1. Introduction
It is widely recognized that the response to lifestyle interventions, such as changes in dietary habits and physical activity, can be heterogeneous and largely genetically determined [
1]. Functional variants of genes involved in appetite regulation, energy metabolism and fat storage are considered candidates to confer susceptibility to weight loss, and their identification may be helpful in the prevention of future health problems.
Among several genetic variants investigated in relation to the effectiveness of weight loss therapy in obese subjects are Q223R (or Gln223Arg, rs1137101) and K656N (or Lys656Asn, rs8129183) polymorphisms in the leptin receptor gene (
LEPR) [
2,
3,
4]. Leptin receptor, which is a member of the cytokine receptor family, has several isoforms of different locations and functions [
5]. For example, the weight-regulating effects of leptin are mediated by binding to and activation of the specific receptor (long isoform, OB-R
L) in the brain, while a soluble form of this receptor (one of its short isoforms, sOB-R) modulates leptin levels by binding free leptin in the circulation and preventing the hormone from degradation and clearance [
6].
The Q223R A > G (A668G) and K656N G > C (G1968C) polymorphisms in
LEPR result in nonconservative changes: glutamine to arginine at codon 223 and lysine to asparagine at codon 656, respectively [
7,
8]. These amino acid substitutions result in changes of the electric charge from neutral to positive for Q223→R and from positive to neutral for K656→N and, therefore, are likely to have functional consequences resulting in changed leptin signaling and, in the broader perspective, an affected response to energy restriction [
7]. However, there are only a few studies that have evaluated the impact of these polymorphisms on changes in metabolic parameters during weight loss.
The diverse response of leptin levels and body mass changes depending on polymorphic variants of
LEP and
LEPR has been investigated mainly in obese adults [
2,
3,
9,
10,
11,
12,
13]. There are also studies suggesting a gene-gene interaction between the
LEP and
LEPR variants in a genetic susceptibility to the development of obesity [
14]. A cumulative effect of
LEPR polymorphisms with other obesity susceptibility loci, including variants in the adiponectin locus (
ADIPOQ), on weight loss was also observed in Spanish obese adolescents after a multidisciplinary intervention [
15]. Case-control studies on the association between polymorphisms in genes encoding adipokines with childhood obesity have been conducted [
16,
17,
18,
19,
20], but to our knowledge, there is no data concerning the influence of these polymorphisms on weight loss and adipokine concentrations after multidisciplinary intervention. Therefore, the aim of our study was to investigate the effect of selected functional single nucleotide polymorphisms (SNPs) in
LEPR,
LEP, and
ADIPOQ loci on metabolic response and weight loss after a 3-month lifestyle intervention in prepubertal obese children.
2. Methods
2.1. Subjects
One hundred obese prepubertal Caucasian children aged 5–10 years were recruited between 2010 and 2012 from a group of consecutive patients seeking dietary counseling in the Department of Nutrition at the Institute of Mother and Child in Warsaw. Obesity was classified as standard deviation score-body mass index (SDS-BMI) >2. Exclusion criteria were: (a) presence of endocrine disorders or genetic syndromes, including syndromic obesity; (b) other chronic medical conditions; (c) intake of medications that could affect growth, pubertal development, nutritional or dietary status. Pubertal stage was determined according to the Tanner scale and subjects who showed pubertal development were excluded. Written informed consent was obtained from the parents of all the examined children. The study was performed in accordance with the Helsinki Declaration for Human Research and the study protocol was approved (No. 25/2006) by the Ethics Committee of the Institute of Mother and Child in Warsaw, Poland.
2.2. Dietary Intervention
To evaluate changes in clinical, anthropometric and biochemical parameters in response to lifestyle changes, the obese prepubertal patients underwent a 3-month intervention program consisting of dietary and physical activity modifications and behavioral therapy, including individual psychological care for the child and his/her family. The dietary guidelines, recommending a low-energy diet based on a balanced distribution of carbohydrates, proteins and lipids, for children and their parents, were described in a previous study [
21]. The recommended daily energy intake was 1200–1400 kcal/day. The diet was composed of 20% protein, 30% fat and of 50% carbohydrates. Patients had 3–5 meals every day. Study participants did not receive vitamins or mineral supplements. Patients received instructions concerning physical activity, details of which were also previously described [
22]. Children were advised to reduce sedentary activities, including watching television and playing computer games to less than two hours a day. Two weeks before the visit in the Department of Nutrition, study participants completed a 10–14 days food diary using a standard questionnaire. Next, randomly selected 3-day records: two consecutive weekdays and one weekend day, before (visit T0) and after 3 months of therapy (visit T3) were analyzed. The subjects and their parents were asked to record the type and amount of food and beverages consumed for the selected days. We evaluated the average daily energy intake and the percentage of energy intake from protein, fat and carbohydrates in the children’s diets. Average daily food rations and their nutritional value were calculated using nutritional analysis software (Dietetyk
®, National Food and Nutrition Institute, version 2.0, Warsaw, Poland) [
23]. The age- and sex-specific dietary reference intake (DRI) percentage was calculated using reference values of daily energy intake for children and adolescents according to Jarosz
et al. [
24].
2.3. Anthropometric Parameters
Physical examinations, including body height and weight measurements, were performed before (visit T0) and after 3 months of therapy (visit T3). Body height was measured using a standing stadiometer and recorded with a precision of 1 mm. Body weight was assessed unclothed, to the nearest 0.1 kg, with a calibrated balance scale. BMI was calculated as body weight divided by height squared (kg/m
2). The BMI of each individual was converted to a standard SDS-BMI for the child’s age and sex using Polish reference tables [
25]. The data of this reference population were derived from a study concerning the physical development of Warsaw children and adolescents aged 1–18 years, which was conducted in the years 1996–1999 by the Department of Development of Children and Adolescents at the Institute of Mother and Child. Skinfolds were measured with a Holtain skinfold caliper (Holtain Ltd., Wales, UK). Fat mass percentage was calculated according to Lohman’s formula [
26].
2.4. Biochemical Measurements
Biochemical parameters were determined twice: before (visit T0) and after 3 months of therapy (visit T3). Venous blood samples were collected between 8:00 and 10:00 a.m. after an overnight fast, and centrifuged at 1000× g for 10 min at 4 °C. Serum specimens were stored at −70 °C prior to assay. Commercially available ELISA kits (DRG Diagnostics, Marburg, Germany) were used to determine leptin and sOB-R concentrations. Inter-assay variations (CV%) were 5.3% and 3.3% for total leptin and sOB-R, respectively. Serum levels of total adiponectin and high molecular weight (HMW) adiponectin were determined using ELISA kit (ALPCO Diagnostics, Salem, NH, USA). Adiponectin multimers were selectively measured after sample pretreatment with two proteases that specifically digested the trimeric forms or both the hexameric and trimeric forms. In this assay, total adiponectin and HMW adiponectin levels were determined directly. Inter-assay variations were 5.0% and 5.7% for total adiponectin and HMW adiponectin, respectively. Glucose, total cholesterol, LDL and HDL-cholesterol and triglycerides were measured by standard methods (Roche Diagnostics, Basel, Switzerland). To reduce interassay variance, samples obtained before and after therapy were analyzed in one assay.
2.5. DNA Isolation and Genetic Analysis
DNA was isolated from 4 mL of venous blood samples by a salting-out procedure [
27]. Genotyping of the -2548G > A
LEP (rs7799039), Q223R (
A668
G, rs1137101) and K656N (
G1968
C, rs8129183)
LEPR, as well as -11377C > G (rs266729) and -11426A > G (rs16861194)
ADIPOQ polymorphisms was performed by PCR amplification followed by digestion with an appropriate restriction enzyme [
20]. The obtained restriction fragments were visualized on 2%–3% agarose gels. For quality control of genetic testing: (i) amplification blank controls consisting of only amplification reagents without DNA were performed during each PCR reaction; (ii) randomly selected DNA samples were analyzed by sequencing; (iii) control samples with genotypes identified by sequencing were included into each PCR-RFLP reaction.
2.6. Statistical Analysis
The results are presented as means ± standard deviation (SD) for normally distributed data or medians and interquartile range (25th–75th percentiles) for non-normally distributed variables. The Kolmogorov-Smirnov test and graphical inspections of data were used to evaluate the distribution for normality.
Due to the fact that an increase in height between visit T0 and T3 could be partially responsible for a decrease in SDS-BMI in that time period, weight loss during intervention was defined as SDS-BMI change (Δ) ≤ −0.5. Differences in anthropometric characteristics and biochemical parameters between children who lost weight and those who have not were assessed using the Student t test for normally distributed data and the nonparametric Mann-Whitney test for non-normally distributed data. Paired Student t test or nonparametric Wilcoxon test were used to compare the parameters mentioned above before and after intervention in the whole group and in subgroups of children who lost weight and those that did not.
The Hardy-Weinberg equilibrium analysis was performed using a chi-square test with one degree of freedom. Additive, dominant, recessive and allelic models were tested for every polymorphism for associations with weight loss. Under each model, the odds ratios (ORs) with their 95% confidence intervals (CI) were calculated, and the chi-square test, Fisher exact test and Cochran-Armitage test for trend were used, as appropriate. The procedure described by Benjamini and Hochberg was used to correct
p-values for multiple testing [
28]. Linkage disequilibrium (LD) was analyzed using the pairwise LD measure D’, and haplotype blocks were constructed from population genotype data using Haploview software [
29], with the default algorithm for generating haplotype blocks based on methods established by Gabriel
et al. [
30]. A D’ value of 1 indicates complete LD between the two markers; a D’ value greater than 0.8—strong LD; 0.2–0.8—incomplete LD; whereas a D’ of less than 0.2—negligible LD. Multiple logistic regression analysis was conducted to assess the combined impact of two
LEPR polymorphisms on weight loss during therapy.
Changes in SDS-BMI, body composition and biochemical parameters were expressed as delta variables, calculated by subtracting values at baseline (T0) from values measured after 3 months of therapy (T3). Differences in anthropometric characteristics and biochemical parameters between different genotypes were assessed using the Student t test for normally distributed data and the nonparametric Mann-Whitney test for non-normally distributed data. To study associations of the combined genotypes of the LEPR polymorphisms Q223R and K656N with changes in soluble leptin receptor concentration, the multivariate linear regression model adjusted for age and sex was used. Genetic variants of the studied polymorphisms were treated as a continuous variable and put in order from the exclusive presence of at least one copy of Q223R minor allele to the exclusive presence of at least one copy of K656N minor allele. Test for linear trend was applied.
The correlation coefficients between changes in body composition and changes in SDS-BMI due to the intervention in children with combined genotypes of the LEPR Q223R and K656N polymorphisms were calculated using Pearson correlation.
Statistical significance was set at 0.05. Statistical analysis was performed using SPSS v.18.0 software (SPSS Inc., Chicago, IL, USA) and StatXact-3 software, version 3.1 (Cytel Software Corporation, Cambridge, MA, USA).
4. Discussion
Our study shows that both Q223R and K656N polymorphisms in the leptin receptor gene may modulate the effect of lifestyle intervention in obese individuals during the prepubertal period, whereas other investigated SNPs in genes encoding leptin and adiponectin were not related to a reduction in body mass. We also demonstrated that combinations of genotypes of the investigated polymorphisms in LEPR may determine predisposition to therapy-induced changes in body composition and soluble leptin receptor concentration in prepubertal children.
Based on the finding that obese subjects have higher serum levels of leptin and lower levels of its soluble receptor, it has been suggested that human obesity is associated with leptin resistance [
31]. Since both the Q223R and the K656N polymorphisms are located within the region encoding the extracellular domain of the leptin receptor, the substitution of one amino acid by another can affect the functional characteristics of the receptor leading to its modified signaling capacity and resulting in various levels of leptin in circulation [
1].
In our previous work, consistently with the results of other authors, we did not observe an association between these two
LEPR SNPs and obesity in children and adolescents [
20,
32,
33]. However, none of these studies took into account the combined effect of the Q223R and K656N variants on susceptibility to obesity and the level of serum adipokines. Such an interaction was described by Lu
et al. [
14] who found that a combination of a polymorphism in the
LEP 3′ flanking region with two polymorphisms in
LEPR (K109R and K656N) was associated with obesity in a Chinese population, even though none of them individually were associated with body weight or BMI.
Lifestyle interventions, including modification of dietary habits and increased physical activity, are recommended as an effective therapy for weight reduction as well as for an improvement of biochemical parameters in obese children and adolescents [
34]. Like other authors and in accordance with our previous findings, after therapy-induced weight loss we observed a significant decrease in leptin concentration with a parallel increase in soluble leptin receptor and adiponectin levels [
35,
36,
37]. However, about 30% of the studied group did not have a change in body mass or body composition despite having energy intake and percentage of DRI before and after therapy similar to the children with weight loss. This finding implies that response to lifestyle interventions may be genetically determined.
Given the important role of leptin in the regulation of energy metabolism, we suggest that genetic variants of
LEPR may modulate physiological responses to lifestyle interventions and its influence on health in prepubertal obese children. When analyzed separately, the Q223R and K656N polymorphisms were related to different parameters in children after weight loss therapy: the Q223R
LEPR SNP was associated with changes in BMI and fat mass percentage, while the K656N with changes in soluble leptin receptor serum concentrations. In contrast, we did not find any differences in serum leptin levels in relation to the studied
LEPR polymorphisms in obese children before and after intervention. The results of previous studies on the relationship between
LEPR variants and leptin concentrations in the circulation are ambiguous. Mars
et al. [
10] also demonstrated no effect of the K109R, Q223R and K656N SNPs on an acute decline in leptin levels after only 4 days of energy restriction in men. However, Luis
et al. [
38] observed in Spanish adults a significant decrease in weight, fat mass as well as leptin concentrations in wild-type homozygotes of K656N after a 3months lifestyle intervention, whereas carriers of the minor allele had no decrease in fat mass or leptin levels after this period. In addition, carriers of the minor K656N allele did not respond with a decrease in insulin level or HOMA-IR after weight loss induced by a hypocaloric diet [
3]. Similarly to other authors studying the K656N polymorphism in adults, we also suggest that children with the minor allele, which is associated with a lack of soluble leptin receptor changes after therapy, may be more resistant to multidisciplinary intervention than wild-type homozygotes [
4].
Repasy
et al. [
19] when analyzing the relationship between the Q223R polymorphism and indicators of energy expenditure in obese children (aged 13 ± 2.7 years), also found no association between leptin concentrations and different genotypes of this SNP. However, the wild-type homozygotes compared with other genotypes showed a significantly lower post-absorptive and postprandial respiratory quotient, which could indicate a lower binding capacity of leptin to the soluble form of the receptor in plasma [
19]. In our study, we also did not observe any differences in leptin levels between the carriers of the Q223R genotypes before and after therapy in contrast with an adult population in whom differences in leptin concentrations had been observed [
39]. These differences could be explained by the younger age of Repasy
et al.’s [
19] study population and our patients, in whom the full effect of Q223R polymorphism has not yet been developed. Nevertheless, our study points at the association between this polymorphism and weight loss in children after lifestyle intervention. Carriers of the Q223R wild-type variant lost significantly less weight and less fat mass after therapy than carriers of the minor allele; therefore, we suggest that wild-type variant carriers may be more resistant to weight loss therapy than carriers of the minor variant. Other authors observed that in adults the wild-type variant of Q223R correlated not only with leptin concentrations but also with higher BMI and body fat, impaired glucose metabolism and dyslipidemia, as well as lower respiratory quotient during low-intensity exercise [
40,
41]. Therefore, it is suggested that this variant of
LEPR polymorphism may carry a risk of insulin resistance and liver steatosis in later life. Other genetic variant at the
LEPR locus (rs3790433) was also associated with increased risk of metabolic syndrome and insulin resistance in adults with metabolic syndrome [
12]. In addition, authors found that low plasma level of (
n-3) and high level of (
n-6) polyunsaturated fatty acids exacerbated impact of this polymorphism on the risk of hiperinsulinemia and insulin resistance. Many studies have shown that fatty acids influence leptin expression and concentration, but the influence of genetic variations on this is still unknown [
42,
43]. However, the knowledge of gene-nutrient interactions might be useful in developing personalized dietary recommendations in obese adults as well as obese children.
To our knowledge, the combined influence of both
LEPR polymorphisms on soluble leptin receptor levels and weight loss in obese subjects has not yet been analyzed. Our results indicate that the studied
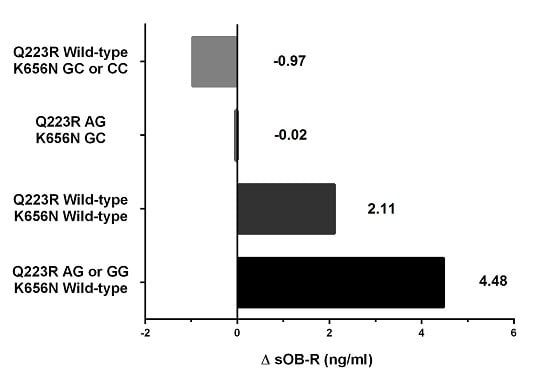
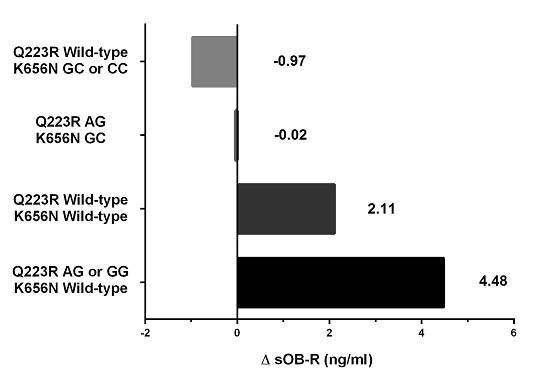
LEPR polymorphisms may have a complementary effect on the weight loss process in obese prepubertal children during lifestyle intervention. Among different combinations of both
LEPR SNPs, we found that children with at least one minor allele in Q223R coupled with wild-type K665N had the greatest likelihood of losing weight (5-fold), the greatest increase in soluble leptin receptor concentrations and decrease in SDS-BMI correlated with a decrease in fat mass during intervention. In contrast, carriers of the wild-type Q223R together with at least one minor allele in K665N, presented no difference in likelihood of weight loss when compared with the reference genotype combination. Moreover, only this tested combination showed a tendency towards a decrease in soluble leptin receptor concentration and towards a decrease in SDS-BMI correlated with a decrease in fat-free mass after therapy. It is known that a change of amino acid physico-chemical properties is one of the mechanisms of SNPs affecting protein stability and function, as well as protein-protein interactions [
44]. Amino acid substitutions in leptin receptor domain due to SNP in
LEPR result in changes of the electric charge from neutral to positive for Q223→R and from positive to neutral for K656→N [
7]. In our study, in the first combination of both
LEPR SNPs, mentioned above, resulting in higher odds of weight loss, 223R and K565 were associated with positive electric charge, while in the latter combination, resulting in the lack of the higher likelihood to lose weight, both SNP variants, Q223 and 565N, were associated with neutral electric charge. Therefore, it seems that the consequence of a substitution of one amino acid by another can be a change in leptin-leptin receptor interactions caused by the differences in charge of amino acids, that may alter the efficiency of the weight loss process in children.
In some studies, functional SNPs in
ADIPOQ were found to have a significant effect on human body composition, but other reports did not confirm an association between these polymorphisms and obesity [
45,
46]. In our previous study we found that the -11377C > G
ADIPOQ polymorphism may modulate the risk of childhood obesity and this polymorphism had a significant impact on the adipokine profile in prepubertal obese children [
20]. In the present study, we observed an increase in adiponectin concentrations in obese children with weight loss, while levels of this adipokine remained unchanged in children without weight loss. However, we did not observe any associations between the two analyzed
ADIPOQ polymorphisms and reduction in body mass in prepubertal children after 3 months of therapy. Sorensen
et al. [
47], when analyzing the -11377C > G
ADIPOQ SNP in a panel of obesity-related candidate genes in approximately 800 adults, suggested its minor role, if any, in modulating weight changes induced by a moderate hypo-energetic low-fat diet. However, other authors studying 2 SNPs in genes encoding adiponectin (-11377C > G
ADIPOQ) and its receptor 1 suggested that these polymorphisms may play a role in the responsiveness to dietary fatty acid modification in adults with metabolic syndrome [
13]. Several
ADIPOQ polymorphisms were also studied in Spanish obese adolescents, but none of them were associated with SDS-BMI and fat mass changes after 3 months of multidisciplinary treatment [
15], which is in concordance with our results.
This study has a few potential limitations. Firstly, adiposity was assessed by indirect estimations (BMI, BMI-SDS, skinfold thickness); nevertheless, the same methods were used before and after intervention, so analyzing differences in these parameters should obviate possible measurement errors. Secondly, our findings were obtained from a relatively small sample of subjects, though thoroughly characterized in terms of phenotypes. Therefore, ethnically matched studies should be performed to confirm our results in different populations of prepubertal obese children. Thirdly, the 3-day records provide only fragmentary information about dietary intake. However, these food records were reported to yield the strongest agreement with actual dietary intake compared with 24-h recall and 5-day food frequency records in children [
48]. Finally, the lifestyle intervention period in this study was only 3 months and a study of a long-term intervention is needed to verify the relationship between the polymorphisms in adipokine genes and adipokine levels in relation to clinical outcomes.

 ,
, 



{kind=link}
{kind=link}