Botulinum Neurotoxin A Complex Recognizes Host Carbohydrates through Its Hemagglutinin Component

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussions
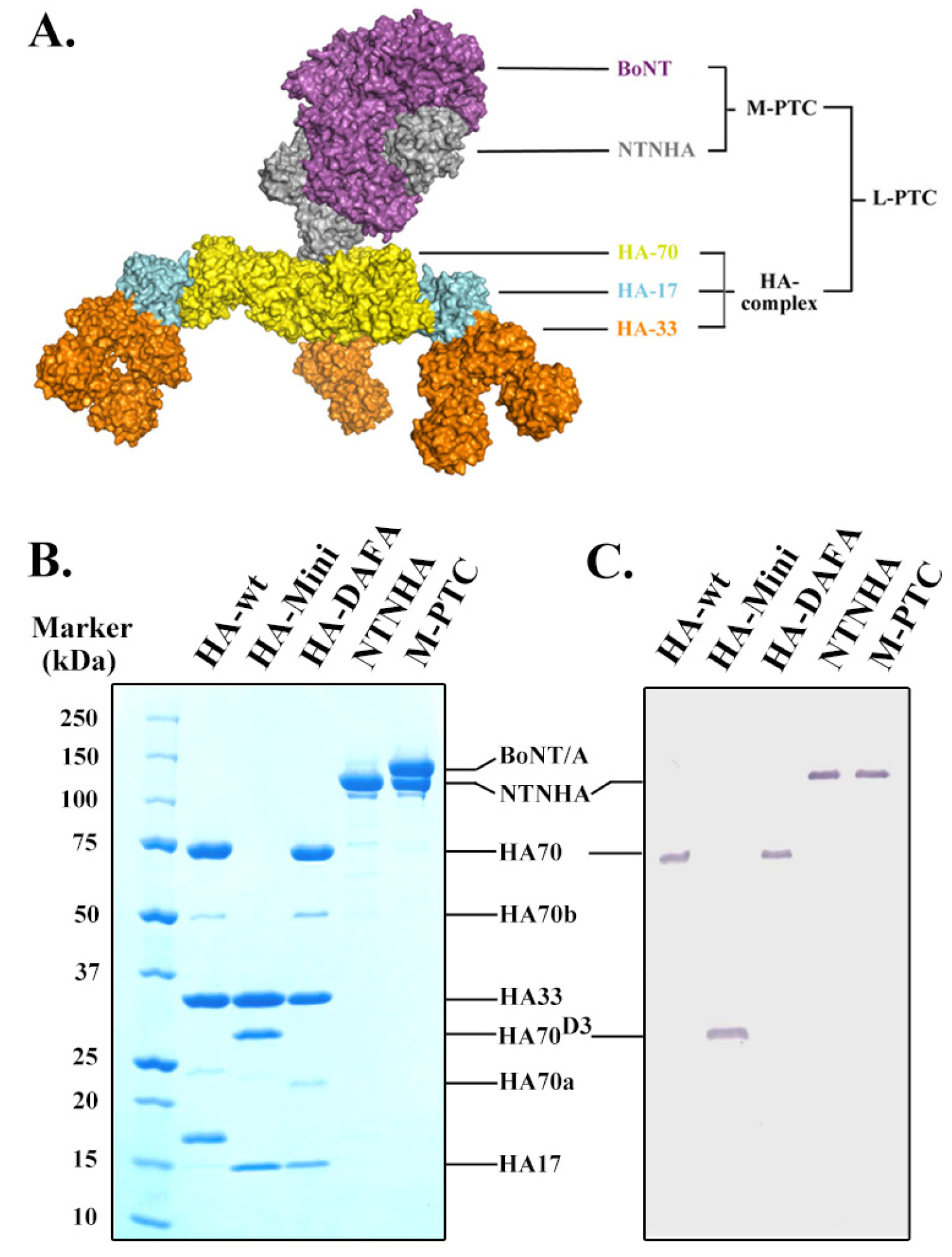
2.1. Homogeneous Recombinant NAPs Are Unique Molecular Probes for Functional Studies
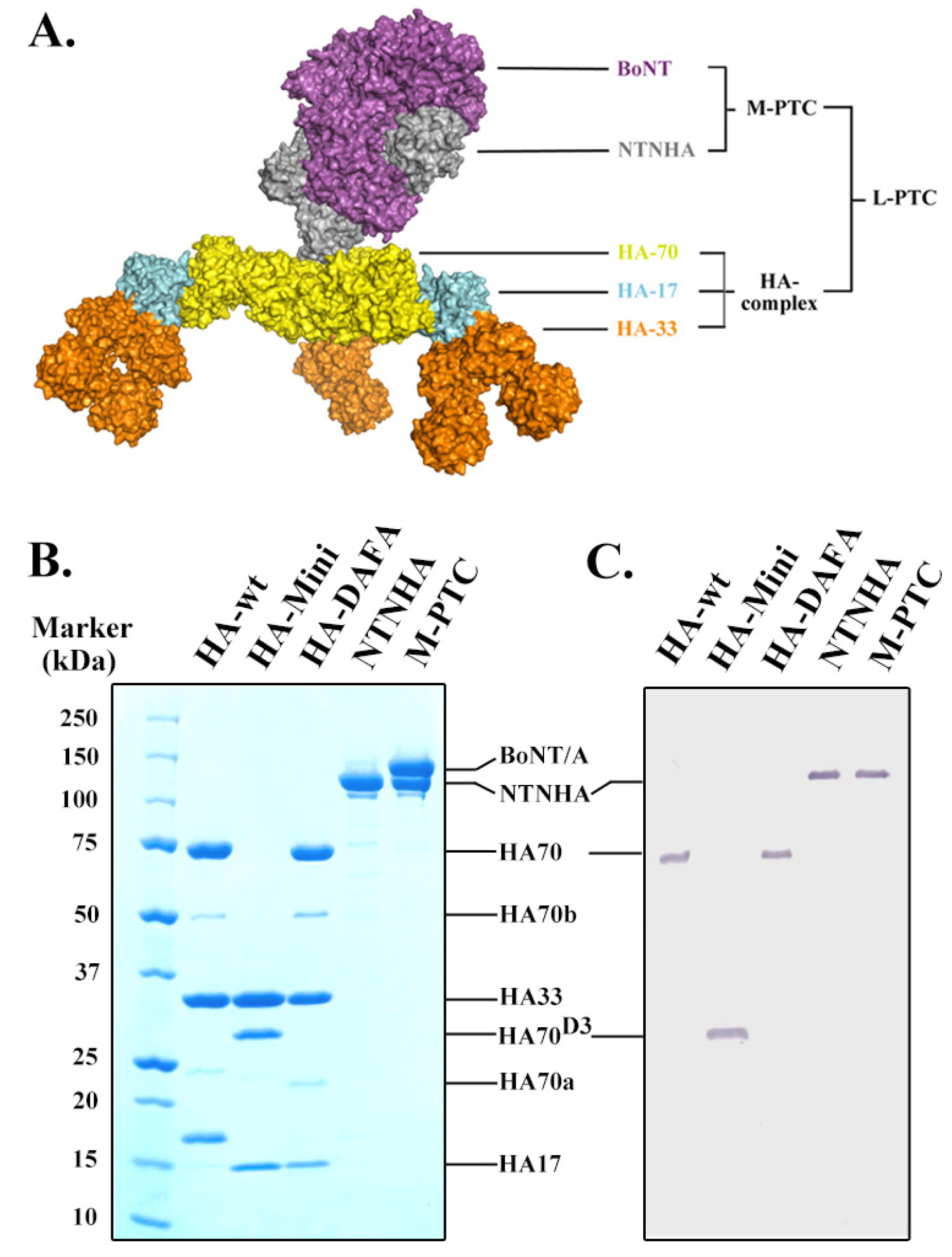
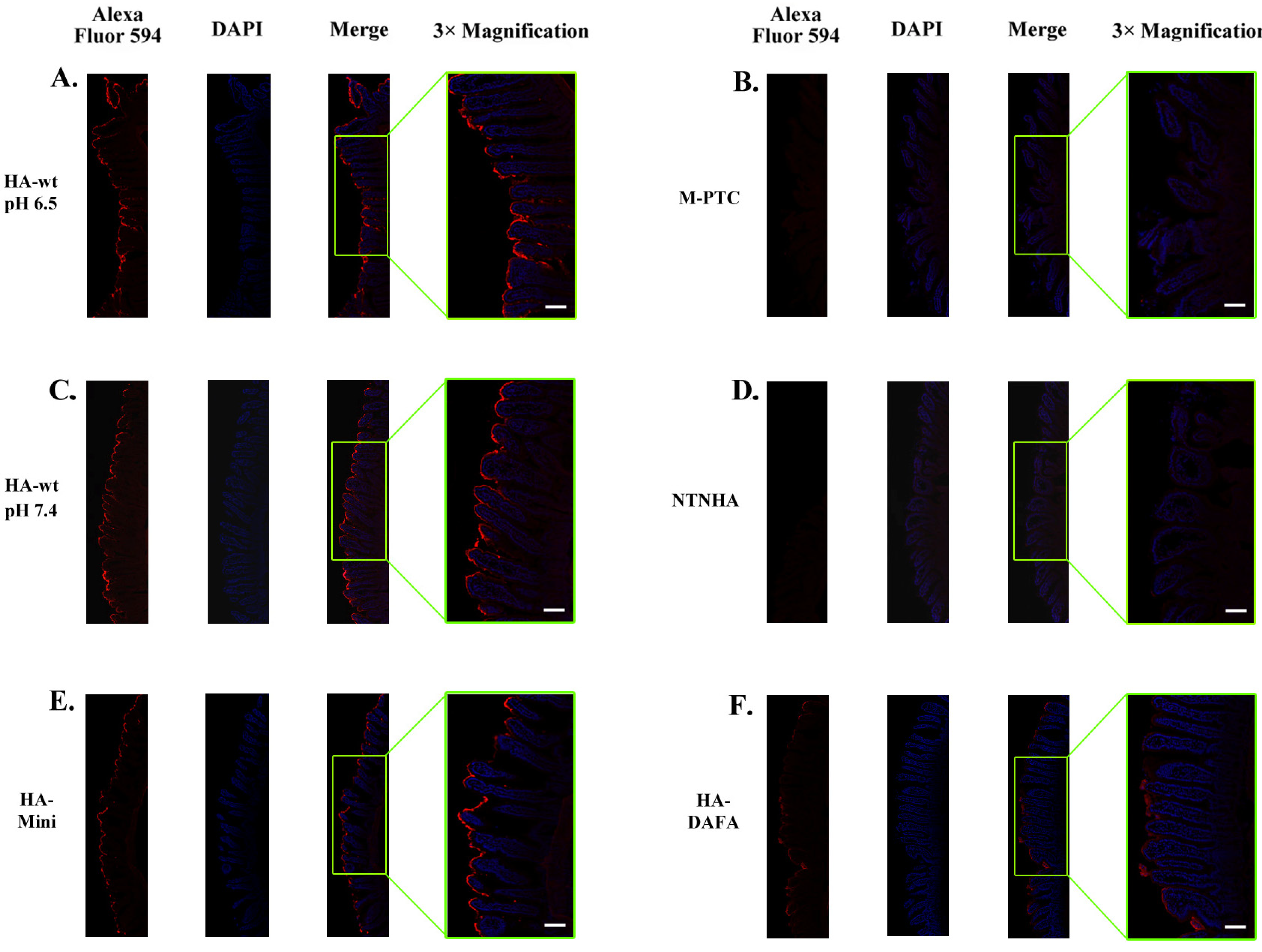
2.2. The HA Complex Mediates the Intestinal Binding of the PTC
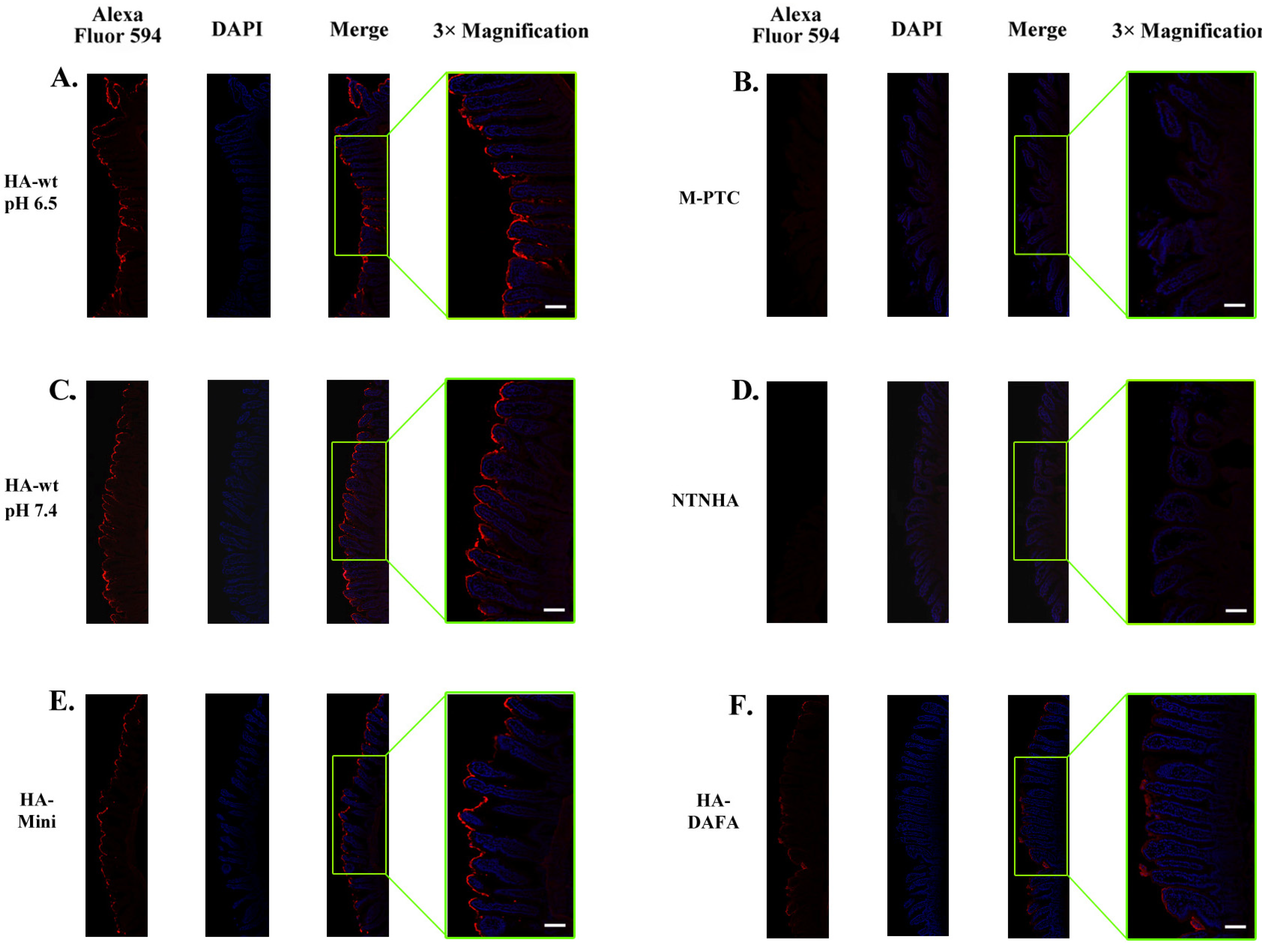
2.3. The Intestinal Binding of the PTC Relies on Carbohydrate Receptors
2.4. The Specific HA–Carbohydrate Recognition Primarily Depends on the Terminal Saccharides

3. Experimental Section
3.1. Cloning, Protein Expression, and Purification
3.2. Immunoblotting Assay
3.3. Ex Vivo Intestinal Binding Assay
3.4. Tissue Preparation and Immunostaining
3.5. Glycan Array Screening
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Bigalke, H.; Rummel, A. Medical aspects of toxin weapons. Toxicology 2005, 214, 210–220. [Google Scholar] [CrossRef]
- Arnon, S.S.; Schechter, R.; Inglesby, T.V.; Henderson, D.A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Fine, A.D.; Hauer, J.; Layton, M.; et al. Botulinum toxin as a biological weapon: Medical and public health management. Jama 2001, 285, 1059–1070. [Google Scholar] [CrossRef]
- Schiavo, G.; Benfenati, F.; Poulain, B.; Rossetto, O.; Polverino de Laureto, P.; DasGupta, B.R.; Montecucco, C. Tetanus and botulinum-B neurotoxins block neurotransmitter release by proteolytic cleavage of synaptobrevin. Nature 1992, 359, 832–835. [Google Scholar] [CrossRef]
- Blasi, J.; Chapman, E.R.; Link, E.; Binz, T.; Yamasaki, S.; de Camilli, P.; Sudhof, T.C.; Niemann, H.; Jahn, R. Botulinum neurotoxin A selectively cleaves the synaptic protein SNAP-25. Nature 1993, 365, 160–163. [Google Scholar] [CrossRef]
- Schiavo, G.; Matteoli, M.; Montecucco, C. Neurotoxins affecting neuroexocytosis. Physiol. Rev. 2000, 80, 717–766. [Google Scholar]
- Hill, K.K.; Smith, T.J. Genetic diversity within Clostridium botulinum serotypes, botulinum neurotoxin gene clusters and toxin subtypes. Curr. Top. Microbiol. Immunol. 2013, 364, 1–20. [Google Scholar]
- Dover, N.; Barash, J.R.; Hill, K.K.; Xie, G.; Arnon, S.S. Molecular characterization of a novel botulinum neurotoxin type H gene. J. Infect. Dis. 2014, 209, 192–202. [Google Scholar]
- Collins, M.D.; East, A.K. Phylogeny and taxonomy of the food-borne pathogen Clostridium botulinum and its neurotoxins. J. Appl. Microbiol. 1998, 84, 5–17. [Google Scholar]
- Lin, G.; Tepp, W.H.; Pier, C.L.; Jacobson, M.J.; Johnson, E.A. Expression of the Clostridium botulinum A2 neurotoxin gene cluster proteins and characterization of the A2 complex. Appl. Environ. Microbiol. 2010, 76, 40–47. [Google Scholar]
- Couesnon, A.; Pereira, Y.; Popoff, M.R. Receptor-mediated transcytosis of botulinum neurotoxin a through intestinal cell monolayers. Cell Microbiol. 2008, 10, 375–387. [Google Scholar]
- Couesnon, A.; Shimizu, T.; Popoff, M.R. Differential entry of botulinum neurotoxin A into neuronal and intestinal cells. Cell Microbiol. 2009, 11, 289–308. [Google Scholar] [CrossRef]
- Couesnon, A.; Molgo, J.; Connan, C.; Popoff, M.R. Preferential entry of botulinum neurotoxin A Hc domain through intestinal crypt cells and targeting to cholinergic neurons of the mouse intestine. PLoS Pathog. 2012, 8, e1002583. [Google Scholar] [CrossRef]
- Cheng, L.W.; Onisko, B.; Johnson, E.A.; Reader, J.R.; Griffey, S.M.; Larson, A.E.; Tepp, W.H.; Stanker, L.H.; Brandon, D.L.; Carter, J.M. Effects of purification on the bioavailability of botulinum neurotoxin type A. Toxicology 2008, 249, 123–129. [Google Scholar] [CrossRef]
- Sakaguchi, G. Clostridium botulinum toxins. Pharmacol. Ther. 1982, 19, 165–194. [Google Scholar] [CrossRef]
- Ohishi, I.; Sugii, S.; Sakaguchi, G. Oral toxicities of Clostridium botulinum toxins in response to molecular size. Infect. Immun. 1977, 16, 107–109. [Google Scholar]
- Gu, S.; Rumpel, S.; Zhou, J.; Strotmeier, J.; Bigalke, H.; Perry, K.; Shoemaker, C.B.; Rummel, A.; Jin, R. Botulinum neurotoxin is shielded by NTNHA in an interlocked complex. Science 2012, 335, 977–981. [Google Scholar] [CrossRef]
- Lee, K.; Gu, S.; Jin, L.; Le, T.T.; Cheng, L.W.; Strotmeier, J.; Kruel, A.M.; Yao, G.; Perry, K.; et al. Structure of a bimodular botulinum neurotoxin complex provides insights into its oral toxicity. PLoS Pathog. 2013, 9, e1003690. [Google Scholar] [CrossRef]
- Amatsu, S.; Sugawara, Y.; Matsumura, T.; Kitadokoro, K.; Fujinaga, Y. Crystal structure of Clostridium botulinum whole hemagglutinin reveals a huge triskelion-shaped molecular complex. J. Biol. Chem. 2013, 288, 35617–35625. [Google Scholar] [CrossRef]
- Maksymowych, A.B.; Reinhard, M.; Malizio, C.J.; Goodnough, M.C.; Johnson, E.A.; Simpson, L.L. Pure botulinum neurotoxin is absorbed from the stomach and small intestine and produces peripheral neuromuscular blockade. Infect. Immun. 1999, 67, 4708–4712. [Google Scholar]
- Bonventre, P.F. Absorption of botulinal toxin from the gastrointestinal tract. Rev. Infect. Dis. 1979, 1, 663–667. [Google Scholar] [CrossRef]
- Gu, S.; Jin, R. Assembly and function of the botulinum neurotoxin progenitor complex. Curr. Top. Microbiol. Immunol. 2013, 364, 21–44. [Google Scholar]
- Fujinaga, Y.; Inoue, K.; Watanabe, S.; Yokota, K.; Hirai, Y.; Nagamachi, E.; Oguma, K. The haemagglutinin of Clostridium botulinum type C progenitor toxin plays an essential role in binding of toxin to the epithelial cells of guinea pig small intestine, leading to the efficient absorption of the toxin. Microbiology 1997, 143, 3841–3847. [Google Scholar] [CrossRef]
- Nishikawa, A.; Uotsu, N.; Arimitsu, H.; Lee, J.C.; Miura, Y.; Fujinaga, Y.; Nakada, H.; Watanabe, T.; Ohyama, T.; Sakano, Y.; et al. The receptor and transporter for internalization of Clostridium botulinum type C progenitor toxin into HT-29 cells. Biochem. Biophys. Res. Commun. 2004, 319, 327–333. [Google Scholar] [CrossRef]
- Kojima, S.; Eguchi, H.; Ookawara, T.; Fujiwara, N.; Yasuda, J.; Nakagawa, K.; Yamamura, T.; Suzuki, K. Clostridium botulinum type A progenitor toxin binds to Intestine-407 cells via N-acetyllactosamine moiety. Biochem. Biophys. Res. Commun. 2005, 331, 571–576. [Google Scholar] [CrossRef]
- Uotsu, N.; Nishikawa, A.; Watanabe, T.; Ohyama, T.; Tonozuka, T.; Sakano, Y.; Oguma, K. Cell internalization and traffic pathway of Clostridium botulinum type C neurotoxin in HT-29 cells. Biochim. Biophys. Acta 2006, 1763, 120–128. [Google Scholar]
- Nakamura, T.; Takada, N.; Tonozuka, T.; Sakano, Y.; Oguma, K.; Nishikawa, A. Binding properties of Clostridium botulinum type C progenitor toxin to mucins. Biochim. Biophys. Acta 2007, 1770, 551–555. [Google Scholar]
- Home: Fuctional Glycomics Gateway. Available online: http://www.functionalglycomics.org/ (accessed on 3 December 2013).
- Heimburg-Molinaro, J.; Song, X.; Smith, D.F.; Cummings, R.D. Preparation and analysis of glycan microarrays. Curr. Protoc. Protein Sci. 2011. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yao, G.; Lee, K.; Gu, S.; Lam, K.-H.; Jin, R. Botulinum Neurotoxin A Complex Recognizes Host Carbohydrates through Its Hemagglutinin Component. Toxins 2014, 6, 624-635. https://doi.org/10.3390/toxins6020624
Yao G, Lee K, Gu S, Lam K-H, Jin R. Botulinum Neurotoxin A Complex Recognizes Host Carbohydrates through Its Hemagglutinin Component. Toxins. 2014; 6(2):624-635. https://doi.org/10.3390/toxins6020624
Chicago/Turabian StyleYao, Guorui, Kwangkook Lee, Shenyan Gu, Kwok-Ho Lam, and Rongsheng Jin. 2014. "Botulinum Neurotoxin A Complex Recognizes Host Carbohydrates through Its Hemagglutinin Component" Toxins 6, no. 2: 624-635. https://doi.org/10.3390/toxins6020624