Interactions of PLA2-s from Vipera lebetina, Vipera berus berus and Naja naja oxiana Venom with Platelets, Bacterial and Cancer Cells

 , and
, and
Abstract
:Abbreviations
| svPLA2 | snake venom PLA2 |
| sPLA2 | secretory PLA2 |
| VLPLA2 | Vipera lebetina phospholipase A2 |
| VBBPLA2 | Vipera berus berus phospholipase A2 |
| NNOPLA2 | Naja naja oxiana phospholipase A2 |
| p-BPB | p-bromophenacylbromide |
| MALDI-TOF MS | matrix assisted laser desorption ionization time-of-flight mass spectrometry |
| lysoPC | lysophosphatidylcholine |
| PRP | platelet-rich plasma |
1. Introduction
2. Results
2.1. Purification and Characterization of sPLA2s

2.2. Inhibition of Human Platelet Aggregation in Vitro

2.3. Inhibitory Effect of Snake Venoms and Their sPLA2s on Bacteria
2.3.1. Acute Toxicity to Vibrio fischeri

{kind=link}
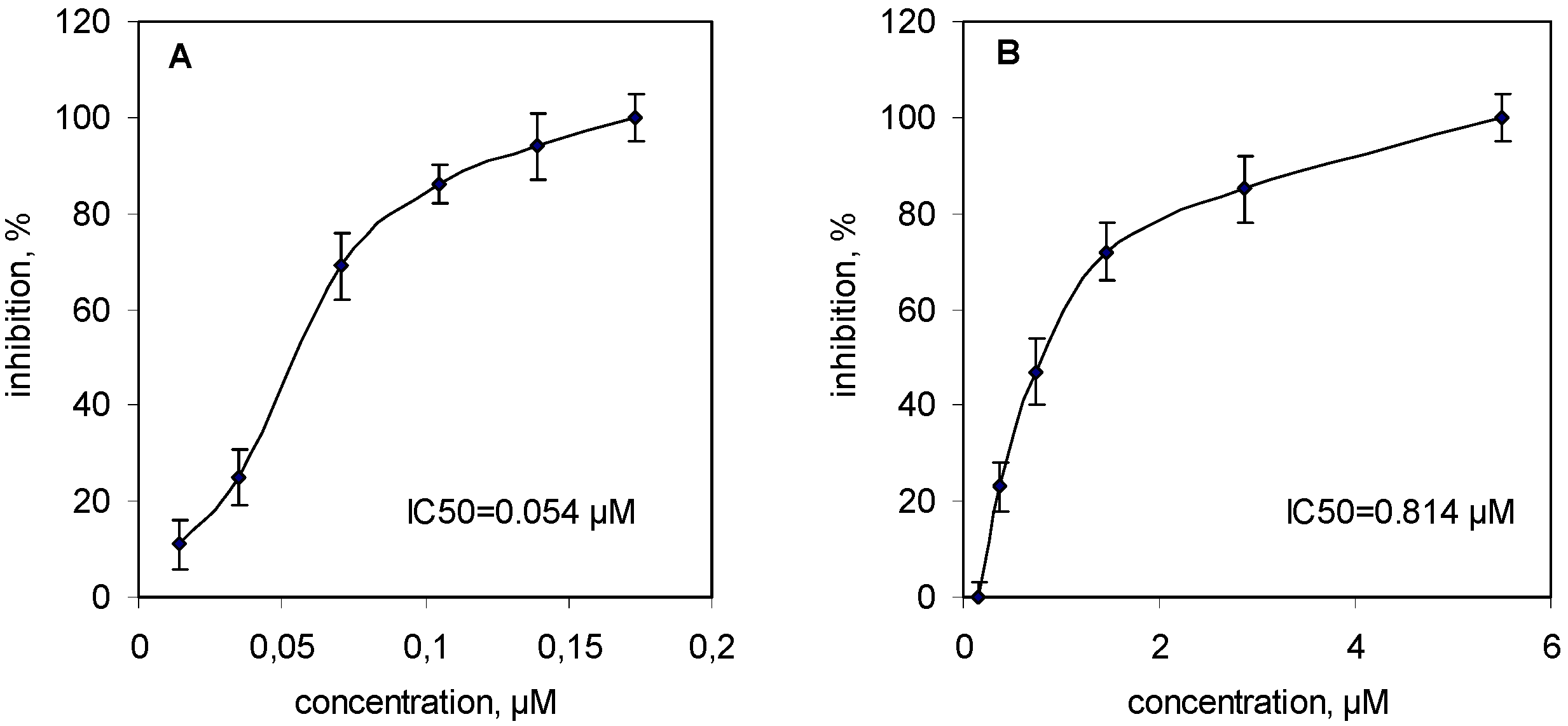{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tested item | Acute toxicity (15-min EC50, μg/mL) | |||
|---|---|---|---|---|
| 3,5-DCP * | V. b. berus | V. lebetina | N. n. oxiana | |
| Venom | 3–4 | 370 | 944 | >1315 |
| PLA2 | 3–4 | >909 (>65.76 μM) | 58 (4.24 μM) | >606 (>45.81 μM) |
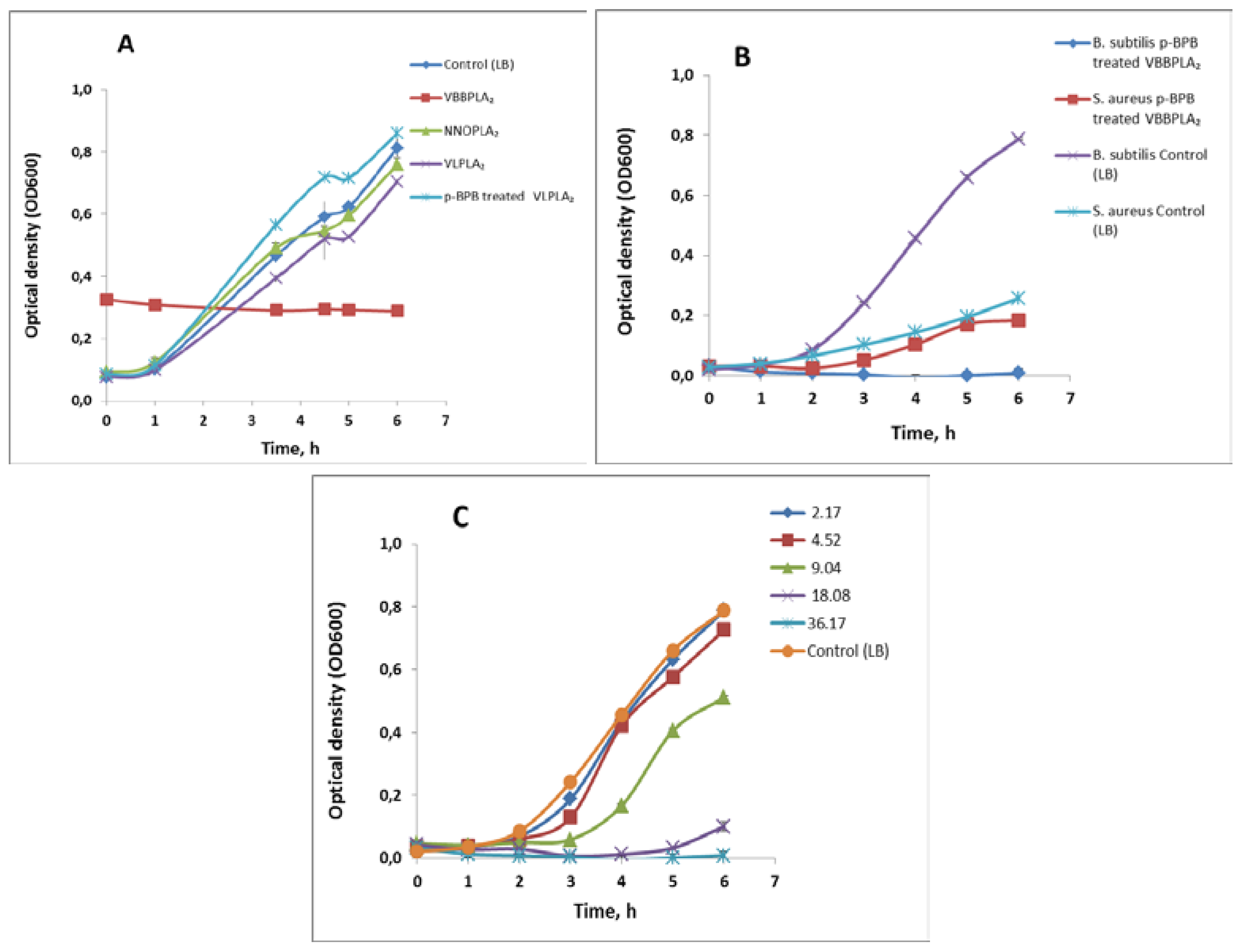
2.3.2. Inhibitory Effect of the Snake Venom PLA2s on Bacterial Growth
| Inhibition of the bacterial growth, % (t = 6 h) | ||||||
|---|---|---|---|---|---|---|
| Bacteria(Gram staining) | svPLA2 | |||||
| V. b. berus | V. b. berus * | V. lebetina | V. lebetina * | N. n. oxiana | ||
| Escherichia coli | Gram (−) | No effect | not tested | No effect | No effect | No effect |
| Bacillus subtilis | Gram (+) | 100% ** | 99% ** | 13% | No effect | Slight effect (6.5%) |
| Staphylococcus aureus | Gram (+) | No effect | 29% | No effect | No effect | 42% |
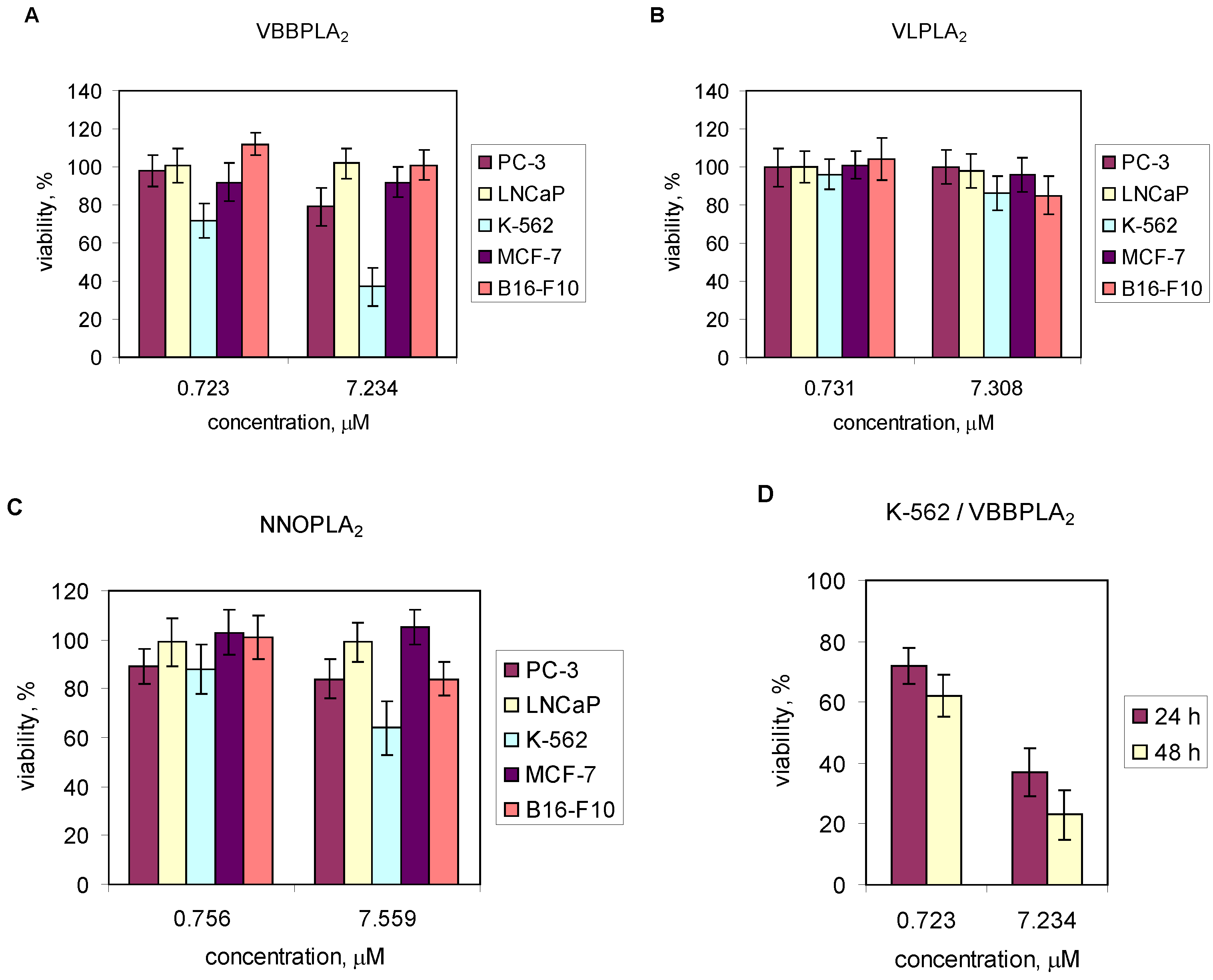
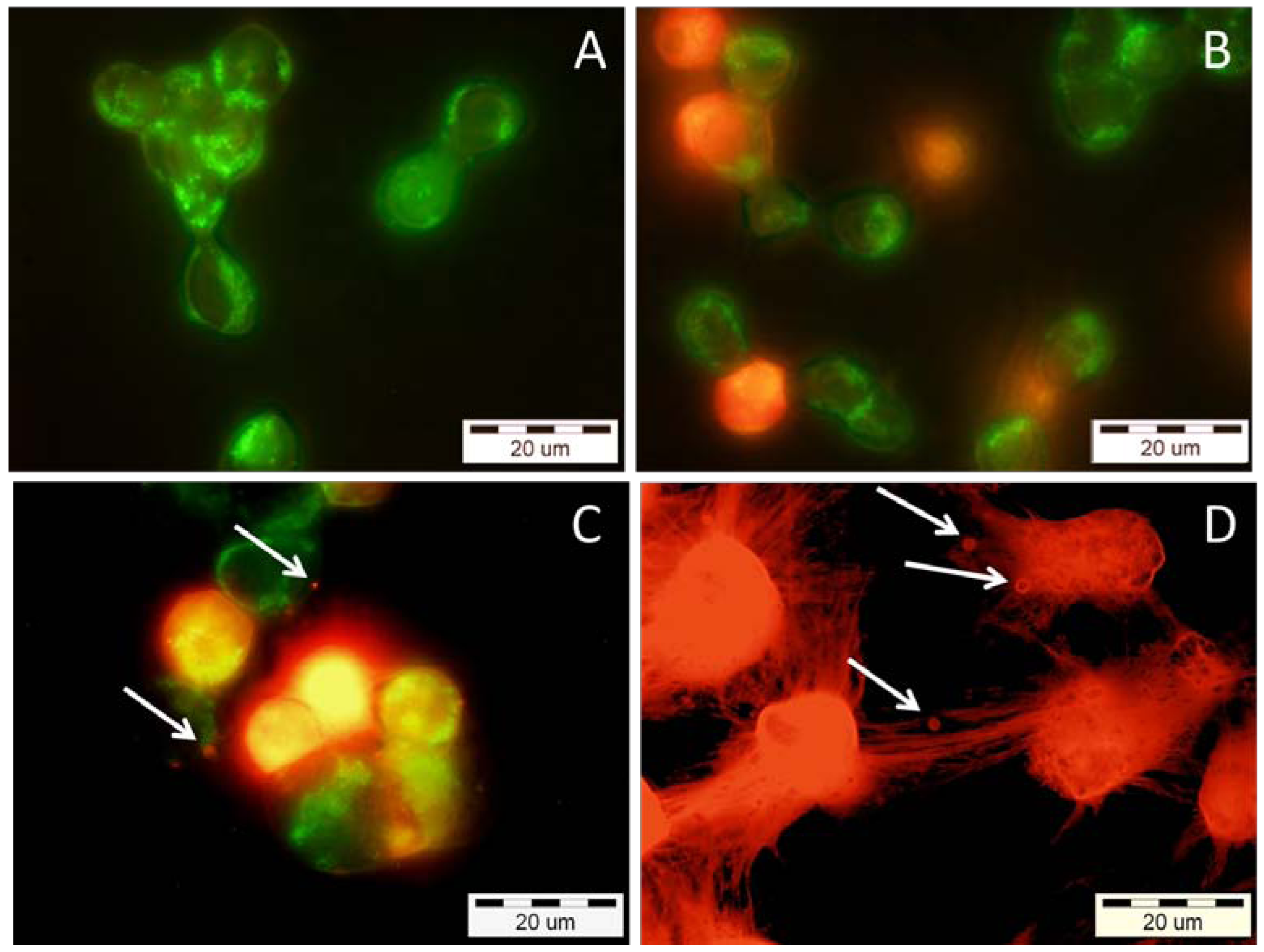
2.4. Effects of Snake Venom PLA2s on Cancer Cells Viability
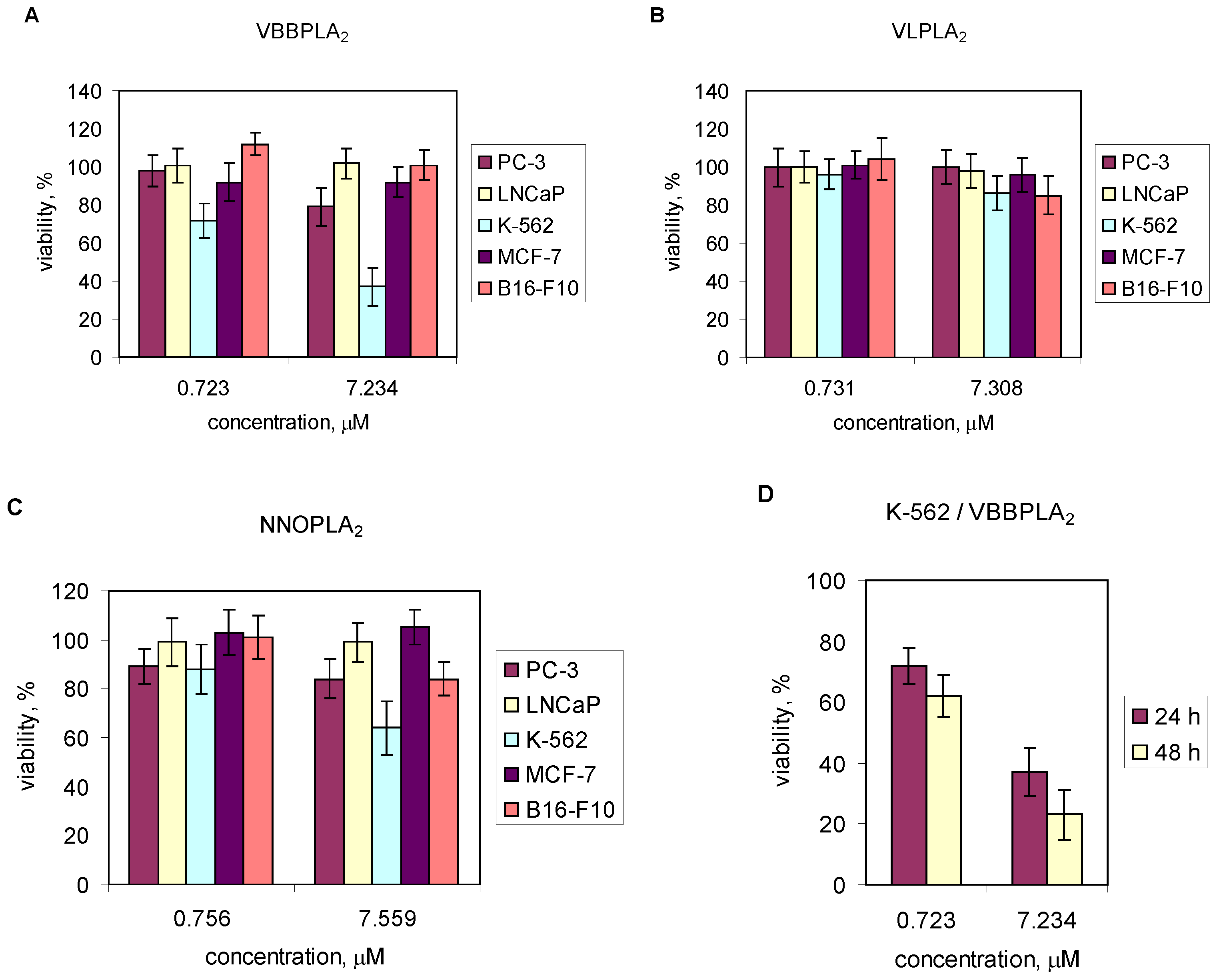
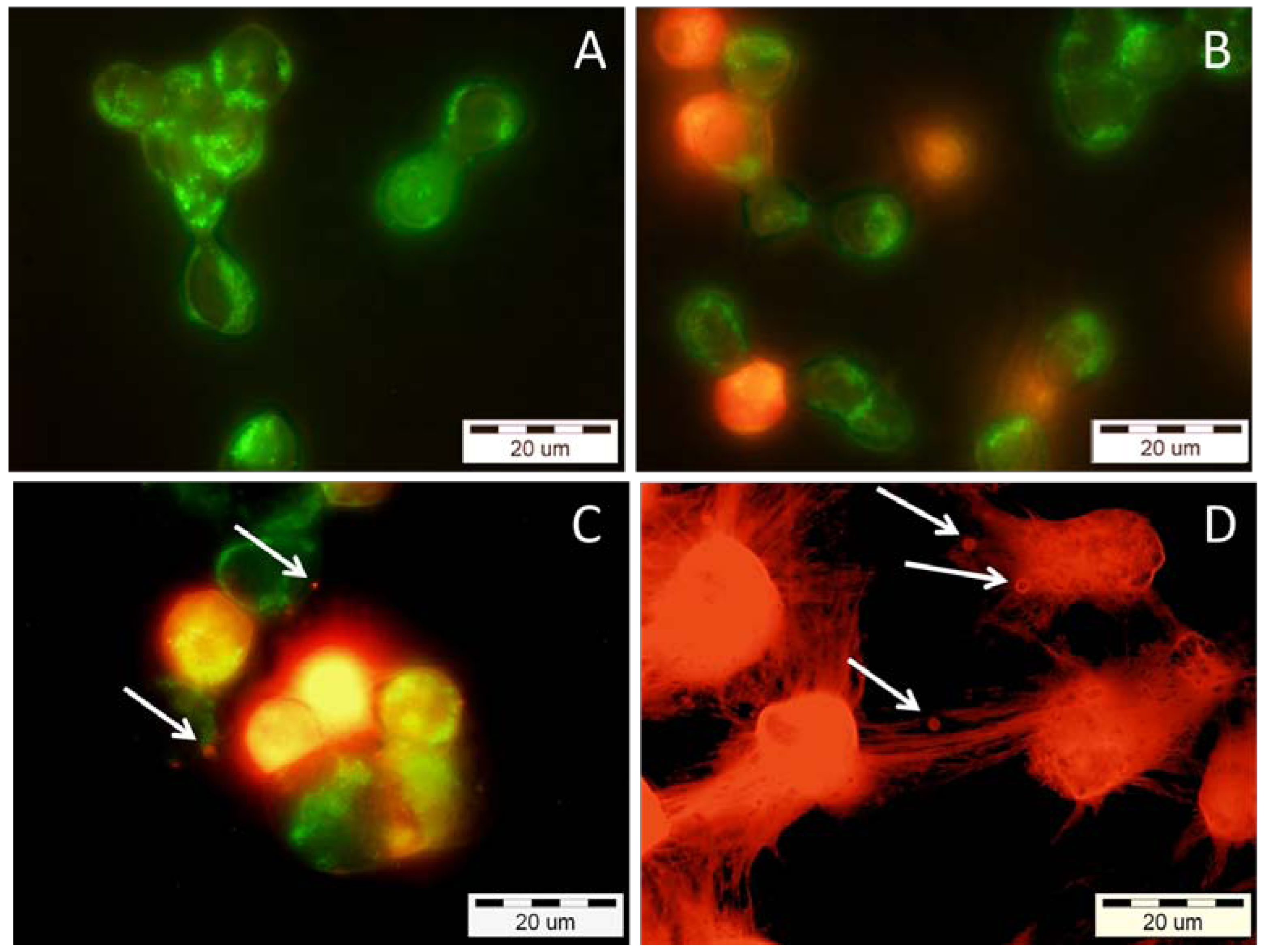
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Purification of Enzymes
4.3. PLA2 Assay
4.4. PLA2 Activity Inhibition with p-bromophenacylbromide (p-BPB)
4.5. Protein Quantification
4.6. Molecular Mass Detection and Isoelectric Focusing of Proteins
4.7. In-Gel Tryptic Digestion and Mass Fingerprinting of Proteins
4.8. Preparation of Human Platelet Suspension and Collagen-Induced Platelet Aggregation Assay
4.9. Antibacterial Activity
4.9.1. Bacterial Strains
4.9.2. Analysis of Antibacterial Activity of PLA2s
4.9.2.1. Bioluminescence Inhibition Assay Using Vibrio fischeri
4.9.2.2. Bacterial Growth Inhibition Assays
4.10. Human Cell Lines and Toxicity Testing of sPLA2s
4.10.1. Analysis of the Viability of the Cells
4.10.1.1. MTT Assay
4.10.1.2. WST-1 Assay
4.10.2. Apoptosis Detection Using Annexin V-FITC and Propidium Iodide (PI)
5. Conclusions
Acknowledgements
Conflict of interest
References
- Murakami, M.; Taketomi, Y.; Miki, Y.; Sato, H.; Hirabayashi, T.; Yamamoto, K. Recent progress in phospholipase A2 research: From cells to animals to humans. Progr. Lipid Res. 2011, 50, 152–192. [Google Scholar] [CrossRef]
- Schaloske, R.H.; Dennis, E.A. The phospholipase A2 superfamily and its group numbering system. Biochim. Biophys. Acta 2006, 1761, 1246–1259. [Google Scholar]
- Burke, J.E.; Dennis, E.A. Phospholipase A2 structure/function, mechanism, and signaling. J. Lipid Res. 2009, 59, S237–S242. [Google Scholar]
- Nevalainen, T.J.; Cardoso, J.C.; Riikonen, P.T. Conserved domains and evolution of secreted phospholipases A(2). FEBS J. 2012, 279, 636–649. [Google Scholar] [CrossRef]
- Lomonte, B.; Angulo, Y.; Sasa, M.; Gutiérrez, J.M. The phospholipase A2 homologues of snake venoms: Biological activities and their possible adaptive roles. Protein Pept. Lett. 2009, 16, 860–876. [Google Scholar] [CrossRef]
- Doley, R.; Zhou, X.; Kini, R.M. Snake Venom Phospholipase A2 Enzymes. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 173–205. [Google Scholar]
- Kini, R.M. Excitement ahead: Structure, function and mechanism of snake venom phospholipase A2 enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef]
- Kini, R.M. Structure-function relationships and mechanism of anticoagulant phospholipase A2 enzymes from snake venoms. Toxicon 2005, 45, 1147–1161. [Google Scholar] [CrossRef]
- Lambeau, G.; Lazdunski, M. Receptors for a growing family of secreted phospholipases A2. Trends Pharmacol. Sci. 1999, 20, 162–170. [Google Scholar] [CrossRef]
- Soares, A.M.; Giglio, J.R. Chemical modifications of phospholipases A2 from snake venoms: Effects on catalytic and pharmacological properties. Toxicon 2003, 42, 855–868. [Google Scholar] [CrossRef]
- Montecucco, C.; Gutiérrez, J.M.; Lomonte, B. Cellular pathology induced by snake venom phospholipase A2 myotoxins and neurotoxins: Common aspects of their mechanism of action. Cell. Mol. Life Sci. 2008, 65, 2897–2912. [Google Scholar] [CrossRef]
- Rodrigues, R.S.; Izidoro, L.F.; de Oliveira, R.J., Jr.; Sampaio, S.V.; Soares, A.M.; Rodrigues, V.M. Snake venom phospholipases A2: A new class of antitumor agents. Protein Pept. Lett. 2009, 16, 894–898. [Google Scholar] [CrossRef]
- Ouyang, C.; Huang, T.F. Potent platelet aggregation inhibitor from Trimeresurus gramineus snake venom. Biochim. Biophys. Acta 1983, 757, 332–341. [Google Scholar] [CrossRef]
- Chen, R.H.; Chen, Y.C. Isolation of an acidic phospholipase A2 from the venom of Agkistrodon acutus (five pace snake) and its effect on platelet aggregation. Toxicon 1989, 27, 675–682. [Google Scholar] [CrossRef]
- Kini, R.M.; Evans, H.J. Effects of Phospholipase A2 Rnzymes on Platelet Sggregation. In Venom phospholipase A2 Rnzymes: Dtructure, Gunction and Mechanism; Kini, R.M., Ed.; John Wiley & Sons: Chichester, UK, 1997; pp. 369–387. [Google Scholar]
- Sim, K.L. Purification and preliminary characterisation of praelongin phospholipases, antiplatelet agents from the snake venom of Acanthophis praelongus. Biochim. Biophys. Acta 1998, 1379, 198–206. [Google Scholar]
- Fuly, A.L.; Soares, A.M.; Marcussi, S.; Giglio, J.R.; Guimarães, J.A. Signal transduction pathways involved in the platelet aggregation induced by a D-49 phospholipase A2 isolated from Bothrops jararacussu snake venom. Biochimie 2004, 86, 731–739. [Google Scholar] [CrossRef]
- Vija, H.; Samel, M.; Siigur, E.; Aaspõllu, A.; Trummal, K.; Tõnismägi, K.; Subbi, J.; Siigur, J. Purification, characterization, and cDNA cloning of acidic platelet aggregation inhibiting phospholipases A2 from the snake venom of Vipera lebetina (Levantine viper). Toxicon 2009, 54, 429–439. [Google Scholar] [CrossRef]
- Tsai, I.H.; Wang, Y.M.; Cheng, A.C.; Starkov, V.; Osipov, A.; Nikitin, I.; Makarova, Y.; Ziganshin, R.; Utkin, Y. cDNA cloning, structural, and functional analyses of venom phospholipases A2 and a Kunitz-type protease inhibitor from steppe viper Vipera ursinii renardi. Toxicon 2011, 57, 332–341. [Google Scholar] [CrossRef]
- Prasad, B.N.; Kemparaju, K.; Bhatt, K.G.; Gowda, T.V. A platelet aggregation inhibitor phospholipase A2 from Russell’s viper (Vipera russelli) venom: Isolation and characterization. Toxicon 1996, 34, 1173–1185. [Google Scholar] [CrossRef]
- Fuly, A.L.; Machado, O.L.T.; Alves, E.W.; Carlini, C.R. Mechanism of inhibitory action on platelet activation of a phospholipase A2 isolated from Lachesis muta (Bushmaster) snake venom. Thromb. Haemost. 1997, 78, 1372–1380. [Google Scholar]
- Chow, G.; Subburaju, S.; Kini, R.M. Purification, characterization, and amino acid sequence determination of acanthins, potent inhibitors of platelet aggregation from Acanthophis antarcticus (common death adder) venom. Arch. Biochem. Biophys. 1998, 354, 232–238. [Google Scholar] [CrossRef]
- Kemparaiu, K.; Krishnakanth, T.P.; Veerabasappa Gowda, T. Purification and characterization of a platelet aggregation inhibitor acidic phospholipase A2 from Indian saw-scaled viper (Echis carinatus) venom. Toxicon 1999, 37, 1659–1671. [Google Scholar] [CrossRef]
- Serrano, S.M.T.; Reichl, A.P.; Mentele, R.; Auerswald, E.A.; Santoro, M.L.; Sampaio, C.A.M.; Camargo, A.C.M.; Assakura, M.T. A novel phospholipase A2, BJ-PLA2, from the venom of the snake Bothrops jararaca: Purification, primary structure analysis, and its characterization as a platelet-aggregation-inhibiting factor. Arch. Biochem. Biophys. 1999, 367, 26–32. [Google Scholar] [CrossRef]
- Roberto, P.G.; Kashima, S.; Marcussi, S.; Pereira, J.O.; Astolfi-Filho, S.; Nomizo, A.; Giglio, J.R.; Fontes, M.R.; Soares, A.M.; Franca, S.C. Cloning and identification of a complete cDNA coding for a bactericidal and antitumoral acidic phospholipase A2 from Bothrops jararacussu venom. Protein J. 2004, 23, 273–285. [Google Scholar] [CrossRef]
- Satish, S.; Tejaswini, J.; Krishnakantha, T.P.; Gowda, T.V. Purification of a Class B1 platelet aggregation inhibitor phospholipase A2 from Indian cobra (Naja Naja) venom. Biochimie 2004, 86, 203–210. [Google Scholar] [CrossRef]
- Páramo, L.; Lomonte, B.; Pizarro-Cerdá, J.; Bengoechea, J.A.; Gorvel, J.P.; Moreno, E. Bactericidal activity of Lys49 and Asp49 myotoxic phospholipases A2 from Bothrops asper snake venom—Synthetic Lys49 myotoxin II-(115–129)-peptide identifies its bactericidal region. Eur. J. Biochem. 1998, 253, 452–461. [Google Scholar]
- Koduri, R.S.; Gronroos, J.O.; Laine, V.J.; le Calvez, C.; Lambeau, G.; Nevalainen, T.J.; Gelb, M.H. Bactericidal properties of human and murine groups I, II, V, X, and XII secreted phospholipases A2. J. Biol. Chem. 2002, 277, 5849–5857. [Google Scholar]
- Oliveira, D.G.; Toyama, M.H.; Novello, J.C; Beriam, L.O.; Marangoni, S. Structural and functional characterization of basic PLA2 isolated from Crotalus durissus terrificus venom. J. Protein Chem. 2002, 21, 161–168. [Google Scholar] [CrossRef]
- Núñez, V.; Arce, V.; Gutiérrez, J.M.; Lomonte, B. Structural and functional characterization of myotoxin I, a Lys49 phospholipase A2 homologue from the venom of the snake Bothrops atrox. Toxicon 2004, 44, 91–101. [Google Scholar] [CrossRef]
- Rodrigues, V.M.; Marcussi, S.; Cambraia, R.S.; de Araújo, A.L.; Malta-Neto, N.R.; Hamaguchi, A.; Ferro, E.A.; Homsi-Brandeburgo, M.I.; Giglio, J.R.; Soares, A.M. Bactericidal and neurotoxic activities of two myotoxic phospholipases A2 from Bothrops neuwiedi pauloensis snake venom. Toxicon 2004, 44, 305–314. [Google Scholar] [CrossRef]
- Santamaría, C.; Larios, S.; Angulo, Y.; Pizarro-Cerda, J.; Gorvel, J.P.; Moreno, E.; Lomonte, B. Antimicrobial activity of myotoxic phospholipases A2 from crotalid snake venoms and synthetic peptide variants derived from their C-terminal region. Toxicon 2005, 45, 807–815. [Google Scholar] [CrossRef]
- Xu, C.; Ma, D.; Yu, H.; Li, Z.; Liang, J.; Lin, G.; Zhang, Y.; Lai, R. A bactericidal homodimeric phospholipases A2 from Bungarus fasciatus venom. Peptides 2007, 28, 969–973. [Google Scholar] [CrossRef]
- Perumal Samy, R.; Gopalakrishnakone, P.; Ho, B.; Chow, V.T. Purification, characterization and bactericidal activities of basic phospholipase A2 from the venom of Agkistrodon halys (Chinese pallas). Biochimie 2008, 90, 1372–1388. [Google Scholar] [CrossRef]
- Perumal Samy, R.; Gopalakrishnakone, P.; Bow, H.; Puspharaj, P.N.; Chow, V.T.K. Identification and characterization of a phospholipase A2 from the venom of the Saw-scaled viper: Novel bactericidal and membrane damaging activities. Biochimie 2010, 92, 1854–1866. [Google Scholar] [CrossRef]
- Costa Torres, A.F.; Dantas, R.T.; Toyama, M.H.; Diz Filho, E.; Zara, F.J.; Rodrigues de Queiroz, M.G.; Pinto Nogueira, N.A.; Rosa de Oliveira, M.; de Oliveira Toyama, D.; Monteiro, H.S.; Martins, A.M. Antibacterial and antiparasitic effects of Bothrops marajoensis venom and its fractions: Phospholipase A2 and L-amino acid oxidase. Toxicon 2010, 55, 795–804. [Google Scholar] [CrossRef]
- Samy, R.P.; Stiles, B.G.; Gopalakrishnakone, P.; Chow, V.T. Antimicrobial proteins from snake venoms: Direct bacterial damage and activation of innate immunity against Staphylococcus aureus skin infection. Curr. Med. Chem. 2011, 18, 5104–5113. [Google Scholar] [CrossRef]
- Toyama, M.H.; Toyama, D.O.; Joazeiro, P.P.; Carneiro, E.M.; Beriam, L.O.; Marangoni, L.S.; Boschero, A.C. Biological and structural characterization of a new PLA2 from the Crotalus durissus collilineatus venom. Protein J. 2005, 24, 103–112. [Google Scholar] [CrossRef]
- Nevalainen, T.J.; Graham, G.G.; Scott, K.F. Antibacterial actions of secreted phospholipases A2. Biochim. Biophys. Acta 2008, 1781, 1–9. [Google Scholar]
- Roberto, P.G.; Kashima, S.; Soares, A.M.; Chioato, L.; Faca, V.M.; Fuly, A.L.; Astolfi-Filho, S.; Pereira, J.O.; Franca, S.C. Cloning and expression of an acidic platelet aggregation inhibitor phospholipase A2 cDNA from Bothrops jararacussu venom gland. Protein Expr. Purif. 2004, 37, 102–108. [Google Scholar] [CrossRef]
- Maity, G.; Mandal, S.; Chatterjee, A.; Bhattacharyya, D. Purification and characterization of a low molecular weight multifunctional cytotoxic phospholipase A2 from Russell's viper venom. J. Chromatogr. B 2007, 845, 232–243. [Google Scholar] [CrossRef]
- Zouari-Kessentini, R.; Luis, J.; Karray, A.; Kallech-Ziri, O.; Srairi-Abid, N.; Bazaa, A.; Loret, E.; Bezzine, S.; el Ayeb, M.; Marrakchi, N. Two purified and characterized phospholipases A2 from Cerastes cerastes venom, that inhibit cancerous cell adhesion and migration. Toxicon 2009, 53, 444–453. [Google Scholar] [CrossRef]
- Chen, K.C.; Liu, W.H.; Chang, L.S. Taiwan cobra phospholipase A2-elicited JNK activation is responsible for autocrine fas-mediated cell death and modulating Bcl-2 and Bax protein expression in human leukemia K562 cells. J. Cell Biochem. 2010, 109, 245–254. [Google Scholar]
- Murakami, T.; Kamikado, N.; Fujimoto, R.; Hamaguchi, K.; Nakamura, H.; Chijiwa, T.; Ohno, M.; Oda-Ueda, N. A [Lys49] phospholipase A2 from Protobothrops flavoviridis venom induces caspase-independent apoptotic cell death accompanied by rapid plasma-membrane rupture in human leukemia cells. Biosci. Biotechnol. Biochem. 2011, 75, 864–870. [Google Scholar] [CrossRef]
- Khunsap, S.; Pakmanee, N.; Khow, O.; Chanhome, L.; Sitprija, V.; Suntravat, M.; Lucena, S.E.; Perez, J.C.; Sánchez, E.E.J. Purification of a phospholipase A(2) from Daboia russelii siamensis venom with anticancer effects. Venom Res. 2011, 2, 42–51. [Google Scholar]
- Bazaa, A.; Luis, J.; Srairi-Abid, N.; Kallech-Ziri, O.; Kessentini-Zouari, R.; Defilles, C.; Lissitzky, J.C.; el Ayeb, M.; Marrakchi, N. MVL-PLA2, a phospholipase A2 from Macrovipera lebetina transmediterranea venom, inhibits tumor cells adhesion and migratio. Matrix Biol. 2009, 28, 188–193. [Google Scholar] [CrossRef]
- Bazaa, A.; Pasquier, E.; Defilles, C.; Limam, I.; Kessentini-Zouari, R.; Kallech-Ziri, O.; el Battari, A.; Braguer, D.; el Ayeb, M.; Marrakchi, N.; Luis, J. MVL-PLA2, a snake venom phospholipase A2, inhibits angiogenesis through an increase in microtubule dynamics and disorganization of focal adhesions. PLoS One 2010, 5, e10124. [Google Scholar]
- Križaj, I.; Siigur, J.; Samel, M.; Cotiĉ, V.; Gubenšek, F. Isolation, partial characterization and complete amino acid sequence of the toxic phospholipase A2 from the venom of the common viper, Vipera berus berus. Biochim. Biophys. Acta 1993, 1157, 81–85. [Google Scholar]
- Ovchinnikov, Y.A.; Miroshnikov, A.I.; Nazimov, I.V.; Apsalaon, U.R.; Soldatova, L.N. Complete amino acid sequence of phospholipase A2 (isozyme E3) from the venom of middle Asian cobra Naja naja oxiana. J. Bioorg. Khim. 1979, 5, 805–813. [Google Scholar]
- Kahru, A. In vitro toxicity testing using marine luminescent bacteria Photobacterium phosphoreum: The BiotoxTM test. Atla-Altern. Lab. Anim. 1993, 21, 210–215. [Google Scholar]
- Yuan, Y.; Jackson, S.P.; Newnham, H.H.; Mitchell, C.A.; Salem, H.H. An essential role for lysophosphatidylcholine in the inhibition of platelet aggregation by secretory phospholipase A2. Blood 1995, 86, 4166–4174. [Google Scholar]
- Fuly, A.L.; de Miranda, A.L.P.; Zingali, R.B.; Guimarães, J.A. Purification and characterization of a phospholipase A2 isoenzyme isolated from Lachesis muta snake venom. Biochem. Pharmacol. 2002, 63, 1589–1597. [Google Scholar] [CrossRef]
- Gao, R.; Kini, R.M.; Gopalakhrishnakone, P. Purification, properties, and amino acid sequence of a hemoglobinuria-inducing phospholipase A2 MiPLA-1, from micropechis ikeheka venom. Arch. Biochem. Biophys. 1999, 369, 181–192. [Google Scholar] [CrossRef]
- Barbosa, P.S.; Martins, A.M.; Havt, A.; Toyama, D.O.; Evangelista, J.S.; Ferreira, D.P.; Joazeiro, P.P.; Beriam, L.O.; Toyama, M.H.; Fonteles, M.C.; Monteiro, H.S. Renal and antibacterial effects induced by myotoxin I and II isolated from Bothrops jararacussu venom. Toxicon 2005, 46, 376–386. [Google Scholar] [CrossRef]
- Costa, T.R.; Menaldo, D.L.; Oliveira, C.Z.; Santos-Filho, N.A.; Teixeira, S.S.; Nomizo, A.; Fuly, A.L.; Monteiro, M.C.; de Souza, B.M.; Palma, M.S.; et al. Myotoxic phospholipases A(2) isolated from Bothrops brazili snake venom and synthetic peptides derived from their C-terminal region: Cytotoxic effect on microorganism and tumor cells. Peptides 2008, 29, 1645–1656. [Google Scholar] [CrossRef]
- Glukhov, E.; Stark, M.; Burrows, L.L.; Deber, C.M. Basis for selectivity of cationic antimicrobial peptides for bacterial versus mammalian membranes. J. Biol. Chem. 2005, 280, 33960–33967. [Google Scholar]
- Cummings, B.S. Phospholipase A2 as targets for anti-cancer drugs. Biochem. Pharmacol. 2007, 74, 949–959. [Google Scholar] [CrossRef]
- Yan, C.H.; Liang, Z.Q.; Gu, Z.L.; Yang, Y.P.; Reid, P.; Qin, Z.H. Contributions of autophagic and apoptotic mechanisms to CrTX-induced death of K562 cells. Toxicon 2006, 47, 521–530. [Google Scholar] [CrossRef]
- Yan, C.H.; Yang, Y.P.; Qin, Z.H.; Gu, Z.L.; Reid, P.; Liang, Z.Q. Autophagy is involved in cytotoxic effects of crotoxin in human breast cancer cell line MCF-7 cells. Acta Pharmacol. Sin. 2007, 28, 540–548. [Google Scholar] [CrossRef]
- Ye, B.; Xie, Y.; Qin, Z.H.; Wu, J.C.; Han, R.; He, J.K. Anti-tumor activity of CrTX in human lung adenocarcinoma cell line A549. Acta Pharmacol. Sin. 2011, 32, 1397–1401. [Google Scholar] [CrossRef]
- Cura, J.E.; Blanzaco, D.P.; Brisson, C.; Cura, M.A.; Carbol, R.; Larrateguy, L.; Mendez, C.; Sechi, J.C.; Silveira, J.S.; Theiller, E.; de Roodt, A.R.; Vidal, J.C. Phase I and pharmacokinetics study crotoxin (cytotoxic PLA2 NSC-624 244) in patients with advanced cancer. Clin. Cancer Res. 2002, 8, 1033–1041. [Google Scholar]
- Higuchi, D.A.; Barbosa, C.M.V.; Bincoletto, C.; Chagas, J.R.; Magalhaes, A.; Richardson, M.; Sanches, E.F.; Pesquero, J.B.; Araujo, R.C.; Pesquero, J.L. Purification and partial characterization of two phospholipases A2 from Bothrops leucurus venom (white-tailed-jararaca) snake venom. Biochimie 2007, 89, 319–328. [Google Scholar] [CrossRef]
- Taketo, M.M.; Sanoshita, M. Phospholipase A2 and apoptosis. Biochim. Biophys. Acta 2002, 1585, 72–76. [Google Scholar]
- Sved, P.; Scott, K.F.; McLeod, D.; King, N.J.; Singh, J.; Tsatralis, T.; Nikolov, B.; Boulas, J.; Nallan, L.; Gelb, M.H.; et al. Oncogenic action of secreted phospholipase A2 in prostate cancer. Cancer Res. 2004, 64, 6934–6940. [Google Scholar] [CrossRef]
- De Haas, G.H.; Postema, N.M.; Nieuwenhuizen, W.; van Deenen, L.L.M. Purification and properties of phospholipase A from porcine pancreas. Biochim. Biophys. Acta 1968, 159, 103–117. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Vesterberg, O. Isoelectric focusing of proteins in polyacrylamide gels. Biochim. Biophys. Acta 1972, 257, 11–19. [Google Scholar] [CrossRef]
- Siigur, E.; Aaspõllu, A.; Trummal, K.; Tõnismägi, K.; Tammiste, I.; Kalkkinen, N.; Siigur, J. Factor X activator from Vipera lebetina venom is synthesized from different genes. Biochim. Biophys. Acta 2004, 1702, 41–51. [Google Scholar]
- Siigur, J.; Samel, M.; Tõnismägi, K.; Subbi, J.; Siigur, E.; Tu, A.T. Biochemical characterization of lebetase, a direct-acting fibrinolytic enzyme from Vipera lebetina snake venom. Thromb. Res. 1998, 90, 39–49. [Google Scholar] [CrossRef]
- Ivask, A.; Rõlova, T.; Kahru, A. A suite of recombinant luminescent bacterial strains for the quantification of bioavailable heavy metals and toxicity testing. BMC Biotechnol. 2009, 9, 41. [Google Scholar]
- Kurvet, I.; Ivask, A.; Bondarenko, O.; Sihtmäe, M.; Kahru, A. LuxCDABE—Transformed Constitutively Bioluminescent Escherichia coli for toxicity Screening: Comparison with naturally luminous Vibrio fischeri. Sensors 2011, 11, 7865–7878. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Samel, M.; Vija, H.; Kurvet, I.; Künnis-Beres, K.; Trummal, K.; Subbi, J.; Kahru, A.; Siigur, J. Interactions of PLA2-s from Vipera lebetina, Vipera berus berus and Naja naja oxiana Venom with Platelets, Bacterial and Cancer Cells. Toxins 2013, 5, 203-223. https://doi.org/10.3390/toxins5020203
Samel M, Vija H, Kurvet I, Künnis-Beres K, Trummal K, Subbi J, Kahru A, Siigur J. Interactions of PLA2-s from Vipera lebetina, Vipera berus berus and Naja naja oxiana Venom with Platelets, Bacterial and Cancer Cells. Toxins. 2013; 5(2):203-223. https://doi.org/10.3390/toxins5020203
Chicago/Turabian StyleSamel, Mari, Heiki Vija, Imbi Kurvet, Kai Künnis-Beres, Katrin Trummal, Juhan Subbi, Anne Kahru, and Jüri Siigur. 2013. "Interactions of PLA2-s from Vipera lebetina, Vipera berus berus and Naja naja oxiana Venom with Platelets, Bacterial and Cancer Cells" Toxins 5, no. 2: 203-223. https://doi.org/10.3390/toxins5020203