Dimethylarginine Dimethylaminohydrolase/Nitric Oxide Synthase Pathway in Liver and Kidney: Protective Effect of Cyanidin 3-O-β-D-Glucoside on Ochratoxin-A Toxicity




Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals
2.2. Animals and Treatment
2.3. Western Blot Analysis
2.4. NO2−/NO3− Quantification
2.5. DDAH Enzyme Activity Assay
2.6. Statistical Analyses
3. Results
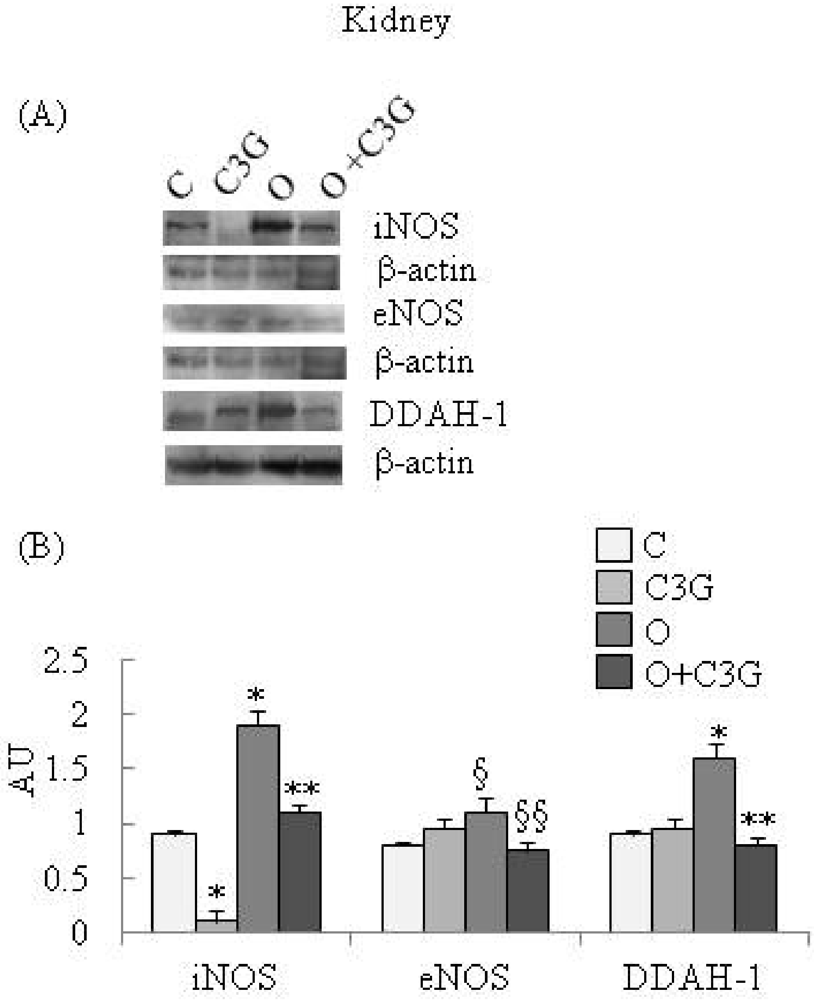

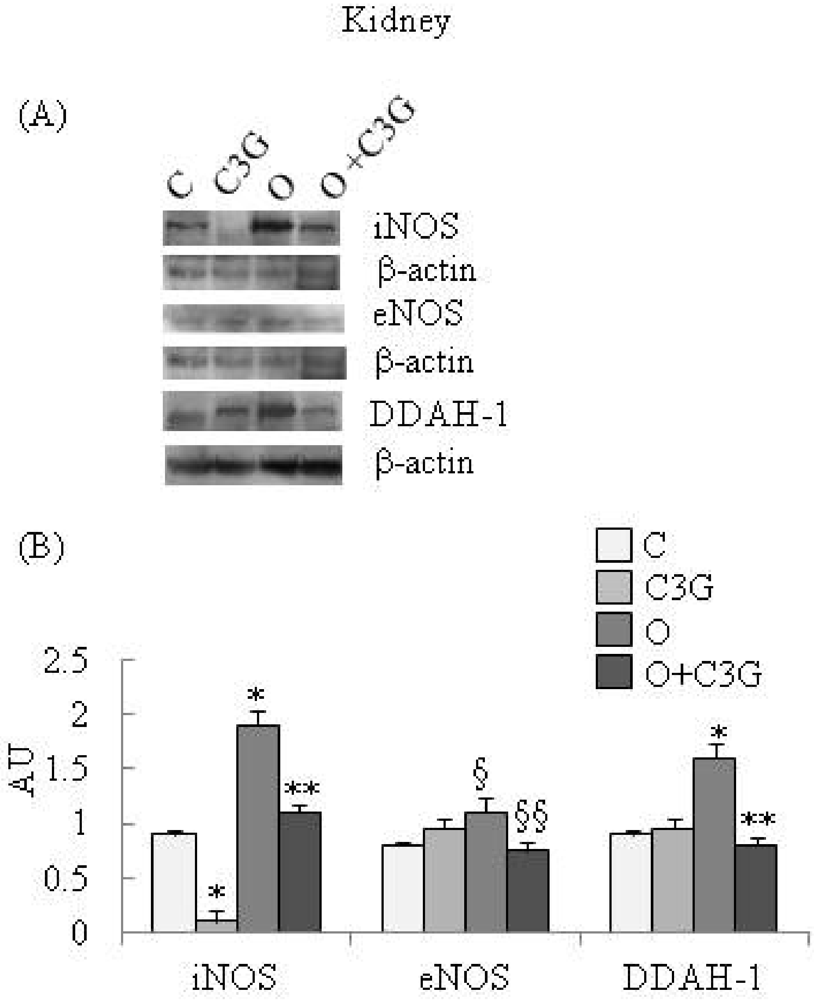{kind=link}
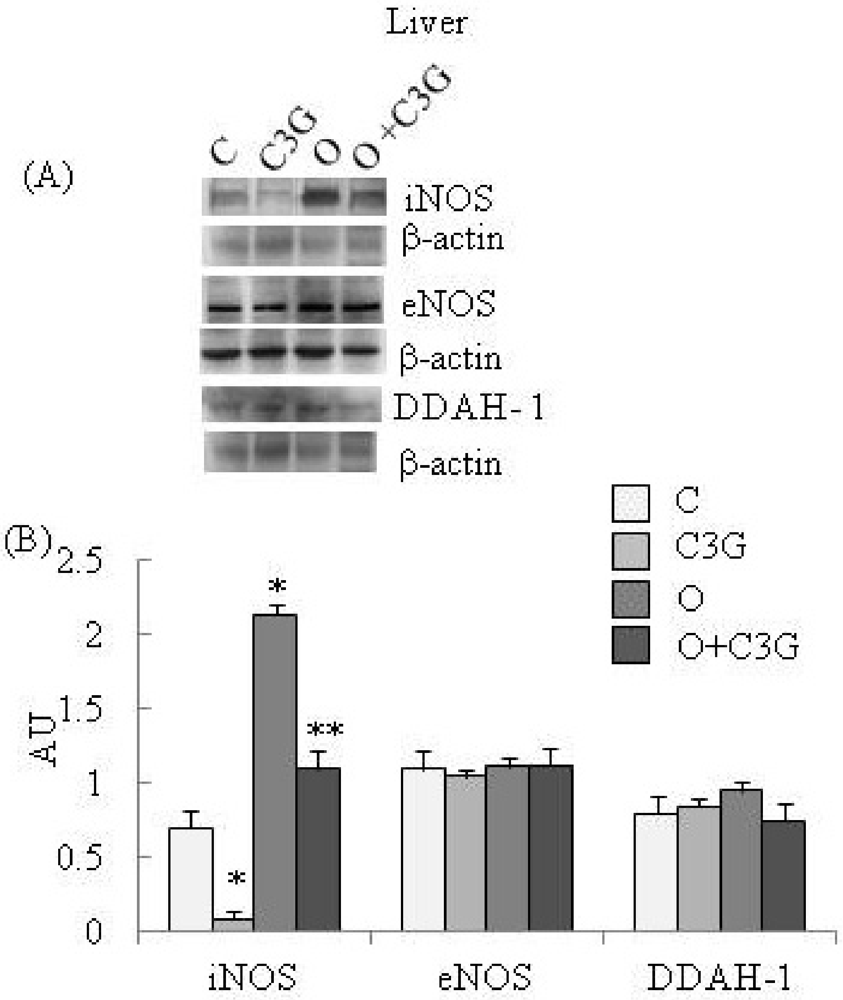
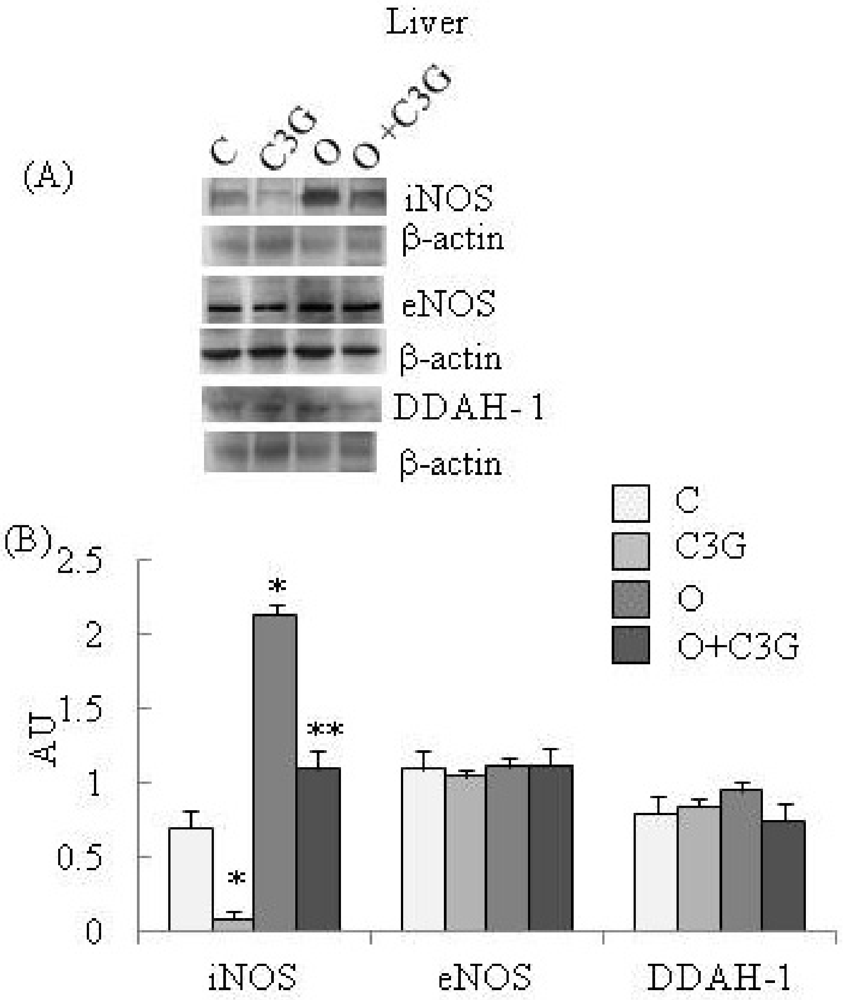{kind=link}
| Kikney | C | C3G | O | O + C3G |
|---|---|---|---|---|
| DDAH activity | 3.59 ± 0.03 | 3.58 ± 0.05 | 5.31 ± 0.04 * | 2.93 ± 0.01 ** |
| nitrite/nitrate levels | 5.15 ± 0.04 | 2.01 ± 0.02 * | 9.37 ± 0.09 * | 6.34 ± 0.08 ** |
| Liver | C | C3G | O | O + C3G |
|---|---|---|---|---|
| DDAH activity | 1.74 ± 0.01 | 1.58 ± 0.03 | 1.50 ± 0.01 | 1.41 ± 0.02 |
| nitrite/nitrate levels | 6.86 ± 0.09 | 3.14 ± 0.02 * | 11.42 ± 0.10 * | 7.10 ± 0.07 ** |
5. Discussion
References
- Clark, H.A.; Snedeker, S.M. Ochratoxin A: Its cancer risk and potential for exposure. J. Toxicol. Environ. Health B Crit. Rev. 2006, 9, 265–296. [Google Scholar]
- Jörgensen, K.; Rasmussen, G.; Thorup, I. Ochratoxin A in Danish cereals 1986–1992 and daily intake by the Danish population. Food Addit. Contam. 1996, 13, 95–104. [Google Scholar]
- Pitt, J.I. Penicillium viridicatum, Penicillium verrucosum, and production of ochratoxin A. Appl. Environ. Microbiol. 1987, 53, 266–269. [Google Scholar]
- Pfohl-Leszkowicz, A.; Manderville, R.A. Ochratoxin A: An overview on toxicity and carcinogenicity in animals and humans. Mol. Nutr. Food Res. 2007, 51, 61–99. [Google Scholar]
- Vrabcheva, T.; Petkova-Bocharova, T.; Grosso, F.; Nikolov, I.; Chernozemsky, I.N.; Castegnaro, M.; Dragacci, S. Analysis of ochratoxin A in foods consumed by in habitants from an area with balkan endemic nephropathy: A 1 month follow-up study. J. Agric. Food Chem. 2004, 52, 2404–2410. [Google Scholar]
- Cavin, C.; Delatour, T.; Marin-Kuan, M.; Holzhauser, D.; Higgins, L.; Bezencon, C.; Guignard, G.; Junod, S.; Richoz-Payot, J.; Gremaud, E.; Hayes, J.D.; Nestler, S.; Mantle, P.; Schilter, B. Reduction in antioxidant defenses may contribute to ochratoxin A toxicity and carcinogenicity. Toxicol. Sci. 2007, 96, 30–39. [Google Scholar]
- Di Giacomo, C.; Acquaviva, R.; Piva, A.; Sorrenti, V.; Vanella, L.; Piva, G.; Casadei, G.; La Fauci, L.; Ritieni, A.; Bognanno, M.; di Renzo, L.; Barcellona, M.L.; Morlacchini, M.; Galvano, F. Protective effect of cyanidin 3-O-β-D-glucoside on ochratoxin A-mediated damage in the rat. Brit. J. Nutr. 2007, 98, 937–943. [Google Scholar]
- Cavin, C.; Delatour, T.; Marin-Kuan, M.; Fenaille, F.; Holzhauser, D.; Guignard, G.; Bezencxon, C.; Piguet, D.; Parisod, V.; Richoz-Payot, J.; Schilter, B. Ochratoxin A-Mediated DNA and protein damage: Roles of nitrosative and oxidative stresses. Toxicol. Sci. 2009, 11, 84–94. [Google Scholar]
- Chinje, E.C.; Stratford, I.J. Role of nitric oxide in growth of solid tumours: a balancing act. Essays Biochem. 1997, 32, 61–72. [Google Scholar]
- Griffioen, A.W.; Molema, G. Angiogenesis: potentials or pharmacologic intervention in the treatment of cancer; cardiovascular diseas and crhonic inflammation. Pharm. Rev. 2000, 52, 237–268. [Google Scholar]
- Chen, G.G.; Lee, T.W.; Xu, H.; Yip, J.H.; Li, M.; Mok, T.S.; Yim, A.P. Increased inducible nitric oxide synthase in lung carcinoma of smokers. Cancer 2008, 112, 372–381. [Google Scholar]
- Lukes, P.; Pàcovà, H.; Kucera, T.; Vesely, D.; Martinek, J.; Asti, J. Expression of endothelial and inducible nitric oxide synthase and caspase-3 in tonsillar cancer; chronic tonsillitis and healthy tonsils. Folia Biol. 2008, 5, 141–145. [Google Scholar]
- Brennan, P.A.; Palacios-Callender, M.; Zaki, G.A.; Spedding, A.V.; Langdon, J.D. Type II nitric oxide synthase (NOS2) expression correlates with lynphnode status in oral squamous carcinoma. J. Oral Pathol. Med. 2001, 30, 129–134. [Google Scholar]
- Lim, K.H.; Ancrile, B.B.; Kashatus, D.F.; Counter, C.M. Tumour maintenance is mediated by eNOS. Nature 2008, 452, 646–649. [Google Scholar]
- Shang, Z.J.; Li, J.R. Expression of endothelial nitric oxide synthase and vascular endothelial growth factor in oral squamous cell carcinoma: its correlation with angiogenesis and disease progression. J. Oral Pathol. Med. 2005, 34, 134–139. [Google Scholar]
- Kostourou, V.; Robinson, S.P.; Cartwright, J.E.; Whitley, G.S. Dimethylarginine dimethylaminohydrolase I enhances tumour growth and angiogenesis. Br. J. Cancer 2002, 87, 673–680. [Google Scholar]
- Leiper, J.; Santa Maria, J.; Chubb, A.; MacAllister, R.J.; Charles, I.G.; Whithley, G.S.J.; Vallance, P. Identification of two human dimethylarginine dimethylaminohydrolases with distinct tissue distributions and homology to microbial arginine deiminases. Biochem. J. 1999, 343, 209–214. [Google Scholar]
- Nijveldt, R.J.; Teerlink, T.; van Guldener, C.; Prins, H.A.; van Lambalgen, A.A.; Stehouwer, C.D.; Rauwerda, J.A.; van Leeuwen, P.A. Handling of asymmetrical dimethylarginine and symmetrical dimethylarginine by the rat kidney under basal conditions and during endotoxaemia. Nephrol. Dial. Transplant. 2003, 18, 2542–2550. [Google Scholar]
- Tran, C.T.; Fox, M.F.; Vallance, P.; Leiper, J.M. Chromosomal localization; gene structure; and expression pattern of DDAH1: comparison with DDAH2 and implications for evolutionary origins. Genomics 2000, 68, 101–105. [Google Scholar]
- Russo, A.; la Fauci, L.; Acquaviva, R.; Campisi, A.; Raciti, G.; Scifo, C.; Renis, M.; Galvano, G.; Vanella, A.; Galvano, F. Ochratoxin A-induced DNA damage in human fibroblast: Protective effect of cyanidin 3-O-β-D-glucoside. J. Nutr. Biochem. 2005, 16, 31–37. [Google Scholar]
- Acquaviva, R.; Campisi, A.; Murabito, P.; Raciti, G.; Avola, R.; Mangiameli, S.; Musumeci, I.; Barcellona, M.L.; Vanella, A.; Li Volti, G. Propofol attenuates peroxynitrite-mediated DNA damage and apoptosis in cultured astrocytes: An alternative protective mechanism. Anesthesiology 2004, 101, 1363–1371. [Google Scholar]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.J.; Randall, R.G. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Knipp, M.; Vasak, M. A colorimetric 96-well microtirer plate assay for the determination of enzymatically formed citrulline. Anal. Biochem. 2000, 286, 257–264. [Google Scholar]
- Grusak, M.A. Phytochemicals in plants: genomics-assisted plant improvement for nutritional and health benefits. Curr. Opin. Biotechnol. 2002, 13, 508–511. [Google Scholar]
- Palombo, E.A. Phytochemicals from traditional medicinal plants used in the treatment of diarrhoea: modes of action and effects on intestinal function. Phytother. Res. 2006, 20, 717–724. [Google Scholar]
- Talalay, P.; Fahey, J.W. Phytochemicals from cruciferous plants protect against cancer by modulating carcinogen metabolism. J. Nutr. 2001, 131, 3027S–3033S. [Google Scholar]
- Noda, Y.; Kaneyuki, T.; Mori, A.; Packer, L. Antioxidant activities of pomegranate fruit extract and its anthocyanidins: delphinidin; cyanidin; and pelargonidin. J. Agric. Food Chem. 2002, 50, 166–171. [Google Scholar] [CrossRef]
- Seeram, N.P.; Momin, R.A.; Nair, M.G.; Bourquin, L.D. Cyclooxygenase inhibitory and antioxidant cyanidin glycosides in cherries and berries. Phytomedcine 2001, 8, 362–369. [Google Scholar]
- Amorini, A.M.; Lazzarino, G.; Galvano, F.; Fazzina, G.; Tavazzi, B.; Galvano, G. Cyanidin-3-O-beta-glucopyranoside protects myocardium and erythrocytes from oxygen radical-mediated damages. Free Radic. Res. 2003, 37, 453–460. [Google Scholar]
- Serraino, I.; Dugo, L.; Dugo, P.; Mondello, L.; Mazzon, E.; Dugo, G.; Caputi, A.P.; Cuzzocrea, S. Protective effects of cyanidin-3-O-glucoside from blackberry extract against peroxynitrite- induced endothelial dysfunction and vascular failure. Life Sci. 2003, 73, 1097–1114. [Google Scholar]
- Ding, M.; Feng, R.; Wang, S.Y.; Bowman, L.; Lu, Y.; Qian, Y.; Castranova, V.; Jiang, B.H.; Shi, X. Cyanidin-3-glucoside; a natural product derived from blackberry; exhibits chemopreventive and chemotherapeutic activity. J. Biol. Chem. 2006, 281, 17359–17668. [Google Scholar]
- World Health Organization (WHO). Evaluation of certain food additives and contaminants. In Proceedings of the Joint FAO/WHO Expert Committee on Food Additives (JECFA), Geneva, Switzerland, 19–28 June 2007; 947, pp. 169–180, WHO Food Additives Series 44.
- Arbillaga, L.; Azqueta, A.; van Delft, J.H.; Lopez de Cerain, A. In vitro gene expression data supporting a DNA non-reactive genotoxic mechanism for ochratoxin A. Toxicol. Appl. Pharmacol. 2007, 220, 216–224. [Google Scholar]
- Pfohl-Leszkowicz, A.; Manderville, R.A. An update on direct genotoxicity as a molecular mechanism of ochratoxin a carcinogenicity. Chem. Res. Toxicol. 2012, 25, 252–262. [Google Scholar]
- Fusi, E.; Rebucci, R.; Pecorini, C.; Campagnoli, A.; Pinotti, L.; Saccone, F.; Cheli, F.; Purup, S.; Sejrsen, K.; Baldi, A. Alpha-tocopherol counteracts the cytotoxicity induced by ochratoxin a in primary porcine fibroblasts. Toxins 2010, 2, 1265–1278. [Google Scholar]
- Manderville, R.A. A case for the genotoxicity of ochratoxin a by bioactivation and covalent DNA adduction. Chem. Res. Toxicol. 2005, 18, 1091–1097. [Google Scholar]
- European Food Safety Authority (EFSA). Opinion of the scientific panel on contaminants in the food chain on a request from the commission related to ochratoxin A in food. EFSA J. 2006, 365, 1–56.
- Petrik, J.; Zanic-Grubisic, T.; Barisic, K.; Pepeljnjak, S.; Radic, B.; Ferencic, Z.; Cepelak, I. Apoptosis and oxidative stress induced by ochratoxin-A in rat kidney. Arch. Toxicol. 2003, 77, 685–693. [Google Scholar]
- Gekle, M.; Sauvant, C.; Schwerdt, G. Ochratoxin A at nanomolar concentrations: A signal modulator in renal cells. Mol. Nutr. Food Res. 2005, 49, 118–130. [Google Scholar]
- Gagliano, N.; Doone, I.D.; Torri, C.; Migliori, M.; Grizzi, F.; Milzani, A.; Filippi, C.; Annoni, G.; Colombo, P.; Costa, F.; Ceva-Grimaldi, G.; Bertelli, A.A.; Giovannini, L.; Gioia, M. Early cytotoxic effects of ochratoxin A in rat liver: A morphological, biochemical and molecular study. Toxicology 2006, 225, 214–224. [Google Scholar]
- Vettorazzi, A.; de Trocóniz, I.F.; González-Peñas, E.; Arbillaga, L.; Corcuera, L.A.; Gil, A.G.; de Cerain, A.L. Kidney and liver distribution of ochratoxin A in male and female F344 rats. Food Chem. Toxicol. 2011, 49, 1935–1942. [Google Scholar]
- Ferrante, M.C.; Mattace Raso, G.; Bilancione, M.; Esposito, E.; Iacono, A.; Meli, R. Differential modification of inflammatory enzymes in J774A.1 macrophages by ochratoxin A alone or in combination with lipopolysaccharide. Toxicol. Lett. 2008, 181, 40–46. [Google Scholar] [CrossRef]
- Radi, R. Nitric oxide, oxidants, and protein tyrosine nitration. Proc. Natl. Acad. Sci. USA 2004, 101, 4003–4008. [Google Scholar]
- Wang, Q.; Xia, M.; Liu, C.; Guo, H.; Ye, Q.; Hu, Y.; Zhang, Y.; Hou, M.; Zhu, H.; Ma, J.; Ling, W. Cyanidin-3-O-β-glucoside inhibits iNOS and COX-2 expression by inducing liver X receptor alpha activation in THP-1 macrophages. Life Sci. 2008, 83, 176–184. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sorrenti, V.; Di Giacomo, C.; Acquaviva, R.; Bognanno, M.; Grilli, E.; D’Orazio, N.; Galvano, F. Dimethylarginine Dimethylaminohydrolase/Nitric Oxide Synthase Pathway in Liver and Kidney: Protective Effect of Cyanidin 3-O-β-D-Glucoside on Ochratoxin-A Toxicity. Toxins 2012, 4, 353-363. https://doi.org/10.3390/toxins4050353
Sorrenti V, Di Giacomo C, Acquaviva R, Bognanno M, Grilli E, D’Orazio N, Galvano F. Dimethylarginine Dimethylaminohydrolase/Nitric Oxide Synthase Pathway in Liver and Kidney: Protective Effect of Cyanidin 3-O-β-D-Glucoside on Ochratoxin-A Toxicity. Toxins. 2012; 4(5):353-363. https://doi.org/10.3390/toxins4050353
Chicago/Turabian StyleSorrenti, Valeria, Claudia Di Giacomo, Rosaria Acquaviva, Matteo Bognanno, Ester Grilli, Nicolantonio D’Orazio, and Fabio Galvano. 2012. "Dimethylarginine Dimethylaminohydrolase/Nitric Oxide Synthase Pathway in Liver and Kidney: Protective Effect of Cyanidin 3-O-β-D-Glucoside on Ochratoxin-A Toxicity" Toxins 4, no. 5: 353-363. https://doi.org/10.3390/toxins4050353