Protective Effect of Resveratrol on Biomarkers of Oxidative Stress Induced by Iron/Ascorbate in Mouse Spermatozoa


Abstract
:1. Introduction
2. Experimental Section
2.1. Chemicals
2.2. Animals
2.3. Experimental Design
2.4. Spermatozoa Quality
2.5. Mitochondrial Membrane Potential (Δψm)
2.6. ROS Level in Spermatozoa
2.7. SOD Activity in Spermatozoa
2.8. GPX Activity in Spermatozoa
2.9. TBARS Concentration in Spermatozoa
2.10. In Vitro Fertilization (IVF)
2.10.1. Egg Recovery
2.10.2. IVF Assay
2.11. Data Analysis
3. Results
3.1. Effect of RVT on Spermatozoa Quality

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sperm parameters | Control | RVT | Ferrous iron/ascorbate | RVT + Ferrous iron/ascorbate |
|---|---|---|---|---|
| Motility (%) | 73.3 ± 5.7 | 73.3 ± 2.8 | 6.66 ± 2.8 * | 53.3 ± 5.7 ** |
| Viability (%) | 65.0 ± 8.6 | 59.6 ± 6.1 | 14.8 ± 3.3 * | 30.5 ± 6.5 ** |
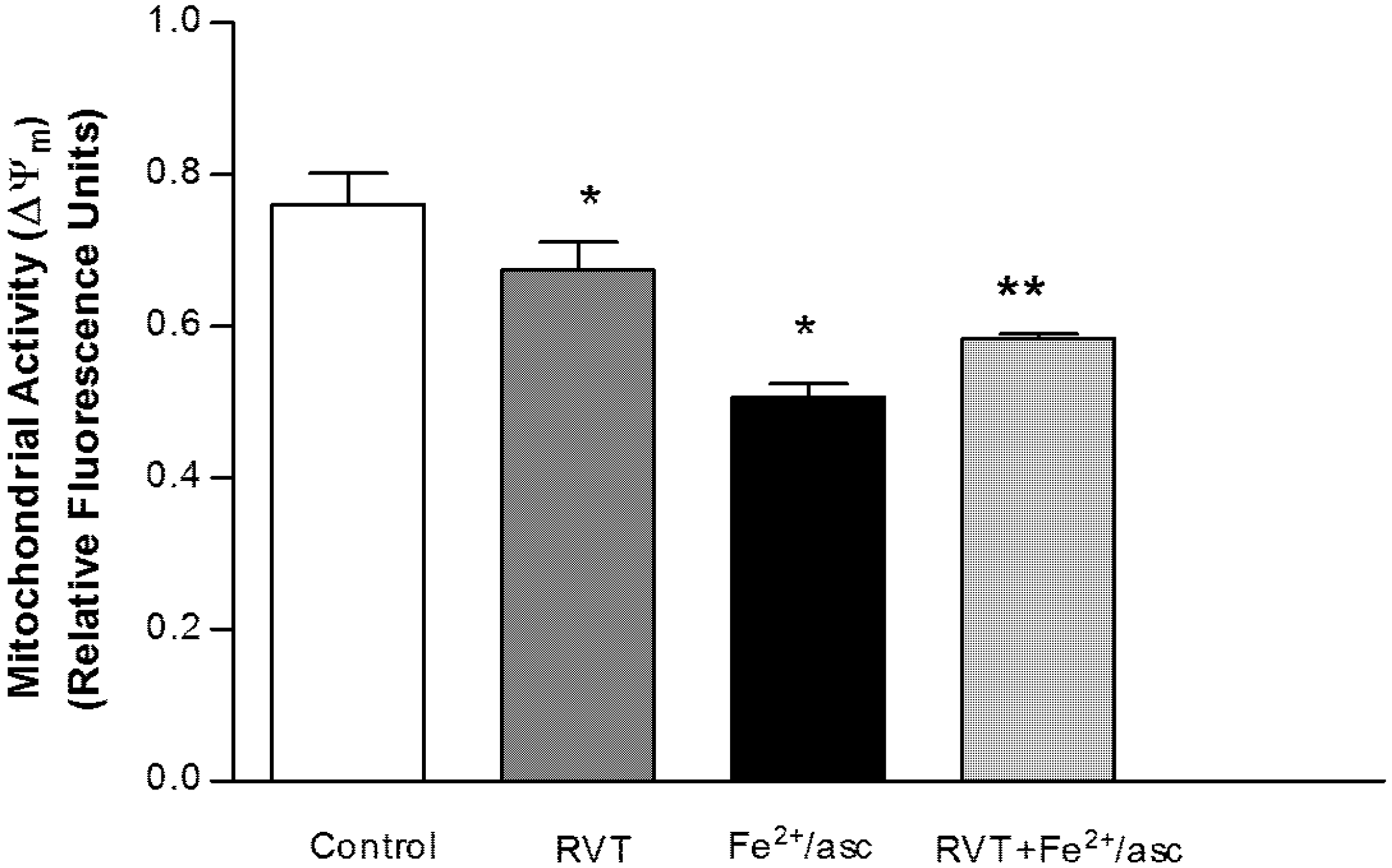
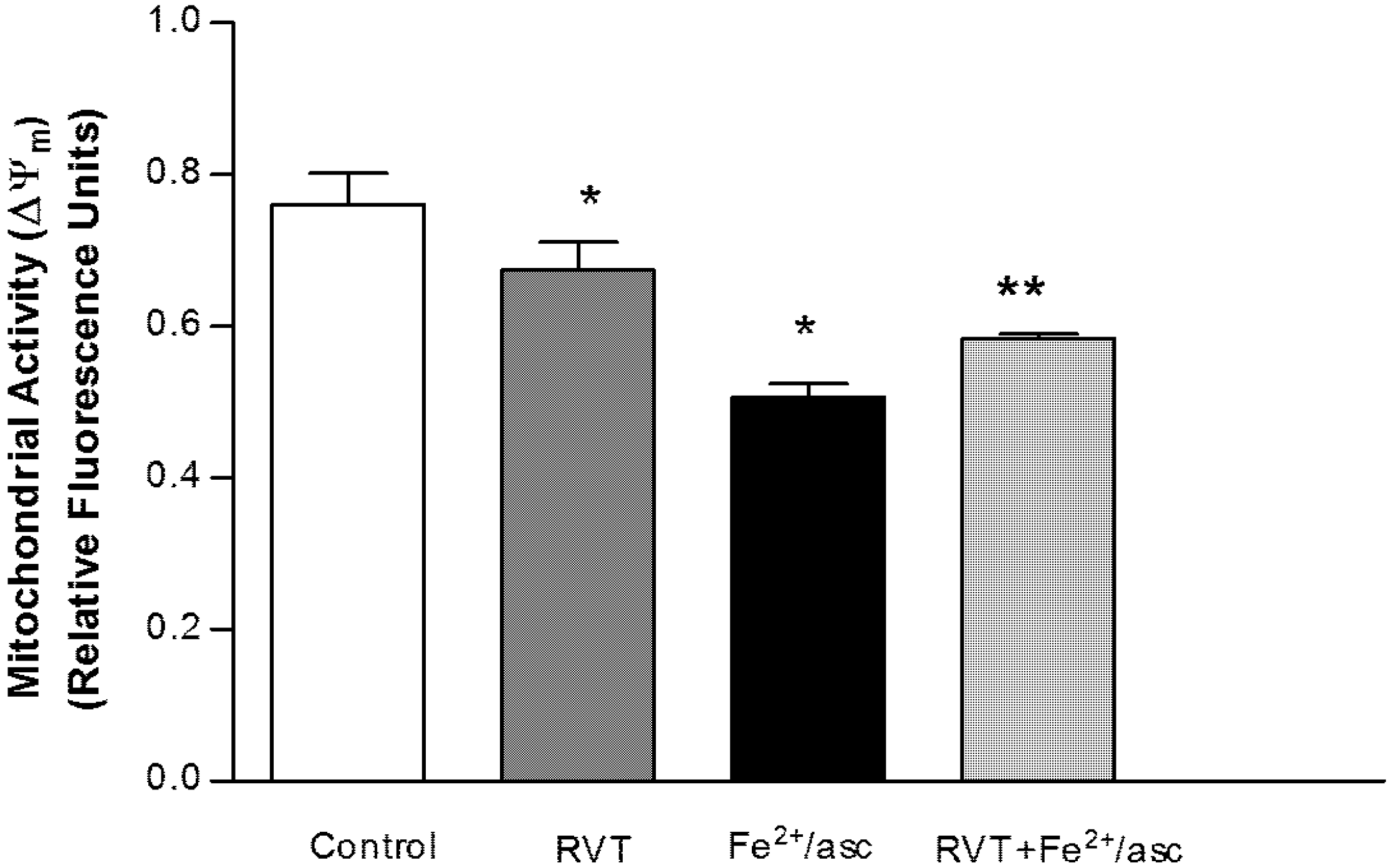
3.2. Protective Effect of Resveratrol on Mitochondrial Function
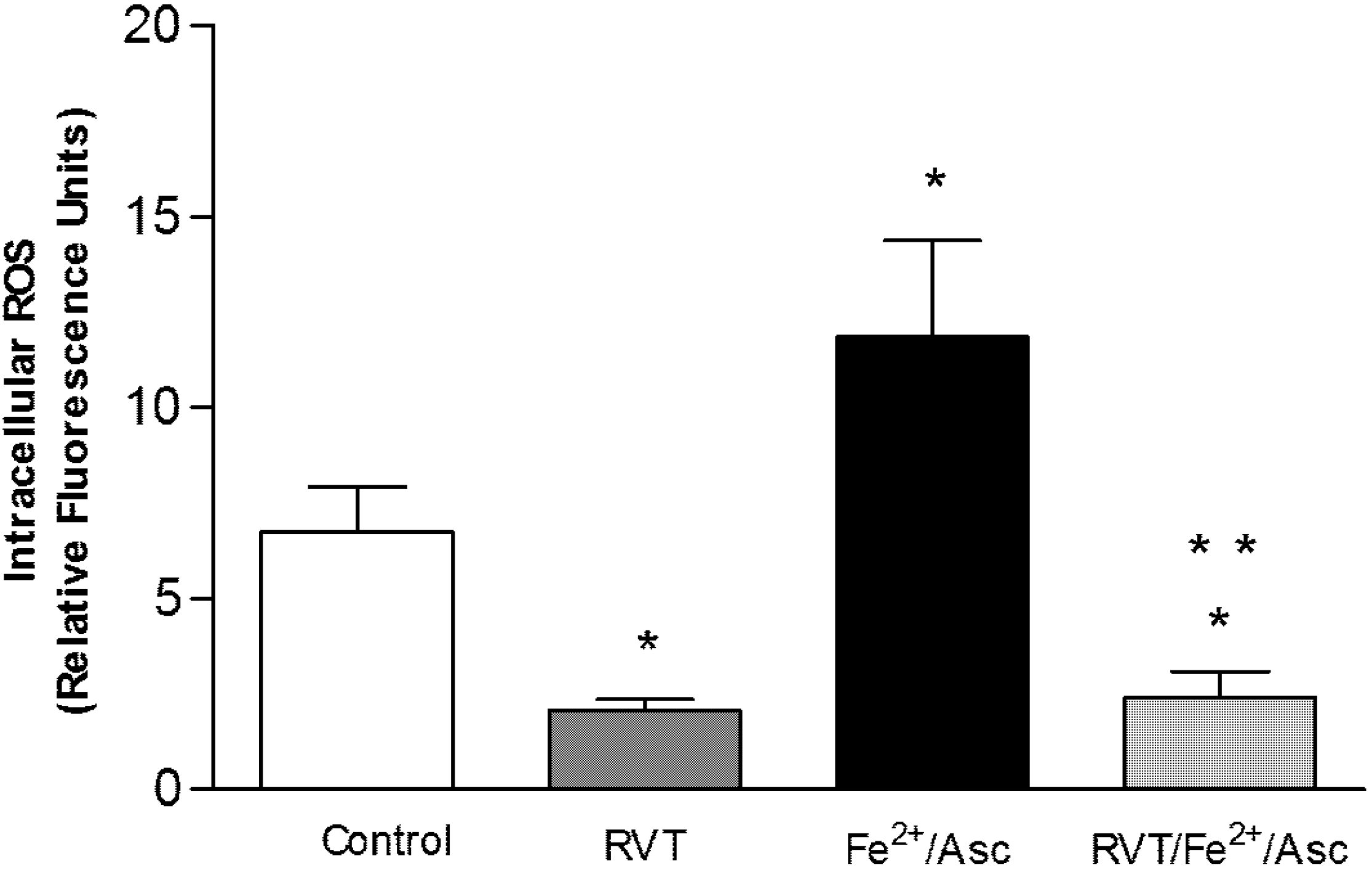
3.3. Resveratrol Diminished Intracellular ROS Generation

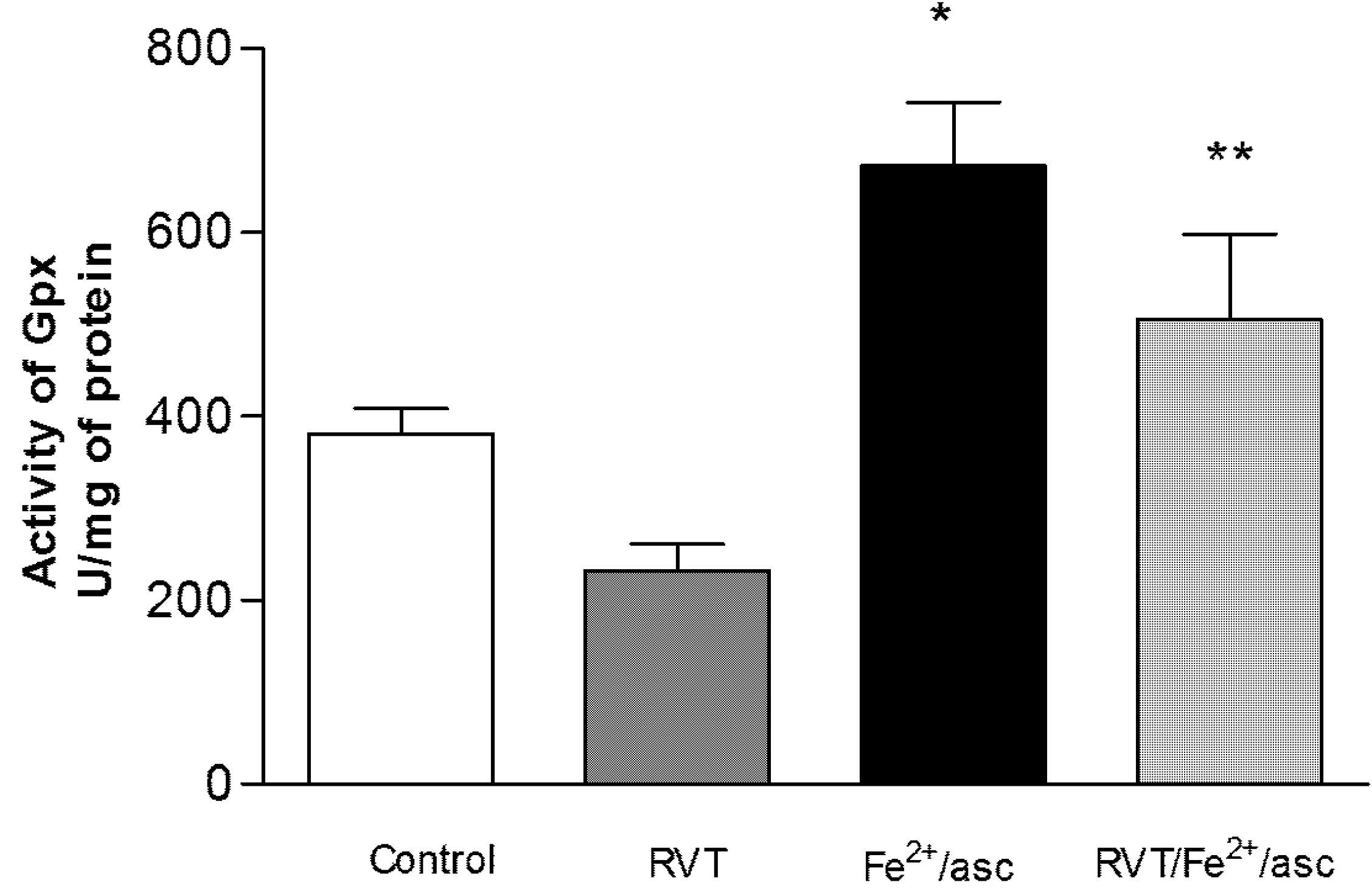
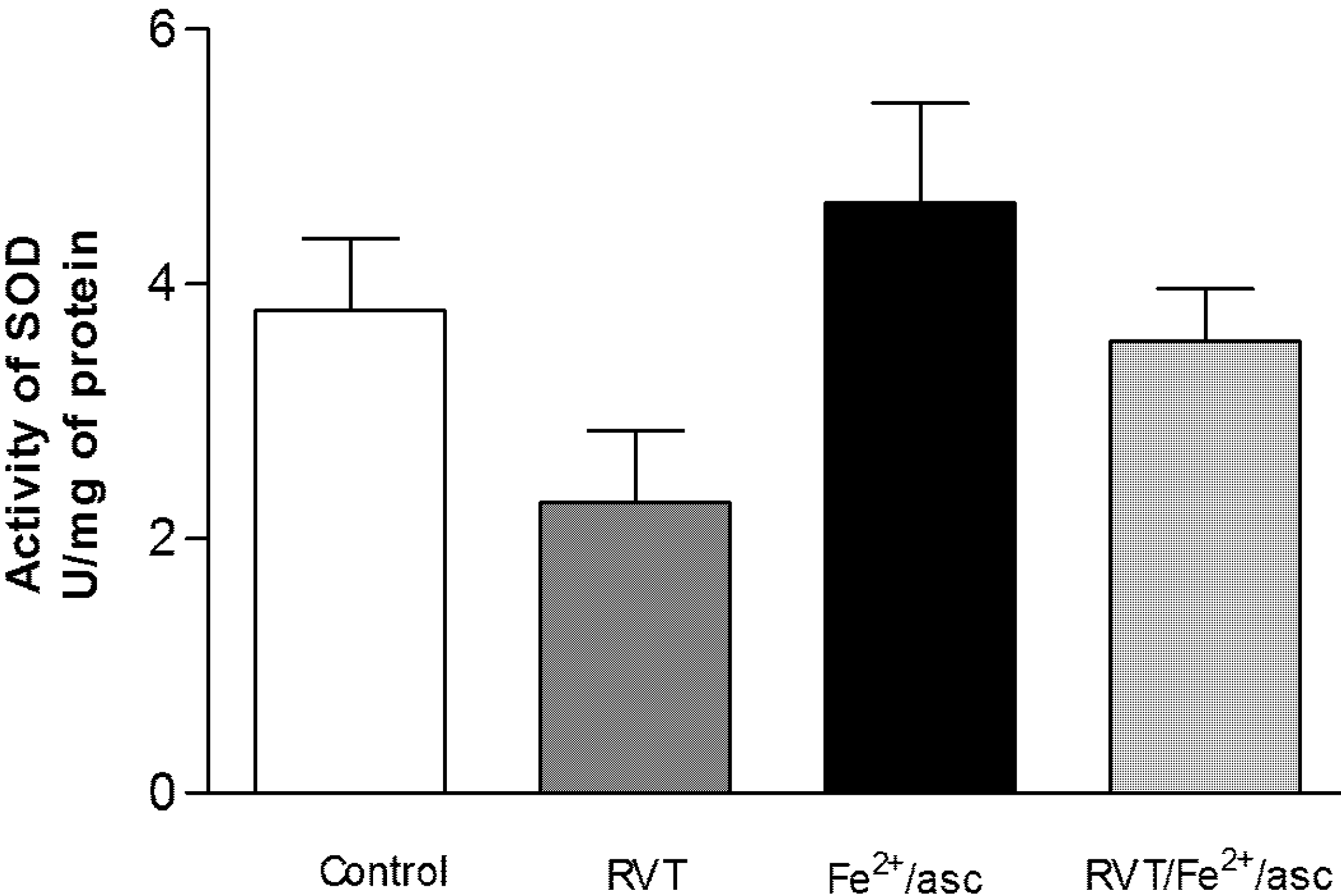
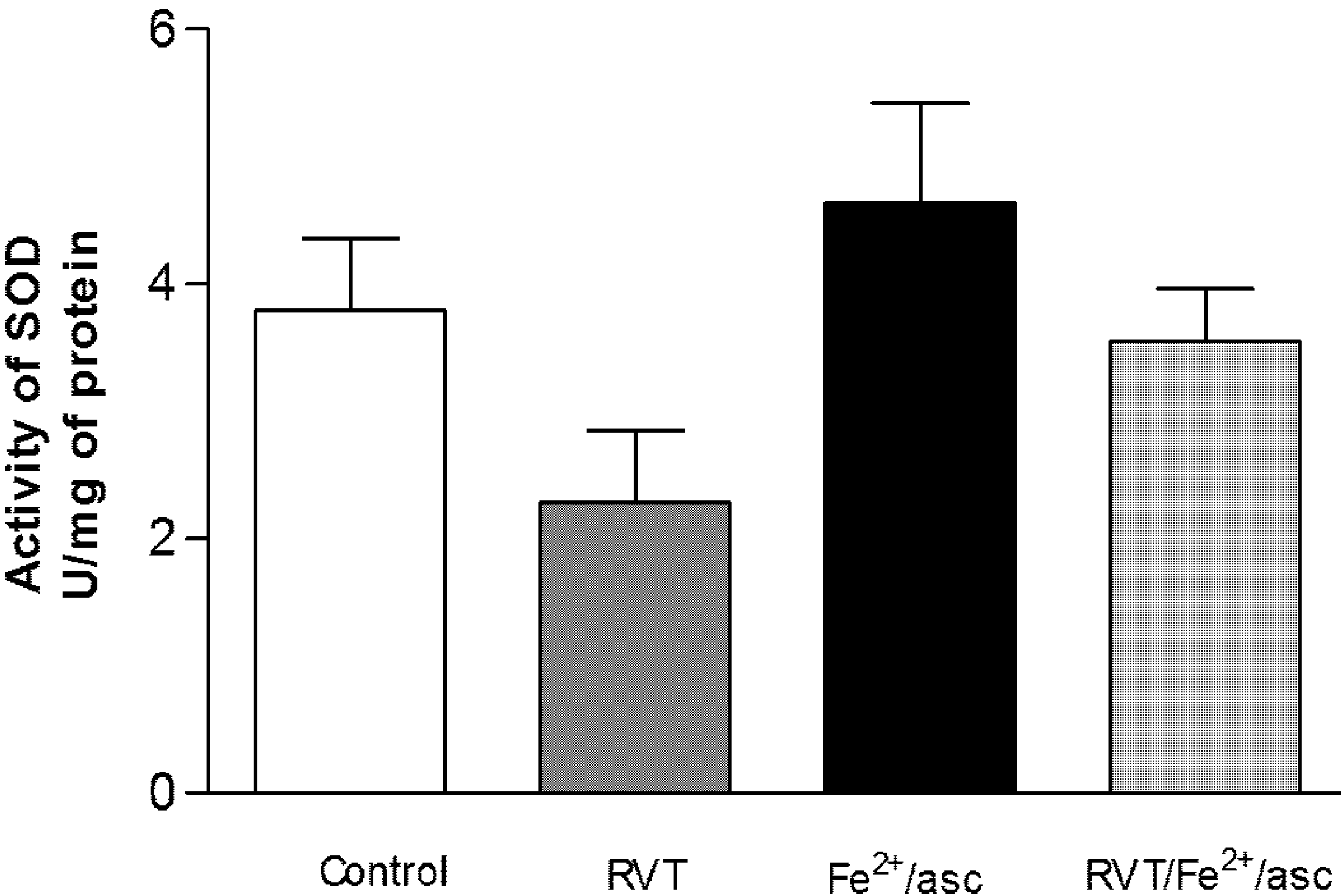
3.4. Effect of RVT on SOD and GPX Activity

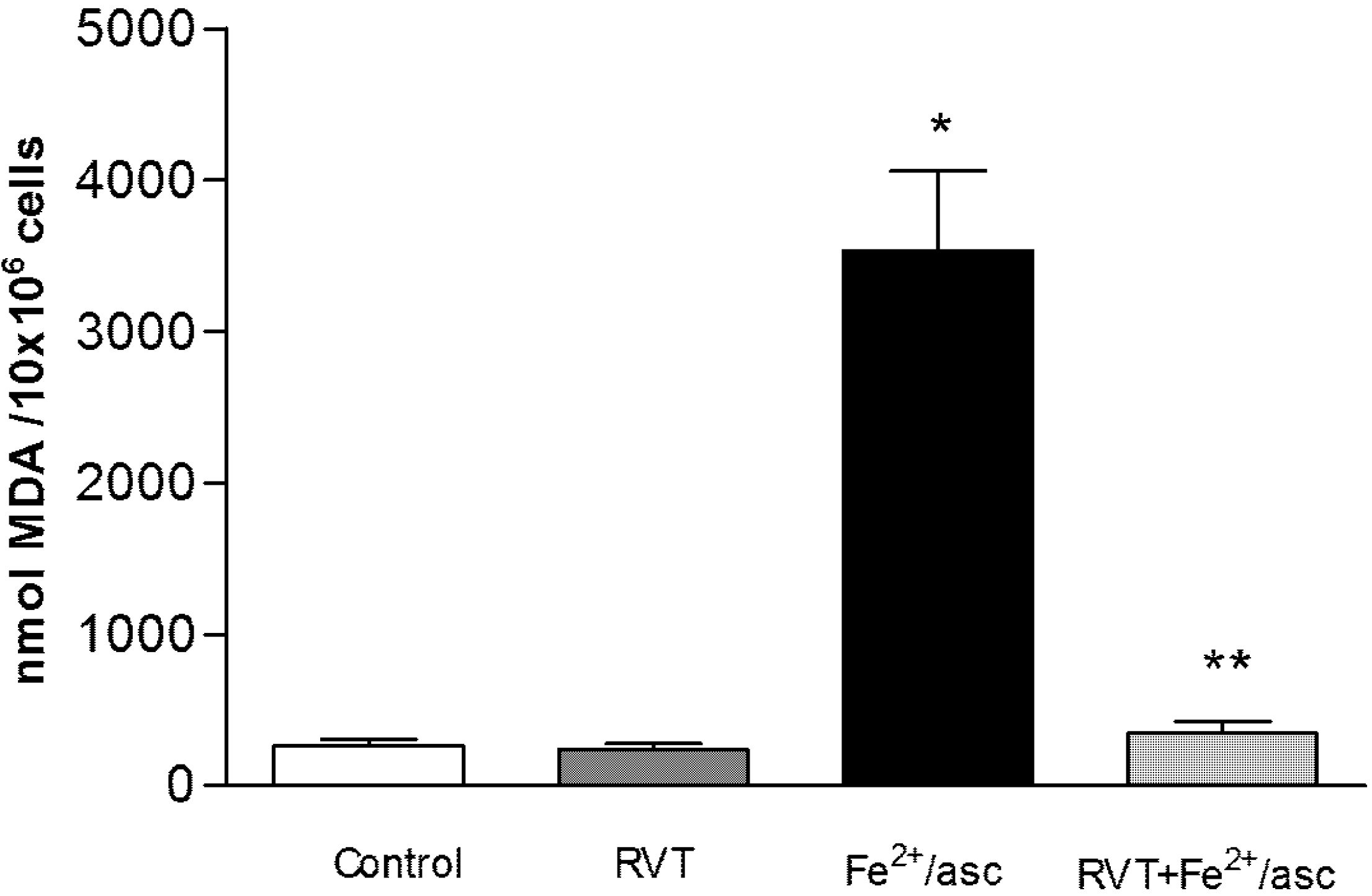
3.5. Effect of RVT on Lipid Peroxidation
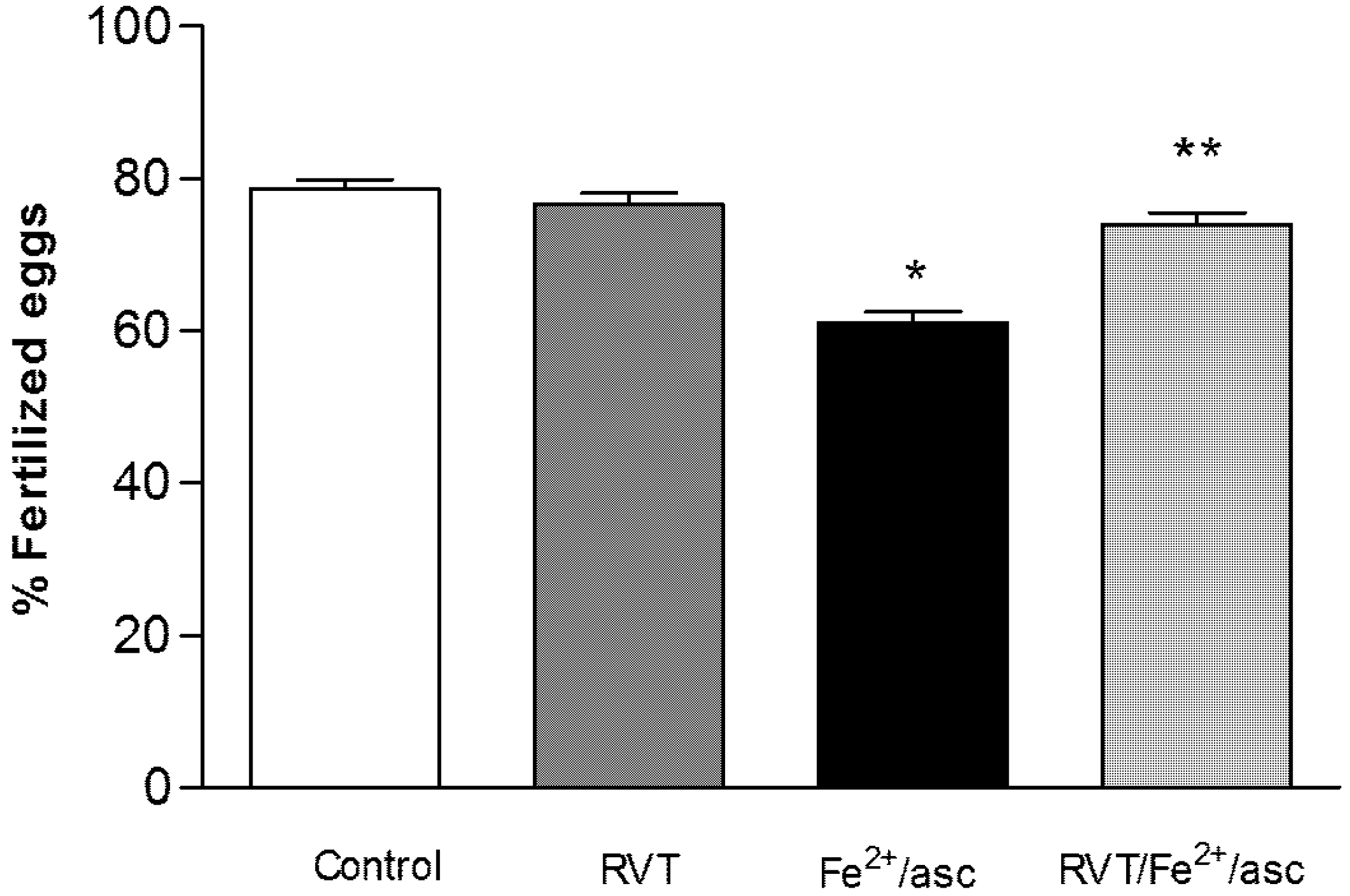
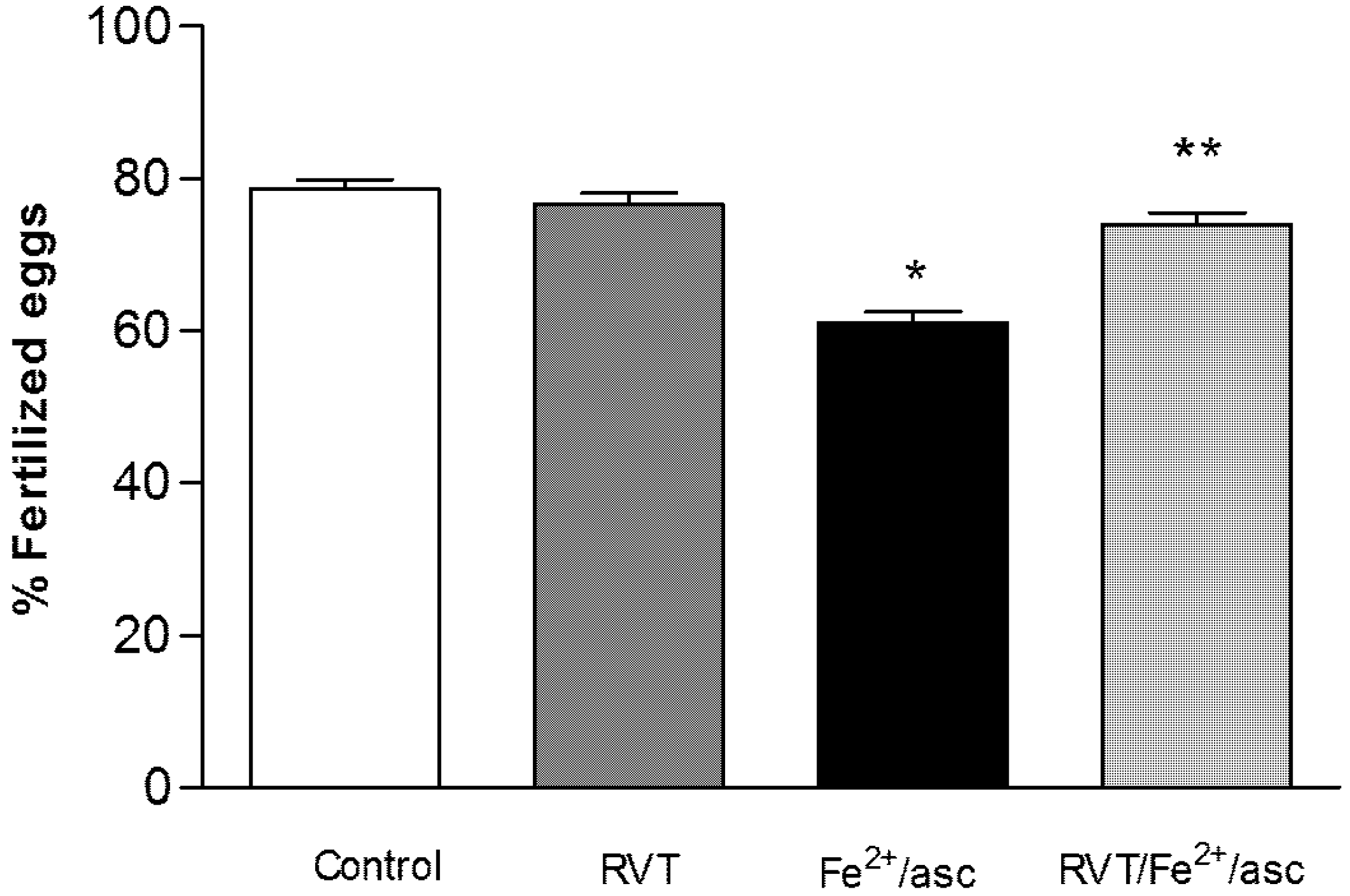
3.6. Resveratrol Retains in Vitro Fertilization Capability
4. Discussion
5. Conclusions
Conflicts of Interest
References
- Storey, K.B. Oxidative stress: Animal adaptations in nature. Braz. J. Med. Biol. Res. 1996, 29, 1715–1733. [Google Scholar]
- Sanocka, D.; Kurpisz, M. Reactive oxygen species and sperm cells. Reprod. Biol. Endocrinol. 2004, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bansal, A.K.; Bilaspuri, G.S. Impacts of oxidative stress and antioxidants on semen functions. Vet. Med. Int. 2011, 2011. [Google Scholar] [CrossRef]
- Kopers, A.J.; de luliis, G.N.; Finnie, J.M.; McLaughlin, E.A.; Aitken, R.J. Significance of mitochondrial reactive oxygen species in the generation of oxidative stress in spermatozoa. Endocr. Res. 2008, 93, 3199–3207. [Google Scholar]
- Aitken, R.J.; Baker, M.A. Oxidative stress, sperm survival and fertility control. Mol. Cell. Endocrinol. 2006, 250, 66–69. [Google Scholar] [CrossRef]
- Agarwal, A.; Gupta, S.; Sikka, S. The role of free radicals and antioxidants in reproduction. Suresh Curr. Opin. Obstet. Gynecol. 2006, 18, 325–332. [Google Scholar] [CrossRef]
- Tremellen, K. Oxidative stress and male infertility a clinical perspective. Hum. Reprod. Update 2008, 14, 243–258. [Google Scholar] [CrossRef]
- Kovacic, P.; Somanathan, R. Multifaceted approach to resveratrol bioactivity: Focus on antioxidant action, cell signaling and safety. Oxid. Med. Cell. Longev. 2010, 3, 86–100. [Google Scholar] [CrossRef]
- Bhat, K.; Kosmeder, J., II; Pezzuto, J. Biological effects of resveratrol. Antioxid. Redox Signal. 2001, 3, 1041–1064. [Google Scholar] [CrossRef]
- Cottart, C.; Antoine, V.; Morizot, C.; Beaudeux, J. Resveratrol bioavailability and toxicity in humans. Mol. Nutr. Food Res. 2010, 54, 7–16. [Google Scholar] [CrossRef]
- Xia, E.; Den, G.; Guo, Y.; Li, H. Biological activities of polyphenols from grapes. Int. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef]
- Juan, M.E.; González-Pons, E.; Munuera, T.; Ballester, J.; Rodríguez-Gil, J.E.; Planas, J.M. Trans-resveratrol, a natural antioxidant from grapes, increases sperm output in healthy rats. J. Nutr. 2005, 135, 757–760. [Google Scholar]
- Shin, S.; Hee, J.J.; Park, D.; Jang, M.J.; Choi, J.H.; Choi, B.H.; Joo, S.S.; Nahm, S.S.; Kim, J.; Kim, Y.B. Trans-resveratrol relaxes the corpus cavernosum ex vivo and enhances testosterone levels and sperm quality in vivo. Arch. Pharm. Res. 2008, 31, 83–87. [Google Scholar] [CrossRef]
- Revel, A.; Raanani, H.; Younglai, E.; Xu, J.; Han, R.; Savouret, J.F.; Casper, R.F. Resveratrol, a natural aryl hydrocarbon receptor antagonist, protects sperm from DNA damage and apoptosis caused by benzo(a)pyrene. Reprod. Toxicol. 2001, 15, 479–486. [Google Scholar] [CrossRef]
- Uguralp, S.; Mizrak, B.; Bay Karabulut, A. Resveratrol reduces ischemia reperfusion injury after experimental testicular torsion. Eur. J. Pediatr. Surg. 2005, 15, 114–119. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Laboratory Manual for the Examination of Human Semen and Semen Cervical Mucus Interaction; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Metheods Enzymol. 1978, 52, 302–310. [Google Scholar] [CrossRef]
- Aitken, R.J.; Curry, B.J. Redox regulation of human sperm function: From the physiological control of sperm capacitation to the etiology of infertility and DNA damage in the germ line. Antioxid. Redox Signal. 2011, 14, 367–381. [Google Scholar] [CrossRef]
- Ford, W.C. Regulation of sperm function by reactive oxygen species. Hum. Reprod. Update 2004, 10, 387–399. [Google Scholar] [CrossRef]
- Griveau, J.F.; Dumont, E.; Renard, P.; Callegari, J.P.; Le Lannou, D. Reactive oxygen species, lipid peroxidation and enzymatic defence systems in human spermatozoa. J. Reprod. Fertil. 1995, 103, 17–26. [Google Scholar] [CrossRef]
- Buffone, M.G.; Ijiri, T.W.; Cao, W.; Merdiushev, T.; Aghajanian, H.K.; Gerton, G.L. Heads or tails? Structural events and molecular mechanisms that promote mammalian sperm acrosomal exocytosis and motility. Mol. Reprod. Dev. 2012, 79, 4–18. [Google Scholar] [CrossRef]
- Lavranos, G.; Balla, M.; Tzortzopoulou, A.; Syriou, V.; Angelopoulou, R. Investigating ROS sources in male infertility: A common end for numerous pathways. Reprod. Toxicol. 2012, 34, 298–307. [Google Scholar] [CrossRef]
- Agarwal, A.; Tamer, M.S. Role of sperm chromatin abnormalities and DNA damage in male infertility. Hum. Reprod. Update 2003, 9, 331–345. [Google Scholar] [CrossRef]
- Zini, A.; San Gabriel, M.; Baazeem, A. Antioxidants and sperm DNA damage: A clinical perspective. J. Assist. Reprod. Genet. 2009, 26, 427–432. [Google Scholar] [CrossRef]
- Zini, A.; Al-Hathal, N. Antioxidant therapy in male infertility: Fact or fiction? Asian J. Androl. 2011, 13, 374–381. [Google Scholar] [CrossRef]
- Lombardo, F.; Sansone, A.; Romanelli, F.; Paoli, D.; Gandini, L.; Lenzi, A. The role of antioxidant therapy in the treatment of male infertility: An overview. Asian J. Androl. 2011, 13, 690–697. [Google Scholar]
- Mínguez-Alarcón, L.; Mendiola, J.; López-Espín, J.J.; Sarabia-Cos, L.; Vivero-Salmerón, G.; Vioque, J.; Navarrete-Muñoz, E.M.; Torres-Cantero, A.M. Dietary intake of antioxidant nutrients is associated with semen quality in young university students. Hum. Reprod. 2012, 27, 2807–2814. [Google Scholar] [CrossRef]
- Zareba, P.; Colaci, D.S.; Afeiche, M.; Gaskins, A.J.; Jørgensen, N.; Mendiola, J.; Swan, S.H.; Chavarro, J.E. Semen quality in relation to antioxidant intake in a healthy male population. Fertil. Steril. 2013, 100, 1572–1579. [Google Scholar] [CrossRef]
- Afeiche, M.; Williams, P.L.; Mendiola, J.; Gaskins, A.J.; Jørgensen, N.; Swan, S.H.; Chavarro, J.E. Dairy food intake in relation to semen quality and reproductive hormone levels among physically active young men. Hum. Reprod. 2013, 28, 2265–2275. [Google Scholar] [CrossRef]
- Ourique, G.M.; Finamor, I.A.; Saccol, E.M.; Riffel, A.P.; Pês, T.S.; Gutierrez, K.; Gonçalves, P.B.; Baldisserotto, B.; Pavanato, M.A.; Barreto, K.P. Resveratrol improves sperm motility, prevents lipid peroxidation and enhances antioxidant defences in the testes of hyperthyroid rats. Reprod. Toxicol. 2013, 37, 31–39. [Google Scholar] [CrossRef]
- Yuluğ, E.; Türedi, S.; Alver, A.; Türedi, S.; Kahraman, C. Effects of resveratrol on methotrexate-induced testicular damage in rats. Sci. World J. 2013, 2013. [Google Scholar] [CrossRef]
- Goldberg, D.M.; Ng, E.; Yan, J.; Karumanchirí, A.; Soleas, G.J.; Diamandis, E.P. Regional differences in resveratrol isomer concentrations of wines from various cultivars. J. Wine Res. 1996, 7, 13–24. [Google Scholar] [CrossRef]
- McMurtrey, K.D. Resveratrol in Wine. In Wine: Nutritional and Therapeutic Benefits; Watkins, T.R., Ed.; Oxford University Press: Oxford, UK, 1997; pp. 45–55. [Google Scholar]
- Juan, M.E.; Vinardell, M.P.; Planas, J.M. The daily oral administration of high doses of trans-resveratrol to rats for 28 days is not harmful. J. Nutr. 2002, 132, 257–260. [Google Scholar]
- Patel, K.R.; Scott, E.; Brown, V.A.; Gescher, A.J.; Steward, W.P.; Brown, K. Clinical trials of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 161–169. [Google Scholar] [CrossRef]
- Silva, E.C.; Cajueiro, J.F.; Silva, S.V.; Soares, P.C.; Guerra, M.M. Effect of antioxidants resveratrol and quercetin on in vitro evaluation of frozen ram sperm. Theriogenology 2012, 77, 1722–1726. [Google Scholar] [CrossRef]
- Aitken, R.J.; Jane, S.; Clarkson, S.; Fishel, S. Generation of reactive oxygen species, lipid peroxidation, and human sperm function. Biol. Reprod. 1989, 40, 183–197. [Google Scholar]
- Martínez-Pastor, F.; Aisen, E.; Fernández-Santos, M.R.; Esteso, M.C.; Maroto-Morales, A.; García-Álvarez, O.; Garde, J.J. Reactive oxygen species generators affect quality parameters and apoptosis markers differently in red deer spermatozoa. Reproduction 2009, 137, 225–235. [Google Scholar]
- Buettner, G.R.; Jurkiewicz, B.A. Catalytic metals, ascorbate and free radicals: Combinations to avoid. Radiat. Res. 1996, 145, 532–541. [Google Scholar] [CrossRef]
- Armstrong, J.S.; Rajasekaran, M.; Chamulitrat, W.; Gatti, P.; Hellstrom, W.J.; Sikka, S.C. Characterization of reactive oxygen species induced effects on human spermatozoa movement and energy metabolism. Free Radic. Biol. Med. 1999, 26, 869–880. [Google Scholar]
- Leonard, S.S.; Xia, C.; Jiang, B.H.; Stinefelt, B.; Klandorf, H.; Harris, G.K.; Shi, X. Resveratrol scavenges reactive oxygen species and effects radical-induced cellular responses. Biochem. Biophys. Res. Commun. 2003, 309, 1017–1026. [Google Scholar] [CrossRef]
- Gravance, C.G.; Garner, D.L.; Miller, M.G.; Berger, T. Fluorescent probes and flow cytometry to assess rat sperm integrity and mitochondrial function. Reprod. Toxicol. 2001, 15, 5–10. [Google Scholar]
- Van Ginkel, P.R.; Sareen, D.; Subramanian, L.; Walker, Q.; Darjatmoko, S.R.; Lindstrom, M.J.; Kulkarni, A.; Albert, D.M.; Polans, A.S. Resveratrol inhibits tumor growth of human neuroblastoma and mediates apoptosis by directly targeting mitochondria. Clin. Cancer Res. 2007, 13, 5162–5169. [Google Scholar] [CrossRef]
- Zini, R.; Morin, C.; Bertelli, A.; Bertelli, A.A.; Tillement, J.P. Effects of resveratrol on the rat brain respiratory chain. Drugs Exp. Clin. Res. 1999, 25, 87–97. [Google Scholar]
- Collodel, G.; Federico, M.G.; Geminiani, M.; Martini, S.; Bonechi, C.; Rossi, C.; Figura, N. Effect of trans-resveratrol on induced oxidative stress in human sperm and in rat germinal cells. Reprod. Toxicol. 2011, 31, 239–246. [Google Scholar] [CrossRef]
- Berrougui, H.; Grenier, G.; Loued, S.; Drouin, G.; Khalil, A. A new insight into resveratrol as an atheroprotective compound: Inhibition of lipid peroxidation and enhancement of cholesterol efflux. Atherosclerosis 2009, 207, 420–427. [Google Scholar] [CrossRef]
- López-Vélez, M.; Martinez-Martinez, F.; Del Valle-Ribes, C. The study of phenolic compounds as natural antioxidants in wine. Crit. Rev. Food Sci. Nutr. 2003, 43, 233–244. [Google Scholar]
- Visconti, P.E.; Ning, X.; Fornes, M.W.; Alvarez, J.G.; Stein, P.; Connors, S.A.; Kopf, G.S. Cholesterol efflux-mediated signal transduction in mammalian sperm: Cholesterol release signals an increase in protein tyrosine phosphorylation during mouse sperm capacitation. Dev. Boil. 1999, 214, 429–443. [Google Scholar] [CrossRef]
- Gray, J.E.; Starmer, J.; Lin, V.S.; Dickinson, B.C.; Magnuson, T. Mitochondrial hydrogen peroxide and defective cholesterol efflux prevent in vitro fertilization by cryopreserved inbred mouse sperm. Biol. Reprod. 2013, 89, 17. [Google Scholar] [CrossRef]
- Kwak, S.S.; Cheonga, S.A.; Jeona, Y.; Leeb, E.; Choic, K.C.; Jeungc, E.B.; Hyun, S.H. The effects of resveratrol on porcine oocyte in vitro maturation and subsequent embryonic development after parthenogenetic activation and in vitro fertilization. Theriogenology 2012, 78, 86–101. [Google Scholar] [CrossRef]
- Lee, K.; Wang, C.; Chaille, J.M.; Machaty, Z. Effect of resveratrol on the development of porcine embryos produced in vitro. J. Reprod. Dev. 2010, 56, 330–335. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mojica-Villegas, M.A.; Izquierdo-Vega, J.A.; Chamorro-Cevallos, G.; Sánchez-Gutiérrez, M. Protective Effect of Resveratrol on Biomarkers of Oxidative Stress Induced by Iron/Ascorbate in Mouse Spermatozoa. Nutrients 2014, 6, 489-503. https://doi.org/10.3390/nu6020489
Mojica-Villegas MA, Izquierdo-Vega JA, Chamorro-Cevallos G, Sánchez-Gutiérrez M. Protective Effect of Resveratrol on Biomarkers of Oxidative Stress Induced by Iron/Ascorbate in Mouse Spermatozoa. Nutrients. 2014; 6(2):489-503. https://doi.org/10.3390/nu6020489
Chicago/Turabian StyleMojica-Villegas, María Angélica, Jeannett Alejandra Izquierdo-Vega, Germán Chamorro-Cevallos, and Manuel Sánchez-Gutiérrez. 2014. "Protective Effect of Resveratrol on Biomarkers of Oxidative Stress Induced by Iron/Ascorbate in Mouse Spermatozoa" Nutrients 6, no. 2: 489-503. https://doi.org/10.3390/nu6020489