Effect of the Novel Polysaccharide PolyGlycopleX® on Short-Chain Fatty Acid Production in a Computer-Controlled in Vitro Model of the Human Large Intestine

 ,
, 
Abstract
:1. Introduction
2. Experimental Section
2.1. Test and Control Fiber
2.2. Intestinal Conditions of TIM-2
2.3. Addition of Test and Control Fibers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium Component | Day 1/Day 2/Day 3 | Day 1/Day 2/Day 3 | Day 1/Day 2/Day 3 |
|---|---|---|---|
| Casein | 2.6 g | 2.6 g | 2.6 g |
| Tween 80 | 1.9 g | 1.9 g | 1.9 g |
| Bactopepton | 2.6 g | 2.6 g | 2.6 g |
| Ox-bile | 42 mg | 42 mg | 42 mg |
| K2HPO4·3H2O | 282 mg | 282 mg | 282 mg |
| NaCl | 0.5 g | 0.5 g | 0.5 g |
| FeSO4·7H2O | 0.54 mg | 0.54 mg | 0.54 mg |
| MgSO4·7H2O | 42 mg | 42 mg | 42 mg |
| CaCl2·2H2O | 48 mg | 48 mg | 48 mg |
| Haemin | 1.2 mg | 1.2 mg | 1.2 mg |
| Cysteine·HCl | 18 mg | 18 mg | 18 mg |
| Vitamins * | 90 mg | 90 mg | 90 mg |
| FOS | 4.0 g/2.5 g/3.5 g | ||
| PGX® | 4.0 g/2.5 g/3.5 g | ||
| Cellulose | 4.0 g/2.5 g/3.5 g |
2.4. Sampling from TIM-2
2.5. Fermentation and pH
2.6. Short-Chain and Branched Chain Fatty Acid Analysis
2.7. Statistical Analyses
3. Results
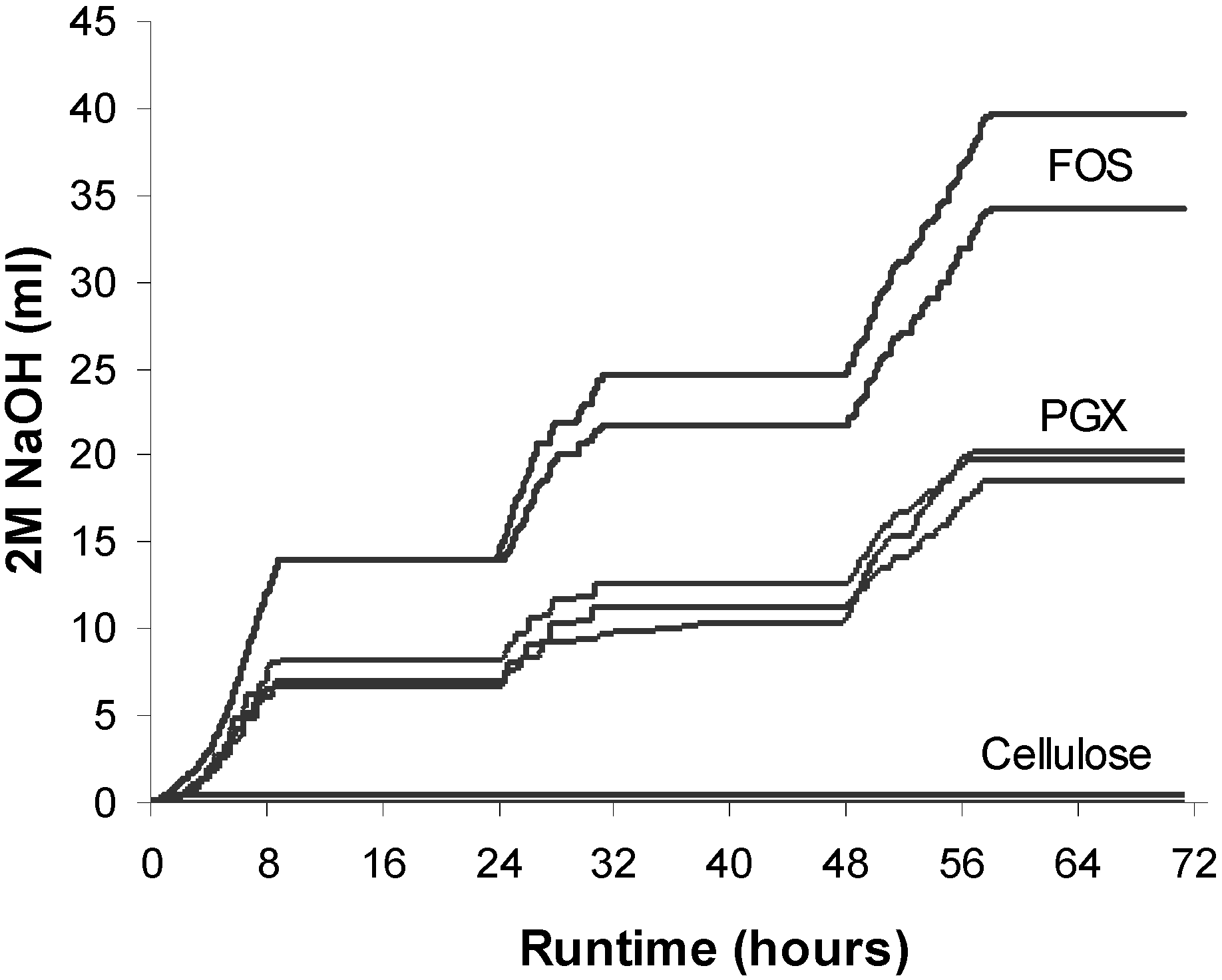
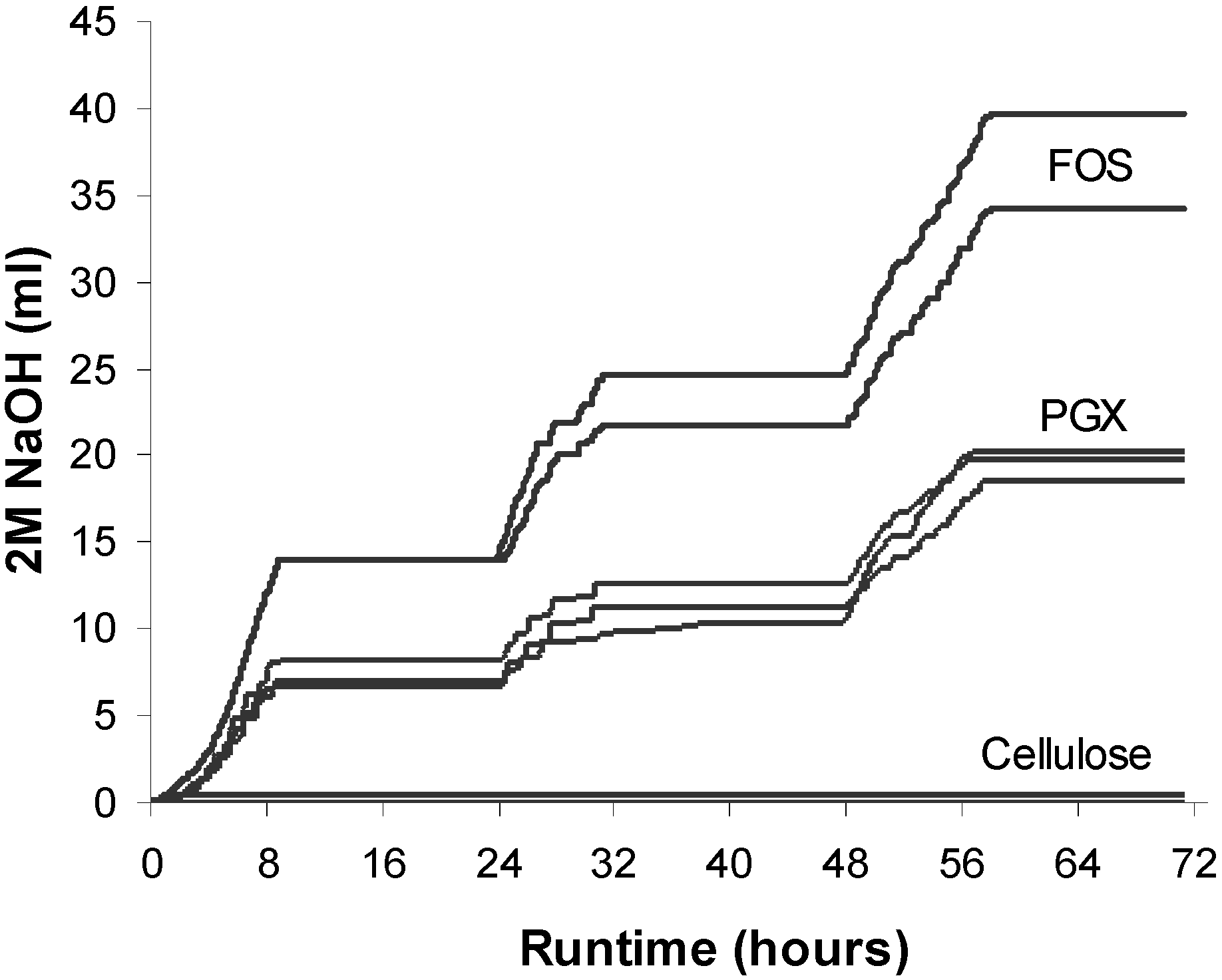
3.1. Sodium Hydroxide Usage
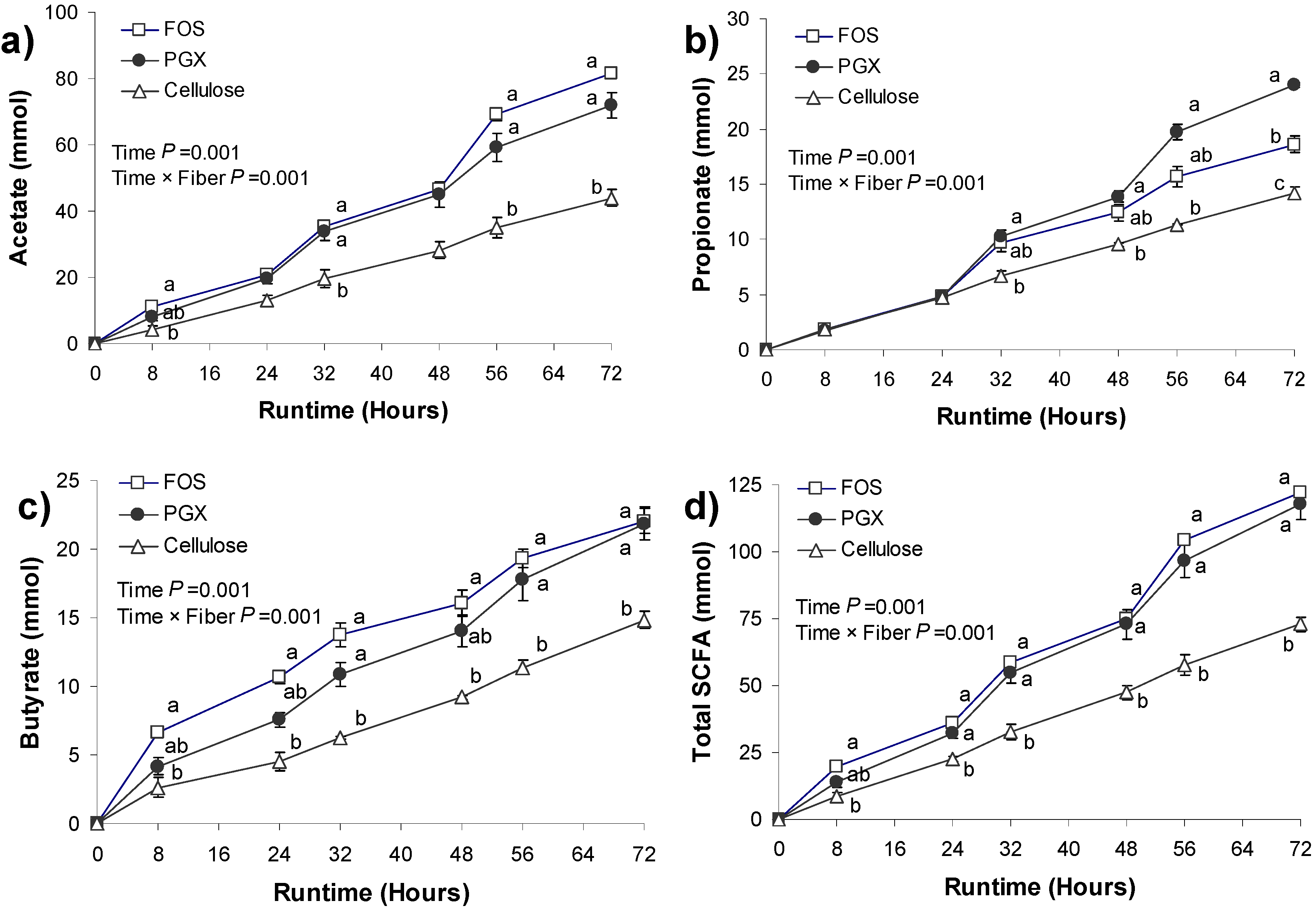
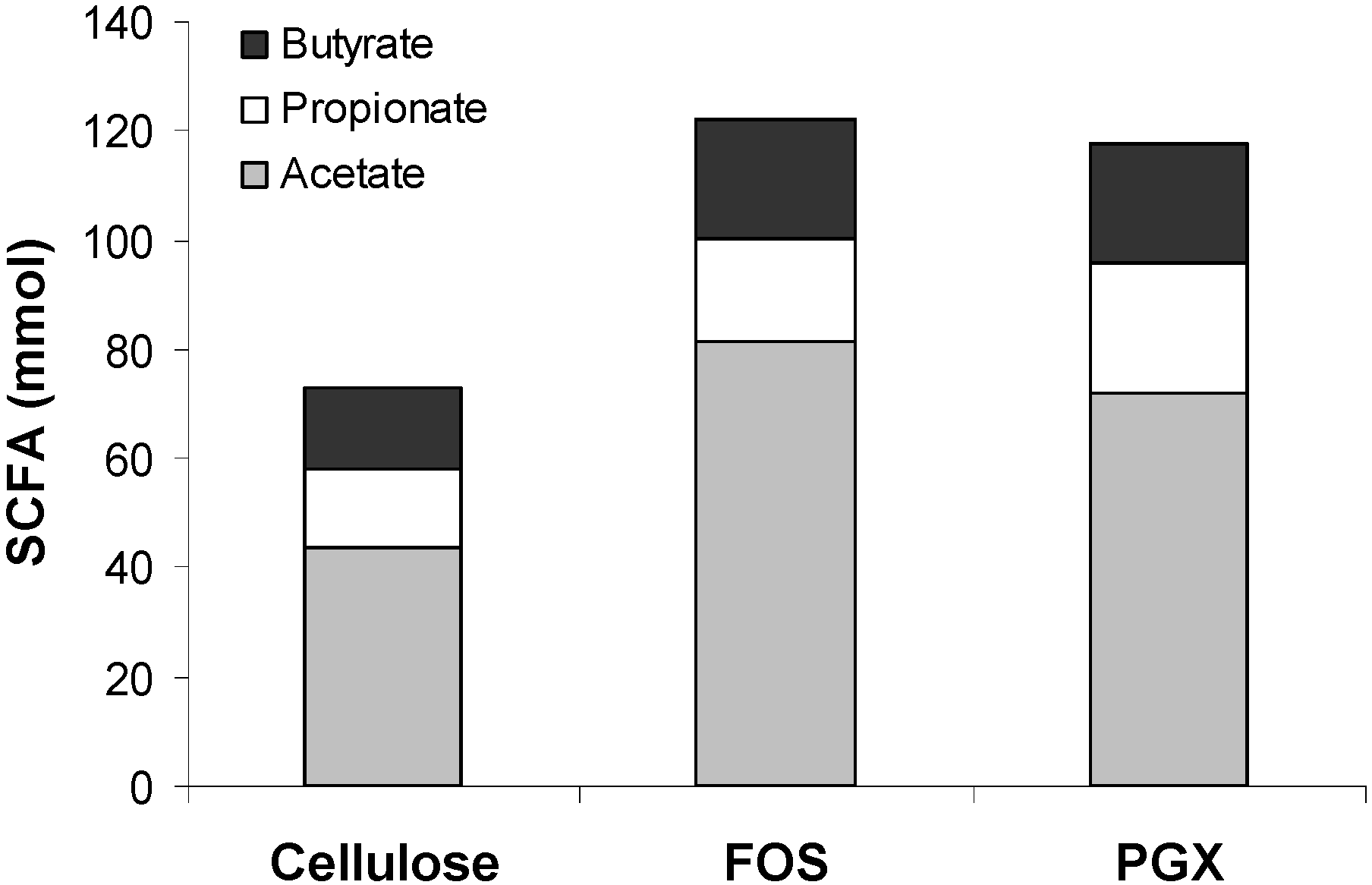
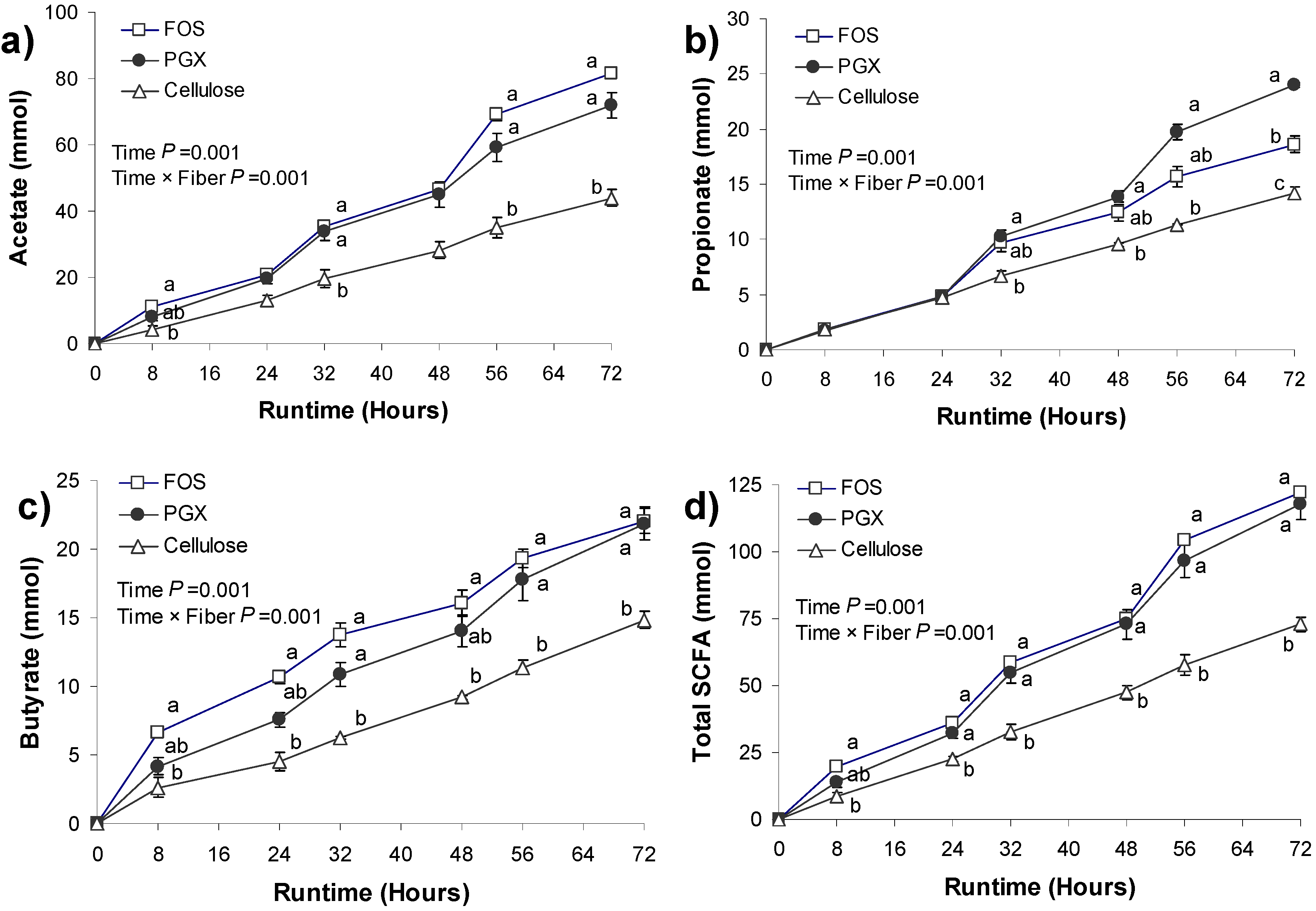
3.2. SCFA Production

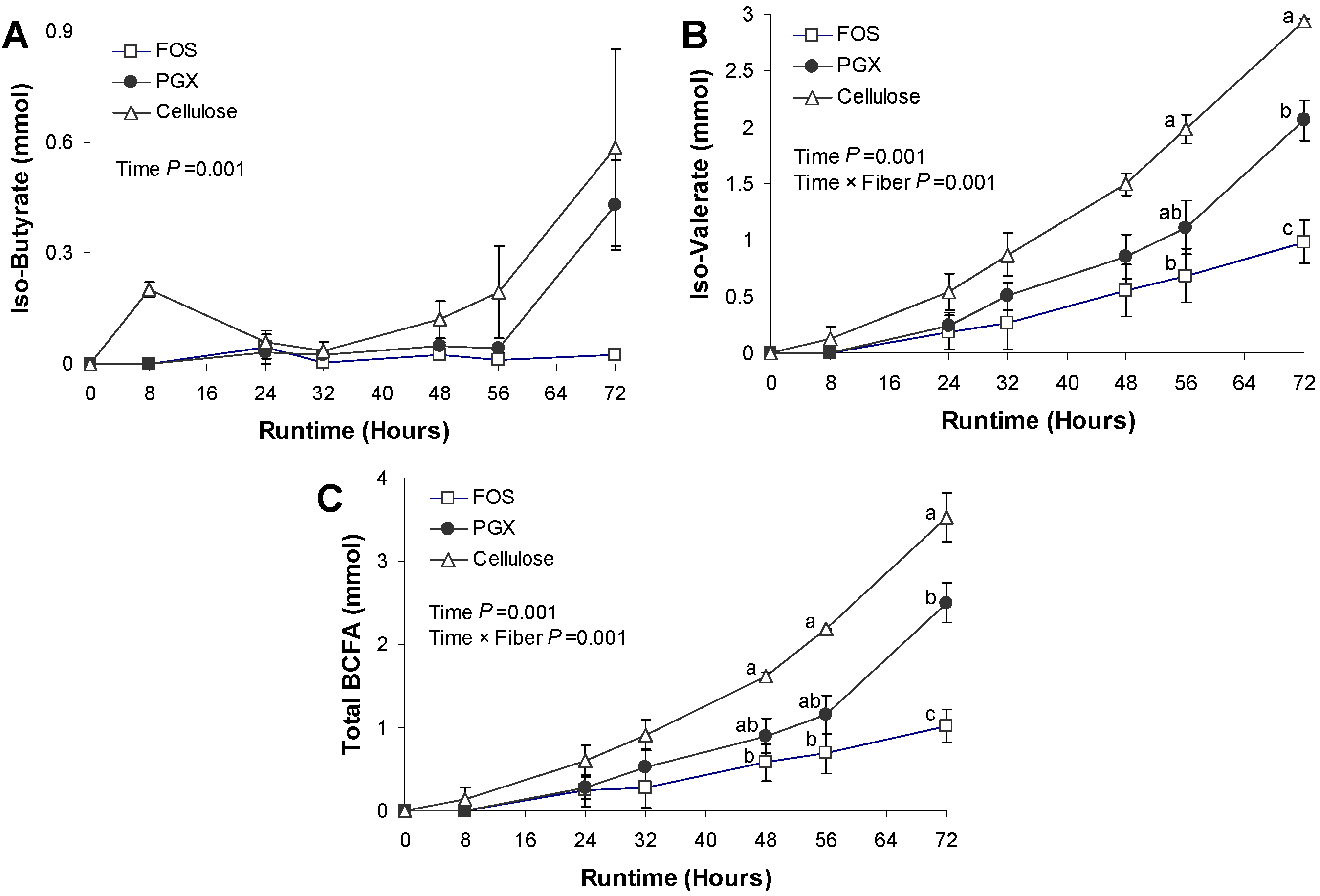
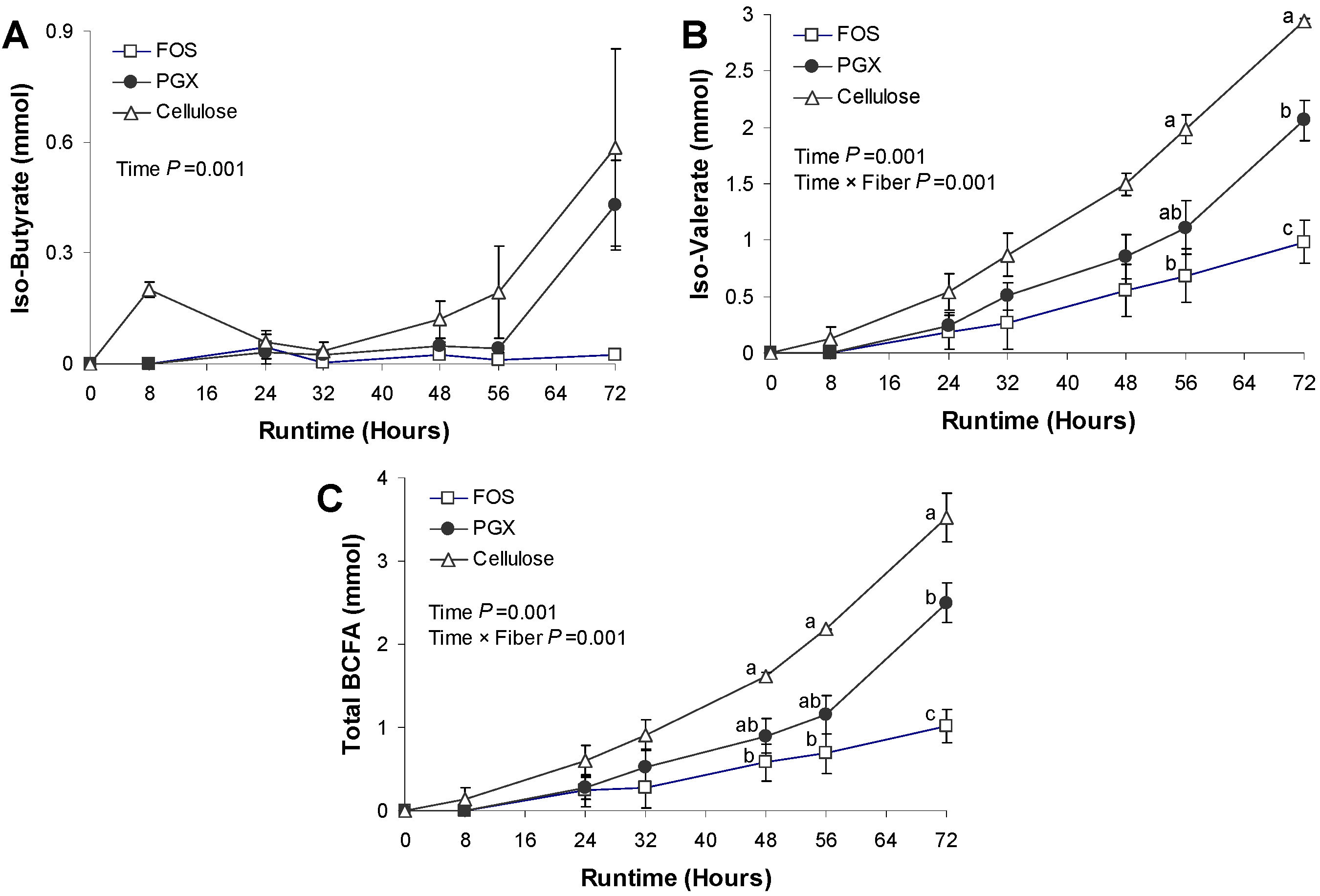
3.3. BCFA Production
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Wong, J.M.; de Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Bureau of Nutritional Sciences; Health Canada. Proposed Policy: Definition and Energy Value for Dietary Fibre; Health Canada: Ottawa, Canada, 2010. [Google Scholar]
- Reimer, R.A.; Pelletier, X.; Carabin, I.G.; Lyon, M.R.; Gahler, R.J.; Wood, S. Faecal short chain fatty acids in healthy subjects participating in a randomised controlled trial examining a soluble highly viscous polysaccharide versus control. J. Hum. Nutr. Diet. 2012, 25, 373–377. [Google Scholar] [CrossRef]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar]
- Minekus, M.; Smeets-Peeters, M.; Bernalier, A.; Marol-Bonnin, S.; Havenaar, R.; Marteau, P.; Alric, M.; Fonty, G.; Huis in’t Veld, J.H. A computer-controlled system to simulate conditions of the large intestine with peristaltic mixing, water absorption and absorption of fermentation products. Appl. Microbiol. Biotechnol. 1999, 53, 108–114. [Google Scholar] [CrossRef]
- Venema, K.; van Nuenen, M.H.M.C.; van den Heuvel, E.G.; Pool, W.; van der Vossen, J.M.B.M. The effect of lactulose on the composition of the intestinal microbiota and short-chain fatty acid production in human volunteers and a computer controlled model of the proximal large intestine. Microb. Ecol. Health Dis. 2003, 15, 94–105. [Google Scholar]
- Van Nuenen, H.M.C.; Meyer, P.D.; Venema, K. The effect of various inulins and clostridium difficile on the metabolic activity of the human colonic microbiota in vitro. Micro. Ecol. Health Dis. 2003, 15, 137–144. [Google Scholar] [CrossRef]
- Macfarlane, S.; Macfarlane, G.T. Regulation of short-chain fatty acid production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef]
- Roberfroid, M.B. Inulin-Type Fructans: Functional Food Ingredients; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Macfarlane, G.T.; Macfarlane, S. Bacteria, colonic fermentation, and gastrointestinal health. J. AOAC Int. 2012, 95, 50–60. [Google Scholar]
- Kumar, V.; Sinha, A.K.; Makkar, H.P.; de Boeck, G.; Becker, K. Dietary roles of non-starch polysachharides in human nutrition: A review. Crit. Rev. Food Sci. Nutr. 2012, 52, 899–935. [Google Scholar] [CrossRef]
- Abdelhameed, A.S.; Ang, A.; Morris, G.A.; Smith, I.; Lawson, C.; Gahler, R.; Wood, S.; Harding, S.E. An analytical ultracentrifuge study on ternary mixtures of konjac glucomannan supplemented with sodium alginate and xanthan gum. Carbohydr. Polym. 2010, 81, 145–148. [Google Scholar] [CrossRef]
- Harding, S.E.; Smith, I.H.; Lawson, C.J.; Gahler, R.J.; Wood, S. Studies on macromolecular interactions in ternary mixtures of konjac glucomannan, xanthan gum and sodium alginate. Carbohydr. Polym. 2011, 83, 329–338. [Google Scholar] [CrossRef]
- Harding, S.E.; Almutairi, F.; Adams, G.G.; Morris, G.; Lawson, C.J.; Gahler, R.J.; Wood, S. Dilute solution viscometry studies on a therapeutic mixture of non-digestible carbohydrates. Int. J. Biotech. Wellness Ind. 2012, 1, 107–114. [Google Scholar]
- Maathuis, A.; Hoffman, A.; Evans, A.; Sanders, L.; Venema, K. The effect of the undigested fraction of maize products on the activity and composition of the microbiota determined in a dynamic in vitro model of the human proximal large intestine. J. Am. Coll. Nutr. 2009, 28, 657–666. [Google Scholar] [CrossRef]
- Gibson, G.R.; Cummings, J.H.; Macfarlane, G.T. Use of a three-stage continuous culture system to study the effect of mucin on dissimilatory sulphate reduction and methanogenesis by mixed populations of human gut bacteria. Appl. Environ. Microbiol. 1998, 54, 2750–2755. [Google Scholar]
- Reimer, R.A.; Grover, G.J.; Koetzner, L.; Gahler, R.; Juneja, P.; Lyon, M.R.; Wood, S. Sitagliptin reduces hyperglycemia and increases satiety hormone secretion more effectively when used with a novel polysaccharide in obese zucker rats. J. Nutr. 2012, 142, 1812–1820. [Google Scholar] [CrossRef]
- Reimer, R.A.; Grover, G.J.; Koetzner, L.; Gahler, R.; Lyon, M.; Wood, S. The soluble fiber complex polyglycoplex lowers serum triglycerides and reduces hepatic steatosis in high-sucrose-fed rats. Nutr. Res. 2011, 31, 296–301. [Google Scholar] [CrossRef]
- Grover, G.J.; Koetzner, L.; Wicks, J.; Gahler, R.; Lyon, M.R.; Reimer, R.A.; Wood, S. Effects of the soluble fiber complex polyglycoplex on glucose homeostasis and body weight in young zucker diabetic rats. Front. Pharmacol. 2011, 2, 47. [Google Scholar]
- Lyon, M.R.; Reichert, R.G. The effect of a novel viscous polysaccharide along with lifestyle changes on short-term weight loss and associated risk factors in overweight and obese adults: An observational retrospective clinical program analysis. Altern. Med. Rev. 2010, 15, 68–75. [Google Scholar]
- Martens, E.C.; Koropatkin, N.M.; Smith, T.J.; Gordon, J.I. Complex glycan catabolism by the human gut microbiota: The bacteroidetes sus-like paradigm. J. Biol. Chem. 2009, 284, 24673–24677. [Google Scholar]
- Macy, J.M.; Ljungdahl, L.G.; Gottschalk, G. Pathway of succinate and propionate formation in bacteroides fragilis. J. Bacteriol. 1978, 134, 84–91. [Google Scholar]
- Arora, T.; Sharma, R.; Frost, G. Propionate: Anti-obesity and satiety enhancing factor? Appetite 2011, 56, 511–515. [Google Scholar] [CrossRef]
- Bugaut, M.; Bentejac, M. Biological effects of short-chain fatty acids in nonruminant mammals. Annu. Rev. Nutr. 1993, 12, 217–241. [Google Scholar] [CrossRef]
- Beaulieu, K.E.; McBurney, M.I. Changes in pig serum lipids, nutrient digestibility and sterol excretion during cecal infusion of propionate. J. Nutr. 1992, 122, 241–245. [Google Scholar]
- Wright, R.S.; Anderson, J.W.; Bridges, S.R. Propionate inhibits hepatocyte lipid synthesis. Proc. Soc. Exp. Biol. Med. 1990, 195, 26–29. [Google Scholar] [CrossRef]
- Imaizumi, K.; Hirata, K.; Yasni, S.; Sugano, M. Propionate enhances synthesis and secretion of bile acids in primary cultured rat hepatocytes via succinyl coa. Biosci. Biotechnol. Biochem. 1992, 56, 1894–1896. [Google Scholar] [CrossRef]
- Al-Lahham, S.; Roelofsen, H.; Rezaee, F.; Weening, D.; Hoek, A.; Vonk, R.; Venema, K. Propionic acid affects immune status and metabolism in adipose tissue from overweight subjects. Eur. J. Clin. Investig. 2012, 42, 357–364. [Google Scholar] [CrossRef]
- Al-Lahham, S.H.; Roelofsen, H.; Priebe, M.; Weening, D.; Dijkstra, M.; Hoek, A.; Rezaee, F.; Venema, K.; Vonk, R.J. Regulation of adipokine production in human adipose tissue by propionic acid. Eur. J. Clin. Investig. 2010, 40, 401–407. [Google Scholar] [CrossRef]
- Wolever, T.M.; Schrade, K.B.; Vogt, J.A.; Tsihlias, E.B.; McBurney, M.I. Do colonic short-chain fatty acids contribute to the long-term adaptation of blood lipids in subjects with type 2 diabetes consuming a high-fiber diet? Am. J. Clin. Nutr. 2002, 75, 1023–1030. [Google Scholar]
- Cummings, J.H.; Macfarlane, G.T. The control and consequences of bacterial fermentation in the human colon. J. Appl. Bacteriol. 1991, 70, 443–459. [Google Scholar] [CrossRef]
- Nyangale, E.P.; Mottram, D.S.; Gibson, G.R. Gut microbial activity, implications for health and disease: The potential role of metabolite analysis. J. Proteome Res. 2012, 11, 5573–5585. [Google Scholar]
- Swanson, K.S.; Grieshop, C.M.; Flickinger, E.A.; Bauer, L.L.; Chow, J.; Wolf, B.W.; Garleb, K.A.; Fahey, G.C., Jr. Fructooligosaccharides and lactobacillus acidophilus modify gut microbial populations, total tract nutrient digestibilities and fecal protein catabolite concentrations in healthy adult dogs. J. Nutr. 2002, 132, 3721–3731. [Google Scholar]
- Slavin, J.L.; Brauer, P.M.; Marlett, J.A. Neutral detergent fiber, hemicellulose and cellulose digestibility in human subjects. J. Nutr. 1981, 111, 287–297. [Google Scholar]
- Birkett, A.; Muir, J.; Phillips, J.; Jones, G.; O’Dea, K. Resistant starch lowers fecal concentrations of ammonia and phenols in humans. Am. J. Clin. Nutr. 1996, 63, 766–772. [Google Scholar]
- Boler, B.M.; Serao, M.C.; Bauer, L.L.; Staeger, M.A.; Boileau, T.W.; Swanson, K.S.; Fahey, G.C., Jr. Digestive physiological outcomes related to polydextrose and soluble maize fibre consumption by healthy adult men. Br. J. Nutr. 2011, 106, 1861–1871. [Google Scholar]
- Duncan, S.H.; Louis, P.; Flint, H.J. Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microbiol. 2004, 70, 5810–5817. [Google Scholar] [CrossRef]
- Vinolo, M.A.R.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short-chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar]
- Cani, P.D.; Osto, M.; Geurts, L.; Everard, A. Involvement of gut microbiota in the development of low-grade inflammation and type 2 diabetes associated with obesity. Gut Microbes 2012, 3, 279–288. [Google Scholar]
- Cluny, N.L.; Reimer, R.A.; Sharkey, K.A. Cannabinoid signalling regulates inflammation and energy balance: The importance of the brain-gut axis. Pharmacol. Biochem. Behav. 2012, 97, 537–543. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Reimer, R.A.; Maathuis, A.J.H.; Venema, K.; Lyon, M.R.; Gahler, R.J.; Wood, S. Effect of the Novel Polysaccharide PolyGlycopleX® on Short-Chain Fatty Acid Production in a Computer-Controlled in Vitro Model of the Human Large Intestine. Nutrients 2014, 6, 1115-1127. https://doi.org/10.3390/nu6031115
Reimer RA, Maathuis AJH, Venema K, Lyon MR, Gahler RJ, Wood S. Effect of the Novel Polysaccharide PolyGlycopleX® on Short-Chain Fatty Acid Production in a Computer-Controlled in Vitro Model of the Human Large Intestine. Nutrients. 2014; 6(3):1115-1127. https://doi.org/10.3390/nu6031115
Chicago/Turabian StyleReimer, Raylene A., Annet J. H. Maathuis, Koen Venema, Michael R. Lyon, Roland J. Gahler, and Simon Wood. 2014. "Effect of the Novel Polysaccharide PolyGlycopleX® on Short-Chain Fatty Acid Production in a Computer-Controlled in Vitro Model of the Human Large Intestine" Nutrients 6, no. 3: 1115-1127. https://doi.org/10.3390/nu6031115