Dog Poisonings Associated with a Microcystis aeruginosa Bloom in the Netherlands



Abstract
:1. Introduction
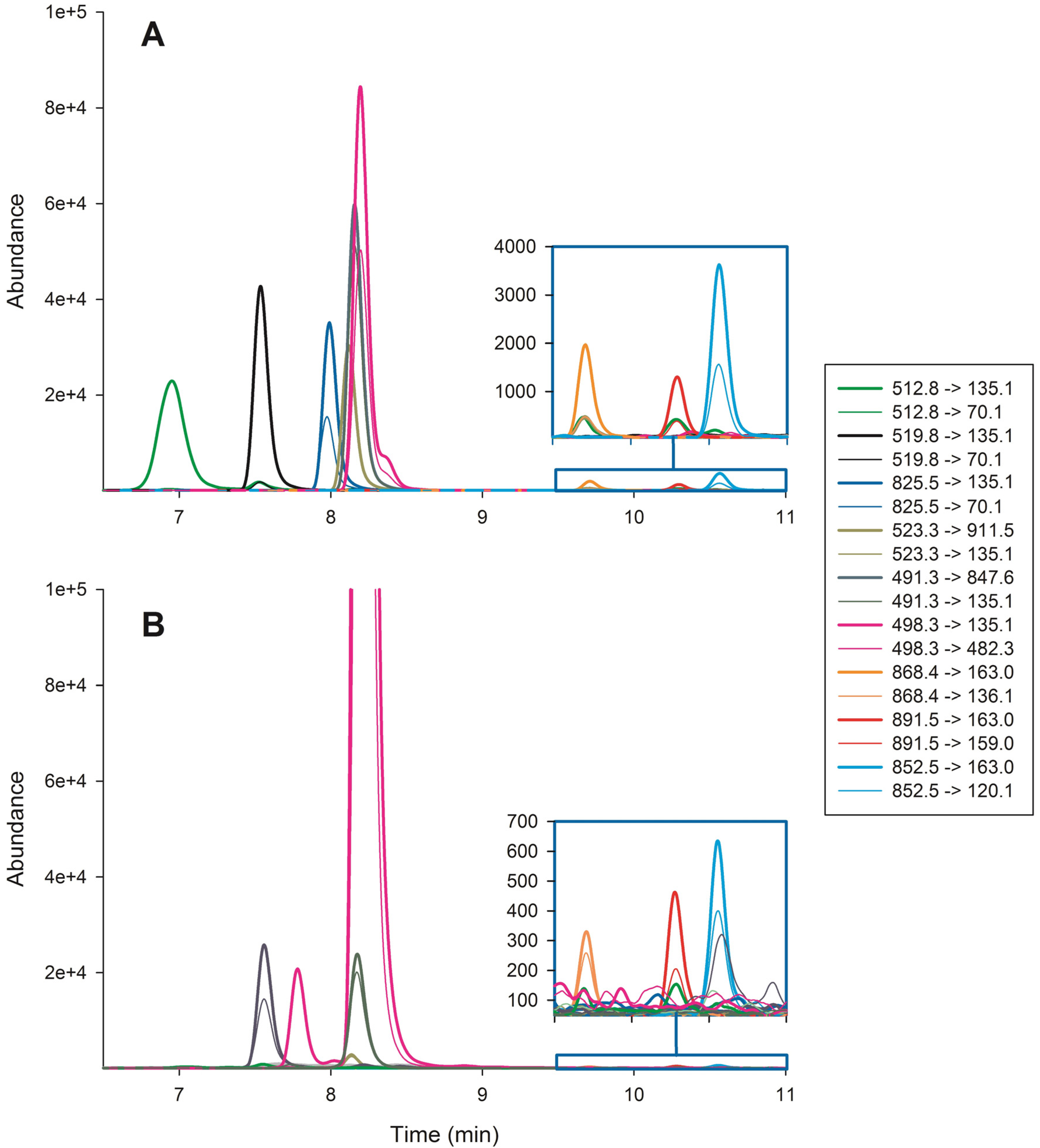
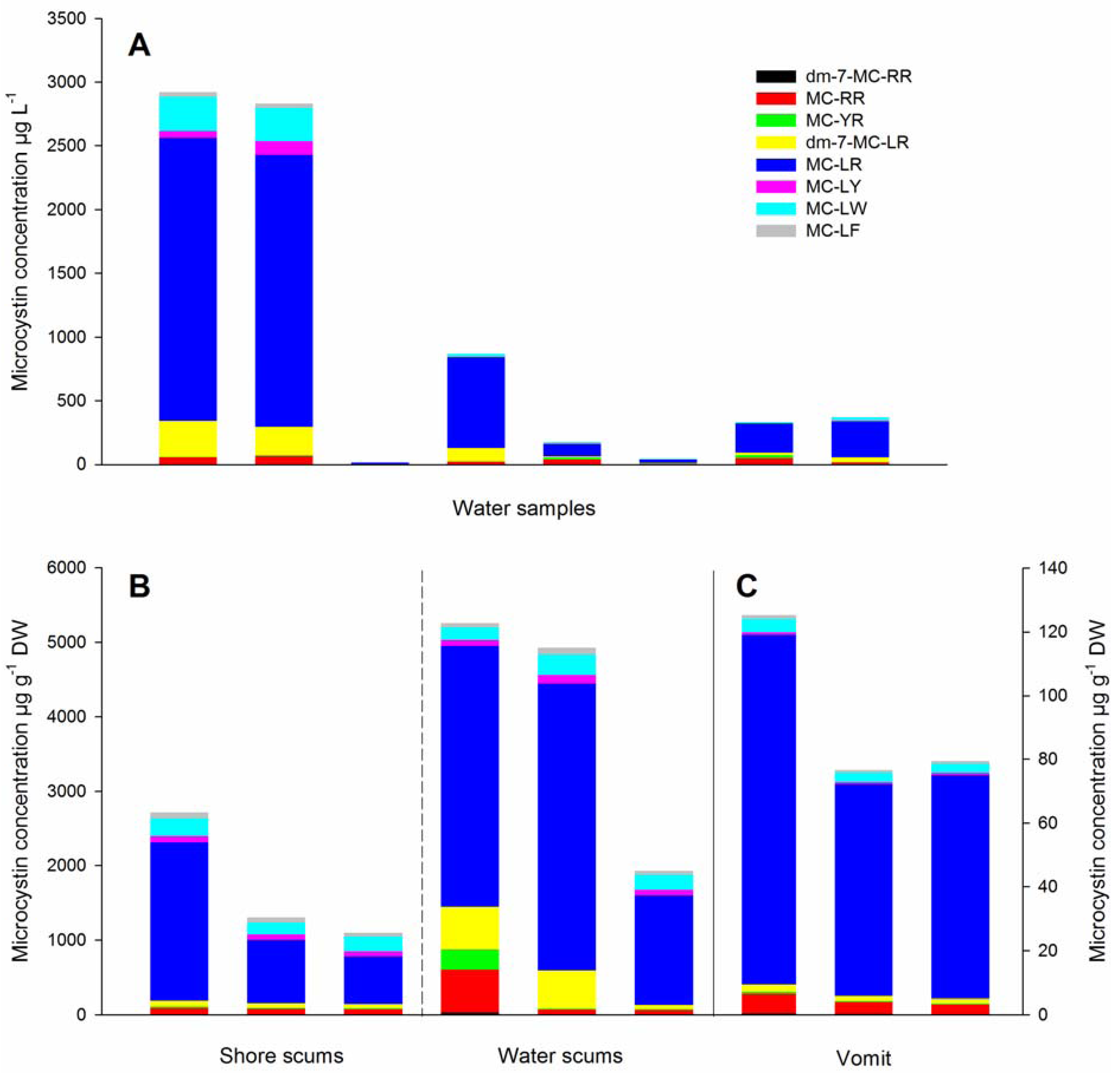
2. Results and Discussion

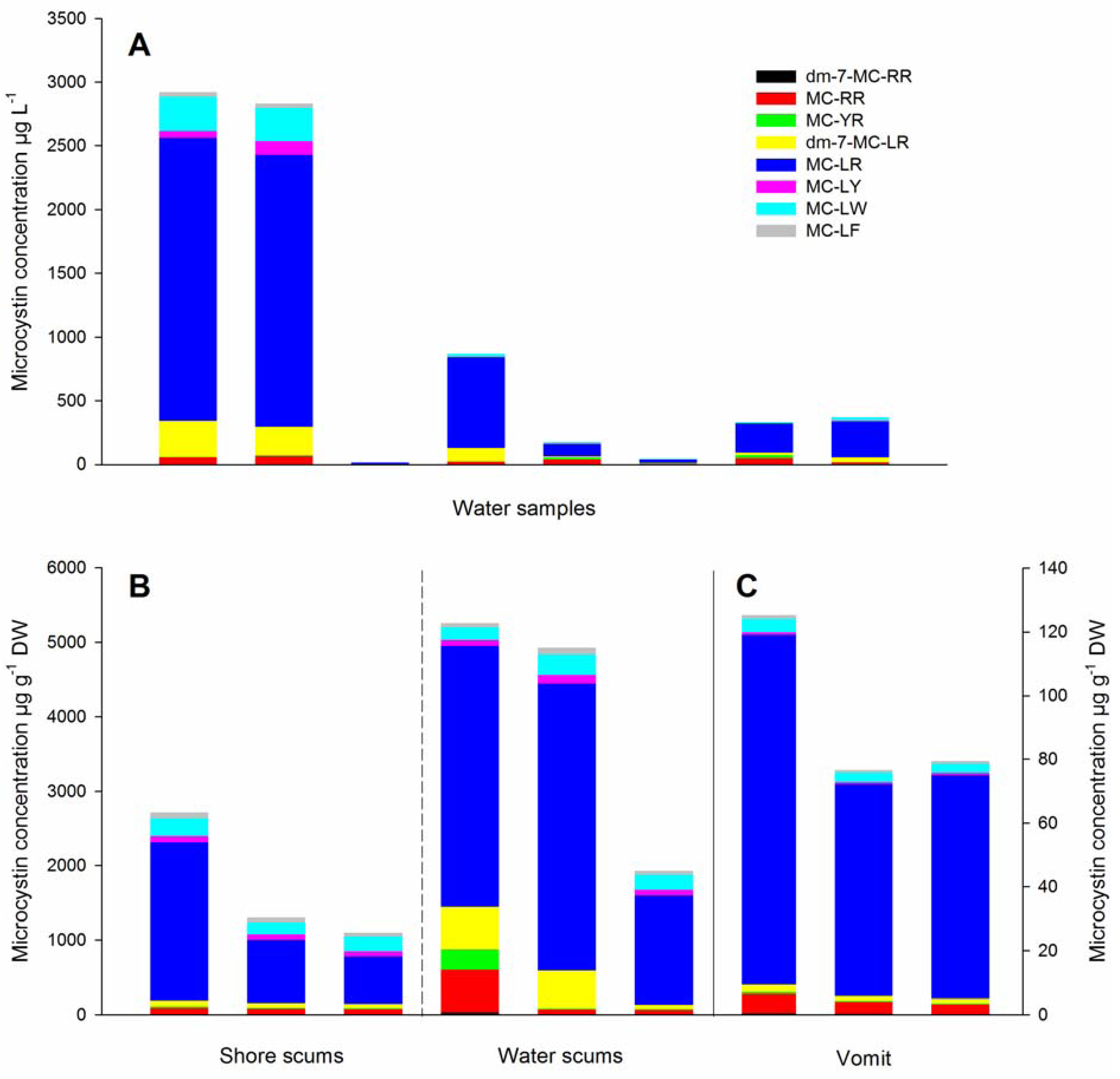

{kind=link}
{kind=link}
| Lake (country) | MC Concentration (µg g−1 dry-weight) | reference |
|---|---|---|
| Lake Baringo (Kenya) | 19800 | [31] |
| Bautzen Reservoir (Germany) | 14700 | [32] |
| Beaver Dam Lake 2 (USA) | 12800 | [33] |
| Lake Winnebago 1 (USA) | 10240 | [33] |
| Lalla Takerkoust (Morocco) | 8800 | [34] |
| Fish pond S2 Wuhan (China) | 7280 | [35] |
| River Guadiana (Portugal) | 7100 | [36] |
| Not specified (Germany) | 5595 | [37] |
| Lake Amstelmeer (Netherlands) | 5265 | This study |
| Lake Grand-Lieu (France) | 5060 | [38] |
| Lake Oubeira (Algeria) | 4590 | [39] |
| Laguna de Bay (Philippines) | 4049 | [40] |
3. Experimental Section
| Compound | Position 2 | Position 4 |
|---|---|---|
| dm-7-MC-RR 1 | Arginine | Arginine |
| MC-RR | Arginine | Arginine |
| NOD | n.a. | n.a. |
| MC-YR | Tyrosine | Arginine |
| dm-7-MC-LR 1 | Leucine | Arginine |
| MC-LR | Leucine | Arginine |
| MC-LY | Leucine | Tyrosine |
| MC-LW | Leucine | Tryptophan |
| MC-LF | Leucine | Phenylalanine |
| Compound | Retention time (min) | Precursor ion (m/z) | Fragmentor (V) | Quantifier ion (m/z) | CE 1 quantifier (V) | Qualifier ion (m/z) | CE 1 qualifier (V) | Ratio 2 (%) |
|---|---|---|---|---|---|---|---|---|
| dm-7-MC-RR | 6.93 | 512.8 | 135 | 135.1 | 26 | 70.1 | 85 | 1.2 |
| MC-RR | 7.62 | 519.8 | 151 | 135.1 | 30 | 70.1 | 75 | 2.7 |
| NOD | 8.03 | 825.5 | 220 | 135.1 | 70 | 70.1 | 95 | 44.2 |
| MC-YR | 8.16 | 523.3 | 102 | 911.5 | 5 | 135.1 | 6 | 103.6 |
| dm-7-MC-LR | 8.21 | 491.3 | 88 | 847.6 | 5 | 135.1 | 6 | 84.0 |
| MC-LR | 8.24 | 498.3 | 88 | 135.1 | 6 | 482.3 | 6 | 56.7 |
| MC-LY | 9.67 | 868.4 | 170 | 163.0 | 35 | 136.1 | 75 | 29.0 |
| MC-LW | 10.22 | 891.5 | 146 | 163.0 | 31 | 159.0 | 75 | 26.9 |
| MC-LF | 10.47 | 852.5 | 140 | 163.0 | 31 | 120.1 | 79 | 39.1 |
| Compound | LOD 1 (fmole inj−1) | LOQ 2 (fmole inj−1) | Calibration curve range (μg L−1) | Curve shape | Recovery (%) |
|---|---|---|---|---|---|
| dm-7-MC-RR | 132 | 132 | 14–338 | Quadratic | 100 |
| MC-RR | 92 | 92 | 10–949 | Quadratic | 96 |
| NOD | <17 | 17 | 1–368 | Linear | 105 |
| MC-YR | <10 | 10 | 1–518 | Linear | 75 |
| dm-7-MC-LR | <12 | 12 | 1–589 | Linear | 78 |
| MC-LR | <19 | 19 | 2–921 | Linear | 79 |
| MC-LY | 165 | 165 | 16–824 | Linear | 73 |
| MC-LW | 77 | 154 | 16–791 | Linear | 54 |
| MC-LF | 37 | 37 | 4–900 | Linear | 64 |
| Compound | Retention time | Peak area 1 | Ratio 2 |
|---|---|---|---|
| dm-7-MC-RR | 0.6 | 5.4 | 5.8 |
| MC-RR | 0.1 | 6.6 | 6.1 |
| NOD | 0.1 | 3.7 | 1.5 |
| MC-YR | 0.0 | 1.3 | 0.9 |
| dm-7-MC-LR | 0.1 | 1.5 | 1.4 |
| MC-LR | 0.0 | 1.4 | 2.8 |
| MC-LY | 0.1 | 1.6 | 7.2 |
| MC-LW | 0.1 | 3.4 | 11.2 |
| MC-LF | 0.0 | 4.9 | 7.4 |
4. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Smith, V.H.; Schindler, D.W. Eutrophication science: Where do we go from here? Trends Ecol. Evol. 2009, 24, 201–207. [Google Scholar] [CrossRef]
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef]
- Dittmann, E.; Wiegand, C. Cyanobacterial toxins—Occurrence, biosynthesis and impact on human affairs. Mol. Nutr. Food Res. 2006, 50, 7–17. [Google Scholar] [CrossRef]
- Stewart, I.; Seawright, A.A.; Shaw, G.R. Cyanobacterial poisoning in livestock, wild mammals and birds—An overview. Adv. Exp. Med. Biol. 2008, 619, 613–637. [Google Scholar]
- Codd, G.A.; Edwards, C.; Beattie, K.A.; Barr, W.M.; Gunn, G.J. Fatal attraction to cyanobacteria? Nature 1992, 359, 110–111. [Google Scholar]
- Edwards, C.; Beattie, K.A.; Scrimgeour, C.M.; Codd, G.A. Identification of anatoxin-a in benthic cyanobacteria (blue-green algae) and in associated dog poisonings at Loch Insh, Scotland. Toxicon 1992, 30, 1165–1175. [Google Scholar] [CrossRef]
- Gugger, M.; Lenoir, S.; Berger, C.; Ledreux, A.; Druart, J.-C.; Humbert, J.-F.; Guette, C.; Bernard, C. First report in a river in France of the benthic cyanobacterium Phormidium favosum producing anatoxin-a associated with dog neurotoxicosis. Toxicon 2005, 45, 919–928. [Google Scholar] [CrossRef]
- Wood, S.A.; Selwood, A.I.; Rueckert, A.; Holland, P.T.; Milne, J.R.; Smith, K.F.; Smits, B.; Watts, L.F.; Cary, C.S. First report of homoanatoxin-a and associated dog neurotoxicosis in New Zealand. Toxicon 2007, 50, 292–301. [Google Scholar] [CrossRef]
- Puschner, B.; Hoff, B.; Tor, E.R. Diagnosis of anatoxin-a poisoning in dogs from North America. J. Vet. Diagn. Investig. 2008, 20, 89–92. [Google Scholar] [CrossRef]
- Faassen, E.J.; Harkema, L.; Begeman, L.; Lurling, M. First report of (homo)anatoxin-a and dog neurotoxicosis after ingestion of benthic cyanobacteria in The Netherlands. Toxicon 2012, 60, 378–384. [Google Scholar] [CrossRef]
- Francis, G. Poisonous Australian lake. Nature 1878, 18, 11–12. [Google Scholar] [CrossRef]
- Nehring, S. Mortality of dogs associated with a mass development of Nodularia spumigena (Cyanophyceae) in a brackish lake at the German North Sea coast. J. Plankton Res. 1993, 15, 867–872. [Google Scholar] [CrossRef]
- Simola, O.; Wiberg, M.; Jokela, J.; Wahlsten, M.; Sivonen, K.; Syrjä, P. Pathologic findings and toxin identification in cyanobacterial (Nodularia spumigena) intoxication in a dog. Vet. Pathol. 2012, 49, 755–759. [Google Scholar] [CrossRef]
- O’Donoghue, J.G.; Wilton, G.S. Algal poisoning in Alberta. Can. J. Comp. Med. 1951, 15, 193–198. [Google Scholar]
- Senior, V.E. Algal poisoning in Saskatchewan. Can. J. Comp. Med. 1960, 24, 26–31. [Google Scholar]
- Sivonen, K.; Jones, G. Cyanobacterial Toxins. In Toxic Cyanobacteria in Water. A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; E&FN Spon: London, UK, 1999; pp. 41–111. [Google Scholar]
- De Figueiredo, D.R.; Azeiteiro, U.M.; Esteves, S.M.; Gonçalves, F.J.M.; Pereira, M.J. Microcystin-producing blooms-a serious global public health issue. Ecotoxicol. Environ. Saf. 2004, 59, 151–163. [Google Scholar] [CrossRef]
- Doekel, S.; Marahiel, M.A. Biosynthesis of natural products on molecular peptide synthetases. Metab. Eng. 2001, 3, 64–77. [Google Scholar] [CrossRef]
- Steis, J.E.; White, J.C. Respiratory alkalosis and primary hypocapnia in Labrador Retrievers participating in field trials in high-ambient-temperature conditions. Am. J. Vet. Res. 2008, 69, 1262–1267. [Google Scholar] [CrossRef]
- Pobel, D.; Godon, J.J.; Humbert, J.F.; Robin, J. High-frequency monitoring of the genetic diversity and the potential toxicity of a Microcystis aeruginosa bloom in a French shallow lake. FEMS Microbiol. Ecol. 2012, 79, 132–141. [Google Scholar]
- Hotto, A.M.; Satchwell, M.F.; Berry, D.L.; Gobler, C.J.; Boyer, G.L. Spatial and temporal diversity of microcystins and microcystin-producing genotypes in Oneida Lake, NY. Harmful Algae 2008, 7, 671–681. [Google Scholar] [CrossRef]
- Van de Waal, D.B.; Verspagen, J.M.H.; Lürling, M.; van Donk, E.; Visser, P.M.; Huisman, J. The ecological stoichiometry of toxins produced by harmful cyanobacteria: An experimental test of the carbon-nutrient balance hypothesis. Ecol. Lett. 2009, 12, 1326–1335. [Google Scholar] [CrossRef]
- Feurstein, D.; Holst, K.; Fisher, A.; Dietrich, D.R. Oatp-associated uptake and toxicity of microcystins in primary murine whole brain cells. Toxicol. Appl. Pharmacol. 2009, 234, 247–255. [Google Scholar] [CrossRef]
- Feurstein, D.; Stemmer, K.; Kleinteich, J.; Speicher, D.; Dietrich, D.R. Microcystin congener- and concentration-dependent induction of murine neuron apoptosis and neurite degeneration. Toxicol. Sci. 2011, 124, 424–431. [Google Scholar] [CrossRef]
- Fisher, A.; Hoeger, S.J.; Stemmer, K.; Feurstein, D.J.; Knobeloch, D.; Nussler, A.; Dietrich, D.R. The role of organic anion transporting polypeptides (OATPs/SLCOs) in the toxicity of different microcystin congeners in vitro: A comparison of primary human hepatocytes and OATP-transferred HEK293 cells. Toxicol. Appl. Pharmacol. 2010, 245, 9–20. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Azevedo, S.M.F.O.; An, J.S.; Molica, R.J.R.; Jochimsen, E.L.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesam, G.K. Human fatalities from cyanobacteria: Chemical and biological evidence for cyanotoxins. Environ. Health Perspect. 2001, 109, 663–668. [Google Scholar] [CrossRef]
- Otten, T.G.; Xu, H.; Qin, B.; Zhu, G.; Paerl, H.W. Spatiotemporal patterns and ecophysiology of toxigenic Microcystis blooms in Lake Taihu, China: Implications for water quality management. Environ. Sci. Technol. 2012, 46, 3480–3488. [Google Scholar] [CrossRef]
- Pobel, D.; Robin, J.; Humbert, J.-F. Influence of sampling strategies on the monitoring of cyanobacteria in shallow lakes: Lessons from a case study in France. Water Res. 2011, 45, 1005–1014. [Google Scholar] [CrossRef]
- Sabart, M.; Pobel, D.; Briand, E.; Combourieu, B.; Salençon, M.J.; Humbert, J.F.; Latour, D. Spatiotemporal variations in microcystin concentrations and in the proportions of microcystin-producing cells in several Microcystis aeruginosa populations. Appl. Environ. Microbiol. 2010, 76, 4750–4759. [Google Scholar] [CrossRef] [Green Version]
- Lürling, M.; Faassen, E.J. Controlling toxic cyanobacteria: Effects of dredging and phosphorus-binding clay on cyanobacteria and microcystins. Water Res. 2012, 46, 1447–1459. [Google Scholar] [CrossRef]
- Ballot, A.; Pflugmacher, S.; Wiegand, C.; Kotut, K.; Krienitz, L. Cyanobacterial toxins in Lake Baringo, Kenya. Limnologica 2003, 33, 2–9. [Google Scholar] [CrossRef]
- Jungmann, D.; Ludwichowski, K.-U.; Faltin, V.; Benndorf, J. A field study to investigate environmental factors that could effect microcystin synthesis of a Microcystis population in the Bautzen Reservoir. Int. Rev. Gesamten Hydrobiol. Hydrogr. 1996, 81, 493–501. [Google Scholar] [CrossRef]
- McDermott, C.M.; Feola, R.; Plude, J. Detection of cyanobacterial toxins (microcystins) in waters of northeastern Wisconsin by a new immunoassay technique. Toxicon 1995, 33, 1433–1442. [Google Scholar] [CrossRef]
- Oudra, B.; Loudiki, M.; Sbiyyaa, B.; Martins, R.; Vasconcelos, V.; Namikoshi, N. Isolation, characterization and quantification of microcystins (heptapeptides hepatotoxins) in Microcystis aeruginosa dominated bloom of Lalla Takerkoust lake-reservoir (Morocco). Toxicon 2001, 39, 1375–1381. [Google Scholar] [CrossRef]
- Zhang, Q.-X.; Carmichael, W.W.; Yu, M.-J.; Li, S.-H. Cyclic peptide hepatotoxins from freshwater cyanobacterial (blue-green algae) waterblooms collected in central China. Environ. Toxicol. Chem. 1991, 10, 313–321. [Google Scholar] [CrossRef]
- Vasconcelos, V.M.; Sivonen, O.K.; Evans, W.R.; Carmichael, W.W.; Namikoshi, M. Hepatotoxic microcystin diversity in cyanobacterial blooms collected in Portuguese freshwaters. Water Res. 1996, 30, 2377–2384. [Google Scholar] [CrossRef]
- Fastner, J.; Neumann, U.; Wirsing, B.; Weckesser, J.; Wiedner, C.; Nixdorf, B.; Chorus, I. Microcystins (hepatotoxic heptapeptides) in German fresh water bodies. Environ. Toxicol. 1999, 14, 13–22. [Google Scholar] [CrossRef]
- Vezie, C.; Brient, L.; Sivonen, K.; Bertru, G.; Lefeuvre, J.-C.; Salkinoja-Salonen, M. Variation of microcystin content of cyanobacterial blooms and isolated strains in Lake Grand-Lieu (France). Microb. Ecol. 1998, 35, 126–135. [Google Scholar]
- Nasri, A.-B.; Bouaïcha, N.; Fastner, J. First report of a microcystin-containing bloom of the cyanobacteria Microcystis spp. in Lake Oubeira, Eastern Algeria. Arch. Environ. Contam. Toxicol. 2004, 46, 197–202. [Google Scholar]
- Cuvin-Aralar, M.; Fastner, J.; Focken, U.; Becker, K.; Aralar, E.V. Microcystins in natural blooms and laboratory cultured Microcystis aeruginosa from Laguna de Bay, Philippines. Syst. Appl. Microbiol. 2002, 25, 179–182. [Google Scholar] [CrossRef]
- Vreman, B.J. Zwemwaterprofiel Amstelmeer (Lutjestrand) 2007 t/m 2010. 2011. (in Dutch). Available online: http://www.hhnk.nl/aspx/download.aspx?File=/contents/pages/214671/250211_ definitief_zwemwaterprofiel_lutjestrand_actualisatie_2011.pdf (accessed on 23 December 2012).
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lürling, M.; Faassen, E.J. Dog Poisonings Associated with a Microcystis aeruginosa Bloom in the Netherlands. Toxins 2013, 5, 556-567. https://doi.org/10.3390/toxins5030556
Lürling M, Faassen EJ. Dog Poisonings Associated with a Microcystis aeruginosa Bloom in the Netherlands. Toxins. 2013; 5(3):556-567. https://doi.org/10.3390/toxins5030556
Chicago/Turabian StyleLürling, Miquel, and Elisabeth J. Faassen. 2013. "Dog Poisonings Associated with a Microcystis aeruginosa Bloom in the Netherlands" Toxins 5, no. 3: 556-567. https://doi.org/10.3390/toxins5030556