Using Cholinesterases and Immobilized Luminescent Photobacteria for the Express-Analysis of Mycotoxins and Estimating the Efficiency of Their Enzymatic Hydrolysis


Abstract
:1. Introduction
2. Results
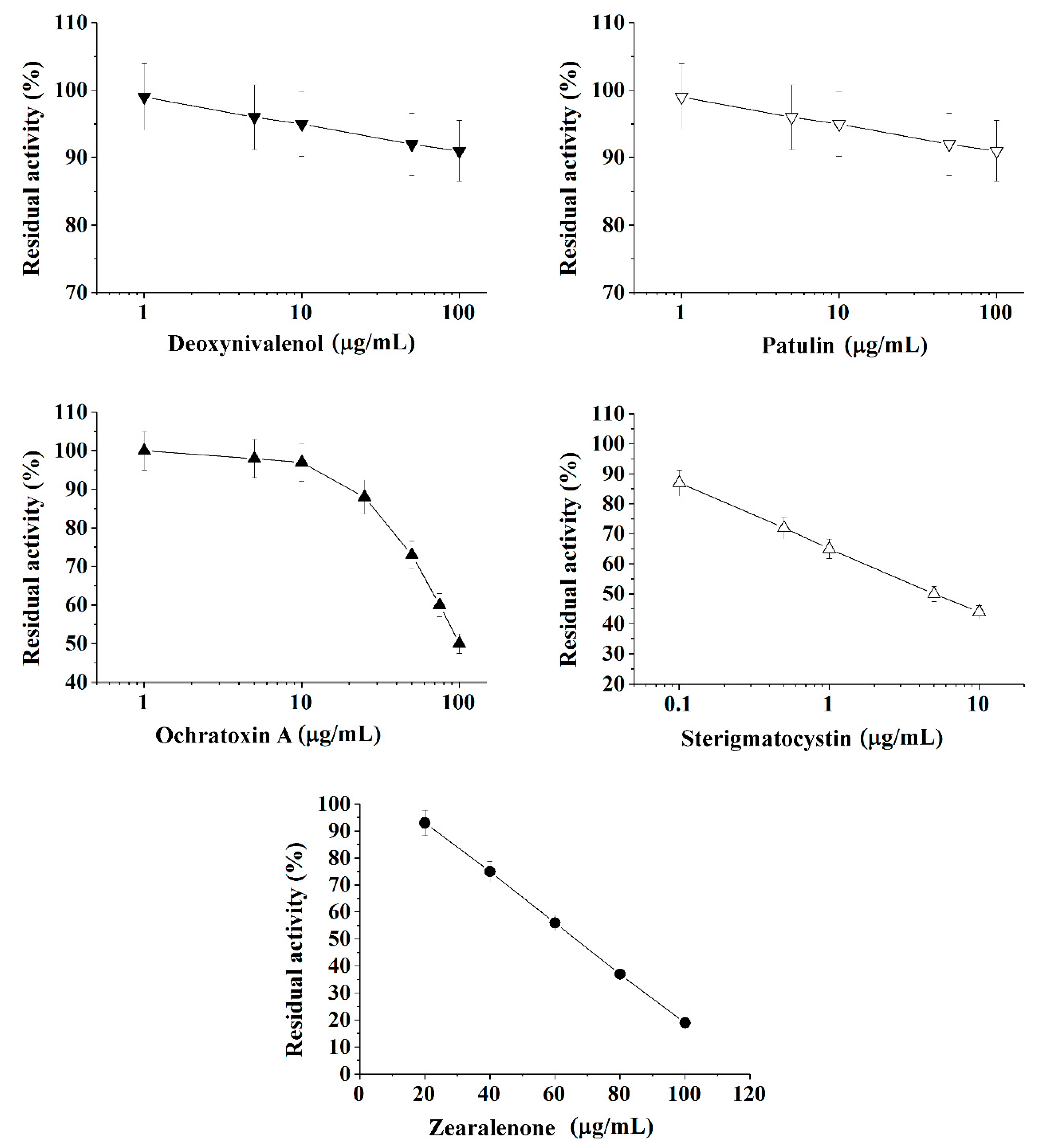
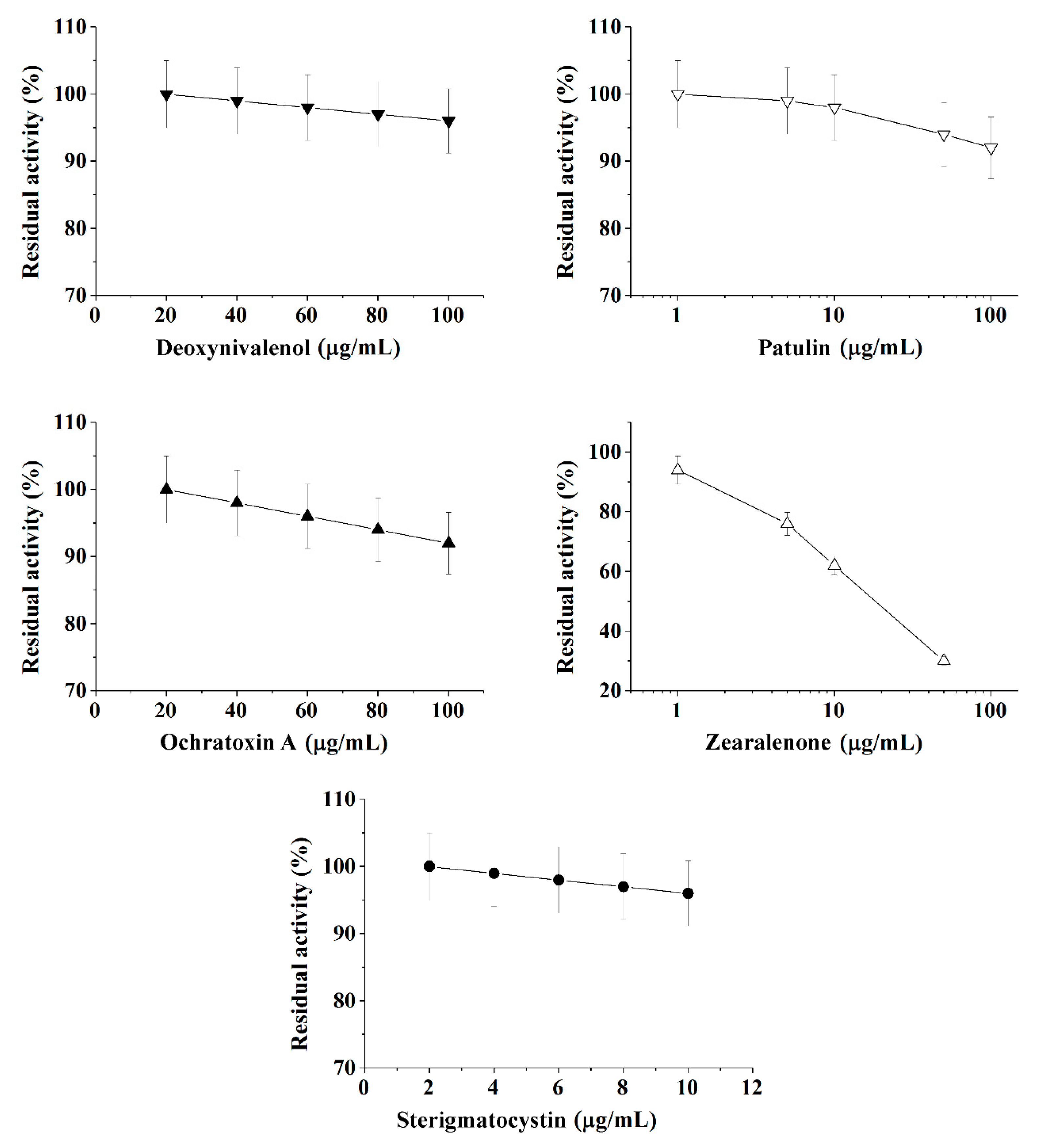
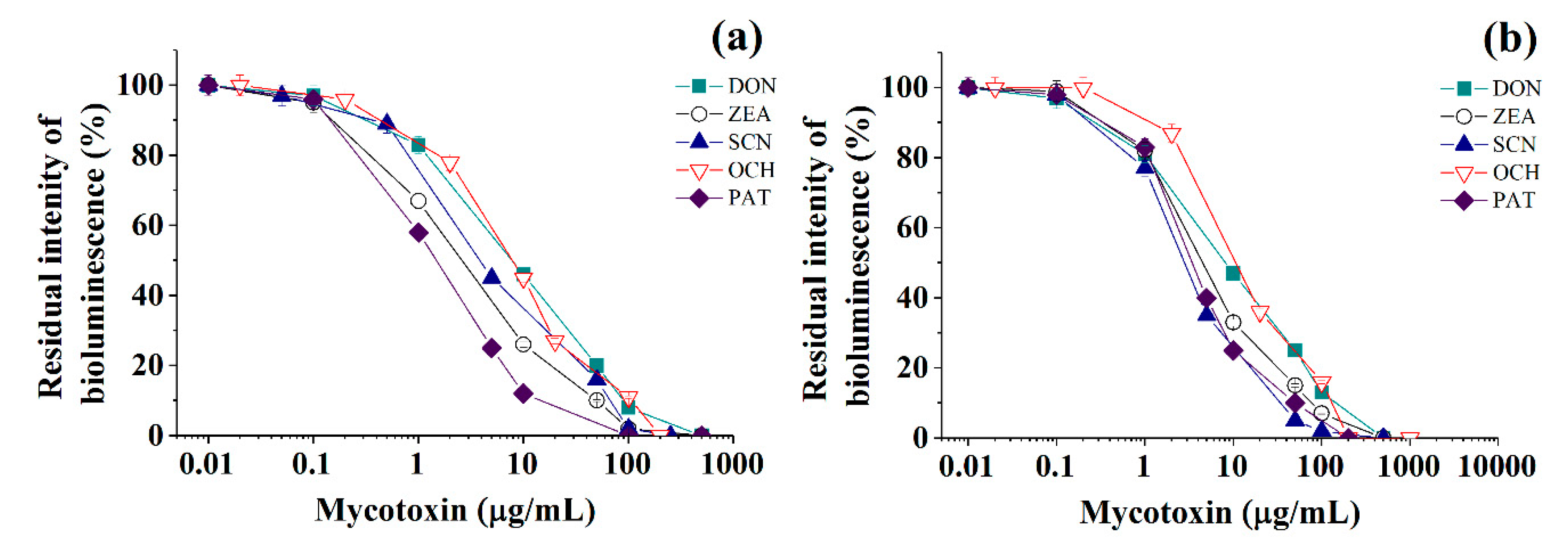
2.1. The Quantitative Express-Analysis of Mycotoxins in Liquid Media Involving Cholinesterases or Immobilized Bioluminescent Photobacterial Cells
2.2. Assessment of Toxicity of the Reaction Medium Obtained after Hydrolysis of Zearalenone by His6-OPH in the Media with Different pH
2.3. Zearalenone Biodegradation in Feed Grain Mixture under the Action of the Enzyme His6-OPH
3. Discussion
4. Materials and Methods
4.1. Chemicals and Strains
4.2. Growth Cells Conditions, Immobilization and Luminescence Measurements
4.3. Mycotoxins Analyses with Cholinesterase Enzymes (AChE and BChE)
4.4. Hydrolysis of Zearalenone in Medium with Different pH under the Action of the His6-OPH
4.5. Hydrolysis of Zearalenone in Feed Grain Mixture under the Action of the His6-OPH
4.6. Determination of Zearalenone by Enzyme-Linked Immunosorbent Assay (ELISA) Test Kit
4.7. Calculations
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kluczkovski, A.M. Fungal and mycotoxin problems in the nut industry. Curr. Opin. Food Sci. 2019, 29, 56–63. [Google Scholar] [CrossRef]
- Janik, E.; Niemcewicz, M.; Ceremuga, M.; Stela, M.; Saluk-Bijak, J.; Siadkowski, A.; Bijak, M. Molecular aspects of mycotoxins—A serious problem for human health. Int. J. Mol. Sci. 2020, 21, 8187. [Google Scholar] [CrossRef] [PubMed]
- Mamo, F.T.; Abate, B.A.; Tesfaye, K.; Nie, C.; Wang, G.; Liu, Y. Mycotoxins in Ethiopia: A review on prevalence, economic and health impacts. Toxins 2020, 12, 648. [Google Scholar] [CrossRef] [PubMed]
- Kunz, B.M.; Wanko, F.; Kemmlein, S.; Bahlmann, A.; Rohn, S.; Maul, R. Development of a rapid multi-mycotoxin LC-MS/MS stable isotope dilution analysis for grain legumes and its application on 66 market samples. Food Control 2020, 109, 106949. [Google Scholar] [CrossRef]
- Tran, V.N.; Viktorová, J.; Ruml, T. Mycotoxins: Biotransformation and bioavailability assessment using caco-2 cell monolayer. Toxins 2020, 12, 628. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in occurrence, importance, and mycotoxin control strategies: Prevention and detoxification in foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef]
- Li, P.; Su, R.; Yin, R.; Lai, D.; Wang, M.; Liu, Y.; Zhou, L. Detoxification of mycotoxins through biotransformation. Toxins 2020, 12, 121. [Google Scholar] [CrossRef] [Green Version]
- Adebiyi, J.A.; Kayitesi, E.; Adebo, O.A.; Changwa, R.; Njobeh, P.B. Food fermentation and mycotoxin detoxification: An African perspective. Food Control 2019, 106, 106731. [Google Scholar] [CrossRef]
- Narváez, A.; Rodríguez-Carrasco, Y.; Castaldo, L.; Izzo, L.; Ritieni, A. Ultra-high-performance liquid chromatography coupled with quadrupole Orbitrap high-resolution mass spectrometry for multi-residue analysis of mycotoxins and pesticides in botanical nutraceuticals. Toxins 2020, 12, 114. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Yu, J.; Li, F.; Li, J.; Shen, Z. A calibration curve implanted enzyme-linked immunosorbent assay for simultaneously quantitative determination of multiplex mycotoxins in cereal samples, soybean and peanut. Toxins 2020, 12, 718. [Google Scholar] [CrossRef]
- Lucci, P.; David, S.; Conchione, C.; Milani, A.; Moret, S.; Pacetti, D.; Conte, L. Molecularly imprinted polymer as selective sorbent for the extraction of zearalenone in edible vegetable oils. Foods 2020, 9, 1439. [Google Scholar] [CrossRef] [PubMed]
- Shcherbakova, L.; Rozhkova, A.; Osipov, D.; Zorov, I.; Mikityuk, O.; Statsyuk, N.; Sinitsyna, O.; Dzhavakhiya, V.; Sinitsyn, A. Effective zearalenone degradation in model solutions and infected wheat grain using a novel heterologous lactonohydrolase secreted by recombinant Penicillium canescens. Toxins 2020, 12, 475. [Google Scholar] [CrossRef] [PubMed]
- Lyagin, I.; Efremenko, E. Enzymes for detoxification of various mycotoxins: Origins and mechanisms of catalytic action. Molecules 2019, 24, 2362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Taheur, F.; Kouidhi, B.; Al Qurashi, Y.M.A.; Ben Salah-Abbès, J.; Chaieb, K. Review: Biotechnology of mycotoxins detoxification using microorganisms and enzymes. Toxicon 2019, 160, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Valera, E.; García-Febrero, R.; Elliott, C.T.; Sánchez-Baeza, F.; Marco, M.P. Electrochemical nanoprobe-based immunosensor for deoxynivalenol mycotoxin residues analysis in wheat samples. Anal. Bioanal. Chem. 2019, 411, 1915–1926. [Google Scholar] [CrossRef]
- Stadler, D.; Berthiller, F.; Suman, M.; Schuhmacher, R.; Krska, R. Novel analytical methods to study the fate of mycotoxins during thermal food processing. Anal. Bioanal. Chem. 2020, 412, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Senko, O.; Stepanov, N.; Maslova, O.; Akhundov, R.; Ismailov, A.; Efremenko, E. Immobilized luminescent bacteria for the detection of mycotoxins under discrete and flow-through conditions. Biosensors 2019, 9, 63. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.; Tan, J.; Xu, Z.; Huang, J.; Tian, Y.; Chen, B.; Wu, Y.; Tong, Y.; Zhu, Y. Computational design of enhanced detoxification activity of a zearalenone lactonase from Clonostachysrosea in acidic medium. RSC Adv. 2019, 9, 31284–31295. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Ye, J.; Zhang, Y.; Shuai, C.; Yuan, Q. Computer-aid molecular docking technology in cereal mycotoxin analysis. J. Food Sci. Eng. 2019, 9, 244–253. [Google Scholar] [CrossRef]
- Efremenko, E.N.; Maslova, O.V.; Kholstov, A.V.; Senko, O.V.; Ismailov, A.D. Biosensitive element in the form of immobilized luminescent photobacteria for detecting ecotoxicants in aqueous flow--through systems. Luminescence 2016, 31, 1283–1289. [Google Scholar] [CrossRef]
- Varfolomeev, S.D.; Efremenko, E.N. Organophosphorus Neurotoxins: Monograph; RIOR Publisher: Moscow, Russia, 2020; p. 380. [Google Scholar] [CrossRef]
- Göbel, R.; Lusky, K. Simultaneous determination of aflatoxins, ochratoxin A, and zearalenone in grains by new immunoaffinity column/liquid chromatography. J. AOAC Int. 2004, 87, 411–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, B.; Han, Z.; Cai, Z.; Wu, Y.; Ren, Y. Simultaneous determination of aflatoxins B1, B2, G1, G2, M1 and M2 in peanuts and their derivative products by ultra-high-performance liquid chromatography–tandem mass spectrometry. Anal. Chim. Acta 2010, 662, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Efremenko, E.; Votchitseva, Y.; Plieva, F.; Galaev, I.; Mattiasson, B. Purification of His6-organophosphate hydrolase using monolithic supermacroporous polyacrylamide cryogels developed for immobilized metal affinity chromatography. Appl. Microbiol. Biotechnol. 2006, 70, 558–563. [Google Scholar] [CrossRef]
- Votchitseva, Y.A.; Efremenko, E.N.; Aliev, T.K.; Varfolomeyev, S.D. Properties of hexahistidine-tagged organophosphate hydrolase. Biochemistry 2006, 71, 167–172. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Mycotoxin | Coefficients of the Linearization Equation | R2 | Working Range, µg/mL | Limit of Quantification (LOQ), µg/mL | Limit of Detection (LOD), µg/mL | |
|---|---|---|---|---|---|---|
| a | b | |||||
| AChE | ||||||
| Deoxynivalenol 1 | 97.9 ± 0.1 | 4.0 ± 0.1 | 0.998 | ≥1698 | 1698 | 563 |
| Ochratoxin A 1 | 64.9 ± 5.4 | 180.9 ± 9.9 | 0.979 | 30−354 | 30 | 10 |
| Patulin 1 | 98.9 ± 0.1 | 4.0 ± 0.1 | 0.998 | ≥2951 | 2951 | 984 |
| Sterigmatocystin 1 | 65.3 ± 0.2 | 21.5 ± 6.0 | 0.999 | 0.12−219 | 0.12 | 0.04 |
| Zearalenone 2 | 111.5 ± 0.9 | 0.9 ± 0.01 | 0.999 | 29−103 | 29 | 10 |
| BChE | ||||||
| Deoxynivalenol 2 | 101.0 ± 0.1 | 0.05 ± 0.001 | 0.999 | 320−1720 | 320 | 105 |
| Ochratoxin A 2 | 102.0 ± 0.1 | 0.1 ± 0.01 | 0.999 | 170−870 | 170 | 56 |
| Patulin 1 | 104.0 ± 0.4 | 6.0 ± 0.2 | 0.997 | ≥1548 | 1548 | 511 |
| Sterigmatocystin 1 | 101.0 ± 0.1 | 0.5 ± 0.01 | 0.999 | 32−172 | 32 | 11 |
| Zearalenone 1 | 108.0 ± 0.2 | 45.9 ± 0.1 | 0.999 | 3−107 | 3 | 1 |
| Photobacterium sp. 9.2 cells | ||||||
| Deoxynivalenol 1 | 84.1 ± 1.2 | 38.0 ± 0.6 | 0.999 | 1−66 | 1 | 0.3 |
| Ochratoxin A 1 | 73.6 ± 10.2 | 31.5 ± 5.5 | 0.923 | 0.4−72 | 0.4 | 0.13 |
| Patulin 1 | 57.1 ± 1.3 | 45.2 ± 1.4 | 0.999 | 0.3−8 | 0.3 | 0.1 |
| Sterigmatocystin 1 | 71.8 ± 6.3 | 33.0 ± 3.9 | 0.981 | 0.4−52 | 0.4 | 0.13 |
| Zearalenone 1 | 57.6 ± 9.0 | 28.2 ± 5.6 | 0.934 | 0.2−32 | 0.2 | 0.07 |
| Photobacterium sp. 17 cells | ||||||
| Deoxynivalenol 1 | 82.1 ± 2.6 | 34.4 ± 1.4 | 0.995 | 0.8−89 | 0.8 | 0.27 |
| Ochratoxin A 1 | 87.0 ± 13.3 | 35.8 ± 7.1 | 0.925 | 1.1−102 | 1.1 | 0.37 |
| Patulin 1 | 60.2 ± 10.1 | 29.9 ± 6.4 | 0.912 | 0.2−32 | 0.4 | 0.07 |
| Sterigmatocystin 1 | 63.8 ± 10.2 | 34.7 ± 6.1 | 0.941 | 0.3−25 | 0.4 | 0.1 |
| Zearalenone 1 | 70.5 ± 11.6 | 33.0 ± 7.4 | 0.905 | 0.4−48 | 0.4 | 0.13 |
| Analytical Method * | Zearalenone, μg/mL | * DH, % | Reference | |
|---|---|---|---|---|
| pH 7.4 | pH 8.5 | pH 8.5 | ||
| BChE | 3.8 ± 0.1 | - | - | This work |
| Photobacterium sp. 9.2 | 4.3 ± 0.1 | 0.67 ± 0.03 | 98.8 ± 0.9 | This work |
| Photobacterium sp. 17 | 4.5 ± 0.1 | 0.63 ± 0.03 | 99.0 ± 0.9 | This work |
| MaxSignal® Zearalenone ELISA Test Kit | 3.8 ± 0.1 | 0.58 ± 0.02 | 99.1 ± 0.9 | This work |
| 3.9 ± 0.2 | - | - | [17] | |
| Photobacterium phosphoreum B-1717 | 4.1 ± 0.2 | - | - | [17] |
| Analytical Method | Zearalenone, mg/kg Feed | * R, % | ** D, % | |
|---|---|---|---|---|
| NE | ED | |||
| BChE | 8.5 ± 0.3 | *** | 85 ± 4 | *** |
| Photobacterium sp. 9.2 | 8.1 ± 0.3 | 0.82 ± 0.04 | 81 ± 4 | 89.9 ± 4.1 |
| Photobacterium sp. 17 | 8.2 ± 0.3 | 0.85 ± 0.05 | 82 ± 4 | 89.6 ± 4.2 |
| MaxSignal® Zearalenone ELISA Test Kit | 7.9 ± 0.3 | 0.77 ± 0.03 | 79 ± 4 | 90.3 ± 4.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Efremenko, E.; Maslova, O.; Stepanov, N.; Ismailov, A. Using Cholinesterases and Immobilized Luminescent Photobacteria for the Express-Analysis of Mycotoxins and Estimating the Efficiency of Their Enzymatic Hydrolysis. Toxins 2021, 13, 34. https://doi.org/10.3390/toxins13010034
Efremenko E, Maslova O, Stepanov N, Ismailov A. Using Cholinesterases and Immobilized Luminescent Photobacteria for the Express-Analysis of Mycotoxins and Estimating the Efficiency of Their Enzymatic Hydrolysis. Toxins. 2021; 13(1):34. https://doi.org/10.3390/toxins13010034
Chicago/Turabian StyleEfremenko, Elena, Olga Maslova, Nikolay Stepanov, and Anvar Ismailov. 2021. "Using Cholinesterases and Immobilized Luminescent Photobacteria for the Express-Analysis of Mycotoxins and Estimating the Efficiency of Their Enzymatic Hydrolysis" Toxins 13, no. 1: 34. https://doi.org/10.3390/toxins13010034