ** | - | Citric acid cycle, gluconeogenesis |
| 16 | C | Peroxiredoxin (Prdx) | 287641 | 22,111 | 8.3 | 9 | 102 ** | - | Detoxification of H2O2 |
| 17 | C | Glutathione S-transferase P (GSTP) | 31946 | 23,356 | 5.4 | 13 | 154 ** | - | Detoxification, oxidative stress |
| 18 | C | U2 small nuclear RNA auxiliary factor 2 isoform b | 37545 | 53,501 | 9.2 | 6 | 74 ** | - | mRNA processing |
| 19 | C | Copine-1 isoform a (CPNE1) | 1791257 | 59,059 | 5.5 | 5 | 76 ** | + | Membrane trafficking |
| 20 | C | Glutathione S-transferase (GST) | 121730 | 25,631 | 8.9 | 6 | 81 ** | + | Detoxification, oxidative stress |
| 21 | C | Rho GDP dissociation inhibitor alpha (Rho GDI 1) | 36038 | 23,193 | 5.0 | 7 | 79 ** | + | Activation of the oxygen superoxide-generating NADPH oxidase |
| 22 | N | Albumin, CRA p | 119626079 | 69,348 | 5.92 | 21 | 89 * | + | Unknown |

3. Experimental Section
3.1. Cell Culture
3.2. Cereulide (CER)
3.3. Toxicity Tests
3.4. Lactate Analysis
3.5. Preparation of Protein Fractions for Proteomics
3.5.1. Secreted Protein Fraction
3.5.2. Cytosolic Protein Fraction
3.5.3. Nuclear/Membrane-Bound Protein Fraction
3.6. 1D and 2D Gel Electrophoresis
In-Gel Digestion
3.7. Cap-LC
3.8. Mass Spectrometry
3.9. Protein Identification
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rajkovic, A.; Uyttendaele, M.; Vermeulen, A.; Andjelkovic, M.; Fitz-James, I.; in’t Veld, P.; Denon, Q.; Verhe, R.; Debevere, J. Heat resistance of bacillus cereus emetic toxin, cereulide. Lett. Appl. Microbiol. 2008, 46, 536–541. [Google Scholar] [CrossRef]
- Suwan, S.; Isobe, M.; Ohtani, I.; Agata, N.; Mori, M.; Ohta, M. Structure of cereulide, a cyclic dodecadepsipeptide toxin from bacillus cereus and studies on nmr characteristics of its alkali-metal complexes including a conformational structure of the k+ complex. J. Chem. Soc. Perkin Trans. 1 1995, 765–775. [Google Scholar]
- Horwood, P.F.; Burgess, G.W.; Oakey, H.J. Evidence for non-ribosomal peptide synthetase production of cereulide (the emetic toxin) in bacillus cereus. FEMS Microbiol. Lett. 2004, 236, 319–324. [Google Scholar]
- Toh, M.; Moffitt, M.C.; Henrichsen, L.; Raftery, M.; Barrow, K.; Cox, J.M.; Marquis, C.P.; Neilan, B.A. Cereulide, the emetic toxin of bacillus cereus, is putatively a product of nonribosomal peptide synthesis. J. Appl. Microbiol. 2004, 97, 992–1000. [Google Scholar] [CrossRef]
- Naranjo, M.; Denayer, S.; Botteldoorn, N.; Delbrassinne, L.; Veys, J.; Waegenaere, J.; Sirtaine, N.; Driesen, R.B.; Sipido, K.R.; Mahillon, J.; et al. Sudden death of a young adult associated with bacillus cereus food poisoning. J. Clin. Microbiol. 2011, 49, 4379–4381. [Google Scholar] [CrossRef]
- Dierick, K.; van Coillie, E.; Swiecicka, I.; Meyfroidt, G.; Devlieger, H.; Meulemans, A.; Hoedemaekers, G.; Fourie, L.; Heyndrickx, M.; Mahillon, J. Fatal family outbreak of bacillus cereus-associated food poisoning. J. Clin. Microbiol. 2005, 43, 4277–4279. [Google Scholar] [CrossRef]
- Mahler, H.; Pasi, A.; Kramer, J.M.; Schulte, P.; Scoging, A.C.; Bar, W.; Krahenbuhl, S. Fulminant liver failure in association with the emetic toxin of bacillus cereus. N. Engl. J. Med. 1997, 336, 1142–1148. [Google Scholar] [CrossRef]
- Messelhausser, U.; Kampf, P.; Fricker, M.; Ehling-Schulz, M.; Zucker, R.; Wagner, B.; Busch, U.; Holler, C. Prevalence of emetic bacillus cereus in different ice creams in bavaria. J. Food Prot. 2010, 73, 395–399. [Google Scholar]
- Ceuppens, S.; Rajkovic, A.; Heyndrickx, M.; Tsilia, V.; de Wiele, T.V.; Boon, N.; Uyttendaele, M. Regulation of toxin production by bacillus cereus and its food safety implications. Crit. Rev. Microbiol. 2011, 37, 188–213. [Google Scholar] [CrossRef]
- Wijnands, L.M.; Dufrenne, J.B.; Rombouts, F.M.; in’t Veld, P.H.; van Leusden, F.M. Prevalence of potentially pathogenic bacillus cereus in food commodities in the netherlands. J. Food Prot. 2006, 69, 2587–2594. [Google Scholar]
- Delbrassinne, L.; Andjelkovic, M.; Dierick, K.; Denayer, S.; Mahillon, J.; van Loco, J. Prevalence and levels of bacillus cereus emetic toxin in rice dishes randomly collected from restaurants and comparison with the levels measured in a recent foodborne outbreak. Foodborne Pathog. Dis. 2012, 9, 809–814. [Google Scholar] [CrossRef]
- Chon, J.W.; Kim, J.H.; Lee, S.J.; Hyeon, J.Y.; Song, K.Y.; Park, C.; Seo, K.H. Prevalence, phenotypic traits and molecular characterization of emetic toxin-producing bacillus cereus strains isolated from human stools in korea. J. Appl. Microbiol. 2012, 112, 1042–1049. [Google Scholar] [CrossRef]
- Shiota, M.; Saitou, K.; Mizumoto, H.; Matsusaka, M.; Agata, N.; Nakayama, M.; Kage, M.; Tatsumi, S.; Okamoto, A.; Yamaguchi, S.; et al. Rapid detoxification of cereulide in bacillus cereus food poisoning. Pediatrics 2010, 125, E951–E955. [Google Scholar] [CrossRef]
- European Food Safety Authority. Scientific report of efsa and ecdc: The european union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2010. EFSA J. 2012, 10, 2597. [Google Scholar]
- European Food Safety Authority. Scientific report of efsa and ecdc: The european union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2009. EFSA J. 2011, 9, 2090. [Google Scholar]
- Siddiki, K.F.; Blackburn, S.A. Food poisoning. In Food and famine in the 21st century; Dando, W.A., Ed.; ABC-CLIO, LLC: Santa Barbara, CA, USA, 2012; pp. 199–208. [Google Scholar]
- Rajkovic, A. Microbial toxins and low level of foodborne exposure. Trends Food Sci. Technol. 2014, 38, 149–157. [Google Scholar] [CrossRef]
- Hidalgo, I.J.; Raub, T.J.; Borchardt, R.T. Characterization of the human-colon carcinoma cell-line (Caco-2) as a model system for intestinal epithelial permeability. Gastroenterology 1989, 96, 736–749. [Google Scholar]
- Chantret, I.; Barbat, A.; Dussaulx, E.; Brattain, M.G.; Zweibaum, A. Epithelial polarity, villin expression, and enterocytic differentiation of cultured human-colon carcinoma-cells—A survey of 20 cell-lines. Cancer Res. 1988, 48, 1936–1942. [Google Scholar]
- Neutra, M.; Louvard, A. Differentiation of intestinal cells in vitro. In Modern Cell Biology: Functional Epithelial Cells in Culture; Martlin, K.S., Ed.; Liss: New York, NY, USA, 1989. [Google Scholar]
- Pinto, M.; Robineleon, S.; Appay, M.D.; Kedinger, M.; Triadou, N.; Dussaulx, E.; Lacroix, B.; Simonassmann, P.; Haffen, K.; Fogh, J.; et al. Enterocyte-like differentiation and polarization of the human-colon carcinoma cell-line Caco-2 in culture. Biol. Cell 1983, 47, 323–330. [Google Scholar]
- Richards, I. Principles and Practice of Toxicology in Public Health; Jones and Bartlett Publishers International: London, UK, 2008. [Google Scholar]
- Alschuler, L.N.G.; Gazella, K.A. The Definitive Guide to Cancer, 3rd ed.; The Crown Publishing Group: New York, NY, USA, 2010. [Google Scholar]
- Puerto, M.; Pichardo, S.; Jos, A.; Camean, A.M. Comparison of the toxicity induced by microcystin-rr and microcystin-yr in differentiated and undifferentiated caco-2 cells. Toxicon 2009, 54, 161–169. [Google Scholar] [CrossRef]
- Moal, V.L.L.; Servin, A.L. Pathogenesis of human enterovirulent bacteria: Lessons from cultured, fully differentiated human colon cancer cell lines. Microbiol. Mol. Biol. Rev. 2013, 77, 380–439. [Google Scholar] [CrossRef]
- Virtanen, S.M.; Roivainen, M.; Andersson, M.A.; Ylipaasto, P.; Hoornstra, D.; Mikkola, R.; Salkinoja-Salonen, M.S. In vitro toxicity of cereulide on porcine pancreatic langerhans islets. Toxicon 2008, 51, 1029–1037. [Google Scholar] [CrossRef]
- Andersson, M.A.; Hakulinen, P.; Honkalampi-Hamalainen, U.; Hoornstra, D.; Lhuguenot, J.C.; Maki-Paakkanen, J.; Savolainen, M.; Severin, I.; Stammati, A.L.; Turco, L.; et al. Toxicological profile of cereulide, the bacillus cereus emetic toxin, in functional assays with human, animal and bacterial cells. Toxicon 2007, 49, 351–367. [Google Scholar] [CrossRef]
- Jaaskelainen, E.L.; Teplova, V.; Andersson, M.A.; Andersson, L.C.; Tammela, P.; Andersson, M.C.; Pirhonen, T.I.; Saris, N.E.L.; Vuorela, P.; Salkinoja-Salonen, M.S. In vitro assay for human toxicity of cereulide, the emetic mitochondrial toxin produced by food poisoning bacillus cereus. Toxicol. In Vitro 2003, 17, 737–744. [Google Scholar] [CrossRef]
- Isobe, M.; Ishikawa, T.; Suwan, S.; Agata, N.; Ohta, M. Synthesis and activity of cereulide, a cyclic dodecadepsipeptide ionophore as emetic toxin from bacillus cereus. Bioorg. Med. Chem. Lett. 1995, 5, 2855–2858. [Google Scholar] [CrossRef]
- Keepers, Y.P.; Pizao, P.E.; Peters, G.J.; Vanarkotte, J.; Winograd, B.; Pinedo, H.M. Comparison of the sulforhodamine-b protein and tetrazolium (mtt) assays for invitro chemosensitivity testing. Eur. J. Cancer 1991, 27, 897–900. [Google Scholar] [CrossRef]
- Shappell, N.W. Ergovaline toxicity on caco-2 cells as assessed by mtt, alamarblue, and DNA assays. In Vitro Cell. Dev. Biol. Anim. 2003, 39, 329–335. [Google Scholar] [CrossRef]
- Zagari, F.; Jordan, M.; Stettler, M.; Broly, H.; Wurm, F.M. Lactate metabolism shift in cho cell culture: The role of mitochondrial oxidative activity. New Biotechnol. 2013, 30, 238–245. [Google Scholar] [CrossRef]
- Andersson, M.A.; Mikkola, R.; Rasimus, S.; Hoornstra, D.; Salin, P.; Rahkila, R.; Heikkinen, M.; Mattila, S.; Peltola, J.; Kalso, S.; et al. Boar spermatozoa as a biosensor for detecting toxic substances in indoor dust and aerosols. Toxicol. In Vitro 2010, 24, 2041–2052. [Google Scholar] [CrossRef]
- Rajkovic, A.; Uyttendaele, M.; Debevere, J. Computer aided boar semen motility analysis for cereulide detection in different food matrices. Int. J. Food Microbiol. 2007, 114, 92–99. [Google Scholar] [CrossRef]
- Rajkovic, A.; Uyttendaele, M.; Deley, W.; van Soom, A.; Rijsselaere, T.; Debevere, J. Dynamics of boar semen motility inhibition as a semi-quantitative measurement of bacillus cereus emetic toxin (cereulide). J. Microbiol. Methods 2006, 65, 525–534. [Google Scholar] [CrossRef]
- Delbrassinne, L.; Andjelkovic, M.; Rajkovic, A.; Dubois, P.; Nguessan, E.; Mahillon, J.; Van Loco, J. Determination of bacillus cereus emetic toxin in food products by means of lc-msa(2). Food Anal. Method 2012, 5, 969–979. [Google Scholar] [CrossRef]
- Rajkovic, A.; Uyttendaele, M.; Ombregt, S.A.; Jaaskelainen, E.; Salkinoja-Salonen, M.; Debevere, J. Influence of type of food on the kinetics and overall production of bacillus cereus emetic toxin. J. Food Prot. 2006, 69, 847–852. [Google Scholar]
- Delbrassinne, L.; Andjelkovic, M.; Rajkovic, A.; Bottledoorn, N.; Mahillon, J.; van Loco, J. Follow-up of the bacillus cereus emetic toxin production in penne pasta under household conditions using liquid chromatography coupled with mass spectrometry. Food Microbiol. 2011, 28, 1105–1109. [Google Scholar] [CrossRef]
- Bouchoux, J.; Beilstein, F.; Pauquai, T.; Guerrera, I.C.; Chateau, D.; Ly, N.; Alqub, M.; Klein, C.; Chambaz, J.; Rousset, M.; et al. The proteome of cytosolic lipid droplets isolated from differentiated Caco-2/tc7 enterocytes reveals cell-specific characteristics. Biol. Cell 2011, 103, 499–517. [Google Scholar] [CrossRef]
- Tempelaars, M.H.; Rodrigues, S.; Abee, T. Comparative analysis of antimicrobial activities of valinomycin and cereulide, the bacillus cereus emetic toxin. Appl. Environ. Microbiol. 2011, 77, 2755–2762. [Google Scholar] [CrossRef]
- Chen, Y.L.S.; Li, J.H.; Yu, C.Y.; Lin, C.J.; Chiu, P.H.; Chen, P.W.; Lin, C.C.; Chen, W.J. Novel cationic antimicrobial peptide gw-h1 induced caspase-dependent apoptosis of hepatocellular carcinoma cell lines. Peptides 2012, 36, 257–265. [Google Scholar] [CrossRef]
- Da Costa, A.N.; Mijal, R.S.; Keen, J.N.; Findlay, J.B.C.; Wild, C.P. Proteomic analysis of the effects of the immunomodulatory mycotoxin deoxynivalenol. Proteomics 2011, 11, 1903–1914. [Google Scholar] [CrossRef]
- Razafimanjato, H.; Benzaria, A.; Taieb, N.; Guo, X.J.; Vidal, N.; Di Scala, C.; Varini, K.; Maresca, M. The ribotoxin deoxynivalenol affects the viability and functions of glial cells. Glia 2011, 59, 1672–1683. [Google Scholar] [CrossRef]
- Maresca, M. From the gut to the brain: Journey and pathophysiological effects of the food-associated trichothecene mycotoxin deoxynivalenol. Toxins 2013, 5, 784–820. [Google Scholar] [CrossRef]
- Bonnet, R.; Pavlovic, S.; Lehmann, J.; Rommelspacher, H. The strong inhibition of triosephosphate isomerase by the natural beta-carbolines may explain their neurotoxic actions. Neuroscience 2004, 127, 443–453. [Google Scholar] [CrossRef]
- Duan, Z.; Lamendola, D.E.; Yusuf, R.Z.; Penson, R.T.; Preffer, F.I.; Seiden, M.V. Overexpression of human phosphoglycerate kinase 1 (PGK1) induces a multidrug resistance phenotype. Anticancer Res. 2002, 22, 1933–1941. [Google Scholar]
- Chandra, H.; Gupta, P.K.; Sharma, K.; Mattoo, A.R.; Garg, S.K.; Gade, W.N.; Sirdeshmukh, R.; Maithal, K.; Singh, Y. Proteome analysis of mouse macrophages treated with anthrax lethal toxin. Bba Proteins Proteom 2005, 1747, 151–159. [Google Scholar] [CrossRef]
- Lehti, M.; Donelan, E.; Abplanalp, W.; Al-Massadi, O.; Habegger, K.M.; Weber, J.; Ress, C.; Mansfeld, J.; Somvanshi, S.; Trivedi, C.; et al. High-density lipoprotein maintains skeletal muscle function by modulating cellular respiration in mice. Circulation 2013, 128, 2364–2371. [Google Scholar] [CrossRef]
- Allen, A.M.; Graham, A. Mitochondrial function is involved in regulation of cholesterol efflux to apolipoprotein (apo)A-I from murine RAW 264.7 macrophages. Lipids Health Dis. 2012, 11. [Google Scholar] [CrossRef]
- Suzuki, M.M.; Matsumoto, M.; Omi, H.; Kobayashi, T.; Nakamura, A.; Kishi, H.; Kobayashi, S.; Takagi, T. Interaction of peptide-bound beads with lipopolysaccharide and lipoproteins. J. Microbiol. Methods 2014, 100, 137–141. [Google Scholar] [CrossRef]
- Henning, M.F.; Herlax, V.; Bakás, L. Contribution of the c-terminal end of apolipoprotein ai to neutralization of lipopolysaccharide endotoxic effect. Innate Immun. 2011, 17, 327–337. [Google Scholar] [CrossRef]
- Ma, J.; Liao, X.-L.; Lou, B.; Wu, M.-P. Role of apolipoprotein A-I in protecting against endotoxin toxicity. Acta Biochim. Biophys. Sin. 2004, 36, 419–424. [Google Scholar]
- Jiao, Y.-L.; Wu, M.-P. Apolipoprotein A-I diminishes acute lung injury and sepsis in mice induced by lipoteichoic acid. Cytokine 2008, 43, 83–87. [Google Scholar] [CrossRef]
- Wasan, K.M.; Morton, R.E.; Rosenblum, M.G.; Lopez-Berestein, G. Decreased toxicity of liposomal amphotericin B due to association of amphotericin B with high-density lipoproteins: Role of lipid transfer protein. J. Pharm. Sci. 1994, 83, 1006–1010. [Google Scholar] [CrossRef]
- Contag, B. Epigenetic effectiveness of complete carcinogens: Specific interactions of polycyclic aromatic hydrocarbons and aminoazo dyes with cholesterol and apolipoprotein A-I. Z. Naturforsch. C 2005, 60, 799–806. [Google Scholar]
- Thaveeratitham, P.; Plengpanich, W.; Naen-Udorn, W.; Patumraj, S.; Khovidhunkit, W. Effects of human apolipoprotein A-I on endotoxin-induced leukocyte adhesion on endothelial cells in vivo and on the growth of escherichia coli in vitro. J. Endotoxin Res. 2007, 13, 58–64. [Google Scholar] [CrossRef]
- Stein, E.A.; Honarpour, N.; Wasserman, S.M.; Xu, F.; Scott, R.; Raal, F.J. Effect of the proprotein convertase subtilisin/kexin 9 monoclonal antibody, AMG 145, in homozygous familial hypercholesterolemia. Circulation 2013, 128, 2113–2120. [Google Scholar] [CrossRef]
- Johnson, R.M.; Ho, Y.S.; Yu, D.Y.; Kuypers, F.A.; Ravindranath, Y.; Goyette, G.W. The effects of disruption of genes for peroxiredoxin-2, glutathione peroxidase-1, and catalase on erythrocyte oxidative metabolism. Free Radic. Biol. Med. 2010, 48, 519–525. [Google Scholar] [CrossRef]
- Buratti, F.M.; Scardala, S.; Funari, E.; Testai, E. Human glutathione transferases catalyzing the conjugation of the hepatoxin microcystin-lr. Chem. Res. Toxicol. 2011, 24, 926–933. [Google Scholar] [CrossRef]
- Fatemi, F.; Allameh, A.; Dadkhah, A.; Forouzandeh, M.; Kazemnejad, S.; Sharifi, R. Changes in hepatic cytosolic glutathione s-transferase activity and expression of its class-p during prenatal and postnatal period in rats treated with aflatoxin b1. Arch. Toxicol. 2006, 80, 572–579. [Google Scholar] [CrossRef]
- Tiessen, C.; Fehr, M.; Schwarz, C.; Baechler, S.; Domnanich, K.; Böttler, U.; Pahlke, G.; Marko, D. Modulation of the cellular redox status by the alternaria toxins alternariol and alternariol monomethyl ether. Toxicol. Lett. 2013, 216, 23–30. [Google Scholar] [CrossRef]
- Choi, S.; Park, S.-Y.; Jeong, J.; Cho, E.; Phark, S.; Lee, M.; Kwak, D.; Lim, J.-Y.; Jung, W.-W.; Sul, D. Identification of toxicological biomarkers of di(2-ethylhexyl) phthalate in proteins secreted by HEPG2 cells using proteomic analysis. Proteomics 2010, 10, 1831–1846. [Google Scholar] [CrossRef]
- Roh, T.; Kwak, M.Y.; Kwak, E.H.; Kim, D.H.; Han, E.Y.; Bae, J.Y.; Bang, D.Y.; Lim, D.S.; Ahn, I.Y.; Jang, D.E.; et al. Chemopreventive mechanisms of methionine on inhibition of benzo(a)pyrene–DNA adducts formation in human hepatocellular carcinoma hepg2 cells. Toxicol. Lett. 2012, 208, 232–238. [Google Scholar] [CrossRef]
- Aktories, K.; Schwan, C.; Papatheodorou, P.; Lang, A.E. Bidirectional attack on the actin cytoskeleton. Bacterial protein toxins causing polymerization or depolymerization of actin. Toxicon 2012, 60, 572–581. [Google Scholar] [CrossRef]
- Barbieri, J.T.; Riese, M.J.; Aktories, K. Bacterial toxins that modify the actin cytoskeleton. Annu. Rev. Cell Dev. Biol. 2002, 18, 315–344. [Google Scholar] [CrossRef]
- Satin, B.; Norais, N.; Rappuoli, R.; Telford, J.; Murgia, M.; Montecucco, C.; Papini, E. Effect of helicobacter pylori vacuolating toxin on maturation and extracellular release of procathepsin d and on epidermal growth factor degradation. J. Biol. Chem. 1997, 272, 25022–25028. [Google Scholar]
- Suleiman, S.A. Petroleum hydrocarbon toxicity in vitro: Effect of n-alkanes, benzene and toluene on pulmonary alveolar macrophages and lysosomal enzymes of the lung. Arch. Toxicol. 1987, 59, 402–407. [Google Scholar] [CrossRef]
- Yamada, A.; Yamamoto, T.; Yamazaki, N.; Yamashita, K.; Kataoka, M.; Nagata, T.; Terada, H.; Shinohara, Y. Differential permeabilization effects of Ca2+ and valinomycin on the inner and outer mitochondrial membranes as revealed by proteomics analysis of proteins released from mitochondria. Mol. Cell. Proteomics 2009, 8, 1265–1277. [Google Scholar] [CrossRef]
- El Hage, T.; Lorin, S.; Decottignies, P.; Djavaheri-Mergny, M.; Authier, F. Proteolysis of pseudomonas exotoxin A within hepatic endosomes by cathepsins B and D produces fragments displaying in vitro ADP-ribosylating and apoptotic effects. FEBS J. 2010, 277, 3735–3749. [Google Scholar] [CrossRef]
- Kim, S.; Ock, J.; Kim, A.K.; Lee, H.W.; Cho, J.-Y.; Kim, D.R.; Park, J.-Y.; Suk, K. Neurotoxicity of microglial cathepsin D revealed by secretome analysis. J Neurochem. 2007, 103, 2640–2650. [Google Scholar]
- Metaye, T.; Kraimps, J.L.; Goujon, J.M.; Fernandez, B.; Quellard, N.; Ingrand, P.; Barbier, J.; Begon, F. Expression, localization, and thyrotropin regulation of cathepsin d in human thyroid tissues. J. Clin. Endocrinol. Metab. 1997, 82, 3383–3388. [Google Scholar] [CrossRef]
- Lah, T.T.; Kalman, E.; Najjar, D.; Gorodetsky, E.; Brennan, P.; Somers, R.; Daskal, I. Cells producing cathepsins D, B, and L in human breast carcinoma and their association with prognosis. Hum. Pathol. 2000, 31, 149–160. [Google Scholar] [CrossRef]
- Grant, M.H.; Rodgers, E.H.; Anderson, K.; Haydon, G.H.; Hayes, P.C. The effect of serum from liver cancer patients on the growth and function of primary and immortalised hepatocytes. Int. J. Artif. Organs 2001, 24, 807–813. [Google Scholar]
- Li, J.; Zhang, D.; Jefferson, P.A.; Ward, K.M.; Ayene, I.S. A bioactive probe for glutathione-dependent antioxidant capacity in breast cancer patients: Implications in measuring biological effects of arsenic compounds. J. Pharmacol. Toxicol. Methods 2014, 69, 39–48. [Google Scholar] [CrossRef]
- Hoensch, H.; Peters, W.H.M.; Roelofs, H.M.J.; Kirch, W. Expression of the glutathione enzyme systemof human colon mucosa by localisation, gender and age. Curr. Med. Res. Opin. 2006, 22, 1075–1083. [Google Scholar] [CrossRef]
- Fella, K.; Glückmann, M.; Hellmann, J.; Karas, M.; Kramer, P.-J.; Kröger, M. Use of two-dimensional gel electrophoresis in predictive toxicology: Identification of potential early protein biomarkers in chemically induced hepatocarcinogenesis. Proteomics 2005, 5, 1914–1927. [Google Scholar] [CrossRef]
- Balliet, R.M.; Capparelli, C.; Guido, C.; Pestell, T.G.; Martinez-Outschoorn, U.E.; Lin, Z.; Whitaker-Menezes, D.; Chiavarina, B.; Pestell, R.G.; Howell, A.; et al. Mitochondrial oxidative stress in cancer-associated fibroblasts drives lactate production, promoting breast cancer tumor growth understanding the aging and cancer connection. Cell Cycle 2011, 10, 4065–4073. [Google Scholar] [CrossRef]
- Natoli, M.; Leoni, B.D.; D’Agnano, I.; D’Onofrio, M.; Brandi, R.; Arisi, I.; Zucco, F.; Felsani, A. Cell growing density affects the structural and functional properties of Caco-2 differentiated monolayer. J. Cell. Physiol. 2011, 226, 1531–1543. [Google Scholar] [CrossRef]
- Natoli, M.; Leoni, B.D.; D’Agnano, I.; Zucco, F.; Felsani, A. Good Caco-2 cell culture practices. Toxicol. In Vitro 2012, 26, 1243–1246. [Google Scholar] [CrossRef]
- Biesta-Peters, E.G.; Reij, M.W.; Blaauw, R.H.; in’t Veld, P.H.; Rajkovic, A.; Ehling-Schulz, M.; Abee, T. Quantification of the emetic toxin cereulide in food products by liquid chromatography-mass spectrometry using synthetic cereulide as a standard. Appl. Environ. Microbiol. 2010, 76, 7466–7472. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Chevallet, M.; Diemer, H.; van Dorssealer, A.; Villiers, C.; Rabilloud, T. Toward a better analysis of secreted proteins: The example of the myeloid cells secretome. Proteomics 2007, 7, 1757–1770. [Google Scholar] [CrossRef] [Green Version]
- O’Farrell, P.H. High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem. 1975, 250, 4007–4021. [Google Scholar]
- Anon. PDQuest. Bio-Rad Laboratories. Available online: http://www.bio-rad.com/LifeScience/pdf/DiscoverySeries/PDQuest_v74.pdf (accessed on 24 July 2014).
- Laemmli, U. Cleavage of structural proteins during the assembly of the head of bacteriophage t4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins from silver stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef]
- Perkins, D.N.; Pappin, D.J.C.; Creasy, D.M.; Cottrell, J.S. Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophoresis 1999, 20, 3551–3567. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rajkovic, A.; Grootaert, C.; Butorac, A.; Cucu, T.; De Meulenaer, B.; Van Camp, J.; Bracke, M.; Uyttendaele, M.; Bačun-Družina, V.; Cindrić, M. Sub-Emetic Toxicity of Bacillus cereus Toxin Cereulide on Cultured Human Enterocyte-Like Caco-2 Cells. Toxins 2014, 6, 2270-2290. https://doi.org/10.3390/toxins6082270
Rajkovic A, Grootaert C, Butorac A, Cucu T, De Meulenaer B, Van Camp J, Bracke M, Uyttendaele M, Bačun-Družina V, Cindrić M. Sub-Emetic Toxicity of Bacillus cereus Toxin Cereulide on Cultured Human Enterocyte-Like Caco-2 Cells. Toxins. 2014; 6(8):2270-2290. https://doi.org/10.3390/toxins6082270
Chicago/Turabian StyleRajkovic, Andreja, Charlotte Grootaert, Ana Butorac, Tatiana Cucu, Bruno De Meulenaer, John Van Camp, Marc Bracke, Mieke Uyttendaele, Višnja Bačun-Družina, and Mario Cindrić. 2014. "Sub-Emetic Toxicity of Bacillus cereus Toxin Cereulide on Cultured Human Enterocyte-Like Caco-2 Cells" Toxins 6, no. 8: 2270-2290. https://doi.org/10.3390/toxins6082270