The Growth of SGC-7901 Tumor Xenografts Was Suppressed by Chinese Bayberry Anthocyanin Extract through Upregulating KLF6 Gene Expression


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Purification and Identification of C3G from Chinese Bayberry Fruit
2.3. Animal and Tumor Xenograft Studies
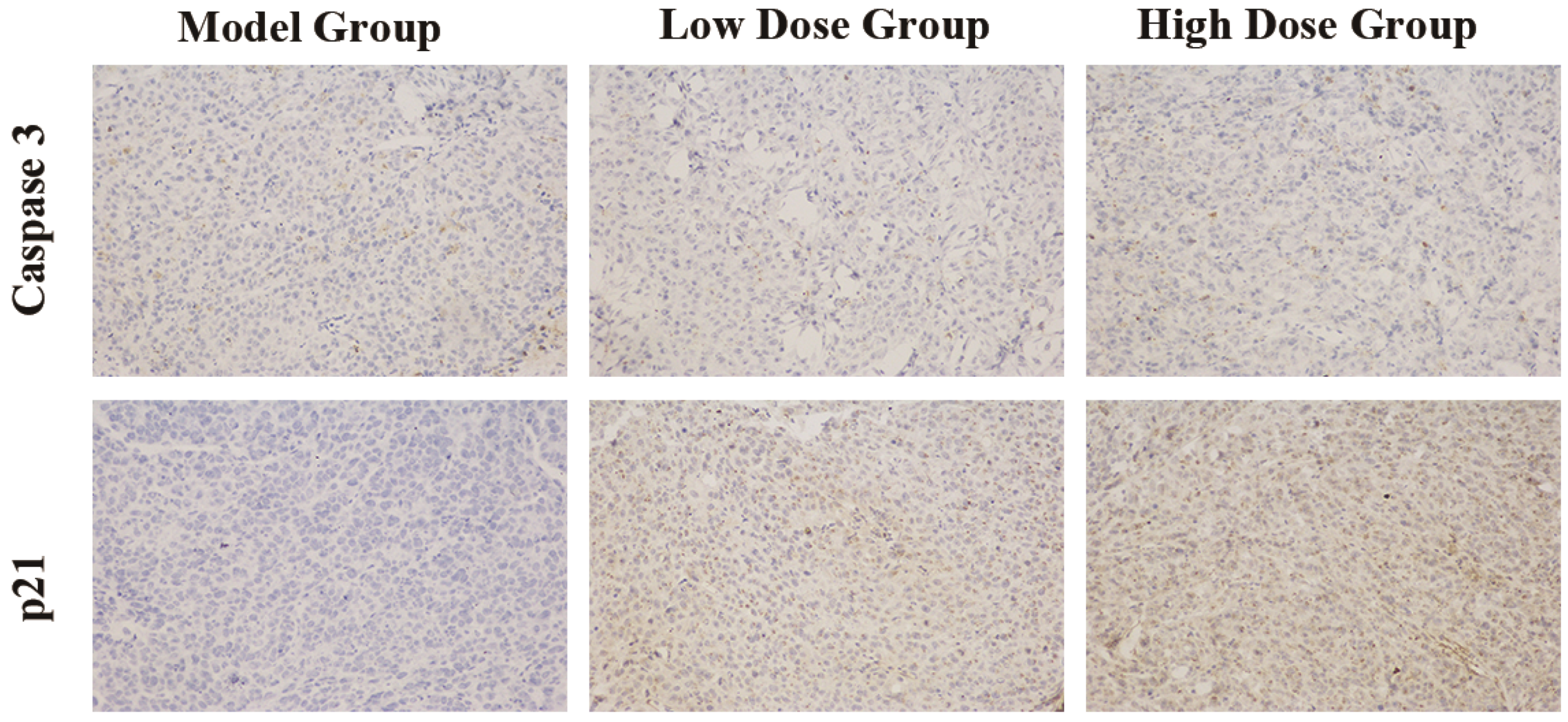
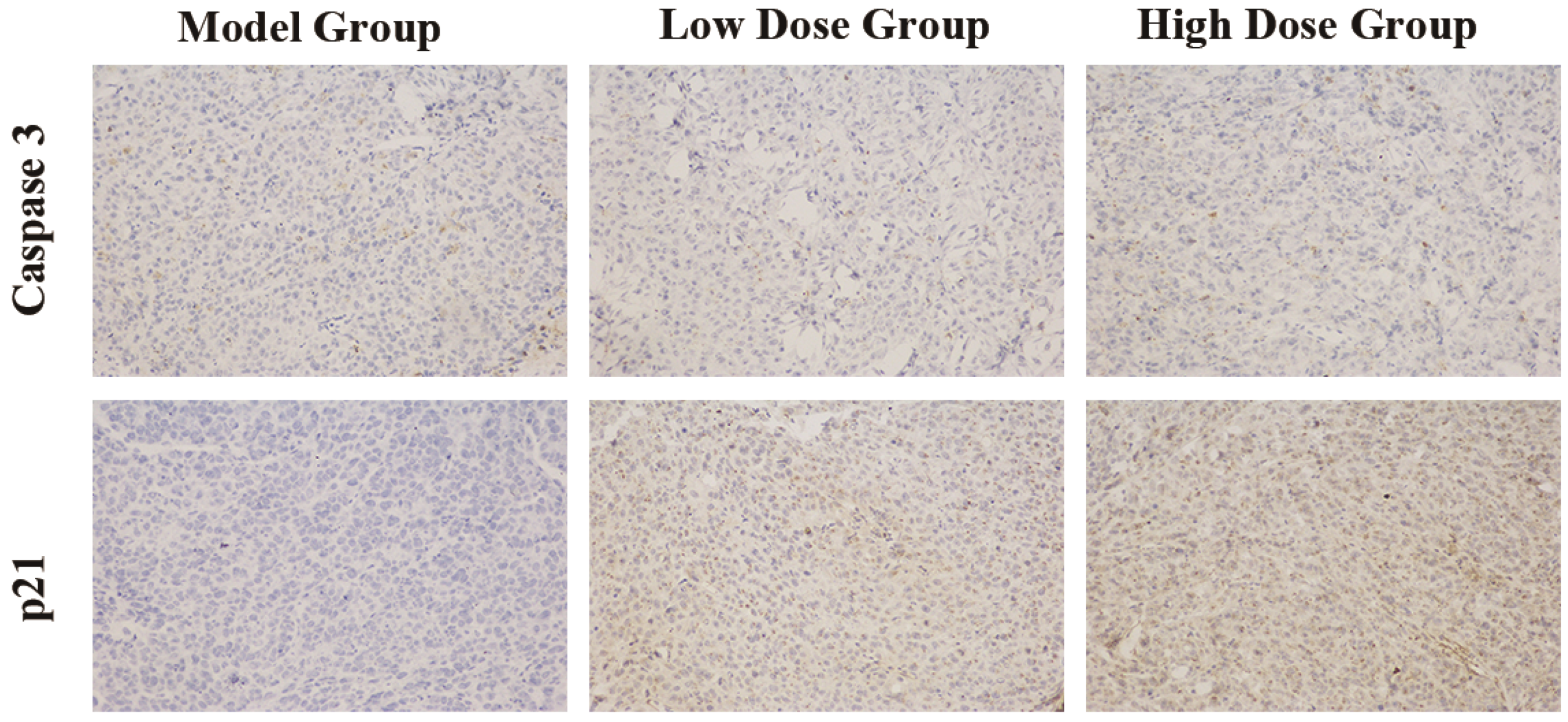
2.4. Immunohistochemical Staining
2.5. Quantitative Real-Time PCR and Western Blot Assay
2.6. Statistics
3. Results
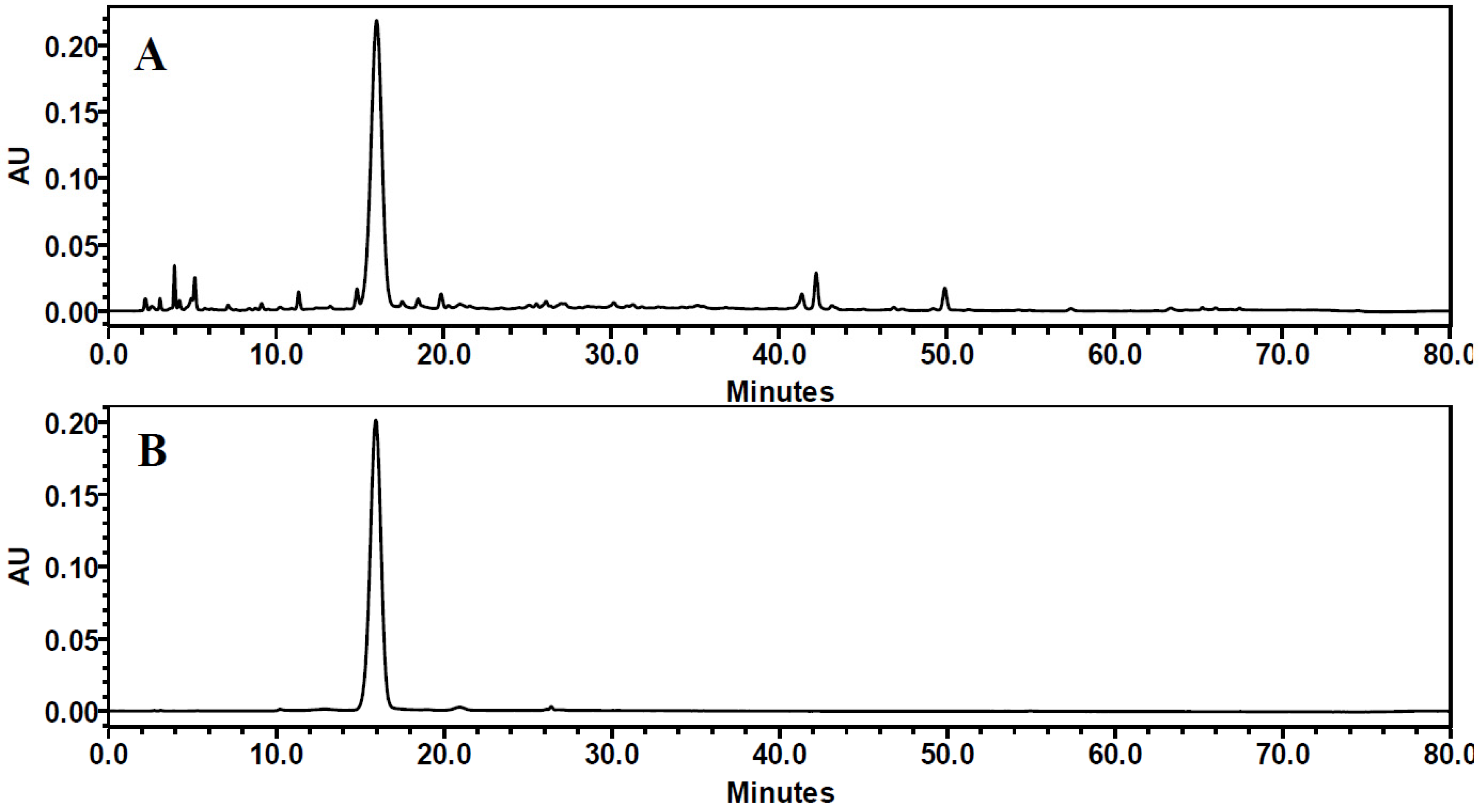
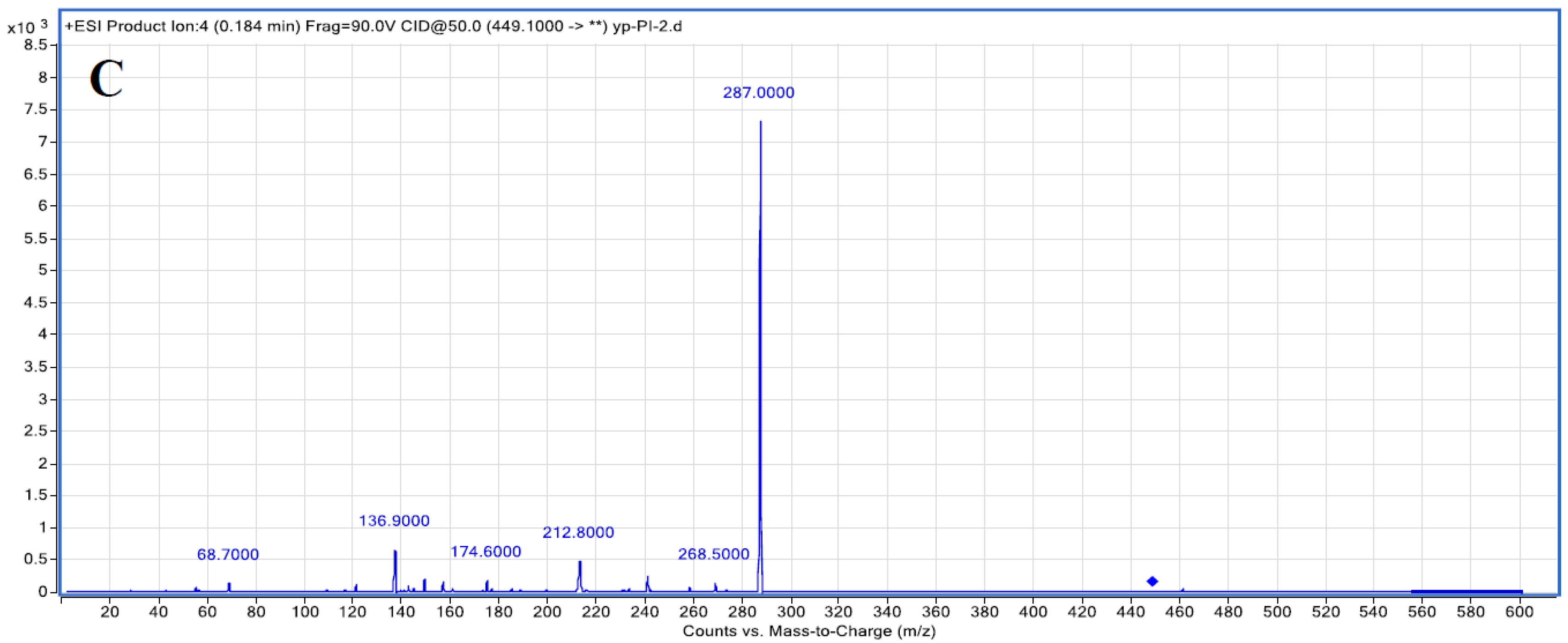
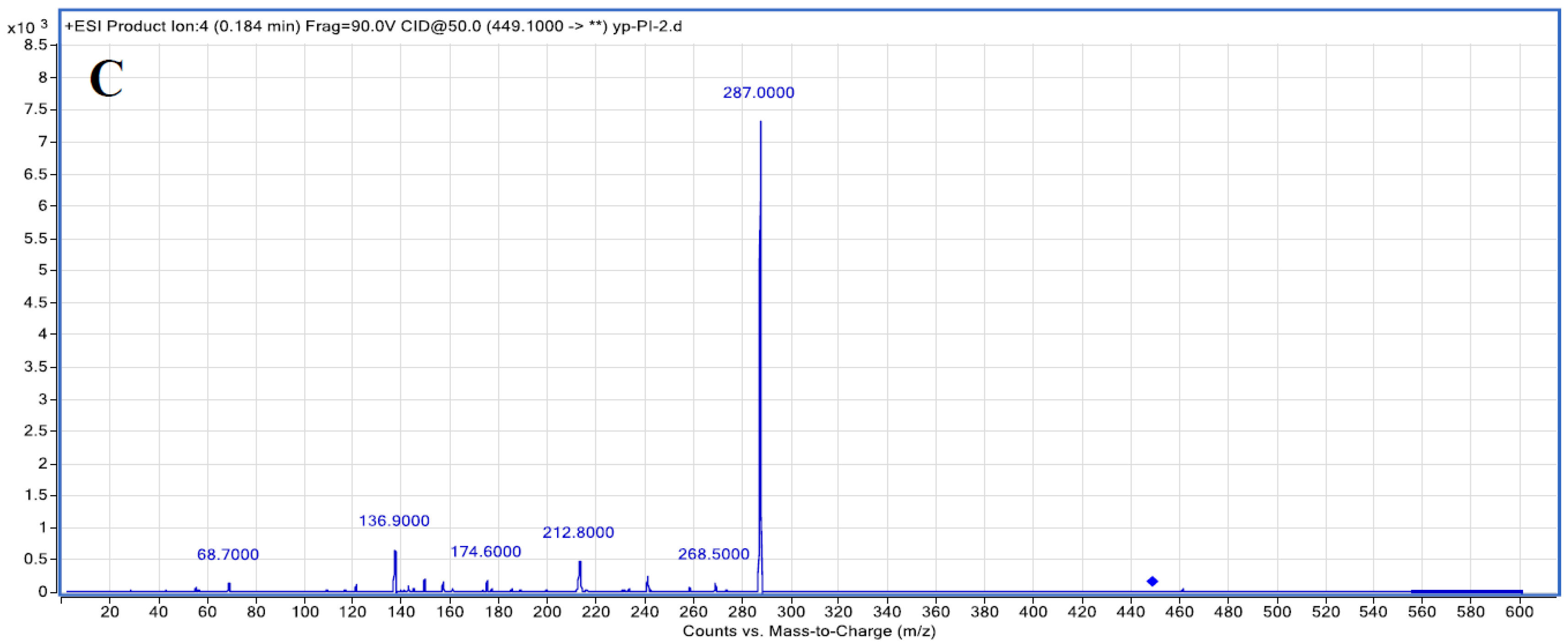
3.1. Purification of C3G Extracted from Chinese Bayberry
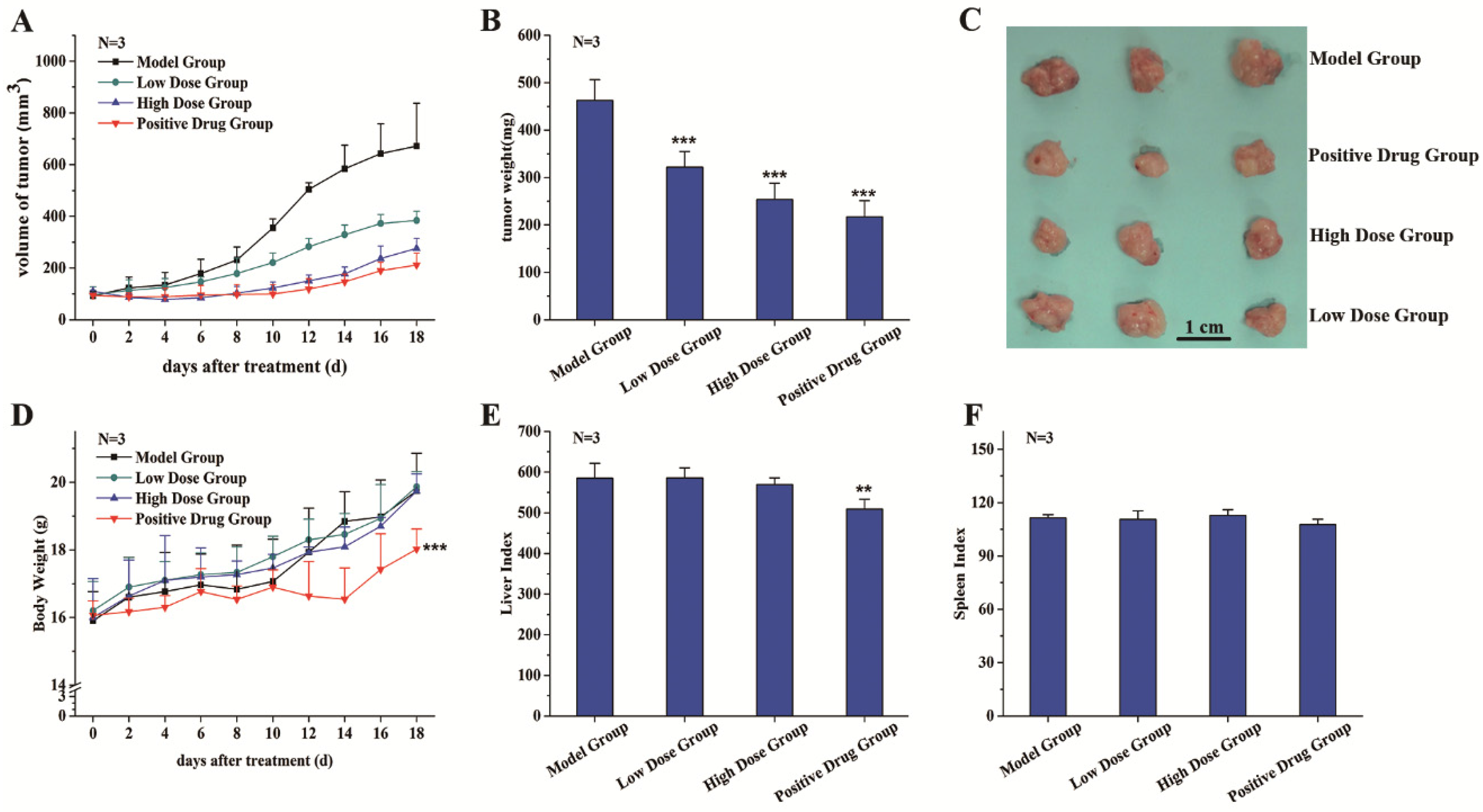
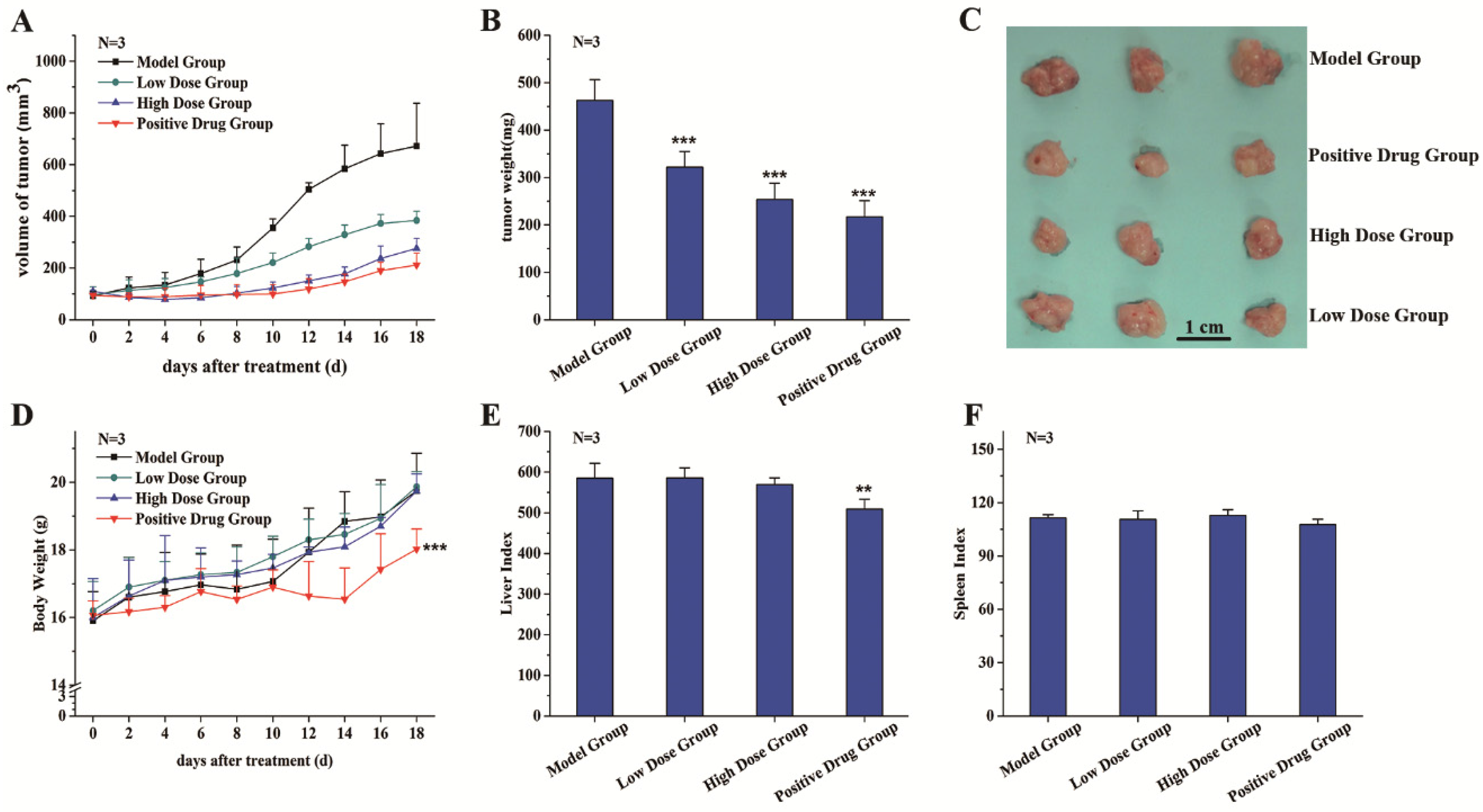
3.2. C3G Suppressed the Growth of SGC-7901 Tumor Xenografts
3.3. C3G Inhibited the Cell Cycle of SGC-7901 Tumor Xenografts
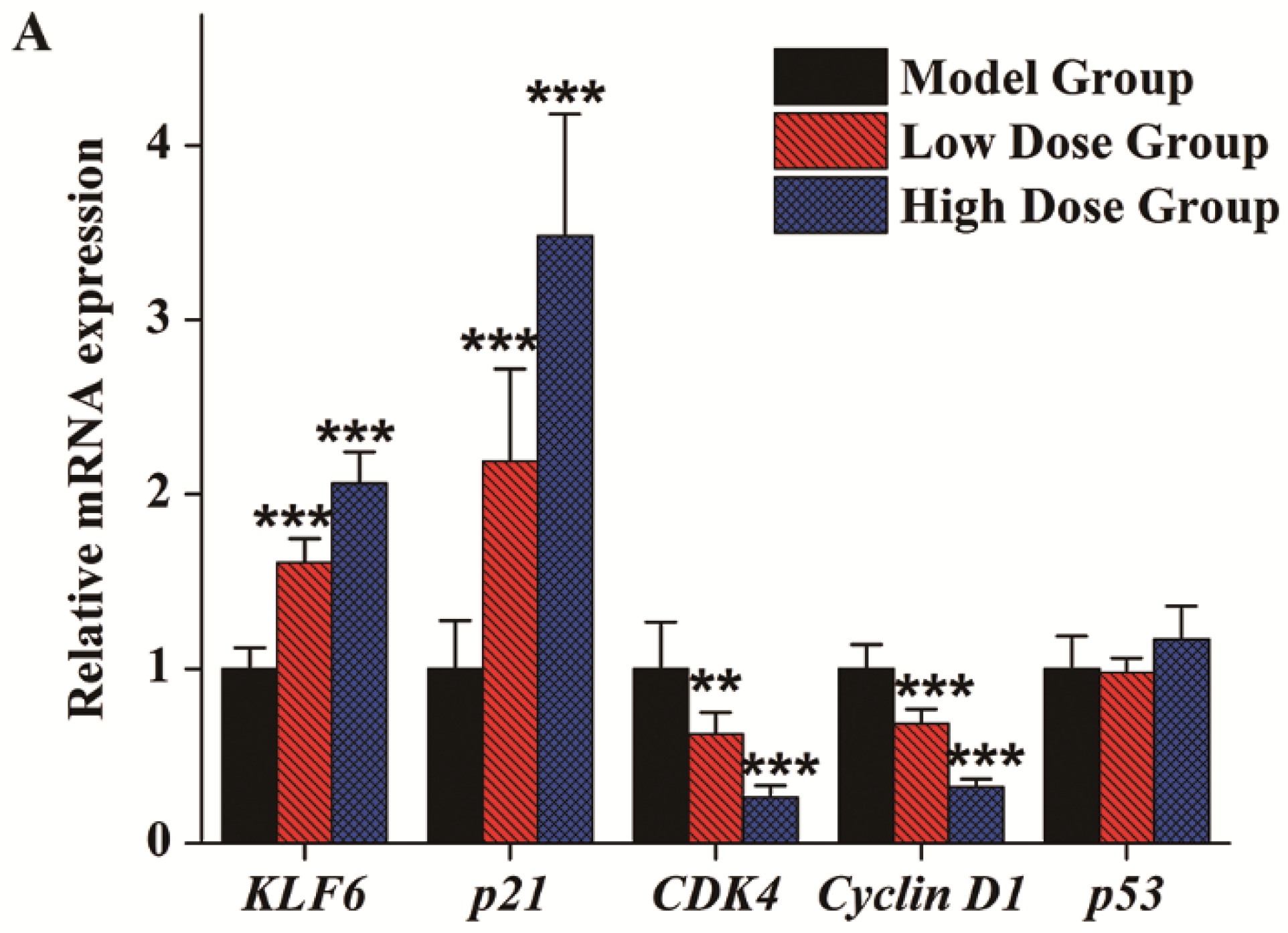
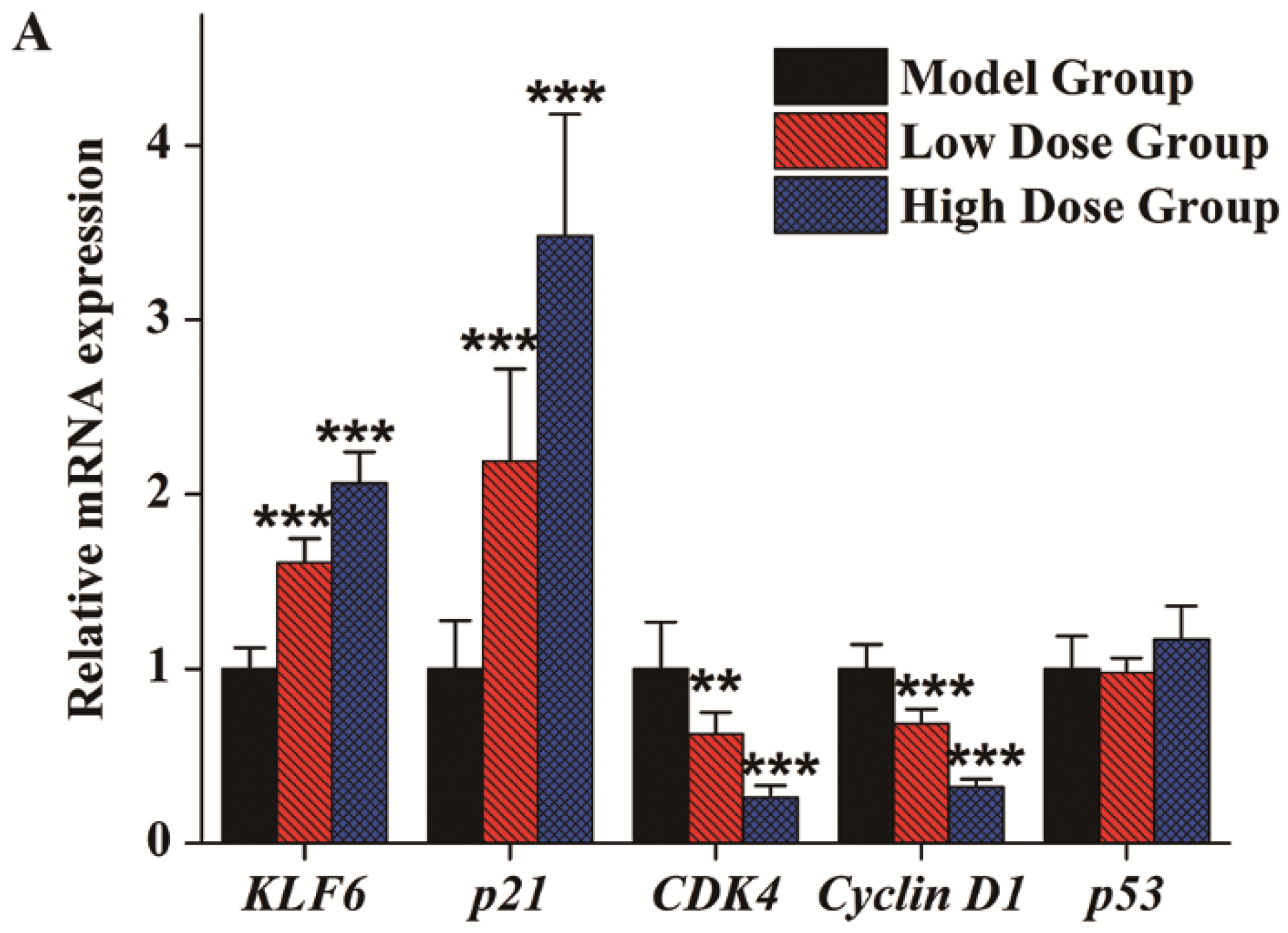
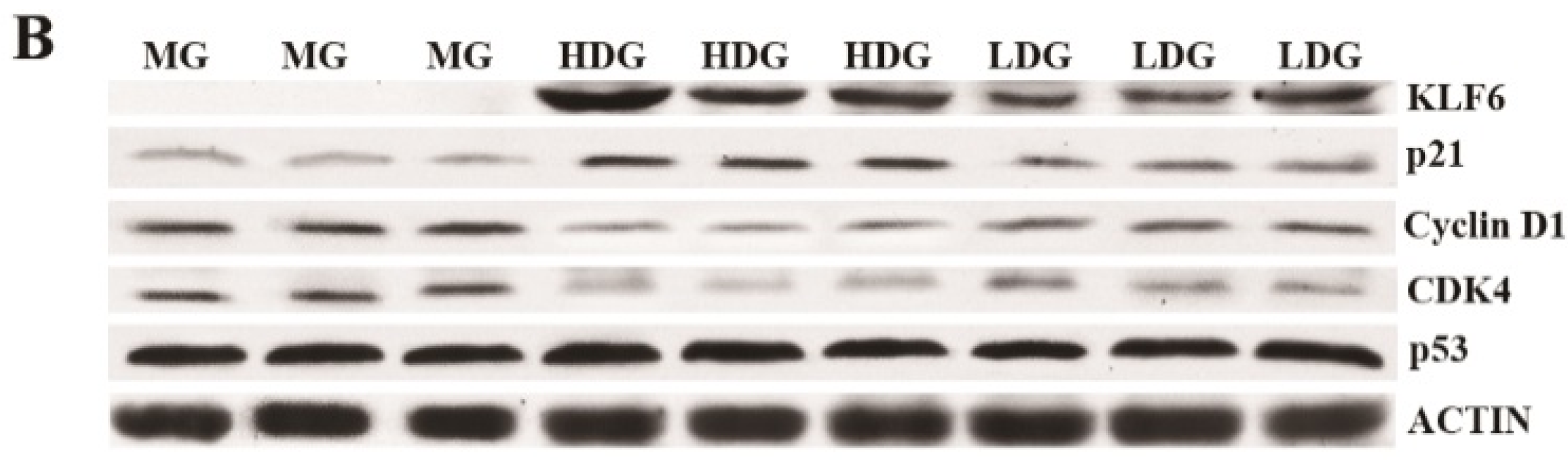
3.4. Gene Expression Analysis
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| C3G | Cyanidin-3-glucoside |
| CDK4 | Cyclin-dependent kinase 4 |
| RIPA | Radio Immunoprecipitation Assay |
| HPLC | High Performance Liquid Chromatography |
| LC-MS | Liquid Chromatograph-Mass Spectrometer |
| 1H-NMR | Hydrogen-Nuclear Magnetic Resonance |
| PBS | Phosphate Buffer Saline |
| PMSF | Phenylmethanesulfonyl Fluoride |
| SDS | Sodium Dodecyl Sulfate |
| PVDF | Polyvinylidene Fluoride |
| KLF6 | Krueppel-Like Factor 6 |
| LSD | Least Significant Difference |
| CSCs | Cancer stem cells |
References
- Bräunlich, M.; Slimestad, R.; Wangensteen, H.; Brede, C.; Malterud, K.E.; Barsett, H. Extracts, anthocyanins and procyanidins from aronia melanocarpa as radical scavengers and enzyme inhibitors. Nutrients 2013, 5, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.N.; Chu, S.C.; Chiou, H.L.; Kuo, W.H.; Chiang, C.L.; Hsieh, Y.S. Mulberry anthocyanins, cyanidin 3-rutinoside and cyanidin 3-glucoside, exhibited an inhibitory effect on the migration and invasion of a human lung cancer cell line. Cancer Lett. 2006, 235, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Meydani, M.; Hasan, S.T. Dietary polyphenols and obesity. Nutrients 2010, 2, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lv, Q.; Jia, S.; Chen, Y.; Sun, C.; Li, X.; Chen, K. Effects of flavonoids-rich chinese bayberry (Morella rubra Sieb. et Zucc.) fruits extract on regulating glucose and lipids metabolism in diabetic KK-Ay mice. Food Funct. 2016, 7, 3130–3140. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zheng, Y.; Chen, Q.; Tang, X.; Jiang, M.; Zhang, J.; Li, X.; Chen, K. Purification and anti-tumour activity of cyanidin-3-O-glucoside from Chinese bayberry fruit. Food Chem. 2012, 131, 1287–1294. [Google Scholar] [CrossRef]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zheng, R.; Baade, P.D.; Zhang, S.; Zeng, H.; Bray, F.; Jemal, A.; Yu, X.Q.; He, J. Cancer statistics in China, 2015. CA Cancer J. Clin. 2016, 66, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Russell, R.M. Nutrition and gastric cancer risk: An update. Nutr. Rev. 2008, 66, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Wu, L.; Guan, W. Dietary nitrates, nitrites, and nitrosamines intake and the risk of gastric cancer: A meta-analysis. Nutrients 2015, 7, 9872–9895. [Google Scholar] [CrossRef] [PubMed]
- Lunet, N.; Lacerda-Vieira, A.; Barros, H. Fruit and vegetables consumption and gastric cancer: A systematic review and meta-analysis of cohort studies. Nutr. Cancer 2005, 53, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nouraie, M.; Pietinen, P.; Kamangar, F.; Dawsey, S.M.; Abnet, C.C.; Albanes, D.; Virtamo, J.; Taylor, P.R. Fruits, vegetables, and antioxidants and risk of gastric cancer among male smokers. Cancer Epidem. Biomark. 2005, 14, 2087–2092. [Google Scholar] [CrossRef] [PubMed]
- González, C.A.; Pera, G.; Agudo, A.; Bueno-de-Mesquita, H.B.; Ceroti, M.; Boeing, H.; Schulz, M.; Del Giudice, G.; Plebani, M.; Carneiro, F. Fruit and vegetable intake and the risk of stomach and oesophagus adenocarcinoma in the European prospective investigation into cancer and nutrition (EPIC–EURGAST). Int. J. Cancer 2006, 118, 2559–2566. [Google Scholar] [CrossRef] [PubMed]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P. Polyphenols and human health: Prevention of disease and mechanisms of action. Nutrients 2010, 2, 1106–1131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, X.; Zheng, J.; Wang, G.; Sun, C.; Ferguson, I.B.; Chen, K. Bioactive components and antioxidant capacity of Chinese bayberry (Myrica rubra Sieb. and Zucc.) fruit in relation to fruit maturity and postharvest storage. Eur. Food Res. Technol. 2008, 227, 1091–1097. [Google Scholar] [CrossRef]
- Sorrenti, V.; Vanella, L.; Acquaviva, R.; Cardile, V.; Giofre, S.; Di Giacomo, C. Cyanidin induces apoptosis and differentiation in prostate cancer cells. Int. J. Oncol. 2015, 47, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Fukamachi, K.; Imada, T.; Ohshima, Y.; Xu, J.; Tsuda, H. Purple corn color suppresses ras protein level and inhibits 7,12-dimethylbenz[a]anthracene-induced mammary carcinogenesis in the rat. Cancer Sci. 2008, 99, 1841–1846. [Google Scholar] [PubMed]
- Shih, P.H.; Yeh, C.T.; Yen, G.C. Effects of anthocyanidin on the inhibition of proliferation and induction of apoptosis in human gastric adenocarcinoma cells. Food Chem. Toxicol. 2005, 43, 1557–1566. [Google Scholar] [CrossRef] [PubMed]
- Malik, M.; Zhao, C.W.; Schoene, N.; Guisti, M.M.; Moyer, M.P.; Magnuson, B.A. Anthocyanin-rich extract from Aronia meloncarpa E induces a cell cycle block in colon cancer but not normal colonic cells. Nutr. Cancer 2003, 46, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Bower, K.A.; Wang, S.; Frank, J.A.; Chen, G.; Ding, M.; Wang, S.; Shi, X.; Ke, Z.; Luo, J. Cyanidin-3-glucoside inhibits ethanol-induced invasion of breast cancer cells overexpressing ErbB2. Mol. Cancer 2010, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Narla, G.; Heath, K.E.; Reeves, H.L.; Li, D.; Giono, L.E.; Kimmelman, A.C.; Glucksman, M.J.; Narla, J.; Eng, F.J.; Chan, A.M.; et al. KLF6, a candidate tumor suppressor gene mutated in prostate cancer. Science 2001, 294, 2563–2566. [Google Scholar] [CrossRef] [PubMed]
- Sangodkar, J.; Shi, J.; DiFeo, A.; Schwartz, R.; Bromberg, R.; Choudhri, A.; McClinch, K.; Hatami, R.; Scheer, E.; Kremer-Tal, S. Functional role of the KLF6 tumour suppressor gene in gastric cancer. Eur. J. Cancer 2009, 45, 666–676. [Google Scholar] [CrossRef] [PubMed]
- Miyaki, M.; Yamaguchi, T.; Iijima, T.; Funata, N.; Mori, T. Difference in the role of loss of heterozygosity at 10p15 (KLF6 locus) in colorectal carcinogenesis between sporadic and familial adenomatous polyposis and hereditary nonpolyposis colorectal cancer patients. Oncol. Basel 2007, 71, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Mukai, S.; Hiyama, T.; Tanaka, S.; Yoshihara, M.; Arihiro, K.; Chayama, K. Involvement of kruppel-like factor 6 (KLF6) mutation in the development of nonpolypoid colorectal carcinoma. World J. Gastroenterol. 2007, 13, 3932–3938. [Google Scholar] [CrossRef] [PubMed]
- DiFeo, A.; Narla, G.; Hirshfeld, J.; Camacho-Vanegas, O.; Narla, J.; Rose, S.L.; Kalir, T.; Yao, S.; Levine, A.; Birrer, M.J. Roles of KLF6 and KLF6-SV1 in ovarian cancer progression and intraperitoneal dissemination. Clin. Cancer Res. 2006, 12, 3730–3739. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Kim, C.J.; Cho, Y.G.; Kim, S.Y.; Nam, S.W.; Lee, S.H.; Yoo, N.J.; Lee, J.Y.; Park, W.S. Genetic and epigenetic alterations of the KLF6 gene in hepatocellular carcinoma. J. Gastroenterol. Hepatol. 2006, 21, 1286–1289. [Google Scholar] [CrossRef] [PubMed]
- Yea, S.; Narla, G.; Zhao, X. Ras promotes growth by alternative splicing-mediated inactivation of the KLF6 tumor suppressor in hepatocellular carcinoma. Gastroenterology 2008, 135, 326. [Google Scholar] [CrossRef] [PubMed]
- Ozdemir, F.; Koksal, M.; Ozmen, V.; Aydin, I.; Buyru, N. Mutations and kruppel-like factor 6 (KLF6) expression levels in breast cancer. Tumor Biol. 2014, 35, 5219–5225. [Google Scholar] [CrossRef] [PubMed]
- Benzeno, S.; Narla, G.; Allina, J.; Cheng, G.Z.; Reeves, H.L.; Banck, M.S.; Odin, J.A.; Diehl, J.A.; Germain, D.; Friedman, S.L. Cyclin-dependent kinase inhibition by the KLF6 tumor suppressor protein through interaction with Cyclin D1. Cancer Res. 2004, 64, 3885–3891. [Google Scholar] [CrossRef] [PubMed]
- Schieber, A.; Keller, P.; Carle, R. Determination of phenolic acids and flavonoids of apple and pear by high-performance liquid chromatography. J. Chromatogr. A 2001, 910, 265–273. [Google Scholar] [CrossRef]
- Schuh, R.; Aicher, W.; Gaul, U.; Côte, S.; Preiss, A.; Maier, D.; Seifert, E.; Nauber, U.; Schröder, C.; Kemler, R. A conserved family of nuclear proteins containing structural elements of the finger protein encoded by krüppel, a drosophila segmentation gene. Cell 1986, 47, 1025–1032. [Google Scholar] [CrossRef]
- Pearson, R.; Fleetwood, J.; Eaton, S.; Crossley, M.; Bao, S. Krüppel-like transcription factors: A functional family. Int. J. Biochem. Cell Biol. 2008, 40, 1996–2001. [Google Scholar] [CrossRef] [PubMed]
- Koritschoner, N.P.; Bocco, J.L.; Panzetta-Dutari, G.M.; Dumur, C.I.; Flury, A.; Patrito, L.C. A novel human zinc finger protein that interacts with the core promoter element of a tata box-less gene. J. Biol. Chem. 1997, 272, 9573–9580. [Google Scholar] [PubMed]
- Reddy, M.K.; Alexander-Lindo, R.L.; Nair, M.G. Relative inhibition of lipid peroxidation, cyclooxygenase enzymes, and human tumor cell proliferation by natural food colors. J. Agric. Food Chem. 2005, 53, 9268–9273. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.N.; Chu, S.C.; Chiou, H.L.; Chiang, C.L.; Yang, S.F.; Hsieh, Y.S. Cyanidin 3-glucoside and peonidin 3-glucoside inhibit tumor cell growth and induce apoptosis in vitro and suppress tumor growth in vivo. Nutr. Cancer 2005, 53, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.P.; Chang, Y.C.; Wu, C.H.; Hung, C.N.; Wang, C.J. Anthocyanin-rich mulberry extract inhibit the gastric cancer cell growth in vitro and xenograft mice by inducing signals of p38/p53 and c-jun. Food Chem. 2011, 129, 1703–1709. [Google Scholar] [CrossRef]
- Charepalli, V.; Reddivari, L.; Radhakrishnan, S.; Vadde, R.; Agarwal, R.; Vanamala, J.K.P. Anthocyanin-containing purple-fleshed potatoes suppress colon tumorigenesis via elimination of colon cancer stem cells. J. Nutr. Biochem. 2015, 26, 1641–1649. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.P.; Lee, Y.K.; Lee, W.S.; Park, O.J.; Kim, Y.M. The involvement of AMPK/GSK3-beta signals in the control of metastasis and proliferation in hepato-carcinoma cells treated with anthocyanins extracted from Korea wild berry Meoru. BMC Complement. Altern. Med. 2014, 14, 109. [Google Scholar]
- Yang, X.; Luo, E.; Liu, X.; Han, B.; Yu, X.; Peng, X. Delphinidin-3-glucoside suppresses breast carcinogenesis by inactivating the Akt/HOTAIR signaling pathway. BMC Cancer 2016, 16, 423. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, N.; Talcott, S.; Safe, S.; Mertens-Talcott, S.U. Cytotoxicity of pomegranate polyphenolics in breast cancer cells in vitro and vivo: Potential role of miRNA-27a and miRNA-155 in cell survival and inflammation. Breast Cancer Res. Treat. 2012, 136, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.S.; Hecht, S.S.; Carmella, S.G.; Yu, N.; Larue, B.; Henry, C.; McIntyre, C.; Rocha, C.; Lechner, J.F.; Stoner, G.D. Anthocyanins in black raspberries prevent esophageal tumors in rats. Cancer Prev. Res. (Phila) 2009, 2, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.C.; Huang, H.P.; Chang, Y.C.; Wang, C.J. An anthocyanin-rich extract from hibiscus sabdariffa linnaeus inhibits N-nitrosomethylurea-induced leukemia in rats. J. Agric. Food Chem. 2014, 62, 1572–1580. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Xu, J.T.; Kim, J.; Chen, T.Y.; Su, X.Y.; Standard, J.; Carey, E.; Griffin, J.; Herndon, B.; Katz, B.; et al. Role of anthocyanin-enriched purple-fleshed sweet potato p40 in colorectal cancer prevention. Mol. Nutr. Food Res. 2013, 57, 1908–1917. [Google Scholar] [CrossRef] [PubMed]
- Duthie, S.J.; Gardner, P.T.; Morrice, P.C.; Wood, S.G.; Pirie, L.; Bestwick, C.C.; Milne, L.; Duthie, G.G. DNA stability and lipid peroxidation in vitamin e-deficient rats in vivo and colon cells in vitro—Modulation by the dietary anthocyanin, cyanidin-3-glycoside. Eur. J. Nutr. 2005, 44, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Yu, B.; Yu, X.P.; Yi, L.; Chen, C.Y.; Mi, M.T.; Ling, W.H. Anticancer activities of an anthocyanin-rich extract from black rice against breast cancer cells in vitro and in vivo. Nutr. Cancer 2010, 62, 1128–1136. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′ to 3′) | Reverse Primer (3′ to 5′) |
|---|---|---|
| p53 | AGGCCTTGGAACTCAAGGAT | CCCTTTTTGGACTTCAGGTG |
| KLF6 | GACAGCTCCGAGGAACTTTCT | CACGCAACCCCACAGTTGA |
| P21 | TGGAGACTCTCAGGGTCGAAAA | GGCGTTTGGAGTGGTAGAAATCT |
| CDK4 | ACAGTTCGTGAGGTGGCTTTA | TCAGATCCTTGATCGTTTCG |
| Cyclin D1 | GAACACGGCTCACGCTTACC | GCCCAGACCCTCAGACTTGC |
| β-actin | TGACGTGGACATCCGCAAAG | CTGGAAGGTGGACAGCGAGG |
| Cyanidin-3-Glucoside | 1H |
|---|---|
| Aglycone | |
| 4 | 9.02 (s) |
| 6 | 6.63 (d, J = 2.5 Hz) |
| 8 | 7.06 (d, J = 2 Hz) |
| 2′ | 8.08 (s) |
| 5′ | 7.10 (s) |
| 6′ | 8.26 (s) |
| Glucose | |
| 1″ | 5.30 (d, J = 16 Hz) |
| 2″ | 3.70 (m) |
| 3″ | 3.66 (br, s) |
| 4″ | 3.54 (m) |
| 5″ | 3.63 (s) |
| 6″A | 4.02 (s) |
| 6″B | 3.83 (s) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, X.-n.; Xie, W.-h.; Zheng, Y.-x.; Cao, J.-p.; Cao, P.-r.; Chen, Q.-j.; Li, X.; Sun, C.-d. The Growth of SGC-7901 Tumor Xenografts Was Suppressed by Chinese Bayberry Anthocyanin Extract through Upregulating KLF6 Gene Expression. Nutrients 2016, 8, 599. https://doi.org/10.3390/nu8100599
Wang Y, Zhang X-n, Xie W-h, Zheng Y-x, Cao J-p, Cao P-r, Chen Q-j, Li X, Sun C-d. The Growth of SGC-7901 Tumor Xenografts Was Suppressed by Chinese Bayberry Anthocyanin Extract through Upregulating KLF6 Gene Expression. Nutrients. 2016; 8(10):599. https://doi.org/10.3390/nu8100599
Chicago/Turabian StyleWang, Yue, Xia-nan Zhang, Wen-hua Xie, Yi-xiong Zheng, Jin-ping Cao, Pei-rang Cao, Qing-jun Chen, Xian Li, and Chong-de Sun. 2016. "The Growth of SGC-7901 Tumor Xenografts Was Suppressed by Chinese Bayberry Anthocyanin Extract through Upregulating KLF6 Gene Expression" Nutrients 8, no. 10: 599. https://doi.org/10.3390/nu8100599