Biotoxin Detection Using Cell-Based Sensors



Abstract
:1. Introduction

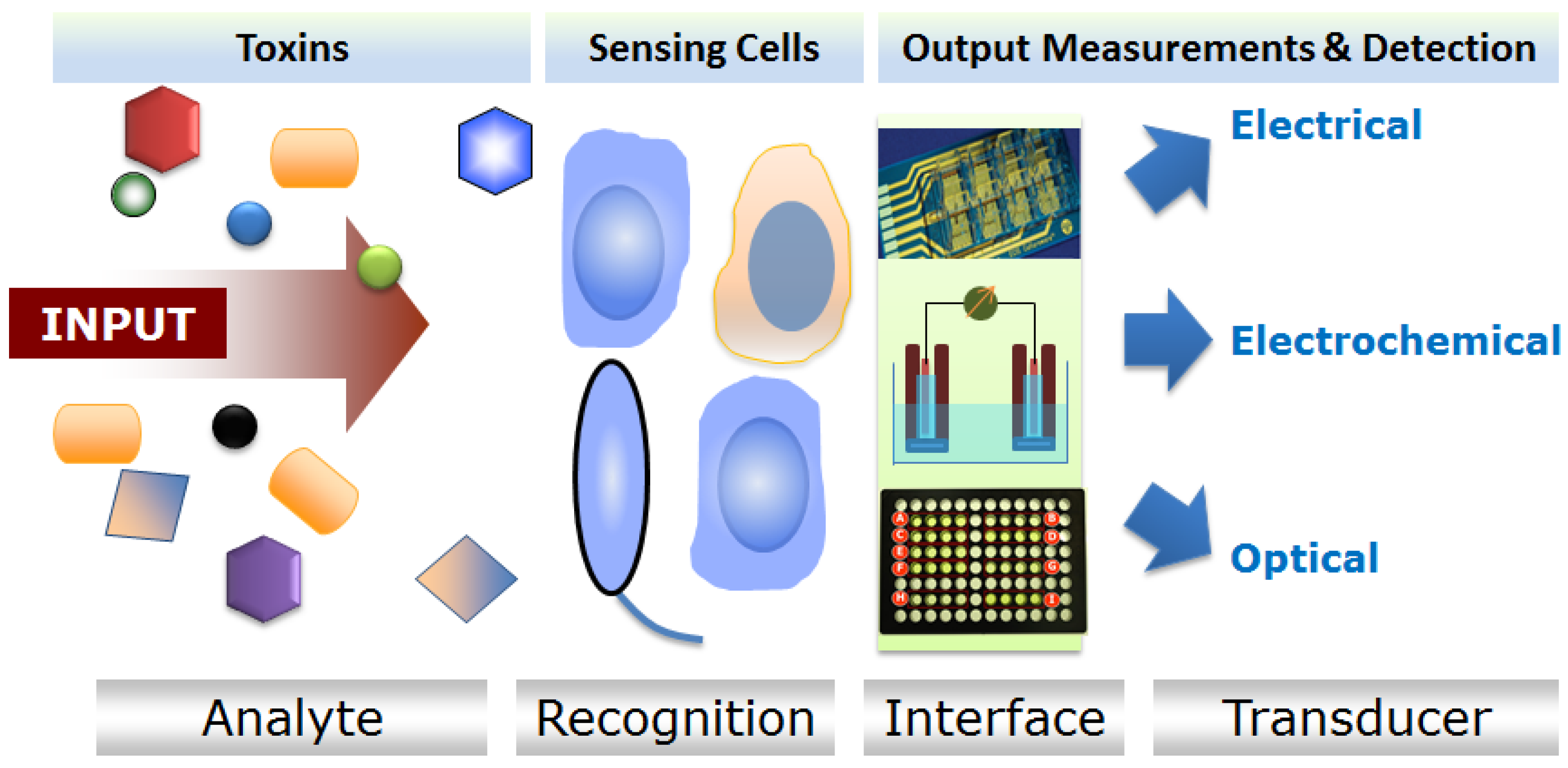{kind=link}
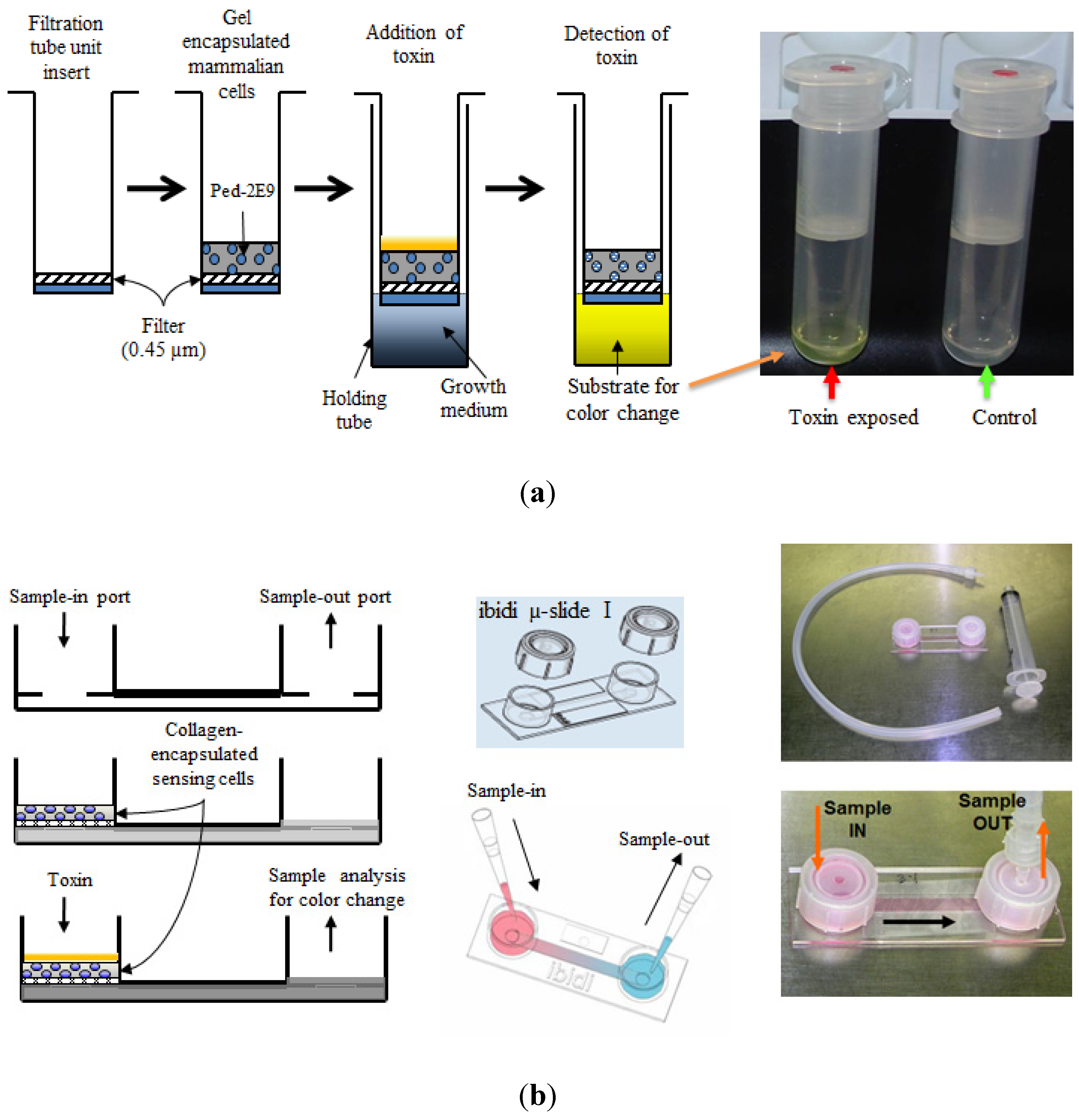{kind=link}
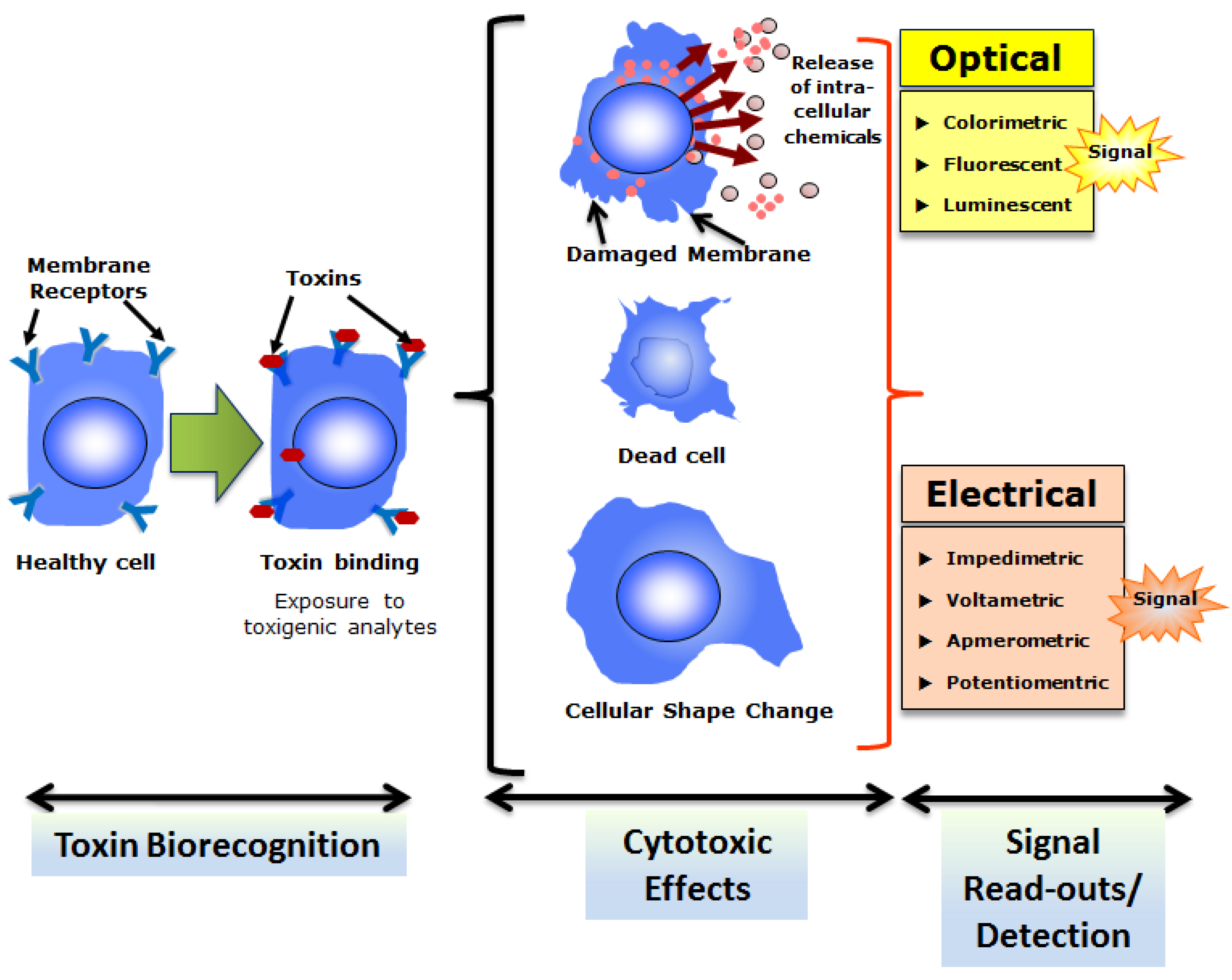{kind=link}
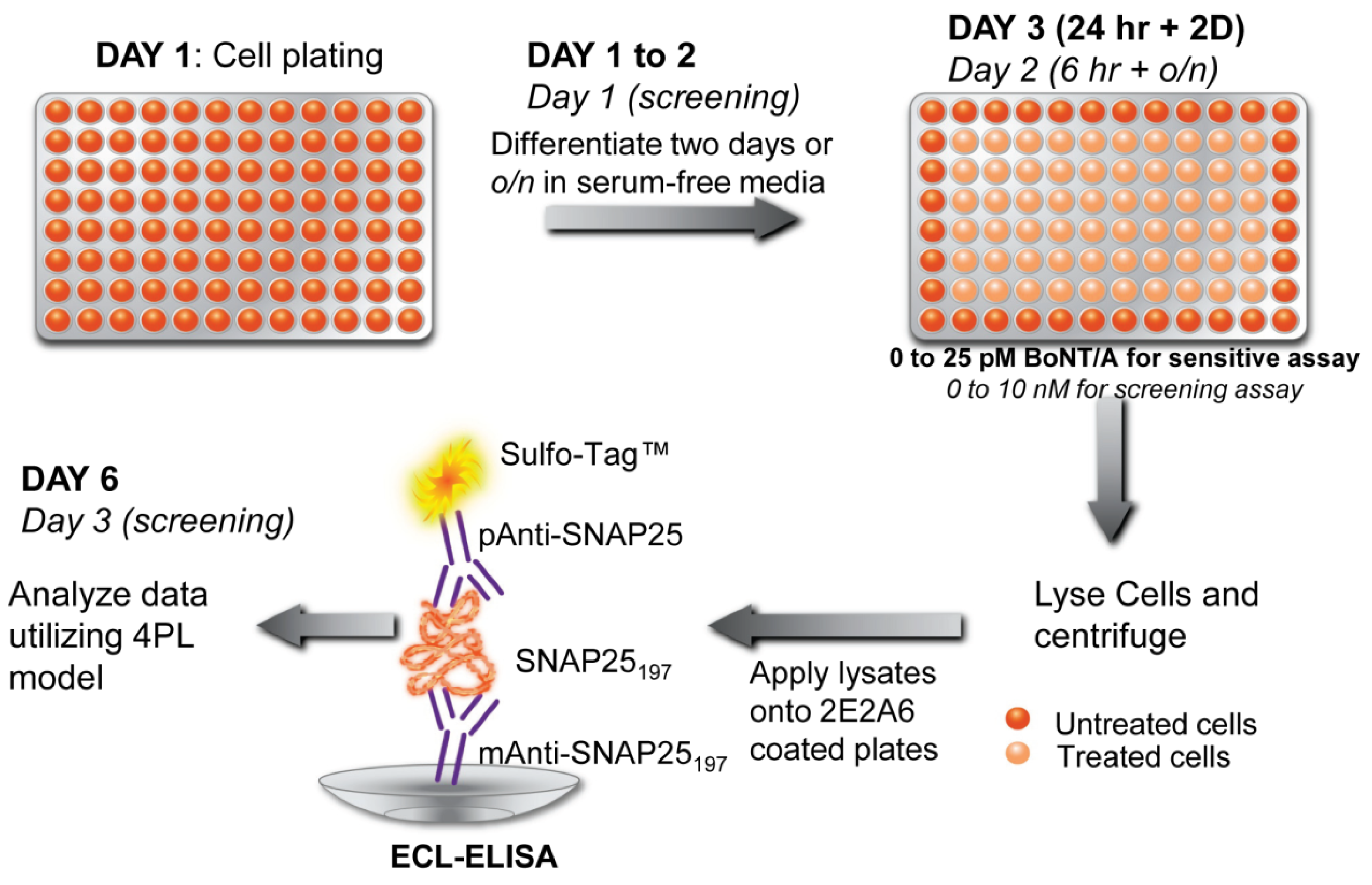{kind=link}
| Cell-Based Biosensors (CBBs) | |||||
|---|---|---|---|---|---|
| Microbial CBB | Invertebrate/Vertebrate/Mammalian CBB | ||||
| Sensor cell | Detection mode | Analyte [Reference] | Sensor cell | Detection mode | Analyte [Reference] |
| E. coli DH5α (pTOLGFP), (pTOLLUX) | Fluorescence, Bioluminescence | BTEX [9] | Hybridoma B-lymphocyte Ped-2E9 | Fluorescence/colorimetry | Listeriolysin O (LLO), B. cereus crude toxin, cytolysin, α-hemolysin, phospholipase C [10,11] |
| Pseudomonas putida KT2440 | Electrochemical | Aromatic hydrocarbons [12] | Green monkey kidney (Vero) | Bio-electric | Aflatoxin M1 [13] |
| E. coli DH5α (pJAMA-arsR) | Bioluminescence | Heavy metals (As, Sb) [14] | Hepatic (HepG2) | Optical/colorimetry | Marine toxins (azaspiracid-1, pectenotoxin-2, okadaic acid) [15,16] |
| P. putida JS444 | Amperometry | Organophosphate nerve agents [17] | Neuronal (Neuro2a) | Optical/colorimetry | Marine toxins (azaspiracid-1, pectenotoxin-2, okadaic acid) [15,16] |
| S. cerevisiae Y190 (modified) | Amperometry | Endocrine disruptor compounds [18] | Lung fibroblast (V79) | Optical/colorimetry | Mycotoxin (14 different types) [19] |
| E. coli (modified) | Amperometry | Genotoxicity [20] | Neuronal (mouse embryonic frontal cortex and spinal cord tissues) | Electric/microelectrode array (MEA) | Botulinum neurotoxin (BoNT/A) [21] |
| Salmonella typhimurium (modified) | Amperometry | Genotoxicity [22] | Vero | Optical/colorimetry | Mycotoxin (T-2, ZEN) [23] |
| Ralstonia eutropha ENV307 (pUTK60) | Bioluminescence | PCBs [24] | Neuronal (PC12) | Fluorescence/FRET | BoNT/A and BoNT/E [25] |
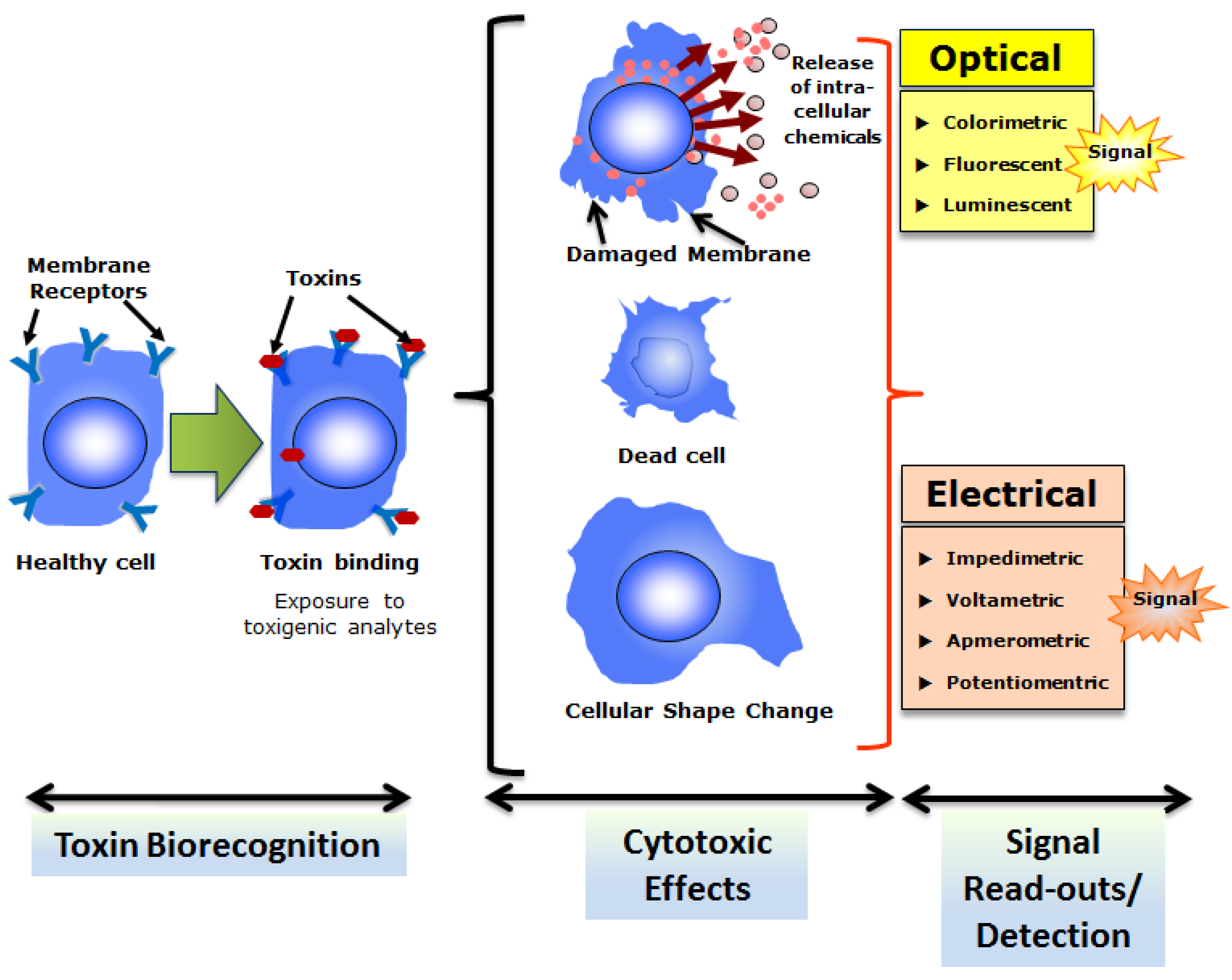
2. Choice of Biological Cells for Toxin Testing
2.1. Microbial CBBs
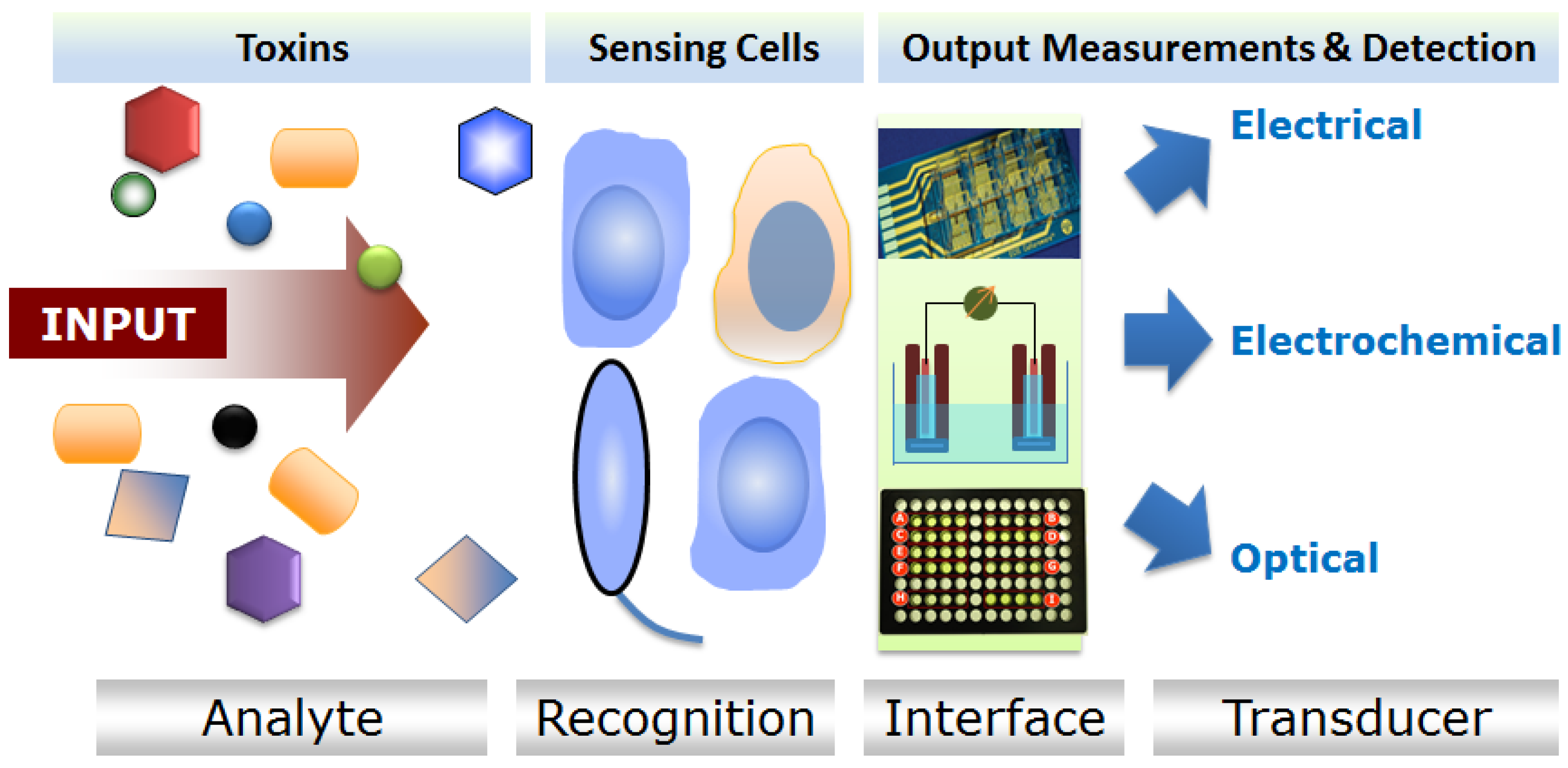
2.2. Higher Eukaryotic (Invertebrate and Vertebrate) CBBs
3. CBBs for Biotoxin Detection: Examples from Major Application Areas
3.1. Detection of Foodborne Toxins
3.1.1. Foodborne Bacterial Toxin Detection

3.1.2. Mycotoxin Detection
3.2. Detection of Marine Toxins
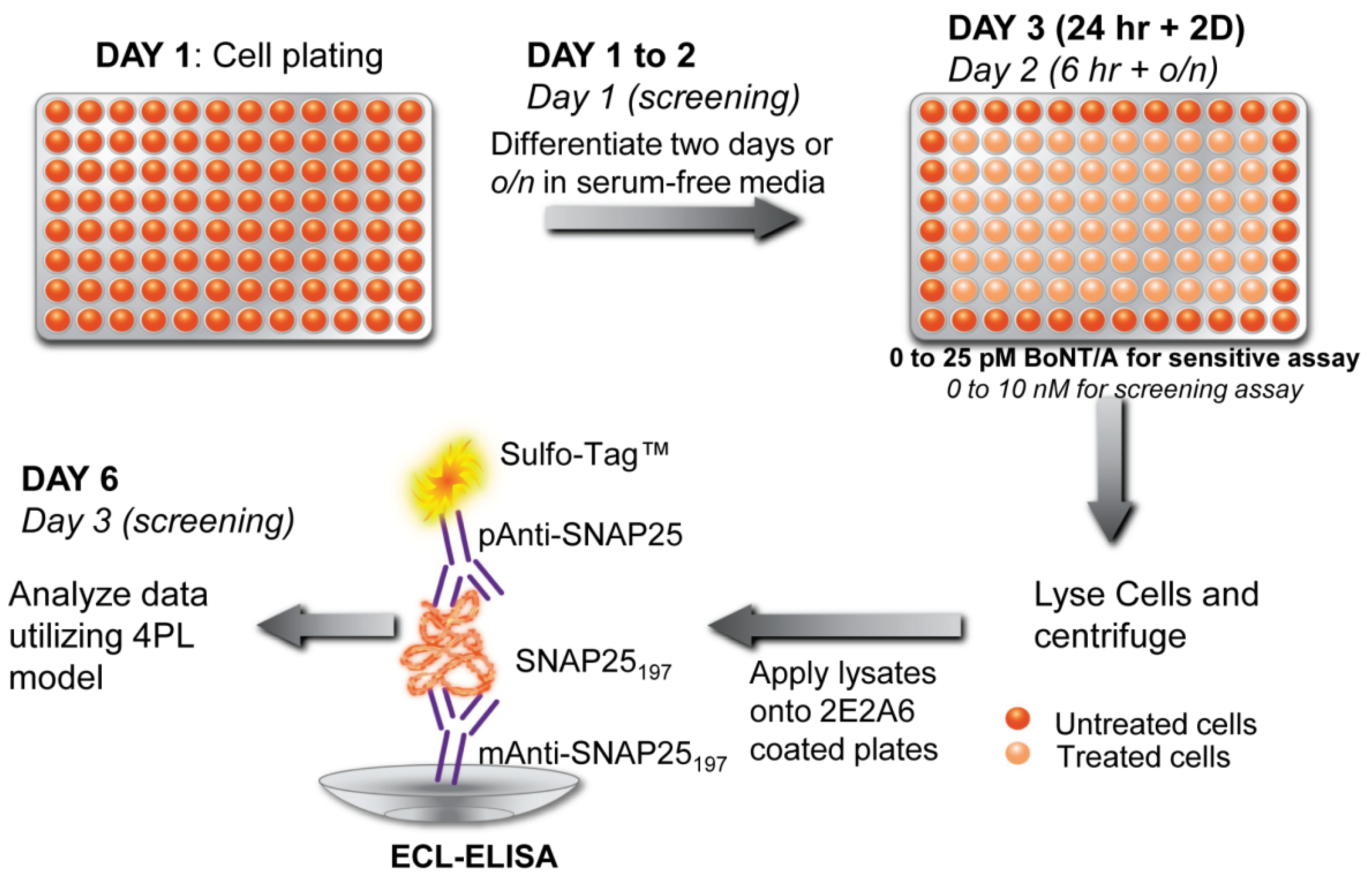
3.3. Detection of Botulinum Neurotoxins
3.4. Other Applications
4. The Challenges and Future of CBBs in Toxin Detection
Acknowledgments
Conflicts of Interest
References
- Pancrazio, J.J.; Whelan, J.P.; Borkholder, D.A.; Ma, W.; Stenger, D.A. Development and application of cell-based biosensors. Ann. Biomed. Eng. 1999, 27, 697–711. [Google Scholar] [CrossRef]
- Banerjee, P.; Bhunia, A.K. Mammalian cell-based biosensors for pathogens and toxins. Trends Biotechnol. 2009, 27, 179–188. [Google Scholar] [CrossRef]
- Banerjee, P.; Franz, B.; Bhunia, A. Mammalian cell-based sensor system. In Advances in Biochemical Engineering/Biotechnology: Whole Cell Sensing Systems I; Belkin, S., Gu, M.B., Eds.; Springer: Berlin Heidelberg, Germany, 2010; Volume 117, pp. 21–55. [Google Scholar]
- Kintzios, S.E. Cell-based biosensors in proteomic analysis. In Frontiers in Drug Design and Discovery; Caldwell, G.W., Rahman, A.U., D’Andrea, M.R., Choudhary, M.I., Eds.; Bentham Science Publishers: Sharjah, United Arab Emirates, 2006; Volume 2, pp. 225–239. [Google Scholar]
- Kintzios, S.E. Cell-based biosensors in clinical chemistry. Mini Rev. Med. Chem. 2007, 7, 1019–1026. [Google Scholar] [CrossRef]
- Stenger, D.A.; Gross, G.W.; Keefer, E.W.; Shaffer, K.M.; Andreadis, J.D.; Ma, W.; Pancrazio, J.J. Detection of physiologically active compounds using cell-based biosensors. Trends Biotechnol. 2001, 19, 304–309. [Google Scholar] [CrossRef]
- Janata, J. Role of analytical chemistry in defense strategies against chemical and biological attack. Ann. Rev. Anal. Chem. 2009, 2, 321–331. [Google Scholar] [CrossRef]
- O’Shaughnessy, T.J.; Pancrazio, J.J. Broadband detection of environmental neurotoxicants. Anal. Chem. 2007, 79, 8838–8845. [Google Scholar] [CrossRef]
- Li, Y.F.; Li, F.Y.; Ho, C.L.; Liao, V.H. Construction and comparison of fluorescence and bioluminescence bacterial biosensors for the detection of bioavailable toluene and related compounds. Environ. Pollut. 2008, 152, 123–129. [Google Scholar] [CrossRef]
- Banerjee, P.; Bhunia, A.K. Cell-based biosensor for rapid screening of pathogens and toxins. Biosens. Bioelectron. 2010, 26, 99–106. [Google Scholar] [CrossRef]
- Banerjee, P.; Lenz, D.; Robinson, J.P.; Rickus, J.L.; Bhunia, A.K. A novel and simple cell-based detection system with a collagen-encapsulated B-lymphocyte cell line as a biosensor for rapid detection of pathogens and toxins. Lab. Invest. 2008, 88, 196–206. [Google Scholar] [CrossRef]
- Paitan, Y.; Biran, I.; Shechter, N.; Biran, D.; Rishpon, J.; Ron, E.Z. Monitoring aromatic hydrocarbons by whole cell electrochemical biosensors. Anal. Biochem. 2004, 335, 175–183. [Google Scholar] [CrossRef]
- Larou, E.; Yiakoumettis, I.; Kaltsas, G.; Petropoulos, A.; Skandamis, P.; Kintzios, S. High throughput cellular biosensor for the ultra-sensitive, ultra-rapid detection of aflatoxin M1. Food Control 2013, 29, 208–212. [Google Scholar] [CrossRef]
- Stocker, J.; Balluch, D.; Gsell, M.; Harms, H.; Feliciano, J.; Daunert, S.; Malik, K.A.; Van der Meer, J.R. Development of a set of simple bacterial biosensors for quantitative and rapid measurements of arsenite and arsenate in potable water. Environ. Sci. Technol. 2003, 37, 4743–4750. [Google Scholar] [CrossRef]
- Sérandour, A.-L.; Ledreux, A.; Morin, B.; Derick, S.; Augier, E.; Lanceleur, R.; Hamlaoui, S.; Moukha, S.; Furger, C.; Biré, R.; et al. Collaborative study for the detection of toxic compounds in shellfish extracts using cell-based assays. Part I: Screening strategy and pre-validation study with lipophilic marine toxins. Anal. Bioanal. Chem. 2012, 403, 1983–1993. [Google Scholar] [CrossRef]
- Ledreux, A.; Sérandour, A.-L.; Morin, B.; Derick, S.; Lanceleur, R.; Hamlaoui, S.; Furger, C.; Biré, R.; Krys, S.; Fessard, V.; et al. Collaborative study for the detection of toxic compounds in shellfish extracts using cell-based assays. Part II: Application to shellfish extracts spiked with lipophilic marine toxins. Anal. Bioanal. Chem. 2012, 403, 1995–2007. [Google Scholar] [CrossRef]
- Lei, Y.; Mulchandani, P.; Wang, J.; Chen, W.; Mulchandani, A. Highly sensitive and selective amperometric microbial biosensor for direct determination of p-nitrophenyl-substituted organophosphate nerve agents. Environ. Sci. Technol. 2005, 39, 8853–8857. [Google Scholar]
- Ino, K.; Kitagawa, Y.; Watanabe, T.; Shiku, H.; Koide, M.; Itayama, T.; Yasukawa, T.; Matsue, T. Detection of hormone active chemicals using genetically engineered yeast cells and microfluidic devices with interdigitated array electrodes. Electrophoresis 2009, 30, 3406–3412. [Google Scholar] [CrossRef]
- Behm, C.; Follmann, W.; Degen, G.H. Cytotoxic potency of mycotoxins in cultures of V79 lung fibroblast cells. J. Toxicol. Environ. Health A 2012, 75, 1226–1231. [Google Scholar] [CrossRef]
- Biran, A.; Ben Yoav, H.; Yagur-Kroll, S.; Pedahzur, R.; Buchinger, S.; Shacham-Diamand, Y.; Reifferscheid, G.; Belkin, S. Microbial genotoxicity bioreporters based on sulA activation. Anal. Bioanal. Chem. 2011, 400, 3013–3024. [Google Scholar] [CrossRef]
- Pancrazio, J.J.; Gopal, K.; Keefer, E.W.; Gross, G.W. Botulinum toxin suppression of CNS network activity in vitro. J. Toxicol. 2013. Available online: http://www.hindawi.com/journals/jt/aip/732913/ (accessed on 26 November 2013).
- Buchinger, S.; Grill, P.; Morosow, V.; Ben-Yoav, H.; Shacham-Diamand, Y.; Biran, A.; Pedahzur, R.; Belkin, S.; Reifferscheid, G. Evaluation of chrono-amperometric signal detection for the analysis of genotoxicity by a whole cell biosensor. Anal. Chim. Acta. 2009, 659, 122–128. [Google Scholar]
- Bouaziz, C.; Bouslimi, A.; Kadri, R.; Zaied, C.; Bacha, H.; Abid-Essefi, S. The in vitro effects of zearalenone and T-2 toxins on Vero cells. Exp. Toxicol. Pathol. 2013, 65, 497–501. [Google Scholar] [CrossRef]
- Layton, A.C.; Muccini, M.; Ghosh, M.M.; Sayler, G.S. Construction of a bioluminescent reporter strain To detect polychlorinated biphenyls. Appl. Environ. Microbiol. 1998, 64, 5023–5026. [Google Scholar]
- Basavanna, U.; Muruvanda, T.; Brown, E.W.; Sharma, S.K. Development of a cell-based functional assay for the detection of clostridium botulinum neurotoxin types A and E. Int. J. Microbiol. 2013, 2013, 593219. [Google Scholar]
- Corcoran, C.A.; Rechnitz, G.A. Cell-based biosensors. Trends Biotechnol. 1985, 3, 92–96. [Google Scholar] [CrossRef]
- Rawson, D.M.; Willmer, A.J.; Turner, A.P. Whole-cell biosensors for environmental monitoring. Biosensors 1989, 4, 299–311. [Google Scholar] [CrossRef]
- Sidwell, J.S.; Rechnitz, G.A. Progress and challenges for biosensors using plant tissue materials. Biosensors 1986, 2, 221–233. [Google Scholar] [CrossRef]
- Williamson, K.J.; Johnson, D.G. A bacterial bioassay for assessment of wastewater toxicity. Water Res. 1981, 15, 383–390. [Google Scholar] [CrossRef]
- Slabbert, J.L.; Steyn, P.L.; Bateman, B.W.; Kfir, R. Rapid detection of toxicity in water using the oxygen uptake rate of mammalian cells as sensor. Water SA 1984, 10, 1–6. [Google Scholar]
- Giaever, I.; Keese, C.R. Monitoring fibroblast behavior in tissue culture with an applied electric field. Proc. Natl. Acad. Sci. USA 1984, 81, 3761–3764. [Google Scholar] [CrossRef]
- Bousse, L. Whole cell biosensors. Sens. Actuators B Chem. 1996, 34, 270–275. [Google Scholar] [CrossRef]
- Belkin, S. Microbial whole-cell sensing systems of environmental pollutants. Curr. Opin. Microbiol. 2003, 6, 206–212. [Google Scholar] [CrossRef]
- Woutersen, M.; Belkin, S.; Brouwer, B.; Van Wezel, A.P.; Heringa, M.B. Are luminescent bacteria suitable for online detection and monitoring of toxic compounds in drinking water and its sources? Anal. Bioanal. Chem. 2011, 400, 915–929. [Google Scholar] [CrossRef]
- King, J.M.; Digrazia, P.M.; Applegate, B.; Burlage, R.; Sanseverino, J.; Dunbar, P.; Larimer, F.; Sayler, G.S. Rapid, sensitive bioluminescent reporter technology for naphthalene exposure and biodegradation. Science 1990, 249, 778–781. [Google Scholar]
- Park, M.; Tsai, S.-L.; Chen, W. Microbial biosensors: engineered microorganisms as the sensing machinery. Sensors 2013, 13, 5777–5795. [Google Scholar] [CrossRef]
- Van der Meer, J.R.; Belkin, S. Where microbiology meets microengineering: Design and applications of reporter bacteria. Nat. Rev. Microbiol. 2010, 8, 511–522. [Google Scholar] [CrossRef]
- Eltzov, E.; Marks, R.S. Whole-cell aquatic biosensors. Anal. Bioanal. Chem. 2011, 400, 895–913. [Google Scholar] [CrossRef]
- Lagarde, F.; Jaffrezic-Renault, N. Cell-based electrochemical biosensors for water quality assessment. Anal. Bioanal. Chem. 2011, 400, 947–964. [Google Scholar] [CrossRef]
- Ivask, A.; Rolova, T.; Kahru, A. A suite of recombinant luminescent bacterial strains for the quantification of bioavailable heavy metals and toxicity testing. BMC Biotechnol. 2009, 9, 41. [Google Scholar] [CrossRef]
- Ramanathan, S.; Ensor, M.; Daunert, S. Bacterial biosensors for monitoring toxic metals. Trends Biotechnol. 1997, 15, 500–506. [Google Scholar] [CrossRef]
- Sochor, J.; Zitka, O.; Hynek, D.; Jilkova, E.; Krejcova, L.; Trnkova, L.; Adam, V.; Hubalek, J.; Kynicky, J.; Vrba, R.; et al. Bio-sensing of cadmium(II) ions using Staphylococcus aureus. Sensors 2011, 11, 10638–10663. [Google Scholar] [CrossRef]
- Mitchell, R.J.; Gu, M.B. Construction and evaluation of nagR-nagAa::lux fusion strains in biosensing for salicylic acid derivatives. Appl. Biochem. Biotechnol. 2005, 120, 183–198. [Google Scholar] [CrossRef]
- Trogl, J.; Kuncova, G.; Kubicova, L.; Parik, P.; Halova, J.; Demnerova, K.; Ripp, S.; Sayler, G.S. Response of the bioluminescent bioreporter Pseudomonas fluorescens HK44 to analogs of naphthalene and salicylic acid. Folia Microbiol. 2007, 52, 3–14. [Google Scholar] [CrossRef]
- Webb, O.F.; Bienkowski, P.R.; Matrubutham, U.; Evans, F.A.; Heitzer, A.; Sayler, G.S. Kinetics and response of a Pseudomonas fluorescens HK44 biosensor. Biotechnol. Bioeng. 1997, 54, 491–502. [Google Scholar] [CrossRef]
- Applegate, B.; Kelly, C.; Lackey, L.; McPherson, J.; Kehrmeyer, S.; Menn, F.M.; Bienkowski, P.; Sayler, G. Pseudomonas putida B2: A tod-lux bioluminescent reporter for toluene and trichloroethylene co-metabolism. J. Ind. Microbiol. Biotechnol. 1997, 18, 4–9. [Google Scholar] [CrossRef]
- Kelly, C.J.; Bienkowski, P.R.; Sayler, G.S. Kinetic analysis of a tod-lux bacterial reporter for toluene degradation and trichloroethylene cometabolism. Biotechnol. Bioeng. 2000, 69, 256–265. [Google Scholar] [CrossRef]
- Turner, K.; Xu, S.; Pasini, P.; Deo, S.; Bachas, L.; Daunert, S. Hydroxylated polychlorinated biphenyl detection based on a genetically engineered bioluminescent whole-cell sensing system. Anal. Chem. 2007, 79, 5740–5745. [Google Scholar] [CrossRef]
- Leedjarv, A.; Ivask, A.; Virta, M.; Kahru, A. Analysis of bioavailable phenols from natural samples by recombinant luminescent bacterial sensors. Chemosphere 2006, 64, 1910–1919. [Google Scholar] [CrossRef]
- Pham, H.T.; Parkinson, J.S. Phenol sensing by Escherichia coli chemoreceptors: A nonclassical mechanism. J. Bacteriol. 2011, 193, 6597–6604. [Google Scholar] [CrossRef]
- Taranova, L.; Semenchuk, I.; Manolov, T.; Iliasov, P.; Reshetilov, A. Bacteria-degraders as the base of an amperometric biosensor for detection of anionic surfactants. Biosens. Bioelectron. 2002, 17, 635–640. [Google Scholar] [CrossRef]
- De Las Heras, A.; De Lorenzo, V. Engineering whole-cell biosensors with no antibiotic markers for monitoring aromatic compounds in the environment. Methods Mol. Biol. 2012, 834, 261–281. [Google Scholar]
- Chan, A.C.; Ager, D.; Thompson, I.P. Resolving the mechanism of bacterial inhibition by plant secondary metabolites employing a combination of whole-cell biosensors. J. Microbiol. Methods. 2013, 93, 209–217. [Google Scholar] [CrossRef]
- Chen, Z.; Lu, M.; Zhuang, G.; Wang, H. Enhanced bacterial biosensor for fast and sensitive detection of oxidatively DNA damaging agents. Anal. Chem. 2011, 83, 3248–3251. [Google Scholar] [CrossRef]
- Bhunia, A.K. Biosensors and bio-based methods for the separation and detection of foodborne pathogens. In Advances in Food and Nutrition Research; Taylor, S., Ed.; Elsevier: San Diego, CA, USA, 2008; Volume 54, pp. 1–44. [Google Scholar]
- Rider, T.H.; Petrovick, M.S.; Nargi, F.E.; Harper, J.D.; Schwoebel, E.D.; Mathews, R.H.; Blanchard, D.J.; Bortolin, L.T.; Young, A.M.; Chen, J.; et al. A B cell-based sensor for rapid identification of pathogens. Science 2003, 301, 213–215. [Google Scholar] [CrossRef]
- Frega, M.; Pasquale, V.; Tedesco, M.; Marcoli, M.; Contestabile, A.; Nanni, M.; Bonzano, L.; Maura, G.; Chiappalone, M. Cortical cultures coupled to micro-electrode arrays: A novel approach to perform in vitro excitotoxicity testing. Neurotoxicol. Teratol. 2012, 34, 116–127. [Google Scholar] [CrossRef]
- Hogberg, H.T.; Sobanski, T.; Novellino, A.; Whelan, M.; Weiss, D.G.; Bal-Price, A.K. Application of micro-electrode arrays (MEAs) as an emerging technology for developmental neurotoxicity: Evaluation of domoic acid-induced effects in primary cultures of rat cortical neurons. Neurotoxicology 2011, 32, 158–168. [Google Scholar] [CrossRef]
- Van Thriel, C.; Westerink, R.H.; Beste, C.; Bale, A.S.; Lein, P.J.; Leist, M. Translating neurobehavioural endpoints of developmental neurotoxicity tests into in vitro assays and readouts. Neurotoxicology 2012, 33, 911–924. [Google Scholar] [CrossRef]
- Asphahani, F.; Thein, M.; Veiseh, O.; Edmondson, D.; Kosai, R.; Veiseh, M.; Xu, J.; Zhang, M. Influence of cell adhesion and spreading on impedance characteristics of cell-based sensors. Biosens. Bioelectron. 2008, 23, 1307–1313. [Google Scholar]
- Asphahani, F.; Thein, M.; Wang, K.; Wood, D.; Wong, S.S.; Xu, J.; Zhang, M. Real-time characterization of cytotoxicity using single-cell impedance monitoring. Analyst 2012, 137, 3011–3019. [Google Scholar] [CrossRef]
- Canete, E.; Diogene, J. Comparative study of the use of neuroblastoma cells (Neuro-2a) and neuroblastomaxglioma hybrid cells (NG108-15) for the toxic effect quantification of marine toxins. Toxicon 2008, 52, 541–550. [Google Scholar] [CrossRef]
- Kovacs, G.T. Electronic sensors with living cellular components. Proc. IEEE 2003, 91, 915–929. [Google Scholar] [CrossRef]
- Ben-Yoav, H.; Melamed, S.; Freeman, A.; Shacham-Diamand, Y.; Belkin, S. Whole-cell biochips for bio-sensing: Integration of live cells and inanimate surfaces. Crit. Rev. Biotechnol. 2011, 31, 337–353. [Google Scholar] [CrossRef]
- Bhunia, A.K.; Banada, P.; Banerjee, P.; Valadez, A.; Hirleman, E.D. Light scattering, fiber optic- and cell-based sensors for sensitive detection of foodborne pathogens. J. Rapid Meth. Autom. Microbiol. 2007, 15, 121–145. [Google Scholar] [CrossRef]
- Banerjee, P.; Morgan, M.T.; Rickus, J.L.; Ragheb, K.; Corvalan, C.; Robinson, J.P.; Bhunia, A.K. Hybridoma Ped-2E9 cells cultured under modified conditions can sensitively detect Listeria monocytogenes and Bacillus cereus. Appl. Microbiol. Biotechnol. 2007, 73, 1423–1434. [Google Scholar] [CrossRef]
- Gray, K.M.; Bhunia, A.K. Specific detection of cytopathogenic Listeria monocytogenes using a two-step method of immunoseparation and cytotoxicity analysis. J. Microbiol. Methods 2005, 60, 259–268. [Google Scholar] [CrossRef]
- Menon, A.; Shroyer, M.L.; Wampler, J.L.; Chawan, C.B.; Bhunia, A.K. In vitro study of Listeria monocytogenes infection to murine primary and human transformed B cells. Comp. Immunol. Microbiol. Infect. Dis. 2003, 26, 157–174. [Google Scholar] [CrossRef]
- Haggblom, M.M.; Apetroaie, C.; Andersson, M.A.; Salkinoja-Salonen, M.S. Quantitative analysis of cereulide, the emetic toxin of Bacillus cereus, produced under various conditions. Appl. Environ. Microbiol. 2002, 68, 2479–2483. [Google Scholar] [CrossRef]
- Beattie, S.H.; Williams, A.G. Detection of toxigenic strains of Bacillus cereus and other Bacillus spp. with an improved cytotoxicity assay. Lett. Appl. Microbiol. 1999, 28, 221–225. [Google Scholar] [CrossRef]
- Rowan, N.J.; Caldow, G.; Gemmell, C.G.; Hunter, I.S. Production of diarrheal enterotoxins and other potential virulence factors by veterinary isolates of Bacillus species associated with nongastrointestinal infections. Appl. Environ. Microbiol. 2003, 69, 2372–2376. [Google Scholar] [CrossRef]
- Schoeni, J.L.; Wong, A.C. Bacillus cereus food poisoning and its toxins. J. Food Prot. 2005, 68, 636–648. [Google Scholar]
- Gray, K.M.; Banada, P.P.; O’Neal, E.; Bhunia, A.K. Rapid Ped-2E9 cell-based cytotoxicity analysis and genotyping of Bacillus species. J. Clin. Microbiol. 2005, 43, 5865–5872. [Google Scholar] [CrossRef]
- Gutleb, A.C.; Morrison, E.; Murk, A.J. Cytotoxicity assays for mycotoxins produced by Fusarium strains: A review. Environ. Toxicol. Pharmacol. 2002, 11, 309–320. [Google Scholar] [CrossRef]
- Nielsen, C.; Casteel, M.; Didier, A.; Dietrich, R.; Martlbauer, E. Trichothecene-induced cytotoxicity on human cell lines. Mycotoxin Res. 2009, 25, 77–84. [Google Scholar] [CrossRef]
- Moschopoulou, G.; Kintzios, S. Application of “membrane-engineering” to bioelectric recognition cell sensors for the ultra-sensitive detection of superoxide radical: A novel biosensor principle. Anal. Chim. Acta. 2006, 573/574, 90–96. [Google Scholar] [CrossRef]
- Moschopoulou, G.; Vitsa, K.; Bem, F.; Vassilakos, N.; Perdikaris, A.; Blouhos, P.; Yialouris, C.; Frosyniotis, D.; Anthopoulos, I.; Mangana, O.; et al. Engineering of the membrane of fibroblast cells with virus-specific antibodies: A novel biosensor tool for virus detection. Biosens. Bioelectron. 2008, 24, 1027–1030. [Google Scholar] [CrossRef]
- Vilariño, N.; Louzao, M.C.; Vieytes, M.R.; Botana, L.M. Biological methods for marine toxin detection. Anal. Bioanal. Chem. 2010, 397, 1673–1681. [Google Scholar] [CrossRef]
- Kulagina, N.V.; Mikulski, C.M.; Gray, S.; Ma, W.; Doucette, G.J.; Ramsdell, J.S.; Pancrazio, J.J. Detection of marine toxins, brevetoxin-3 and saxitoxin, in seawater using neuronal networks. Environ. Sci. Technol. 2006, 40, 578–583. [Google Scholar] [CrossRef]
- Capek, P.; Dickerson, T.J. Sensing the deadliest toxin: Technologies for botulinum neurotoxin detection. Toxins 2010, 2, 24–53. [Google Scholar] [CrossRef]
- Jones, R.G.; Liu, Y.; Halls, C.; Thorpe, S.J.; Longstaff, C.; Matejtschuk, P.; Sesardic, D. Release of proteolytic activity following reduction in therapeutic human serum albumin containing products: Detection with a new neoepitope endopeptidase immunoassay. J. Pharm. Biomed. Anal. 2011, 54, 74–80. [Google Scholar] [CrossRef]
- Pellett, S. Progress in cell based assays for botulinum neurotoxin detection. Curr. Top. Microbiol. Immunol. 2013, 364, 257–285. [Google Scholar]
- Dorner, M.B.; Schulz, K.M.; Kull, S.; Dorner, B.G. Complexity of botulinum neurotoxins: Challenges for detection technology. Curr. Top. Microbiol. Immunol. 2012, 364, 219–255. [Google Scholar]
- Whitemarsh, R.C.; Strathman, M.J.; Chase, L.G.; Stankewicz, C.; Tepp, W.H.; Johnson, E.A.; Pellett, S. Novel application of human neurons derived from induced pluripotent stem cells for highly sensitive botulinum neurotoxin detection. Toxicol. Sci. 2012, 126, 426–435. [Google Scholar] [CrossRef]
- Fernandez-Salas, E.; Wang, J.; Molina, Y.; Nelson, J.B.; Jacky, B.P.; Aoki, K.R. Botulinum neurotoxin serotype A specific cell-based potency assay to replace the mouse bioassay. PLoS One 2012, 7, e49516. [Google Scholar]
- Scarlatos, A.; Cadotte, A.J.; DeMarse, T.B.; Welt, B.A. Cortical networks grown on microelectrode arrays as a biosensor for botulinum toxin. J. Food Sci. 2008, 73, E129–E136. [Google Scholar] [CrossRef]
- Gopal, K.V.; Wu, C.; Shrestha, B.; Campbell, K.C.; Moore, E.J.; Gross, G.W. d-Methionine protects against cisplatin-induced neurotoxicity in cortical networks. Neurotoxicol. Teratol. 2012, 34, 495–504. [Google Scholar] [CrossRef]
- Relman, D.A. Shedding light on microbial detection. N. Engl. J. Med. 2003, 349, 2162–2163. [Google Scholar]
- Curtis, T.M.; Collins, A.M.; Gerlach, B.D.; Brennan, L.M.; Widder, M.W.; Van der Schalie, W.H.; Vo, N.T.K.; Bols, N.C. Suitability of invertebrate and vertebrate cells in a portable impedance-based toxicity sensor: Temperature mediated impacts on long-term survival. Toxicol. In Vitro 2013, 27, 2061–2066. [Google Scholar] [CrossRef]
- Brennan, L.M.; Widder, M.W.; Lee, L.E.J.; Van der Schalie, W.H. Long-term storage and impedance-based water toxicity testing capabilities of fluidic biochips seeded with RTgill-W1 cells. Toxicol. In Vitro 2012, 26, 736–745. [Google Scholar] [CrossRef]
- Petrovick, M.S.; Nargi, F.E.; Towle, T.; Hogan, K.; Bohane, M.; Wright, D.J.; MacRae, T.H.; Potts, M.; Helm, R.F. Improving the long-term storage of a mammalian biosensor cell line via genetic engineering. Biotechnol. Bioeng. 2010, 106, 474–481. [Google Scholar]
- Kiilerich-Pedersen, K.; Rozlosnik, N. Cell-based biosensors: Electrical sensing in microfluidic devices. Diagnostics 2012, 2, 83–96. [Google Scholar] [CrossRef]
- Prabhakarpandian, B.; Shen, M.C.; Pant, K.; Kiani, M.F. Microfluidic devices for modeling cell-cell and particle-cell interactions in the microvasculature. Microvasc. Res. 2011, 82, 210–220. [Google Scholar] [CrossRef]
- Giulitti, S.; Magrofuoco, E.; Prevedello, L.; Elvassore, N. Optimal periodic perfusion strategy for robust long-term microfluidic cell culture. Lab. Chip. 2013, 13, 4430–4441. [Google Scholar] [CrossRef]
- Mairhofer, J.; Roppert, K.; Ertl, P. Microfluidic systems for pathogen sensing: A review. Sensors 2009, 9, 4804–4823. [Google Scholar] [CrossRef]
- Hong, W.S.; Young, E.W.; Tepp, W.H.; Johnson, E.A.; Beebe, D.J. A microscale neuron and Schwann cell coculture model for increasing detection sensitivity of botulinum neurotoxin type A. Toxicol. Sci. 2013, 134, 64–72. [Google Scholar] [CrossRef]
- Gross, G.W.; Pancrazio, J.J. Neuronal network biosensors. In Smart Biosensor Technology; Knopf, G.K., Bassi, A.S., Eds.; Taylor and Francis Publishers, CRC Press: Boca Raton, FL, USA, 2006; pp. 177–201. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Banerjee, P.; Kintzios, S.; Prabhakarpandian, B. Biotoxin Detection Using Cell-Based Sensors. Toxins 2013, 5, 2366-2383. https://doi.org/10.3390/toxins5122366
Banerjee P, Kintzios S, Prabhakarpandian B. Biotoxin Detection Using Cell-Based Sensors. Toxins. 2013; 5(12):2366-2383. https://doi.org/10.3390/toxins5122366
Chicago/Turabian StyleBanerjee, Pratik, Spyridon Kintzios, and Balabhaskar Prabhakarpandian. 2013. "Biotoxin Detection Using Cell-Based Sensors" Toxins 5, no. 12: 2366-2383. https://doi.org/10.3390/toxins5122366