An Extract of Rhodobacter sphaeroides Reduces Cisplatin-Induced Nephrotoxicity in Mice

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
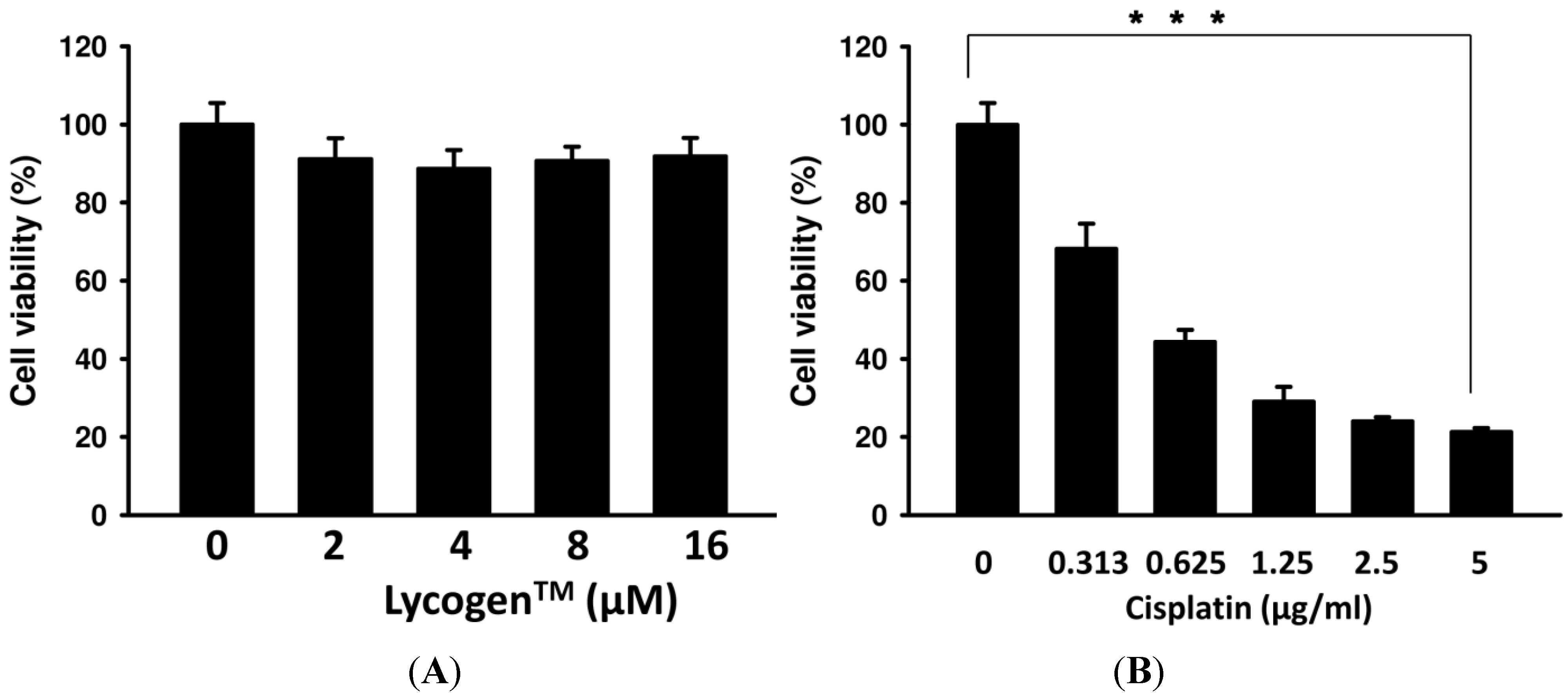
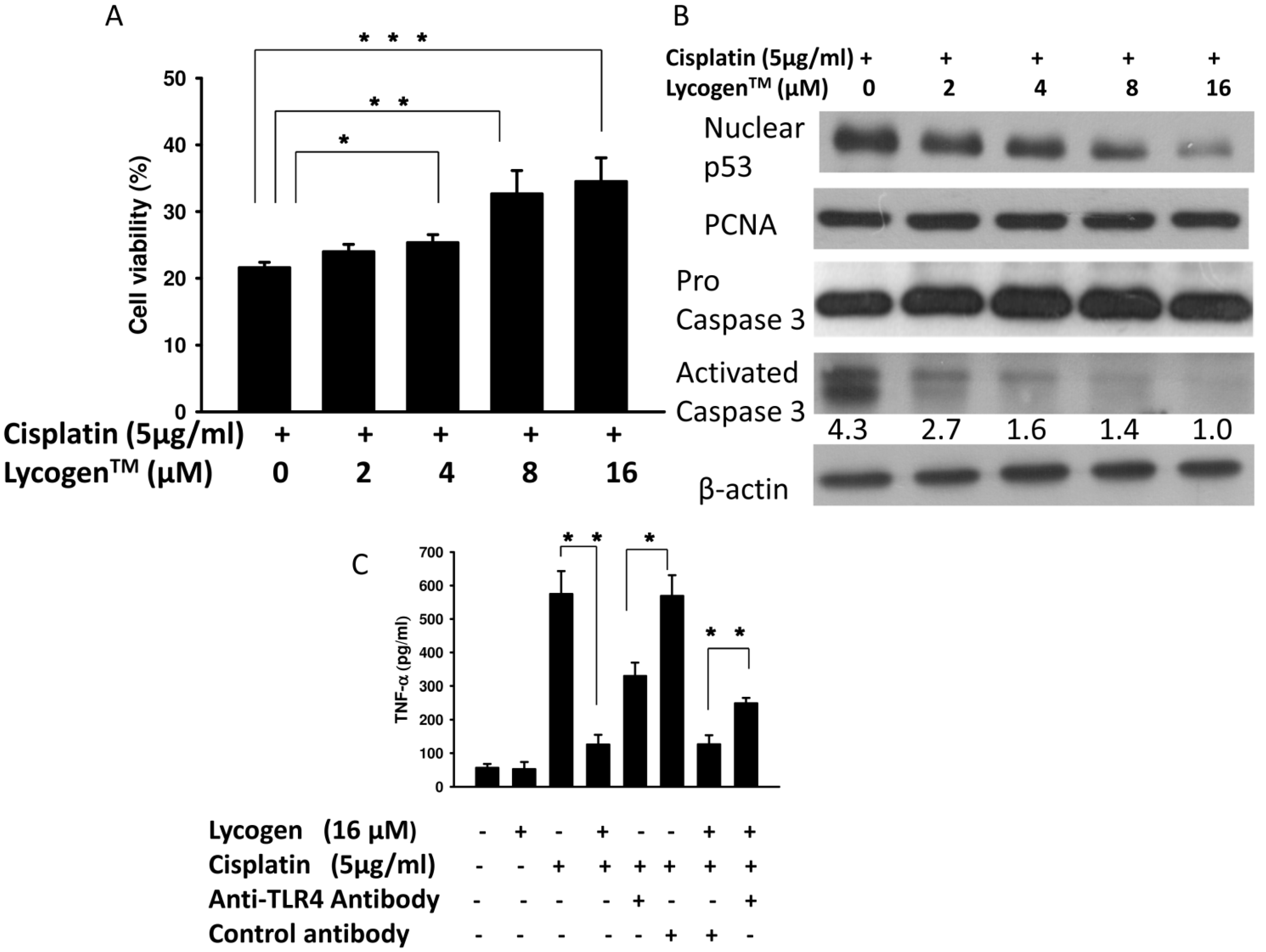
2.1. Evaluation of Cell Viability after Lycogen™ or Cisplatin Treatment in Vitro
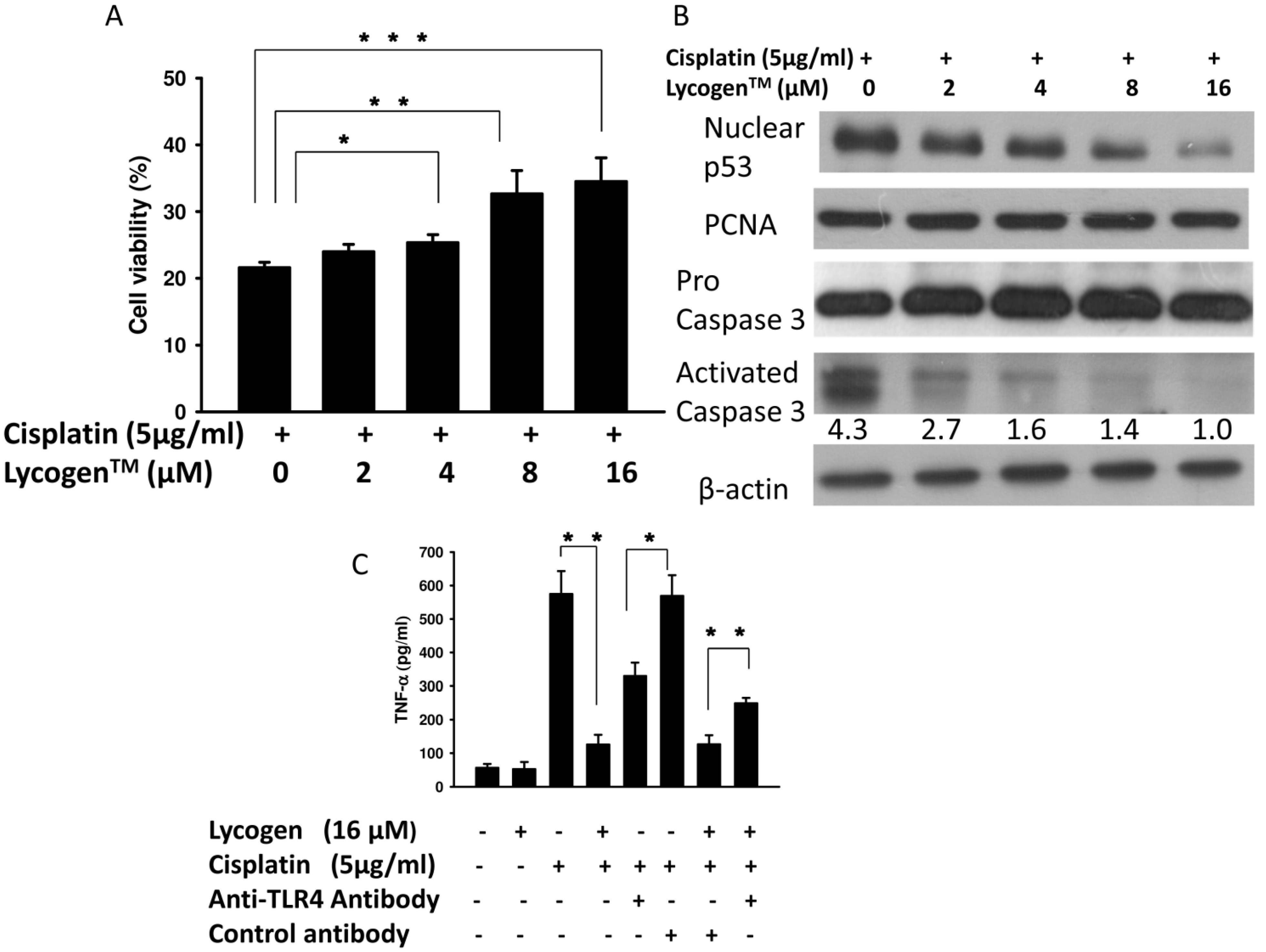
2.2. Lycogen™ Treatment Attenuated Cisplatin-Induced Renal Cell Death

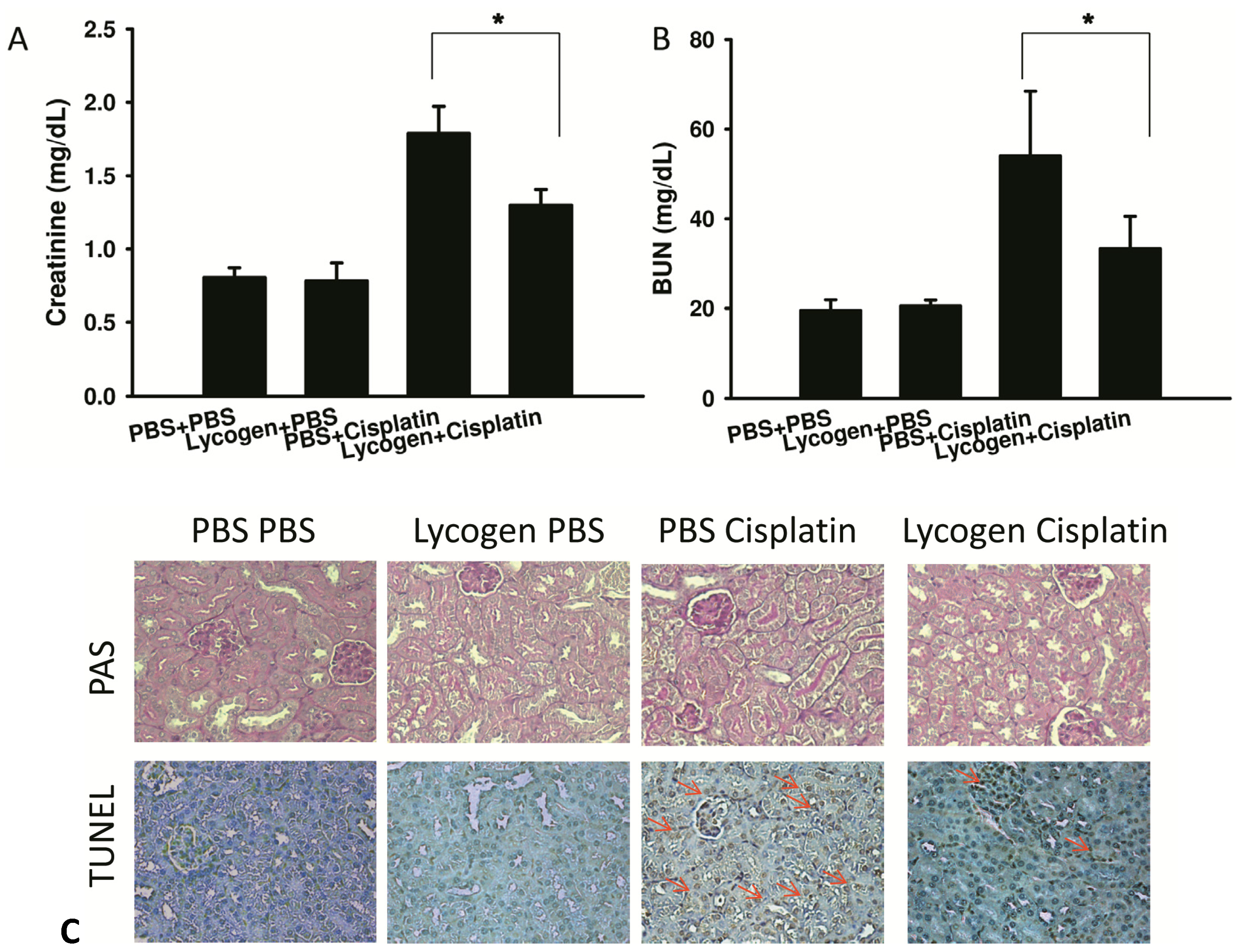
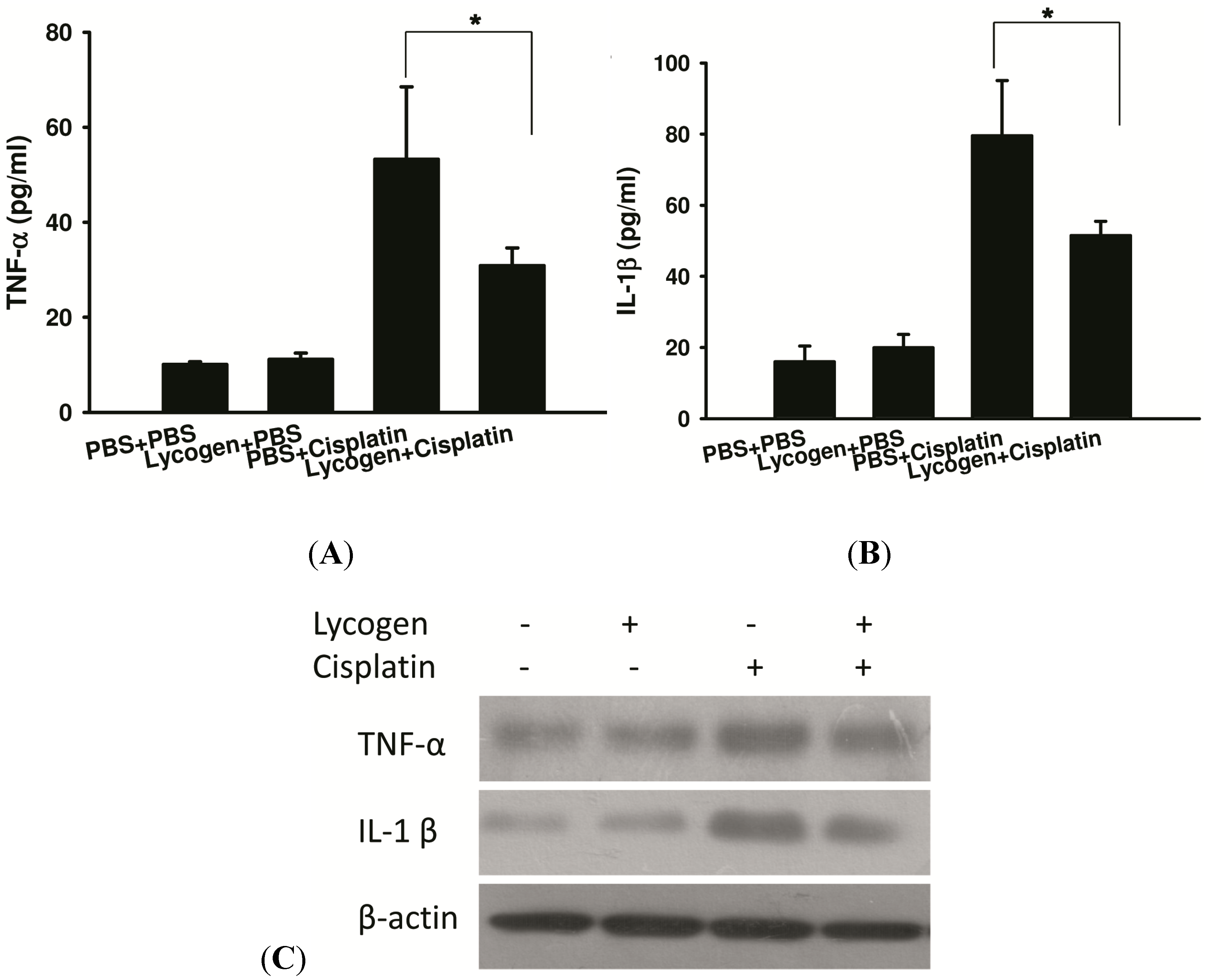
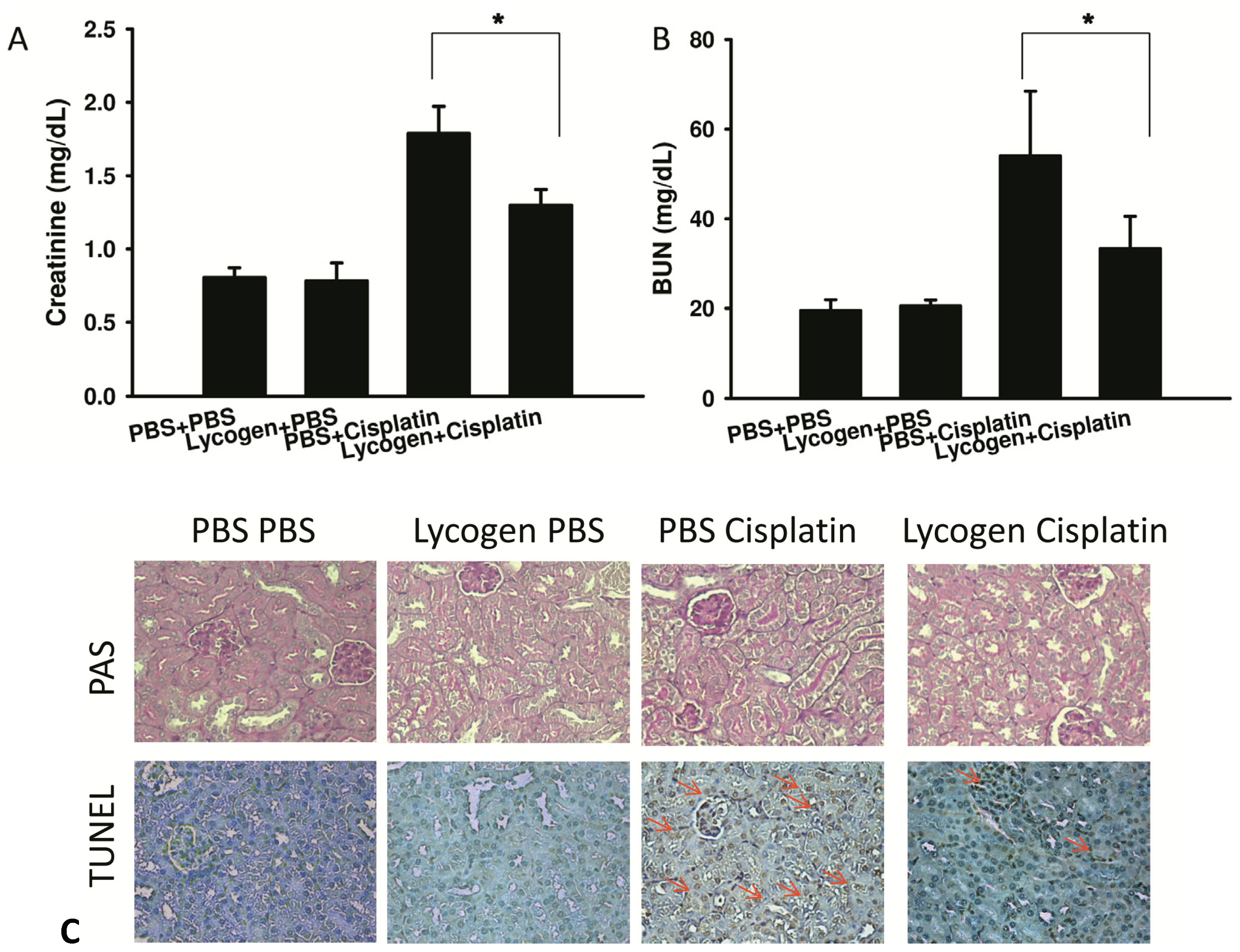
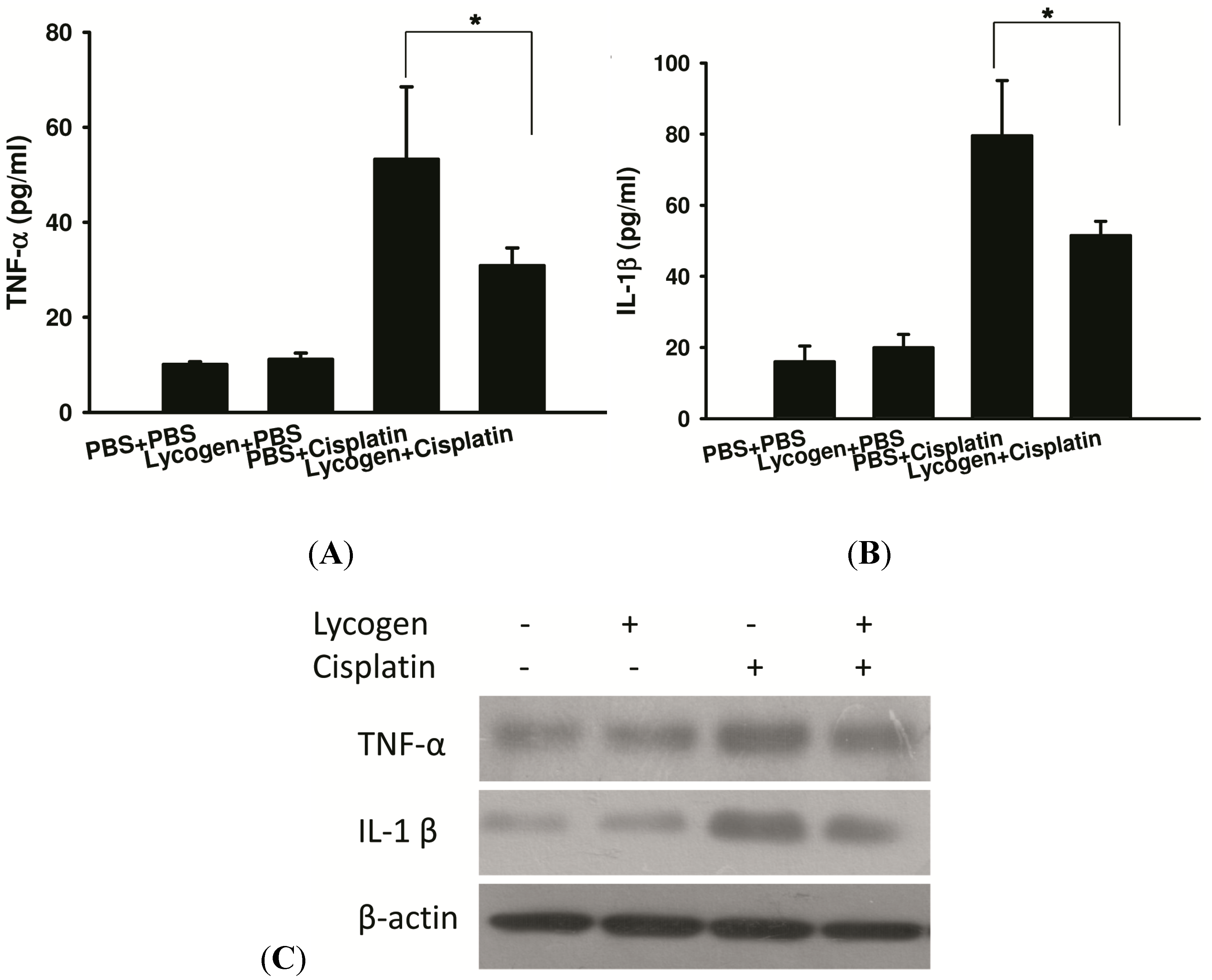
2.3. Lycogen™ Treatment Attenuated Renal Injury in Cisplatin-Treated Mice
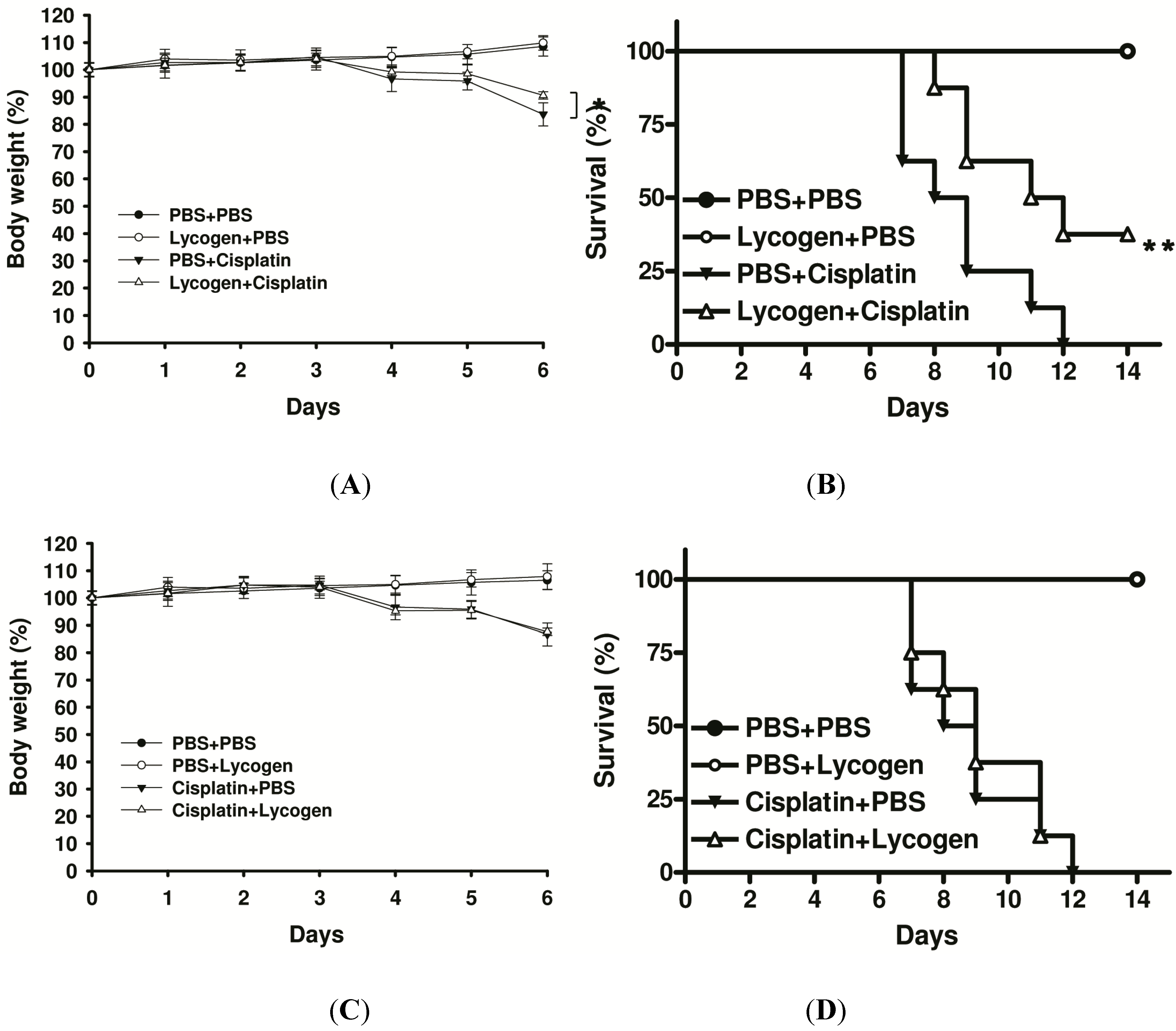
2.4. Lycogen™ Improved Cisplatin-Induced Body Weight Loss and Survival
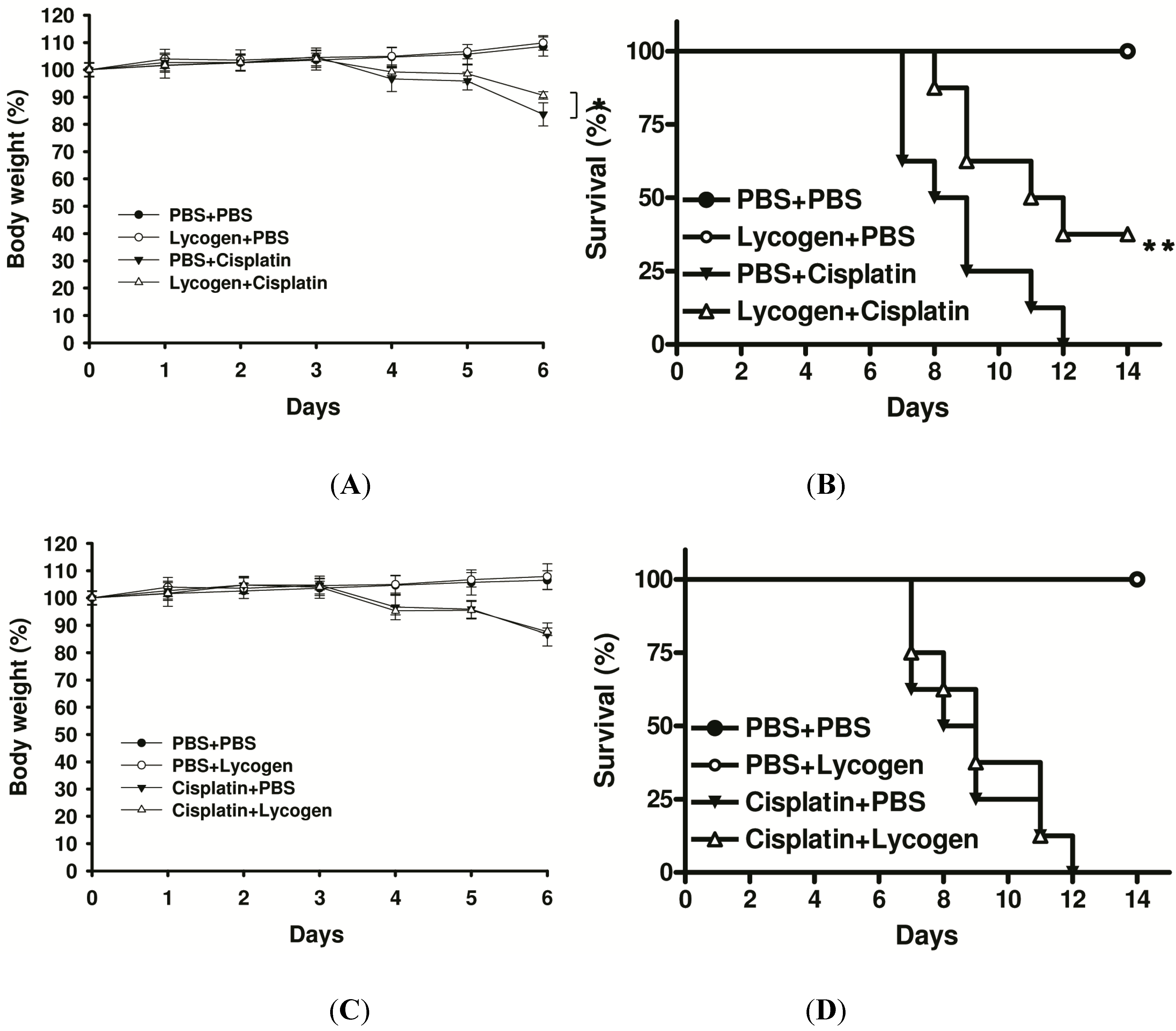
3. Discussion
4. Experimental Section
4.1. Reagents, Cells and Mice
4.2. Cell Proliferation Assay
4.3. Western Blot Analysis
4.4. Establishment of Experimental Renal Injury Model
4.5. Determination of Creatinine and BUN
4.6. Assessment of Cytokines
4.7. Histological Examinations
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Pabla, N.; Dong, Z. Cisplatin neophrotoxicity: Mechanisms and renoprotective strategies. Kindney Int. 2008, 73, 994–1007. [Google Scholar] [CrossRef]
- Wu, S.L.; Chen, J.C.; Li, C.C.; Lo, H.Y.; Ho, T.Y.; Hsiang, C.Y. Vanillin improves and prevents trinitrobenzene sulfonic acid-induced colitis in mice. J. Pharmacol. Exp. Ther. 2009, 330, 370–376. [Google Scholar] [CrossRef]
- Hsiang, C.Y.; Lo, H.Y.; Huang, H.C.; Li, C.C.; Wu, S.L.; Ho, T.Y. Ginger extract and zingerone ameliorated trinitrobenzene sulphonic acid-induced colitis in mice via modulation of nuclear factor-κB activity and interleukin-1β signaling pathway. Food Chem. 2013, 136, 170–177. [Google Scholar]
- Chen, Y.; Graham, A.; Potter, W.; Morgan, J.; Vaughan, L.; Bellnier, D.A.; Henderson, B.W.; Oseroff, A.; Dougherty, T.J.; Pandey, R.K. Bacteriopurpurinimides: Highly stable and potent photosensitizers for photodynamic therapy. J. Med. Chem. 2002, 45, 255–258. [Google Scholar] [CrossRef]
- Deshmukh, S.S.; Tang, K.; Kálmán, L. Lipid binding to the carotenoid binding site in photosynthetic reaction centers. J. Am. Chem. Soc. 2011, 133, 16309–16316. [Google Scholar] [CrossRef]
- Kelsey, L.; Katoch, P.; Johnson, K.E.; Batra, S.K.; Mehta, P.P. Retinoids regulate the formation and degradation of gap junctions in androgen-responsive human prostate cancer cells. PLoS One 2012, 7, e32846. [Google Scholar]
- Liu, W.S.; Chen, M.C.; Chiu, K.H.; Wen, Z.H.; Lee, C.H. Amelioration of dextran sodium sulfate-induced colitis in mice by Rhodobacter sphaeroides extract. Molecules 2012, 17, 13622–13630. [Google Scholar]
- Liu, W.S.; Kuan, Y.D.; Chiu, K.H.; Wang, W.K.; Chang, F.H.; Liu, C.H.; Lee, C.H. The Extract of Rhodobacter sphaeroides Inhibits Melanogenesis through the MEK/ERK Signaling Pathway. Mar. Drugs 2013, 11, 1899–1908. [Google Scholar] [CrossRef]
- Cummings, B.S.; Schnellmann, R.G. Cisplatin-induced renal cell apoptosis: Caspase 3-dependent and -independent pathways. J. Pharmacol. Exp. Ther. 2002, 302, 8–17. [Google Scholar] [CrossRef]
- Sahu, B.D.; Kuncha, M.; Sindura, G.J.; Sistla, R. Hesperidin attenuates cisplatin-induced acute renal injury by decreasing oxidative stress, inflammation and DNA damage. Phytomedicine 2013, 20, 453–460. [Google Scholar] [CrossRef]
- Zhang, B.; Ramesh, G.; Uematsu, S.; Akira, S.; Reeves, W.B. TLR4 signaling mediates inflammation and tissue injury in nephrotoxicity. J. Am. Soc. Nephrol. 2008, 19, 923–932. [Google Scholar]
- Lu, Z.; Zhang, X.; Li, Y.; Jin, J.; Huang, Y. TLR4 antagonist reduces early-stage atherosclerosis in diabetic apolipoprotein E-deficient mice. J. Endocrinol. 2013, 216, 61–71. [Google Scholar] [CrossRef]
- Loppnow, H.; Libby, P.; Freudenberg, M.; Krauss, J.H.; Weckesser, J.; Mayer, H. Cytokine induction by lipopolysaccharide (LPS) corresponds to lethal toxicity and is inhibited by nontoxic Rhodobacter capsulatus LPS. Infect. Immun. 1990, 58, 3743–3750. [Google Scholar]
- Seth, R.; Yang, C.; Kaushal, V.; Shah, S.V.; Kaushal, G.P. p53-dependent caspase-2 activation in mitochondrial release of apoptosis-inducing factor and its role in renal tubular epithelial cell injury. J. Biol. Chem. 2005, 280, 31230–31239. [Google Scholar]
- Yang, C.; Kaushal, V.; Haun, R.S.; Seth, R.; Shah, S.V.; Kaushal, G.P. Transcriptional activation of caspase-6 and -7 genes by cisplatin-induced p53 and its functional significance in cisplatin nephrotoxicity. Cell Death Differ. 2008, 15, 530–544. [Google Scholar] [CrossRef]
- Kaushal, G.P.; Liu, L.; Kaushal, V.; Hong, X.; Melnyk, O.; Seth, R.; Safirstein, R.; Shah, S.V. Regulation of caspase-3 and -9 activation in oxidant stress to RTE by forkhead transcription factors, Bcl-2 proteins, and MAP kinases. Am. J. Physiol. Renal. Physiol. 2004, 287, F1258–F1268. [Google Scholar]
- Kirikae, T.; Schade, F.U.; Kirikae, F.; Qureshi, N.; Takayama, K.; Rietschel, E.T. Diphosphoryl lipid A derived from the lipopolysaccharide (LPS) of Rhodobacter sphaeroides ATCC 17023 is a potent competitive LPS inhibitor in murine macrophage-like J774.1 cells. FEMS Immuno. Med. Microbiol. 1994, 9, 237–243. [Google Scholar]
- Jarvis, B.W.; Lichenstein, H.; Qureshi, N. Diphosphoryl lipid A from Rhodobacter sphaeroides inhibits complexes that form in vitro between lipopolysaccharide (LPS)-binding protein, soluble CD14, and spectrally pure LPS. Infect. Immun. 1997, 65, 3011–3016. [Google Scholar]
- Aida, Y.; Kusumoto, K.; Nakatomi, K.; Takada, H.; Pabst, M.J.; Maeda, K. An analogue of lipid A and LPS from Rhodobacter sphaeroides inhibits neutrophil responses to LPS by blocking receptor recognition of LPS and by depleting LPS-binding protein in plasma. J. Leukoc. Biol. 1995, 58, 675–682. [Google Scholar]
- Albrecht, M.; Klein, A.; Hugueney, P.; Sandmann, G.; Kuntz, M. Molecular cloning and functional expression in E. coli of a novel plant enzyme mediating zeta-carotene desaturation. FEBS Lett. 1995, 372, 199–202. [Google Scholar] [CrossRef]
- Sahin, K.; Tuzcu, M.; Sahin, N.; Ali, A.; Kucuk, O. Nrf2/HO-1 signaling pathway may be the prime target for chemoprevention of cisplatin-induced nephrotoxicity by lycopene. Food Chem. Toxicol. 2010, 48, 2670–2674. [Google Scholar] [CrossRef]
- Lee, C.H.; Lin, Y.H.; Hsieh, J.L.; Chen, M.C.; Kuo, W.L. A polymer coating applied to Salmonella prevents the binding of Salmonella-specific antibodies. Int. J. Cancer 2013, 132, 717–725. [Google Scholar] [CrossRef]
- Chen, M.C.; Chang, W.W.; Kuan, Y.D.; Lin, S.T.; Hsu, H.C.; Lee, C.H. Resveratrol inhibits LPS-induced epithelial-mesenchymal transition in mouse melanoma model. Innate Immun. 2012, 18, 685–693. [Google Scholar] [CrossRef]
- ImageJ. Image Processing and Analysis in Java. Available online: http://rsbweb.nih.gov/ij/ (accessed on 6 September 2013).
- Mitazaki, S.; Hashimoto, M.; Matsuhashi, Y.; Honma, S.; Suto, M.; Kato, N.; Nakagawasai, O.; Tan-No, K.; Hiraiwa, K.; Yoshida, M.; et al. Interleukin-6 modulates oxidative stress produced during the development of cisplatin nephrotoxicity. Life Sci. 2013, 92, 694–700. [Google Scholar] [CrossRef]
- Herzog, C.; Seth, R.; Shah, S.V.; Kaushal, G.P. Role of meprin A in renal tubular epithelial cell injury. Kidney Int. 2007, 71, 1009–1018. [Google Scholar] [CrossRef]
- Lee, C.H.; Hsieh, J.L.; Wu, C.L.; Hsu, H.C.; Shiau, A.L. B cells are required for tumor-targeting Salmonella in host. Appl. Microbiol. Biotechnol. 2011, 92, 1251–1260. [Google Scholar]
- Lee, C.H.; Hsieh, J.L.; Wu, C.L.; Hsu, P.Y.; Shiau, A.L. T cell augments the antitumor activity of tumor-targeting Salmonella. Appl. Microbiol. Biotechnol. 2011, 90, 1381–1388. [Google Scholar] [CrossRef]
- Chang, W.W.; Lai, C.H.; Chen, M.C.; Liu, C.F.; Kuan, Y.D.; Lin, S.T.; Lee, C.H. Salmonella enhance chemosensitivity in tumor through connexin 43 upregulation. Int. J. Cancer 2013, 133, 1926–1935. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chang, W.-W.; Liu, J.-J.; Liu, C.-F.; Liu, W.-S.; Lim, Y.-P.; Cheng, Y.-J.; Lee, C.-H. An Extract of Rhodobacter sphaeroides Reduces Cisplatin-Induced Nephrotoxicity in Mice. Toxins 2013, 5, 2353-2365. https://doi.org/10.3390/toxins5122353
Chang W-W, Liu J-J, Liu C-F, Liu W-S, Lim Y-P, Cheng Y-J, Lee C-H. An Extract of Rhodobacter sphaeroides Reduces Cisplatin-Induced Nephrotoxicity in Mice. Toxins. 2013; 5(12):2353-2365. https://doi.org/10.3390/toxins5122353
Chicago/Turabian StyleChang, Wen-Wei, Jau-Jin Liu, Chi-Fan Liu, Wen-Sheng Liu, Yun-Ping Lim, Yu-Jung Cheng, and Che-Hsin Lee. 2013. "An Extract of Rhodobacter sphaeroides Reduces Cisplatin-Induced Nephrotoxicity in Mice" Toxins 5, no. 12: 2353-2365. https://doi.org/10.3390/toxins5122353