Effects of n-3 PUFAs on Intestinal Mucosa Innate Immunity and Intestinal Microbiota in Mice after Hemorrhagic Shock Resuscitation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Study Design
2.3. Bacterial Culture
2.4. Enzyme-Linked Immunosorbent Assay
2.5. Western Blot Analysis of Lysozyme and Mucin 2
2.6. Quantitative Polymerase Chain Reaction Analysis of the Antibacterial Lectin RegIIIγ and Cryptdin 24
2.7. Immunohistochemistry of Lysozyme in Ileal Tissue Samples
2.8. Periodic Acid-Schiff Staining
2.9. DNA Extraction and 16S rRNA Pyrosequencing
2.10. Statistical Analysis
3. Results
3.1. Bacterial Culture and Colony Counting
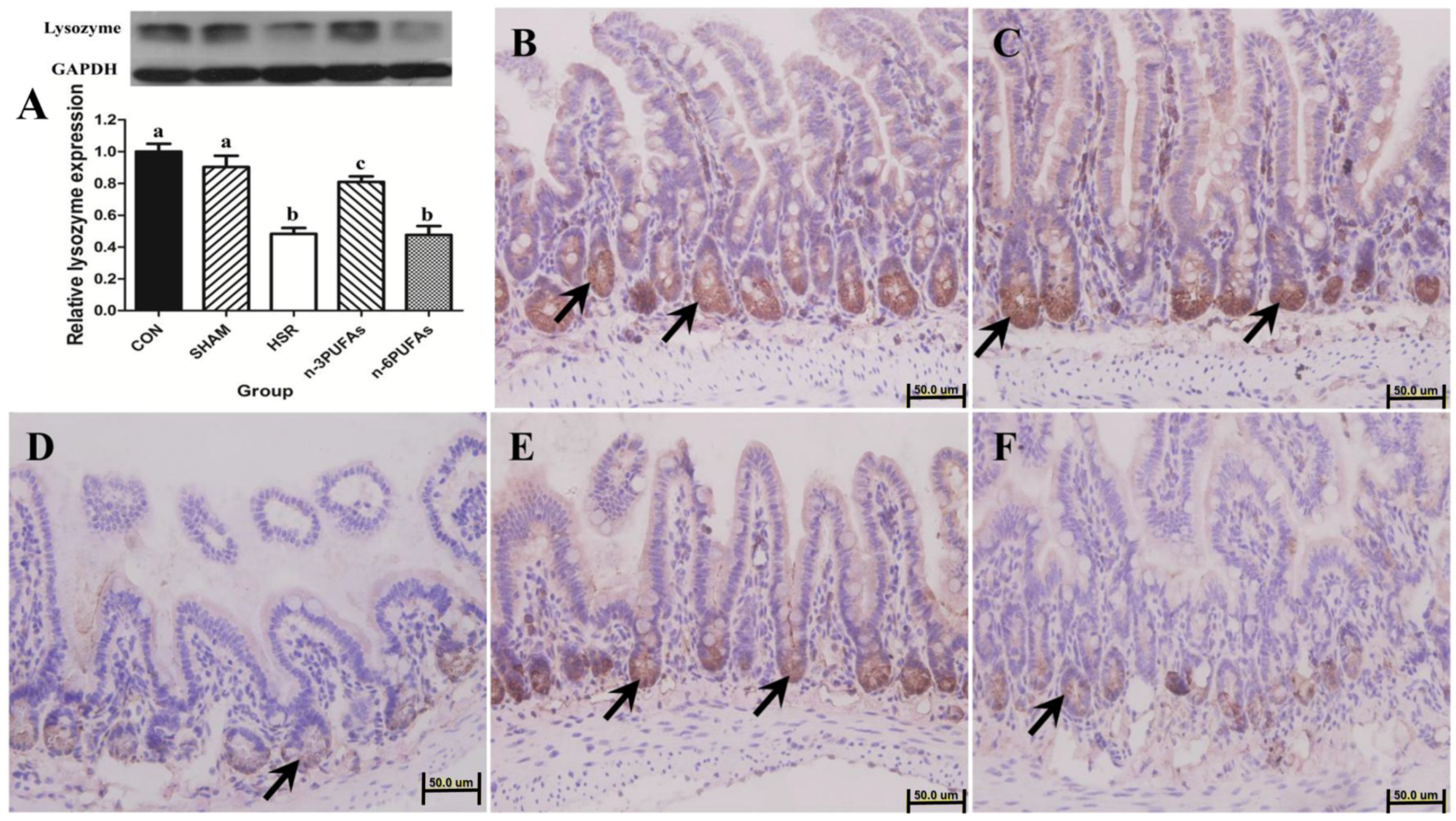
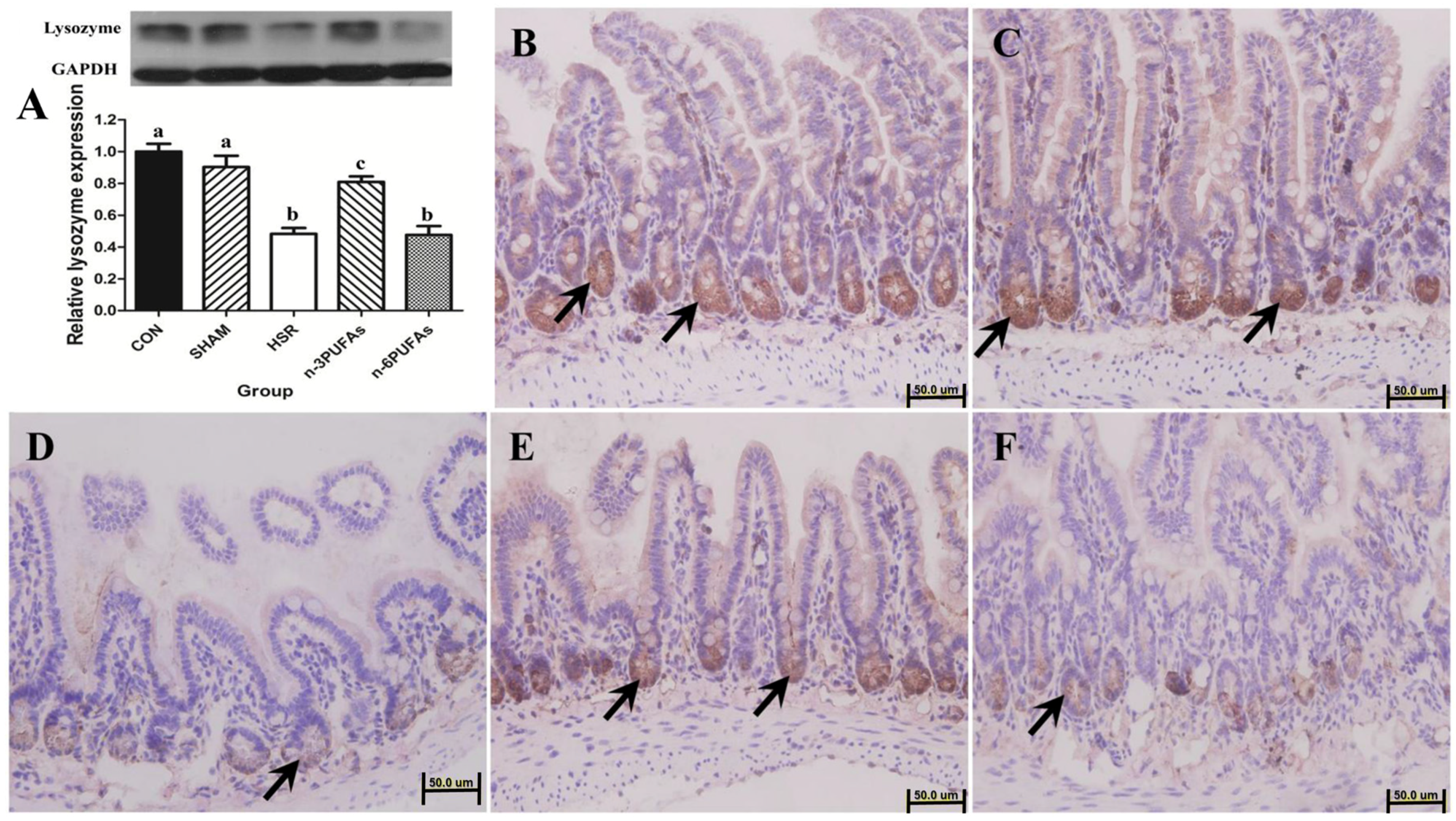
3.2. Expression and Localization of Lysozyme in Ileal Tissue
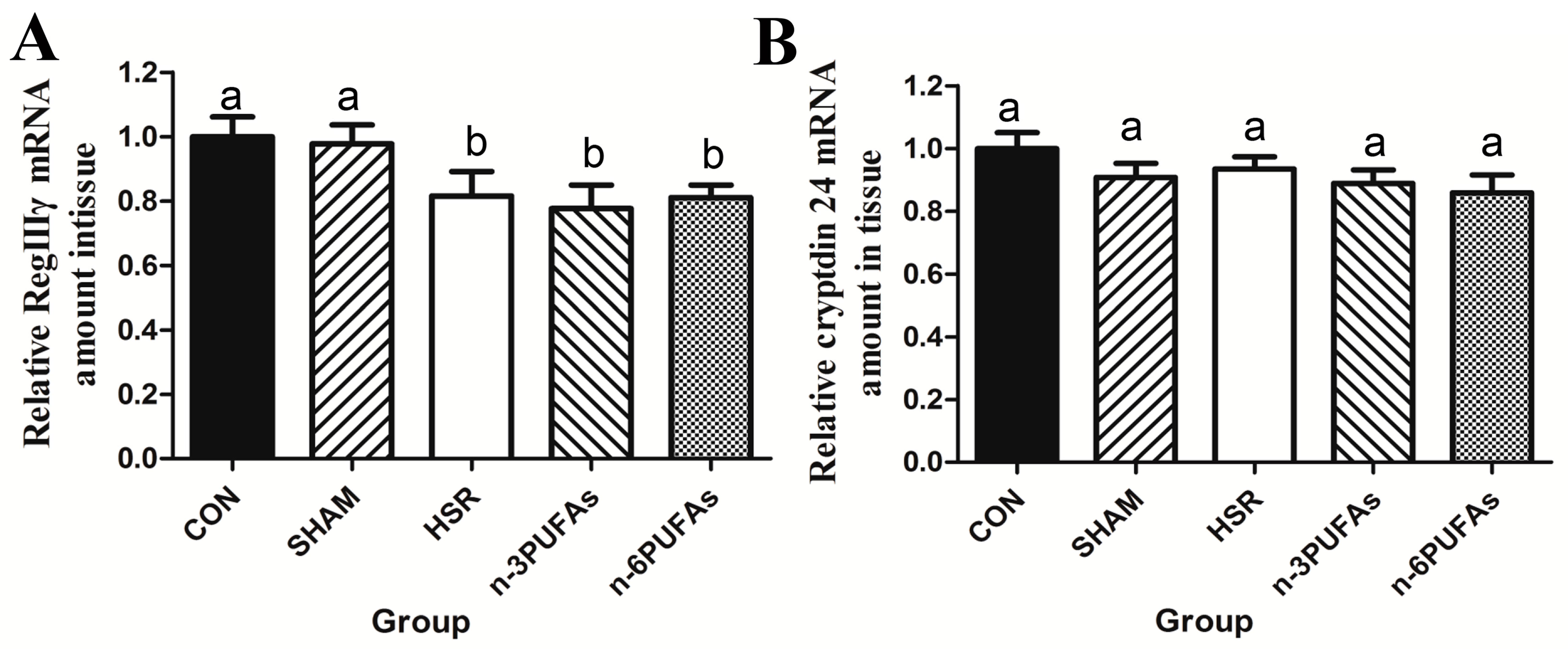
3.3. mRNA Levels of RegIIIγ and Cryptdin 24 in Ileal Tissue
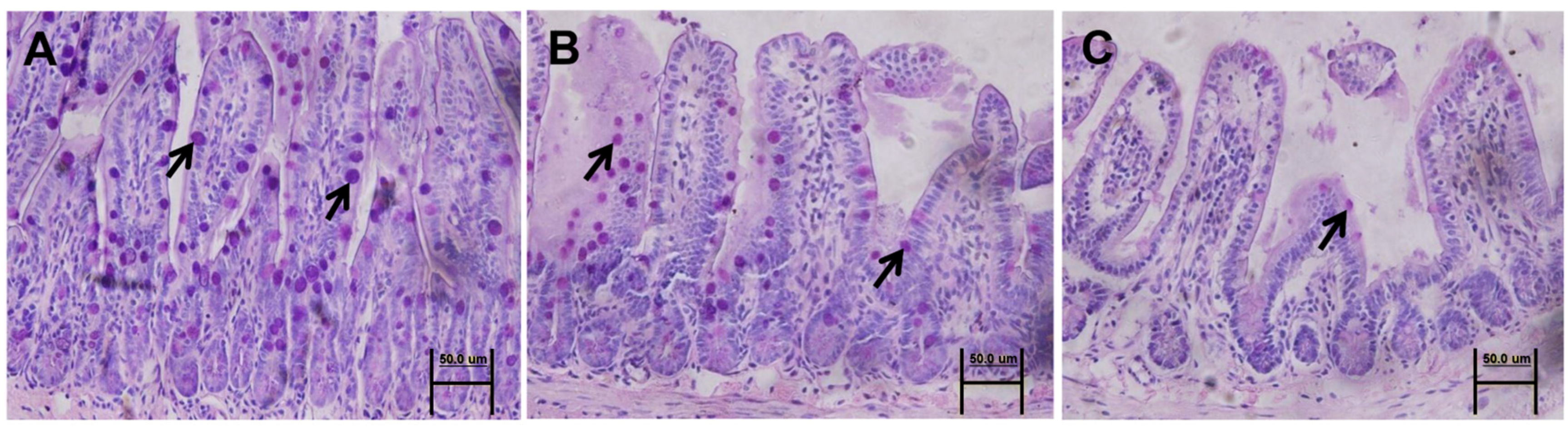
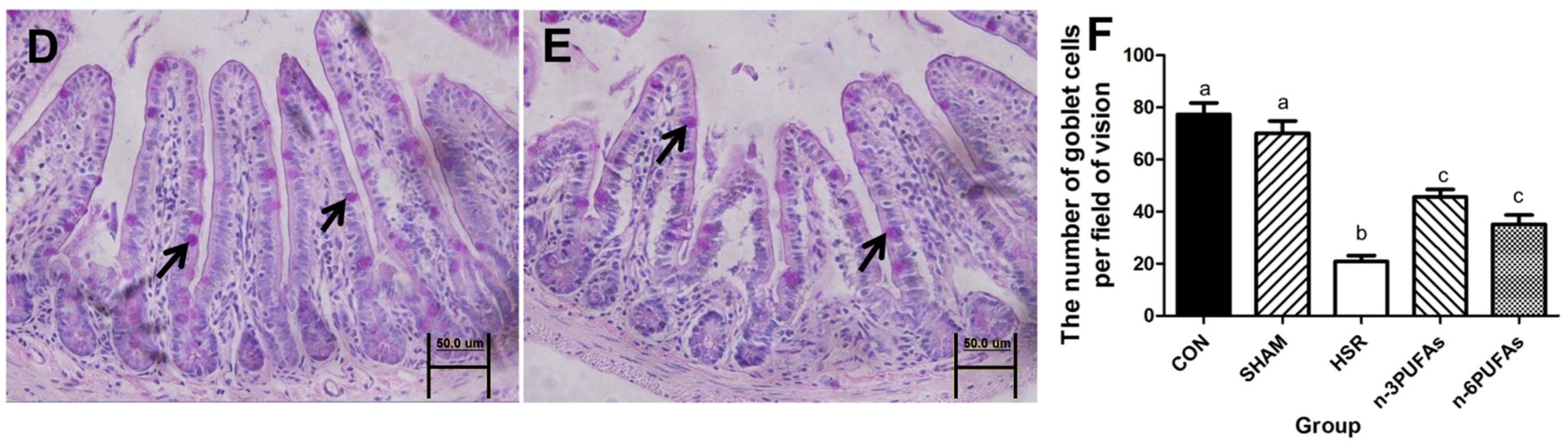
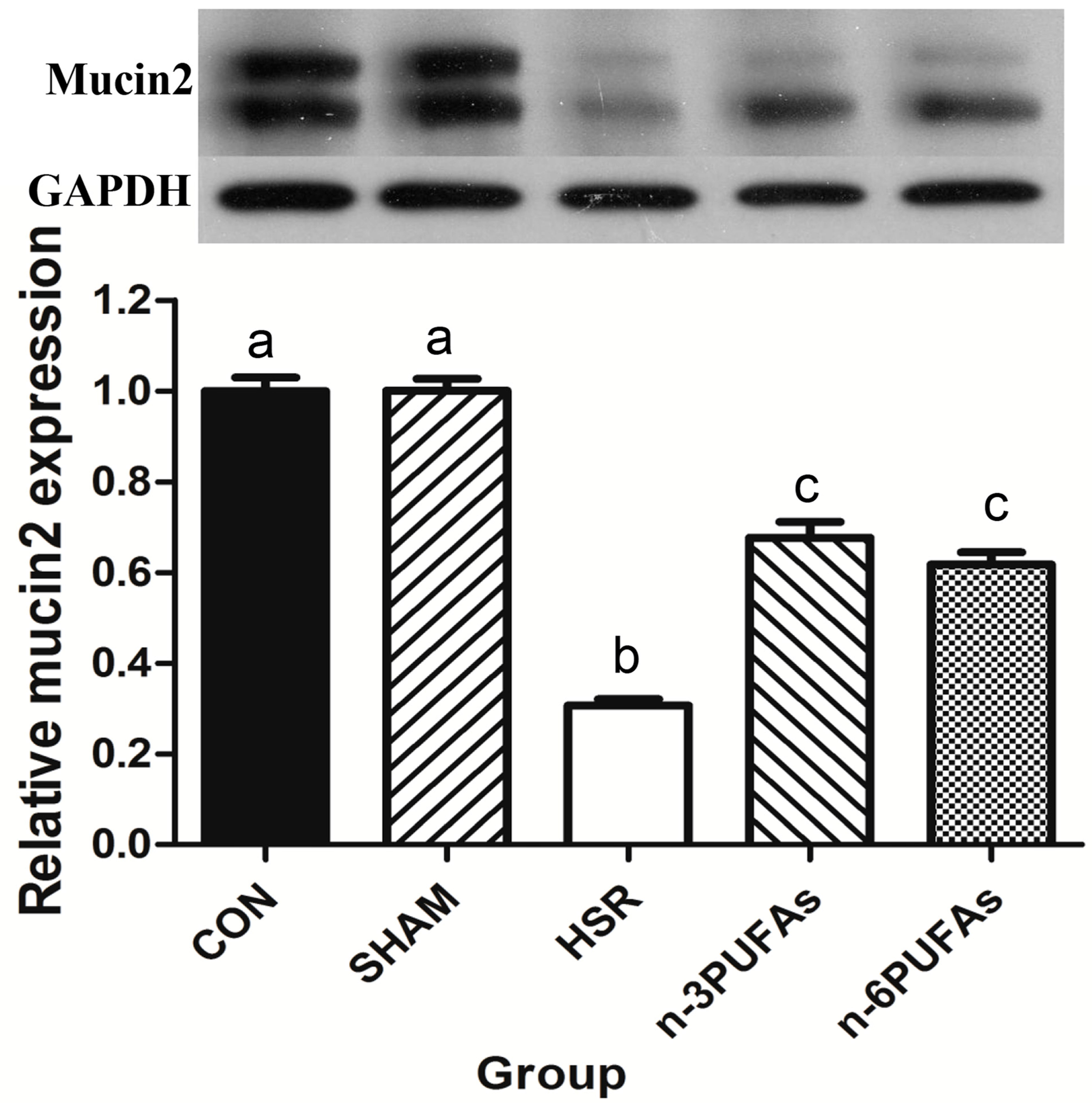
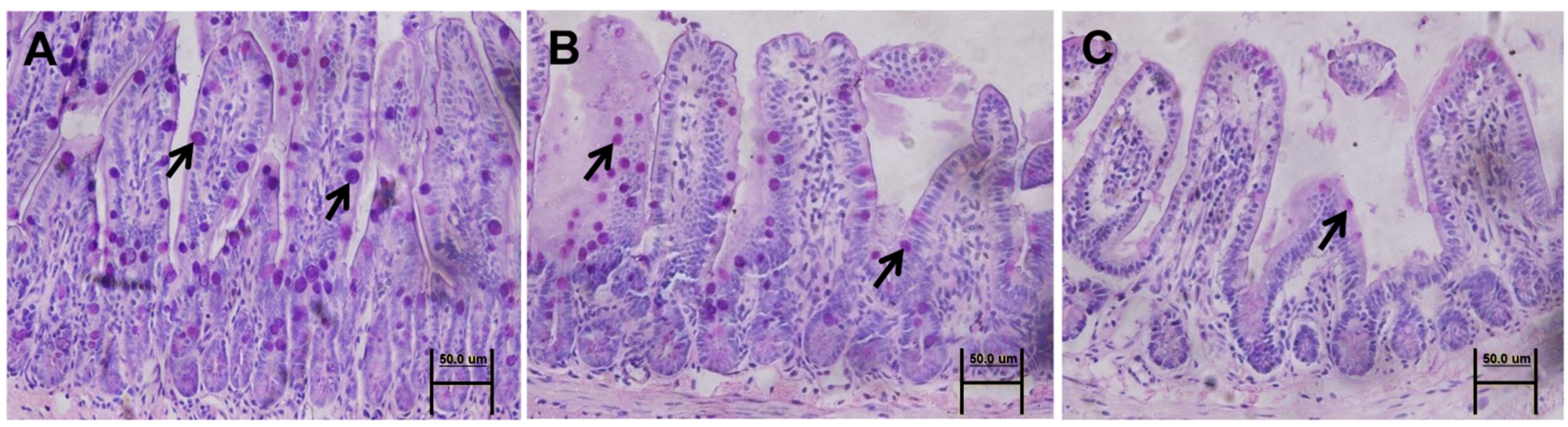
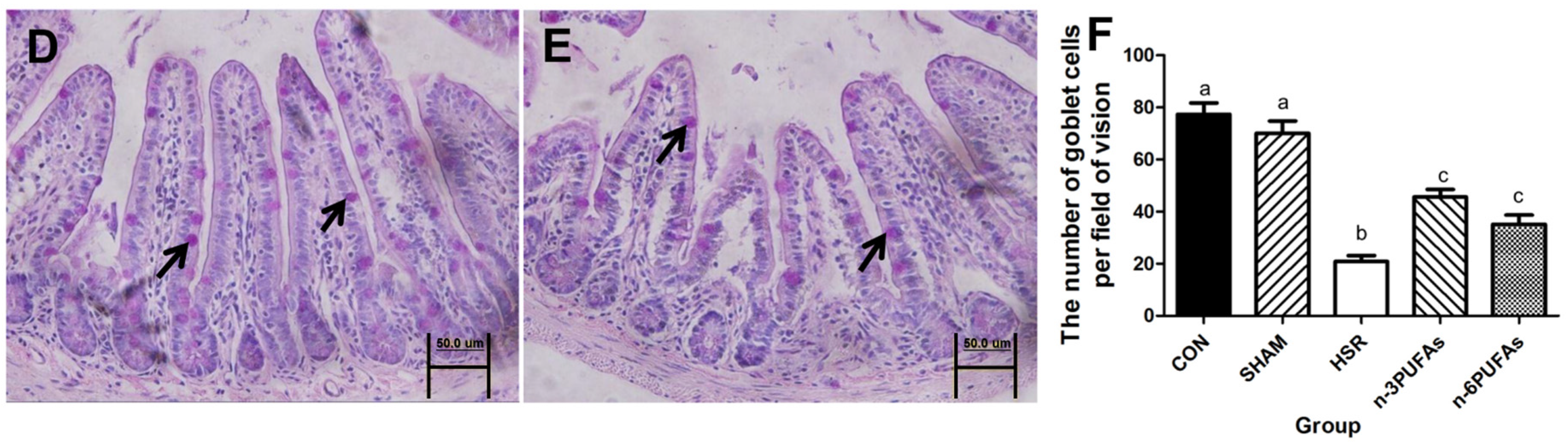
3.4. Mucin 2 Expression and Goblet Cell Count
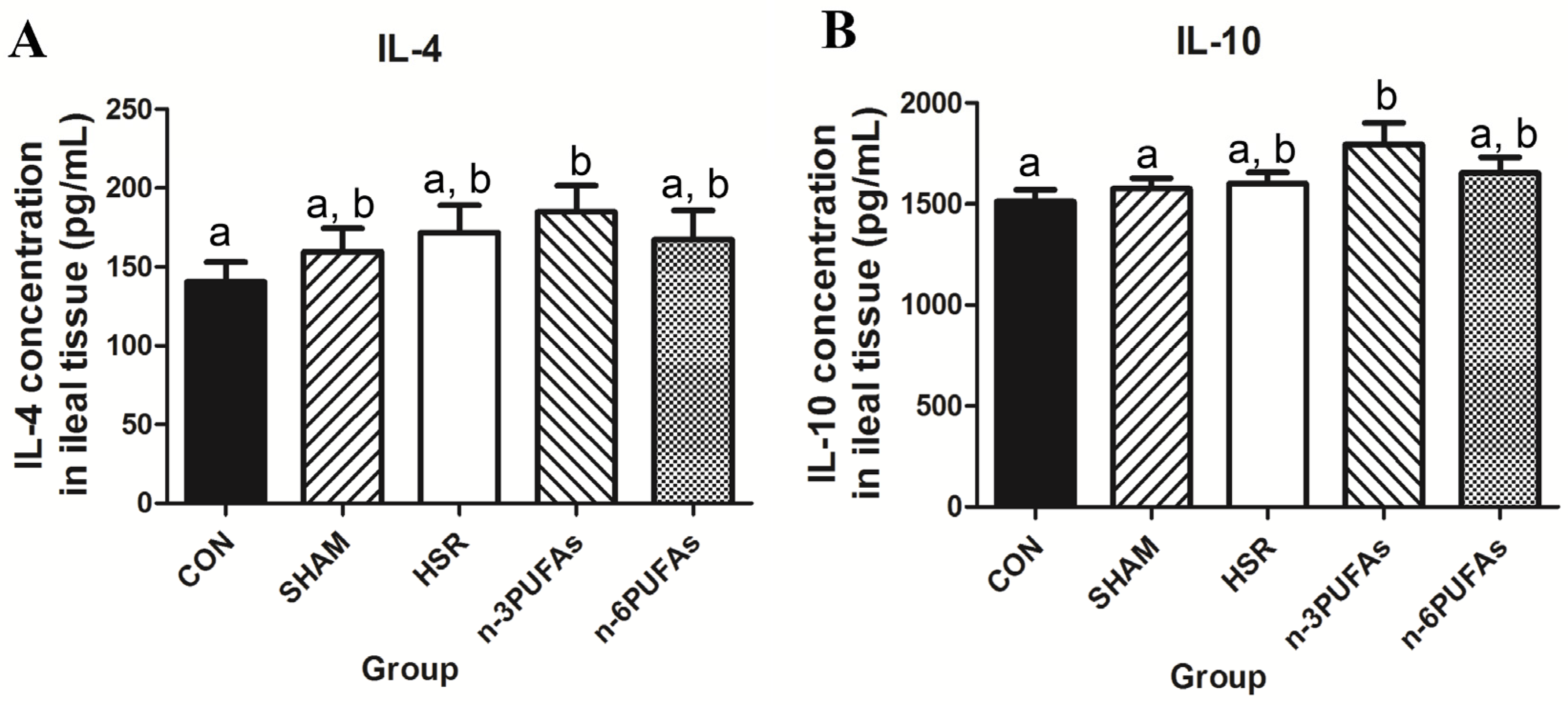
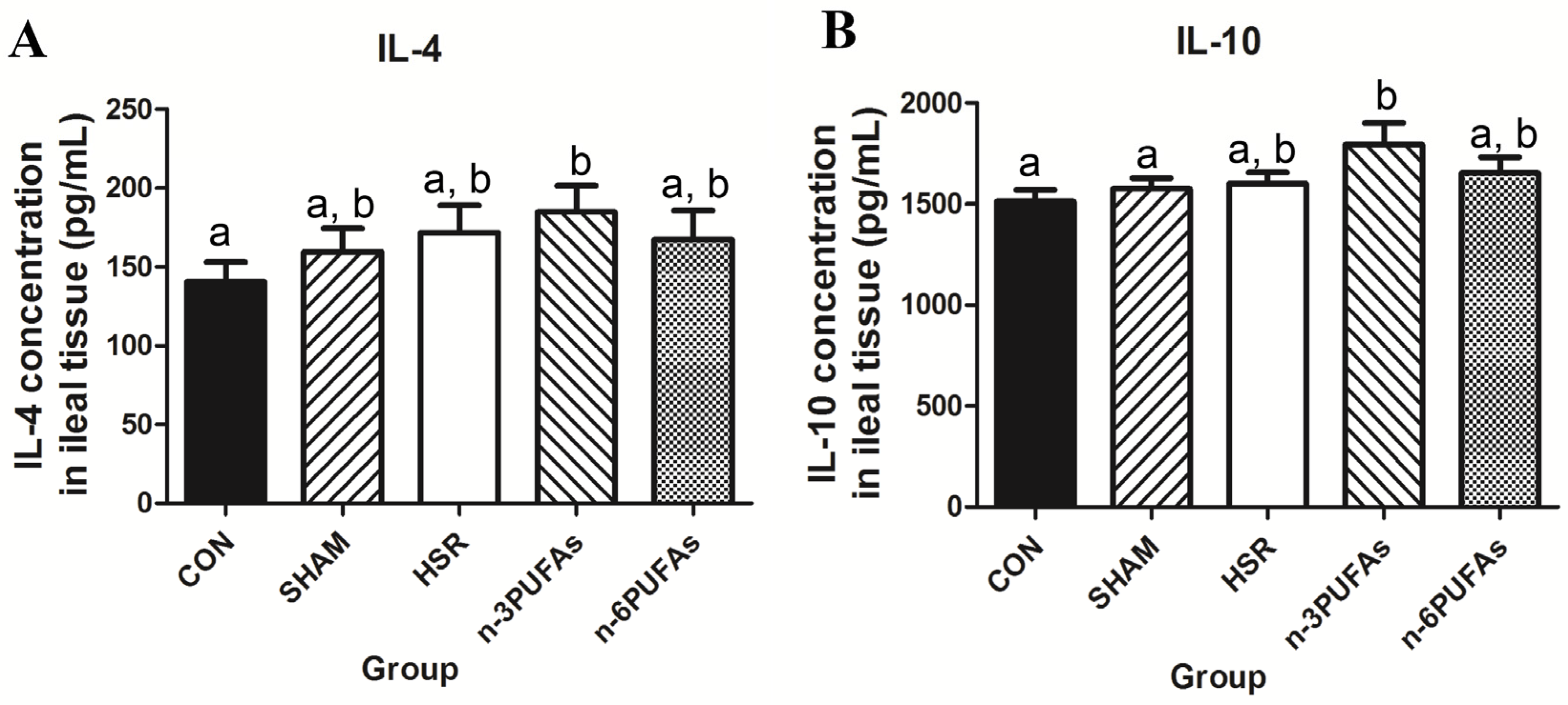
3.5. Levels of IL-4 and IL-10 in Ileal Tissue
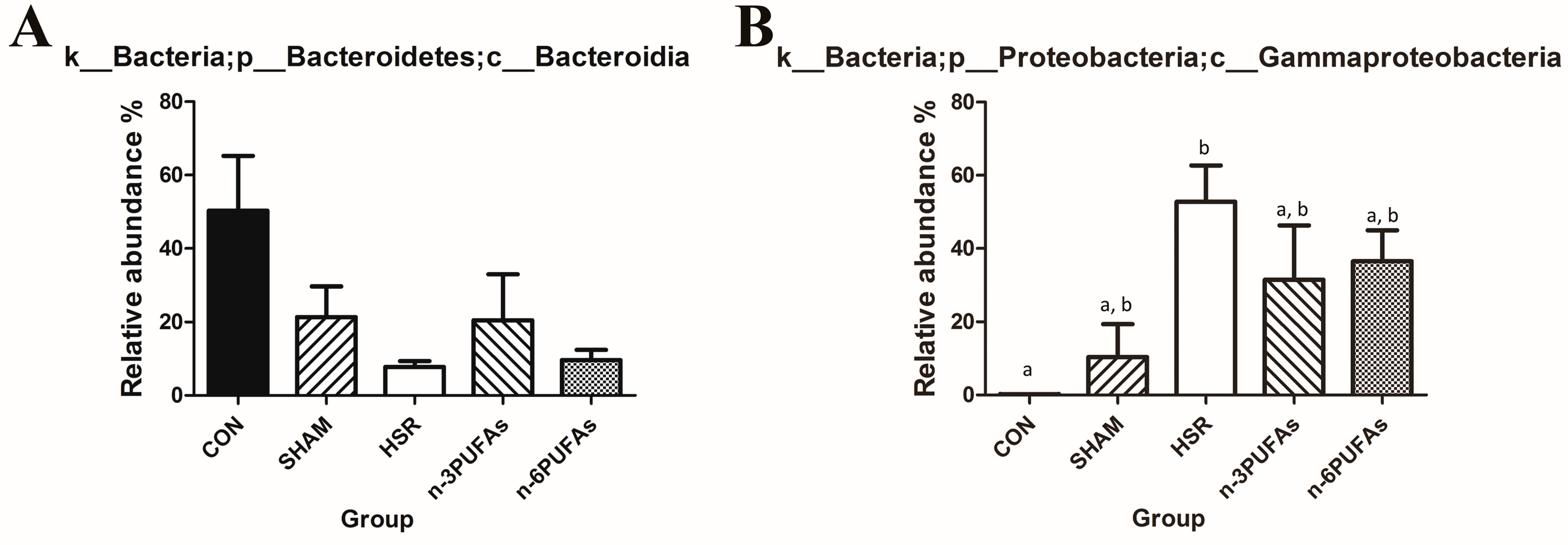
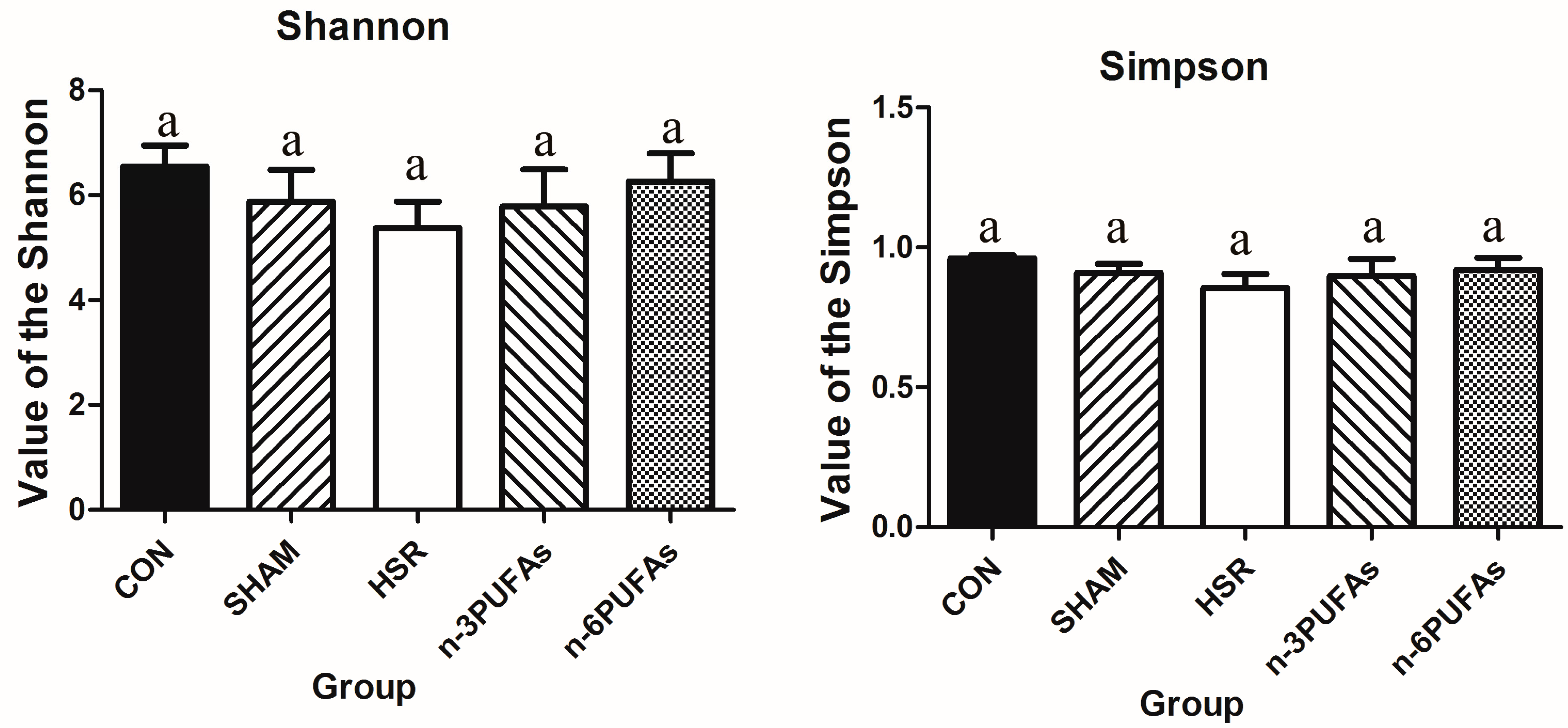
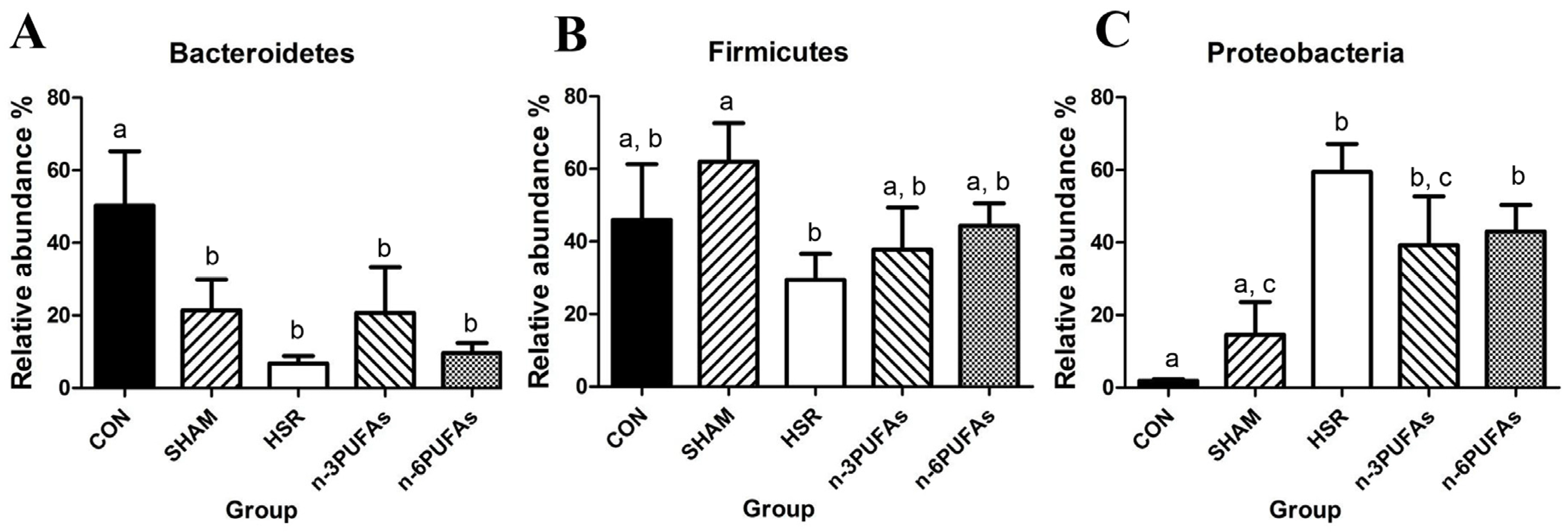
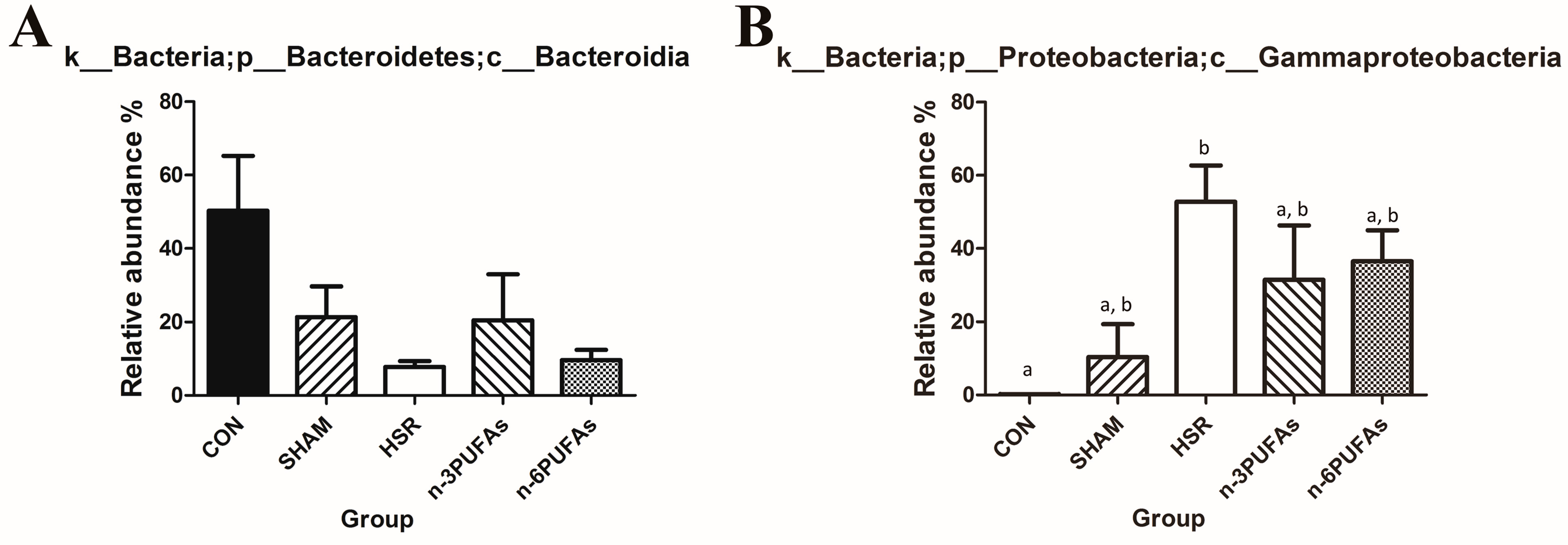
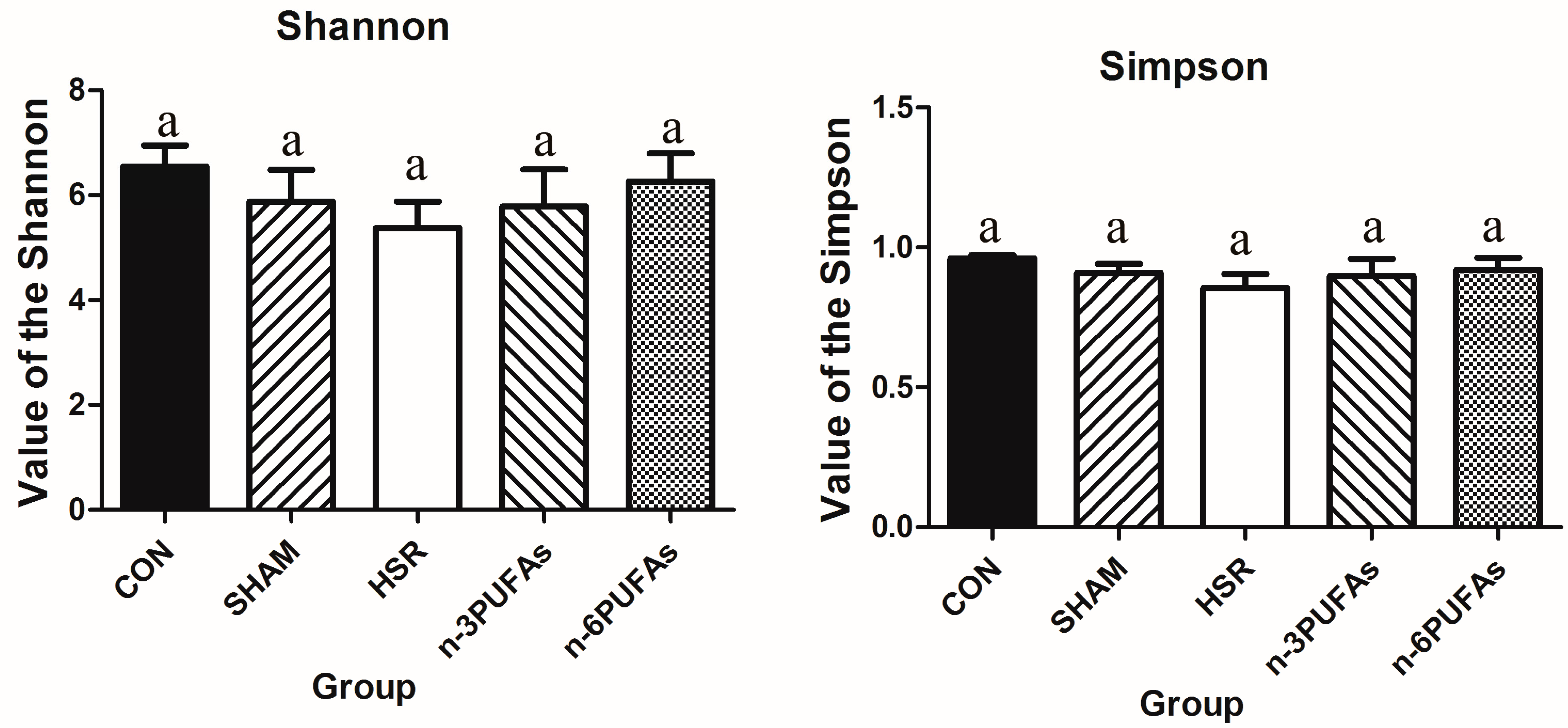
3.6. Relative Abundance of Bacteria in the Intestinal Mucosa
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| PUFAs | Polyunsaturated fatty acids |
| HSR | Hemorrhagic shock resuscitation |
| IRI | Ischemia-reperfusion injury |
| IMII | Intestinal mucosa innate immunity |
| CON | Control |
| MAP | Mean arterial pressure |
| PBS | Phosphate-buffered saline |
| CFUs | Colony-forming units |
| ELISA | Enzyme-linked immunosorbent assay |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
References
- American Gastroenterological Association. American gastroenterological association medical position statement: Guidelines on intestinal ischemia. Gastroenterology 2000, 118, 951–953. [Google Scholar]
- Sapalidis, K.; Papavramidis, T.S.; Gialamas, E.; Deligiannidis, N.; Tzioufa, V.; Papavramidis, S. The role of allopurinol’s timing in the ischemia reperfusion injury of small intestine. J. Emerg. Trauma Shock 2013, 6, 203–208. [Google Scholar] [PubMed]
- Vollmar, B.; Menger, M.D. Intestinal ischemia/reperfusion: Microcirculatory pathology and functional consequences. Langenbecks Arch. Surg. 2011, 396, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Kassahun, W.T.; Schulz, T.; Richter, O.; Hauss, J. Unchanged high mortality rates from acute occlusive intestinal ischemia: Six year review. Langenbecks Arch. Surg. 2008, 393, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Valparaiso, A.P.; Vicente, D.A.; Bograd, B.A.; Elster, E.A.; Davis, T.A. Modeling acute traumatic injury. J. Surg. Res. 2015, 194, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, X.; Li, N.; Li, J. The study of n-3 PUFAs protecting the intestinal barrier in rat HS/R model. Lipids Health Dis. 2014, 13. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pan, L.; Lu, J.; Li, N.; Li, J. N-3 PUFAs attenuate ischemia/reperfusion induced intestinal barrier injury by activating I-FABP-PPARgamma pathway. Clin. Nutr. 2012, 31, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Wang, X.; Li, Q.; Li, N.; Li, J. Essential amino acid enriched high-protein enteral nutrition modulates insulin-like growth factor-1 system function in a rat model of trauma-hemorrhagic shock. PLoS ONE 2013, 8, e77823. [Google Scholar] [CrossRef] [PubMed]
- Anastasilakis, C.D.; Ioannidis, O.; Gkiomisi, A.I.; Botsios, D. Artificial nutrition and intestinal mucosal barrier functionality. Digestion 2013, 88, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Bevins, C.L.; Salzman, N.H. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat. Rev. Microbiol. 2011, 9, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Grootjans, J.; Hundscheid, I.H.; Lenaerts, K.; Boonen, B.; Renes, I.B.; Verheyen, F.K.; Dejong, C.H.; von Meyenfeldt, M.F.; Beets, G.L.; Buurman, W.A. Ischaemia-induced mucus barrier loss and bacterial penetration are rapidly counteracted by increased goblet cell secretory activity in human and rat colon. Gut 2013, 62, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Cullen, T.W.; Schofield, W.B.; Barry, N.A.; Putnam, E.E.; Rundell, E.A.; Trent, M.S.; Degnan, P.H.; Booth, C.J.; Yu, H.; Goodman, A.L. Gut microbiota. Antimicrobial peptide resistance mediates resilience of prominent gut commensals during inflammation. Science 2015, 347, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Grootjans, J.; Hodin, C.M.; de Haan, J.J.; Derikx, J.P.; Rouschop, K.M.; Verheyen, F.K.; van Dam, R.M.; Dejong, C.H.; Buurman, W.A.; Lenaerts, K. Level of activation of the unfolded protein response correlates with paneth cell apoptosis in human small intestine exposed to ischemia/reperfusion. Gastroenterology 2011, 140, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Grau-Carmona, T.; Bonet-Saris, A.; Garcia-de-Lorenzo, A.; Sanchez-Alvarez, C.; Rodriguez-Pozo, A.; Acosta-Escribano, J.; Minambres, E.; Herrero-Meseguer, J.I.; Mesejo, A. Influence of n-3 polyunsaturated fatty acids enriched lipid emulsions on nosocomial infections and clinical outcomes in critically ill patients: Icu lipids study. Crit. Care Med. 2015, 43, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Manzanares, W.; Langlois, P.L.; Dhaliwal, R.; Lemieux, M.; Heyland, D.K. Intravenous fish oil lipid emulsions in critically ill patients: An updated systematic review and meta-analysis. Crit. Care 2015, 19. [Google Scholar] [CrossRef] [PubMed]
- Lei, Q.C.; Wang, X.Y.; Xia, X.F.; Zheng, H.Z.; Bi, J.C.; Tian, F.; Li, N. The role of Omega-3 fatty acids in acute pancreatitis: A meta-analysis of randomized controlled trials. Nutrients 2015, 7, 2261–2273. [Google Scholar] [CrossRef] [PubMed]
- Ticinesi, A.; Meschi, T.; Lauretani, F.; Felis, G.; Franchi, F.; Pedrolli, C.; Barichella, M.; Benati, G.; Di Nuzzo, S.; Ceda, G.; et al. Nutrition and inflammation in older individuals: Focus on vitamin D, n-3 polyunsaturated fatty acids and whey proteins. Nutrients 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.E.; McClave, S.A.; Martindale, R.G.; Warren, M.M.; Johnson, D.R.; Braunschweig, C.; McCarthy, M.S.; Davanos, E.; Rice, T.W.; Cresci, G.A.; et al. Guidelines for the provision and assessment of nutrition support therapy in the adult critically ill patient: Society of critical care medicine (SCCM) and american society for parenteral and enteral nutrition (A.S.P.E.N.). Crit. Care Med. 2016, 44, 390–438. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Bi, J.; Gao, X.; Tian, F.; Wang, X.; Li, N.; Li, J. Partial enteral nutrition preserves elements of gut barrier function, including innate immunity, intestinal alkaline phosphatase (IAP) level, and intestinal microbiota in mice. Nutrients 2015, 7, 6294–6312. [Google Scholar] [CrossRef] [PubMed]
- Aligned 16S rDNA Data and Tools. Available online: http://greengenes.lbl.gov/cgi-bin/nph-index.cgi (accessed on 2 October 2011).
- Wang, X.; Pierre, J.F.; Heneghan, A.F.; Busch, R.A.; Kudsk, K.A. Glutamine improves innate immunity and prevents bacterial enteroinvasion during parenteral nutrition. J. Parenter. Enter. Nutr. 2014, 39, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Hodin, C.M.; Lenaerts, K.; Grootjans, J.; de Haan, J.J.; Hadfoune, M.; Verheyen, F.K.; Kiyama, H.; Heineman, E.; Buurman, W.A. Starvation compromises paneth cells. Am. J. Pathol. 2011, 179, 2885–2893. [Google Scholar] [CrossRef] [PubMed]
- Tani, T.; Fujino, M.; Hanasawa, K.; Shimizu, T.; Endo, Y.; Kodama, M. Bacterial translocation and tumor necrosis factor-alpha gene expression in experimental hemorrhagic shock. Crit. Care Med. 2000, 28, 3705–3709. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.-X.; Chen, S.; Ma, L.-P.; Jiang, L.-Y.; Chen, J.-W.; Chang, R.-M.; Wen, L.-Q.; Wu, W.; Jiang, Z.-P.; Huang, Z.-T. Functional and morphological changes of the gut barrier during the restitution process after hemorrhagic shock. World J. Gastroenterol. 2005, 11, 5485–5491. [Google Scholar] [CrossRef] [PubMed]
- Fishman, J.E.; Levy, G.; Alli, V.; Zheng, X.; Mole, D.J.; Deitch, E.A. The intestinal mucus layer is a critical component of the gut barrier that is damaged during acute pancreatitis. Shock 2014, 42, 264–270. [Google Scholar] [CrossRef] [PubMed]
- De Matos, O.G.; Amaral, S.S.; Marques Pereira da Silva, P.E.; Perez, D.A.; Alvarenga, D.M.; Matos Ferreira, A.V.; Alvarez-Leite, J.; Menezes, G.B.; Cara, D.C. Dietary supplementation with omega-3-PUFA-rich fish oil reduces signs of food allergy in ovalbumin-sensitized mice. Clin. Dev. Immunol. 2012, 2012, 236564. [Google Scholar] [CrossRef] [PubMed]
- Hekmatdoost, A.; Wu, X.; Morampudi, V.; Innis, S.M.; Jacobson, K. Dietary oils modify the host immune response and colonic tissue damage following citrobacter rodentium infection in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G917–G928. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, W.; Zhang, F.; Pan, L.; Li, N.; Li, J. Fish oil-supplemented parenteral nutrition in severe acute pancreatitis patients and effects on immune function and infectious risk: A randomized controlled trial. Inflammation 2009, 32, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Hecker, M.; Ott, J.; Sondermann, C.; Schaefer, M.; Obert, M.; Hecker, A.; Morty, R.E.; Vadasz, I.; Herold, S.; Rosengarten, B.; et al. Immunomodulation by fish-oil containing lipid emulsions in murine acute respiratory distress syndrome. Crit. Care 2014, 18. [Google Scholar] [CrossRef] [PubMed]
- Juman, S.; Hashimoto, M.; Katakura, M.; Inoue, T.; Tanabe, Y.; Arita, M.; Miki, T.; Shido, O. Effects of long-term oral administration of arachidonic acid and docosahexaenoic acid on the immune functions of young rats. Nutrients 2013, 5, 1949–1961. [Google Scholar] [CrossRef] [PubMed]
- Dabbagh, K.; Takeyama, K.; Lee, H.M.; Ueki, I.F.; Lausier, J.A.; Nadel, J.A. IL-4 induces mucin gene expression and goblet cell metaplasia in vitro and in vivo. J. Immunol. 1999, 162, 6233–6237. [Google Scholar] [PubMed]
- Heneghan, A.F.; Pierre, J.F.; Kudsk, K.A. JAK-STAT and intestinal mucosal immunology. JAK-STAT 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Doherty, T.A.; Khorram, N.; Lund, S.; Mehta, A.K.; Croft, M.; Broide, D.H. Lung type 2 innate lymphoid cells express cysteinyl leukotriene receptor 1, which regulates Th2 cytokine production. J. Allergy Clin. Immunol. 2013, 132, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Klose, C.S.; Artis, D. Innate lymphoid cells as regulators of immunity, inflammation and tissue homeostasis. Nat. Immunol. 2016, 17, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Li, N.N.; Zhou, Y.; Qin, X.P.; Chen, Y.; He, D.; Feng, J.Y.; Wu, X.T. Does intravenous fish oil benefit patients post-surgery? A meta-analysis of randomised controlled trials. Clin. Nutr. 2014, 33, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Morris, T.; Rajakariar, R.; Stables, M.; Gilroy, D.W. Not all eicosanoids are bad. Trends Pharmacol. Sci. 2006, 27, 609–611. [Google Scholar] [CrossRef] [PubMed]
- Schwab, J.M.; Serhan, C.N. Lipoxins and new lipid mediators in the resolution of inflammation. Curr. Opin. Pharmacol. 2006, 6, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L. Nitric oxide, aspirin-triggered lipoxins and no-aspirin in gastric protection. Inflamm. Allergy Drug Targets 2006, 5, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Dufton, N.; Perretti, M. Therapeutic anti-inflammatory potential of formyl-peptide receptor agonists. Pharmacol. Ther. 2010, 127, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Savill, J. Resolution of inflammation: The beginning programs the end. Nat. Immunol. 2005, 6, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, T.; Ducheix, S.; le Faouder, P.; Perez, T.; Riols, F.; Boue, J.; Bertrand-Michel, J.; Dubourdeau, M.; Guillou, H.; Perretti, M.; et al. Protective effects of n-6 fatty acids-enriched diet on intestinal ischaemia/reperfusion injury involve lipoxin A4 and its receptor. Br. J. Pharmacol. 2015, 172, 910–923. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, Y.P.; Guo, P.; Ye, X.H.; Wang, J.; Yuan, S.Y.; Yao, S.L.; Shang, Y. A lipoxin A4 analog ameliorates blood-brain barrier dysfunction and reduces MMP-9 expression in a rat model of focal cerebral ischemia-reperfusion injury. J. Mol. Neurosci. 2012, 46, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.P.; Tjonahen, E.; Keledjian, R.; Zhu, M.; Yang, R.; Recchiuti, A.; Pillai, P.S.; Petasis, N.A.; Serhan, C.N. Anti-inflammatory and pro-resolving properties of benzo-lipoxin A4 analogs. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Mobraten, K.; Haug, T.M.; Kleiveland, C.R.; Lea, T. Omega-3 and Omega-6 PUFAs induce the same GPR120-mediated signalling events, but with different kinetics and intensity in Caco-2 cells. Lipids Health Dis. 2013, 12. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Pan, Y.; Yan, R.; Zeng, B.; Wang, H.; Zhang, X.; Li, W.; Wei, H.; Liu, Z. Commensal bacteria direct selective cargo sorting to promote symbiosis. Nat. Immunol. 2015, 16, 918–926. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer | Sequence (5′–3′) |
|---|---|---|
| GAPDH | Forward Primer | AGGCCGGTGCTGAGTATGTC |
| Reverse Primer | TGCCTGCTTCACCACCTTCT | |
| Cryptdin 24 | Forward Primer | TGAAGACACTAATCCTCCTCTCTGC |
| Reverse Primer | GCTCCTCAGTTTTAGTCTCTTCATCTGTA | |
| RegIIIγ | Forward Primer | GTATGATGCAGATATGGCCTG |
| Reverse Primer | ATATTGGCCACTGTTACCAC |
| Control | Sham | HSR | n-3 PUFAs | n-6 PUFAs | |
|---|---|---|---|---|---|
| Liver | 0 a | 0 a | 200 a | 0 a | 0 a |
| Spleen | 0 a | 0 a | 0 a | 0 a | 0 a |
| MLN | 0 a | 0 a | 4000 b | 550 c | 900 c |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, F.; Gao, X.; Zhang, L.; Wang, X.; Wan, X.; Jiang, T.; Wu, C.; Bi, J.; Lei, Q. Effects of n-3 PUFAs on Intestinal Mucosa Innate Immunity and Intestinal Microbiota in Mice after Hemorrhagic Shock Resuscitation. Nutrients 2016, 8, 609. https://doi.org/10.3390/nu8100609
Tian F, Gao X, Zhang L, Wang X, Wan X, Jiang T, Wu C, Bi J, Lei Q. Effects of n-3 PUFAs on Intestinal Mucosa Innate Immunity and Intestinal Microbiota in Mice after Hemorrhagic Shock Resuscitation. Nutrients. 2016; 8(10):609. https://doi.org/10.3390/nu8100609
Chicago/Turabian StyleTian, Feng, Xuejin Gao, Li Zhang, Xinying Wang, Xiao Wan, Tingting Jiang, Chao Wu, Jingcheng Bi, and Qiucheng Lei. 2016. "Effects of n-3 PUFAs on Intestinal Mucosa Innate Immunity and Intestinal Microbiota in Mice after Hemorrhagic Shock Resuscitation" Nutrients 8, no. 10: 609. https://doi.org/10.3390/nu8100609
APA StyleTian, F., Gao, X., Zhang, L., Wang, X., Wan, X., Jiang, T., Wu, C., Bi, J., & Lei, Q. (2016). Effects of n-3 PUFAs on Intestinal Mucosa Innate Immunity and Intestinal Microbiota in Mice after Hemorrhagic Shock Resuscitation. Nutrients, 8(10), 609. https://doi.org/10.3390/nu8100609