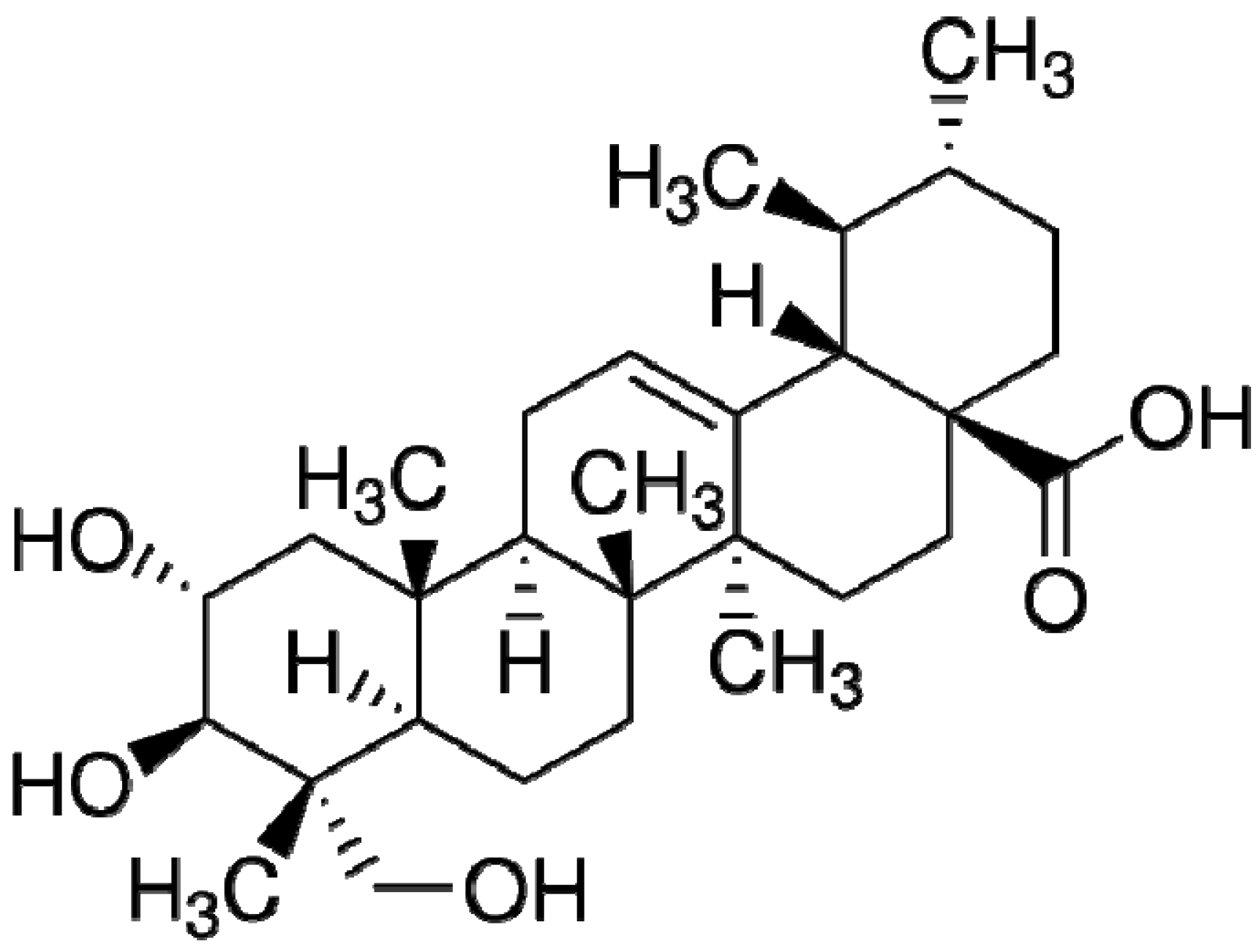
Asiatic Acid Alleviates Hemodynamic and Metabolic Alterations via Restoring eNOS/iNOS Expression, Oxidative Stress, and Inflammation in Diet-Induced Metabolic Syndrome Rats

Abstract
:1. Introduction
2. Experimental Section
2.1. Animals
2.2. Experimental Protocols
2.3. Indirect Measurement of Blood Pressure in Conscious Rats
2.4. Fasting Blood Glucose (FBG), Oral Glucose Tolerance Test (OGTT), and Serum Insulin Assessments
2.5. Fasting Serum Insulin Assessments and HOMA-IR Calculation
2.6. Hemodynamic Measurements
2.7. Measurement of Oxidative Stress Markers
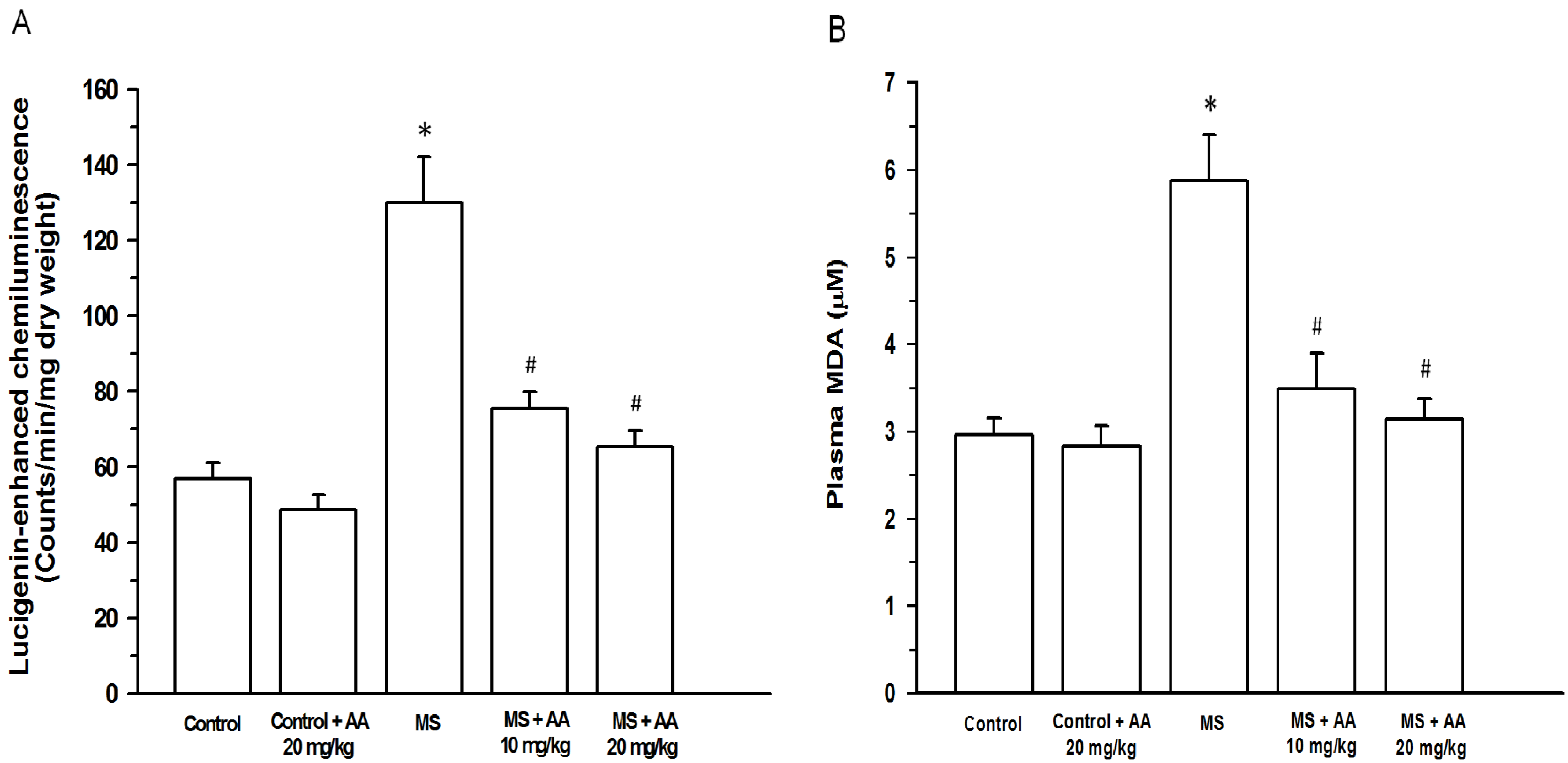
2.7.1. O2•− Production
2.7.2. Plasma MDA
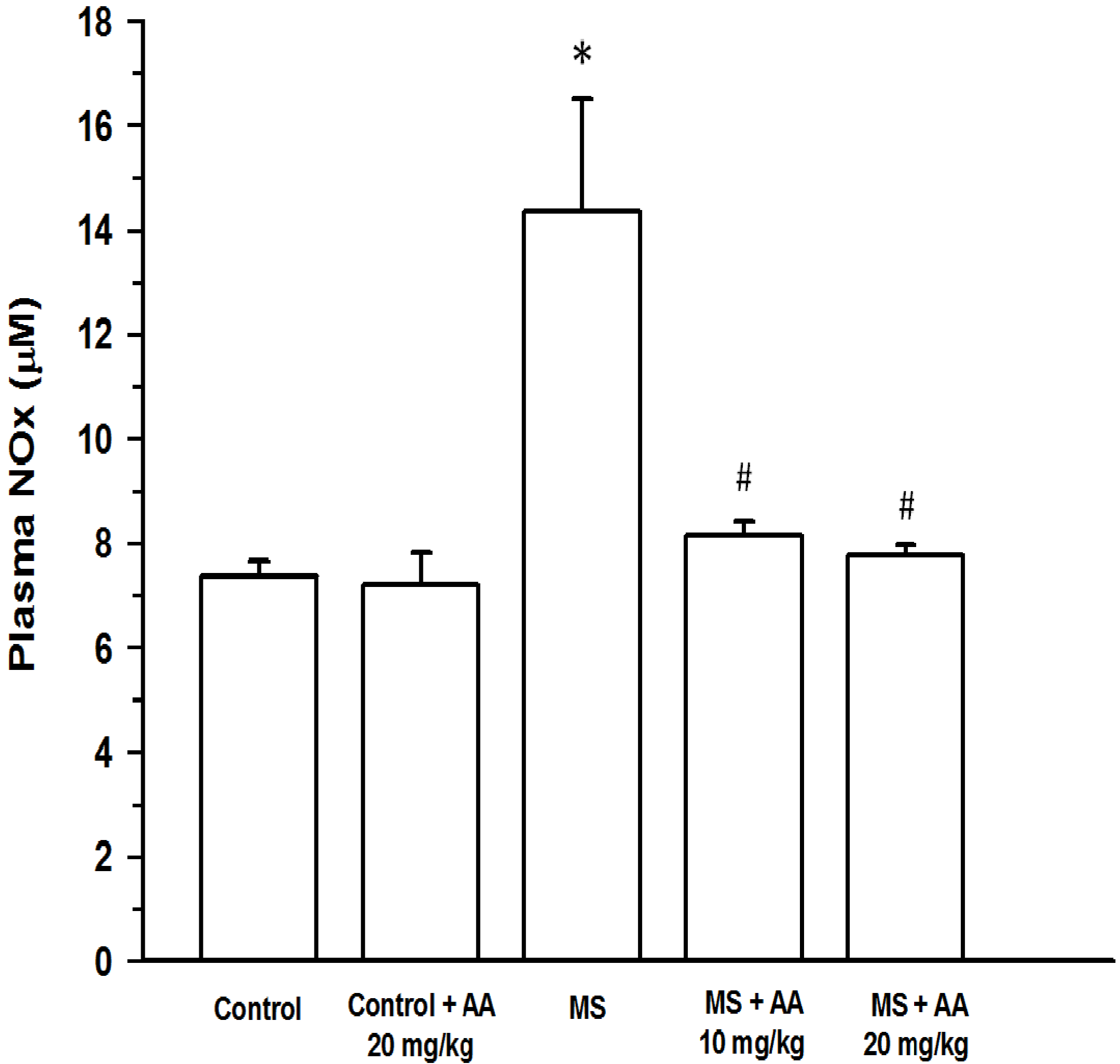
2.7.3. Plasma NOx
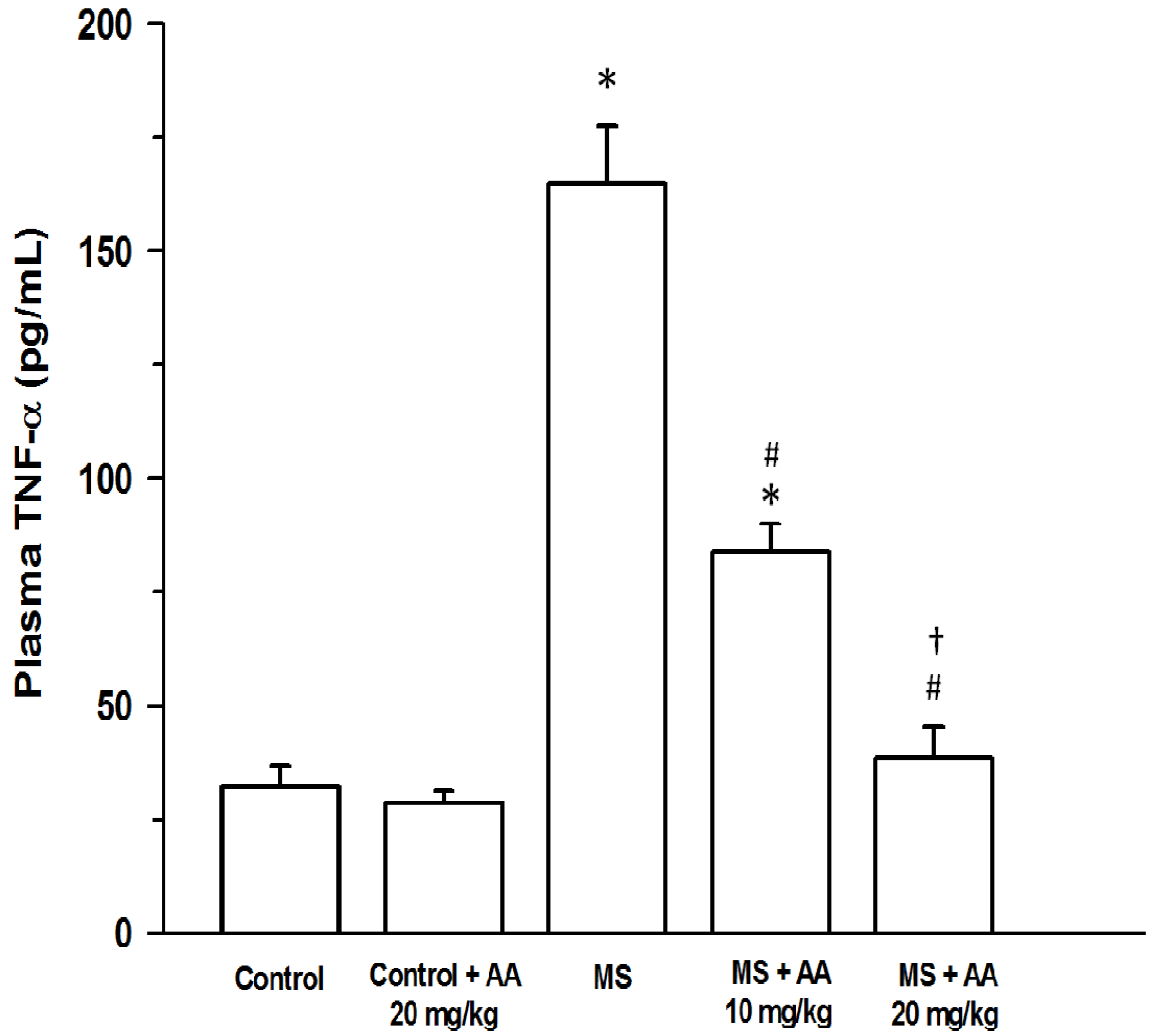
2.7.4. TNF-α
2.8. Western Blot Analysis
2.9. Chemicals
2.10. Statistical Analysis
3. Results
3.1. Effects of the Asiatic Acid on Metabolic Abnormalities

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Control | Control + AA 20 mg/kg | MS | MS + AA 10 mg/kg | MS + AA 20 mg/kg |
|---|---|---|---|---|---|
| Fasting blood glucose (mg/dL) | 81.13 ± 1.3 | 82.88 ± 2.1 | 116.71 ± 2.8 * | 93.50 ± 2.1 # | 89.25 ± 1.5 # |
| Fasting serum insulin (ng/mL) | 0.45 ± 0.06 | 0.47 ± 0.06 | 4.34 ± 0.43 * | 2.19 ± 0.13 *,# | 1.38 ± 0.24 *,#,† |
| AUC (mg/dL/120 min) | 13,065 ± 425 | 13,018 ± 410 | 16,125 ± 202 * | 14,393 ± 173 # | 14,073 ± 180 # |
| HOMA-IR score | 2.27 ± 0.32 | 2.90 ± 0.4 | 28.90 ± 3.60 * | 12.54 ± 0.78 *,# | 6.72 ± 1.22 *,#,† |
| Cholesterol (mg/dL) | 54.00 ± 2.29 | 55.38 ± 2.31 | 89.00 ± 2.31 * | 67.38 ± 2.20 *,# | 61.43 ± 3.26 # |
| Triglycerides (mg/dL) | 24.50 ± 3.08 | 25.75 ± 2.40 | 70.71 ± 2.71 * | 41.71 ± 2.10 *,# | 29.57 ± 2.27 *,#,† |
| HDL-C (mg/dL) | 41.38 ± 1.29 | 41.29 ± 1.27 | 14.86 ± 1.52 * | 26.50 ± 0.63 *,# | 34.13 ± 0.55 *,#,† |
| ALP (U/L) | 73.1 ± 4.5 | 74.5 ± 3.6 | 150.7 ± 1.1 * | 119.0 ± 4.2 *,# | 82.2 ± 3.9 #,† |
| ALT (U/L) | 21.0 ± 0.9 | 19.2 ± 1.6 | 34.7 ± 2.6 * | 25.0 ± 1.2 *,# | 21.5 ± 0.8 # |
| Body weight (g) | 426.57 ± 5.0 | 416.75 ± 5.8 | 423.71 ± 6.5 | 408.43 ± 6.9 | 404.86 ± 10.6 |
| Liver wet weight/body weight (mg/g) | 30.3 ± 1.1 | 30.7 ± 0.6 | 43.3 ± 1.2 * | 33.6 ± 0.4 *,# | 32.2 ± 0.3 #,† |
| Heart wet weight/body weight (mg/g) | 3.37 ± 0.05 | 3.33 ± 0.03 | 3.52 ± 0.07 | 3.39 ± 0.07 | 3.33 ± 0.1 |
| LV wet weight/body weight (mg/g) | 2.21 ± 0.02 | 2.24 ± 0.04 | 2.34 ± 0.04 | 2.22 ± 0.03 | 2.21 ± 0.04 |
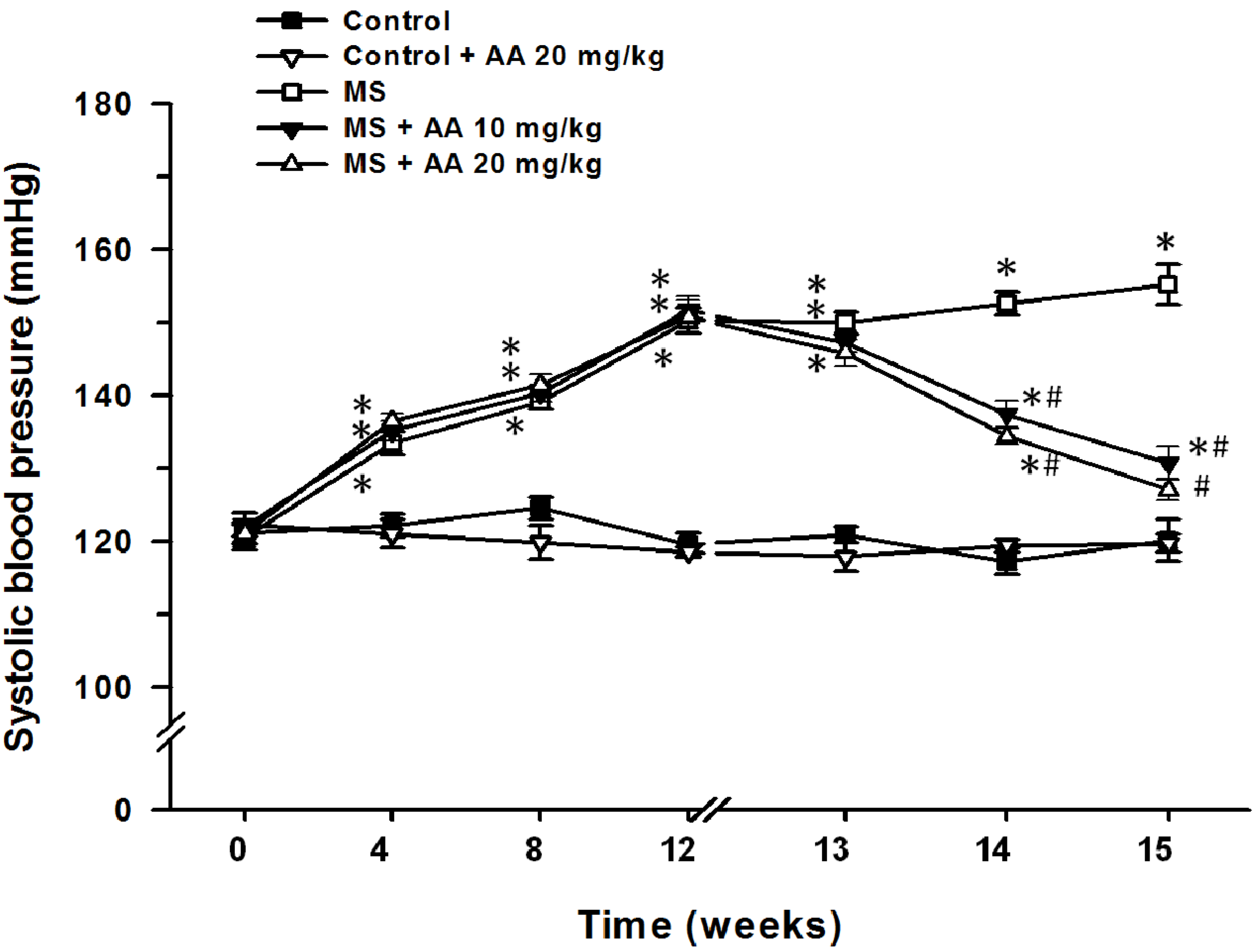
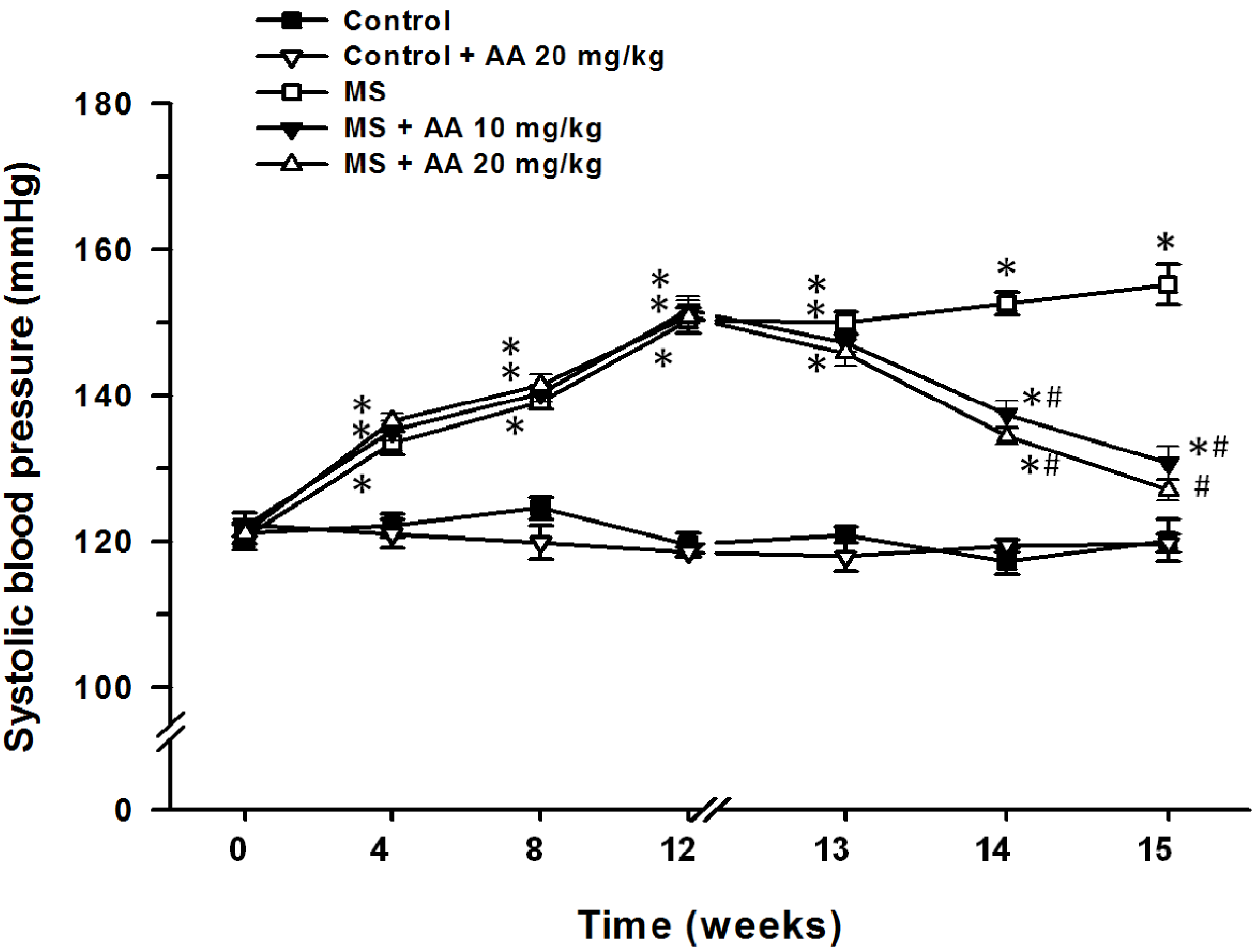
3.2. Effects of Asiatic Acid on SP (Indirect Measurement of Blood Pressure)
3.3. Effects of Asiatic Acid on Hemodynamic Parameters
| Parameters | Control | Control + AA 20 mg/kg | MS | MS + AA 10 mg/kg | MS + AA 20 mg/kg |
|---|---|---|---|---|---|
| SP (mmHg) | 117.83 ± 1.9 | 116.61 ± 1.6 | 149.14 ± 3.6 * | 132.91 ± 1.0 *,# | 128.21 ± 1.7 *,# |
| DP (mmHg) | 76.96 ± 1.7 | 77.07 ± 1.5 | 98.51 ± 2.4 * | 88.47 ± 1.5 *,# | 88.28 ± 1.3 *,# |
| MAP (mmHg) | 90.58 ± 1.7 | 90.25 ± 1.4 | 116.05 ± 2.7 * | 103.08 ± 1.1 *,# | 101.59 ± 1.3 *,# |
| HR (beat/min) | 345.89 ± 8.6 | 344.76 ± 5.7 | 417.32 ± 14.6 * | 359.34 ± 4.9 # | 356.14 ± 6.4 # |
| HBF (mL/min/100 g tissue) | 7.38 ± 0.35 | 7.49 ± 0.40 | 4.00 ± 0.18 * | 5.91 ± 0.28 *,# | 6.33 ± 0.29 # |
| HVR(mmHg/mL/min/100 g tissue) | 12.44 ± 0.53 | 12.31 ± 0.73 | 29.42 ± 1.77 * | 17.73 ± 0.84 *,# | 16.27 ± 0.74 *,# |
3.4. Effects of the Asiatic Acid on Oxidative Stress Markers
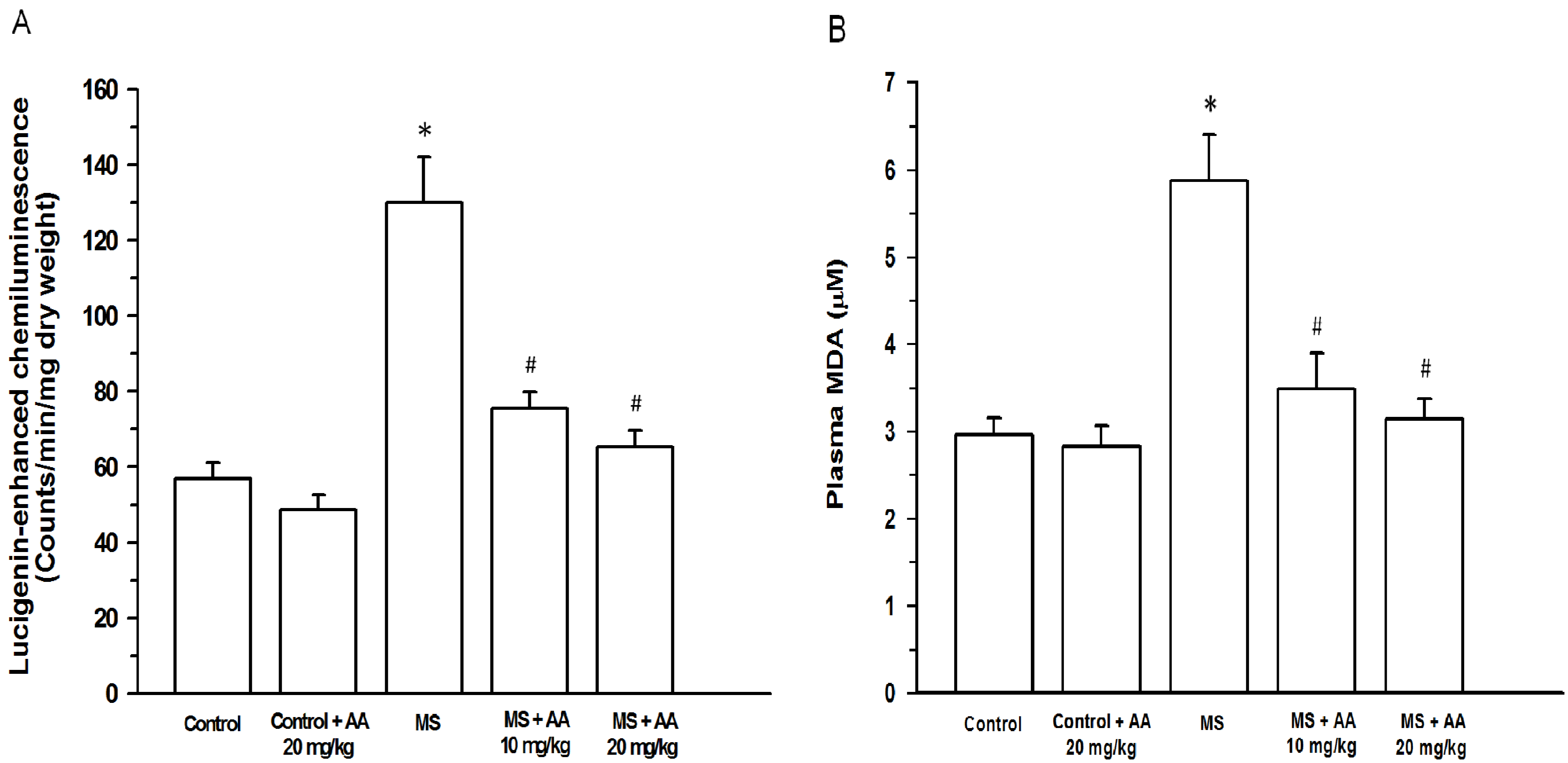
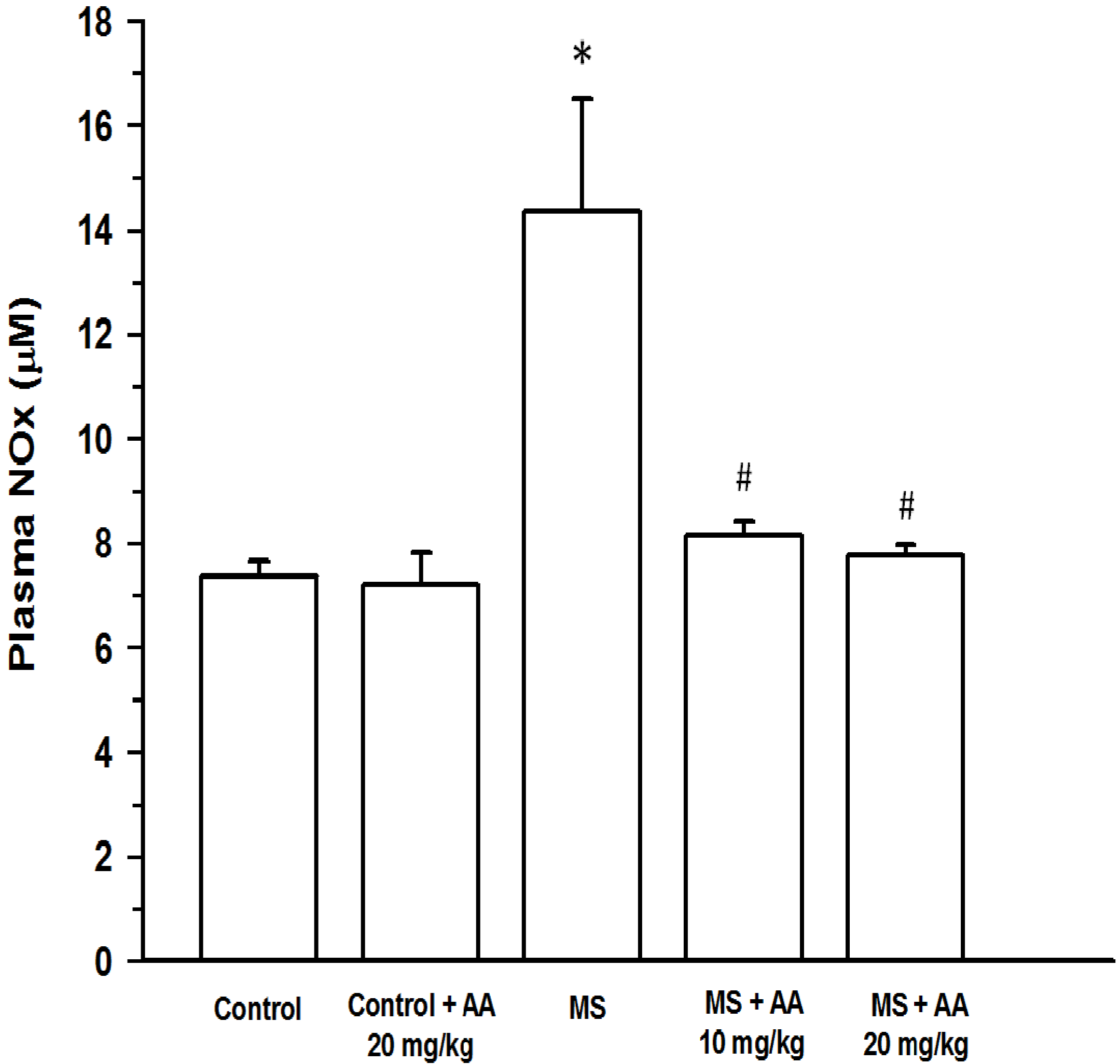
3.5. Effects of Asiatic Acid on Plasma NOx and TNF-α Concentration
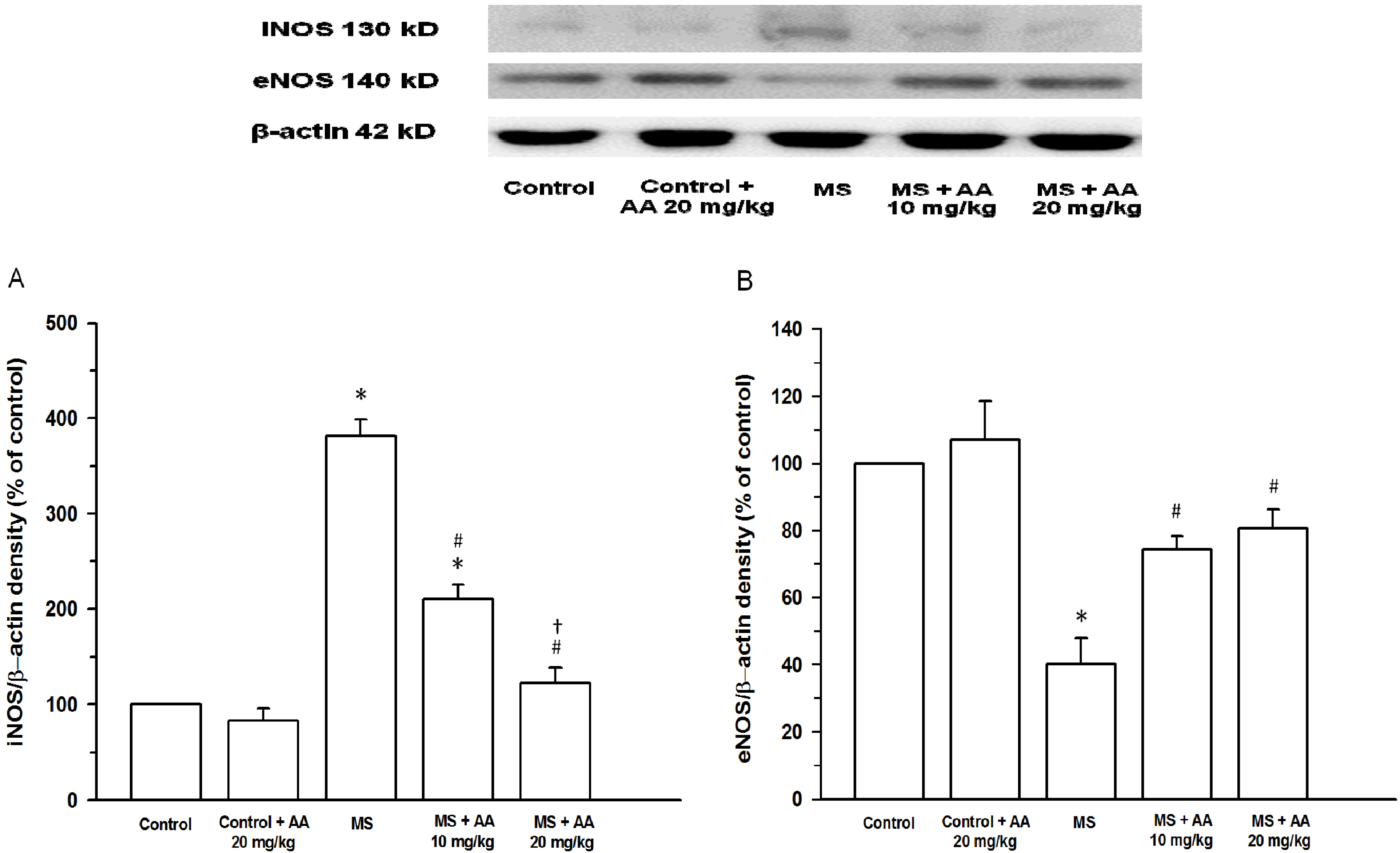
3.6. Effects of Asiatic Acid on Expression of iNOS and eNOS Protein in Aortic Tissues


4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Alam, M.A.; Kauter, K.; Brown, L. Naringin improves diet-induced cardiovascular dysfunction and obesity in high carbohydrate, high fat diet-fed rats. Nutrients 2013, 5, 637–650. [Google Scholar] [CrossRef] [Green Version]
- Poudyal, H.; Kumar, S.A.; Iyer, A.; Waanders, J.; Ward, L.C.; Brown, L. Responses to oleic, linoleic and alpha-linolenic acids in high-carbohydrate, high-fat diet-induced metabolic syndrome in rats. J. Nutr. Biochem. 2013, 24, 1381–1392. [Google Scholar] [CrossRef]
- Wong, W.Y.; Poudyal, H.; Ward, L.C.; Brown, L. Tocotrienols reverse cardiovascular, metabolic and liver changes in high carbohydrate, high fat diet-fed rats. Nutrients 2012, 4, 1527–1541. [Google Scholar] [CrossRef] [Green Version]
- Panchal, S.K.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, M.A.; Diwan, V.; Kauter, K.; Sernia, C.; Campbell, F.; Ward, L.; et al. High-carbohydrate, high-fat diet-induced metabolic syndrome and cardiovascular remodeling in rats. J. Cardiovasc. Pharmacol. 2011, 57, 611–624. [Google Scholar] [CrossRef]
- Zimmet, P.; Alberti, K.G.; Shaw, J. Global and societal implications of the diabetes epidemic. Nature 2001, 414, 782–787. [Google Scholar] [CrossRef]
- Isomaa, B.; Almgren, P.; Tuomi, T.; Forsen, B.; Lahti, K.; Nissen, M.; Taskinen, M.R.; Groop, L. Cardiovascular morbidity and mortality associated with the metabolic syndrome. Diabetes Care 2001, 24, 683–689. [Google Scholar] [CrossRef]
- Roberts, C.K.; Barnard, R.J.; Sindhu, R.K.; Jurczak, M.; Ehdaie, A.; Vaziri, N.D. Oxidative stress and dysregulation of NAD(P)H oxidase and antioxidant enzymes in diet-induced metabolic syndrome. Metabolism 2006, 55, 928–934. [Google Scholar] [CrossRef]
- Li, R.; Wang, W.Q.; Zhang, H.; Yang, X.; Fan, Q.; Christopher, T.A.; Lopez, B.L.; Tao, L.; Goldstein, B.J.; Gao, F.; et al. Adiponectin improves endothelial function in hyperlipidemic rats by reducing oxidative/nitrative stress and differential regulation of eNOS/iNOS activity. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1703–E1708. [Google Scholar] [CrossRef]
- Armutcu, F.; Ataymen, M.; Atmaca, H.; Gurel, A. Oxidative stress markers, C-reactive protein and heat shock protein 70 levels in subjects with metabolic syndrome. Clin. Chem. Lab. Med. 2008, 46, 785–790. [Google Scholar]
- Li, C.B.; Li, X.X.; Chen, Y.G.; Gao, H.Q.; Bu, P.L.; Zhang, Y.; Ji, X.P. Huang-lian-jie-du-tang protects rats from cardiac damages induced by metabolic disorder by improving inflammation-mediated insulin resistance. PLoS One 2013, 8, e67530. [Google Scholar]
- Panchal, S.K.; Ward, L.; Brown, L. Ellagic acid attenuates high-carbohydrate, high-fat diet-induced metabolic syndrome in rats. Eur. J. Nutr. 2013, 52, 559–568. [Google Scholar] [CrossRef]
- Dandona, P.; Weinstock, R.; Thusu, K.; Abdel-Rahman, E.; Aljada, A.; Wadden, T. Tumor necrosis factor-α in sera of obese patients: Fall with weight loss. J. Clin. Endocrinol. Metab. 1998, 83, 2907–2910. [Google Scholar]
- Roberts, C.K.; Barnard, R.J.; Sindhu, R.K.; Jurczak, M.; Ehdaie, A.; Vaziri, N.D. A high-fat, refined-carbohydrate diet induces endothelial dysfunction and oxidant/antioxidant imbalance and depresses NOS protein expression. J. Appl. Physiol. 2005, 98, 203–210. [Google Scholar]
- Lee, M.K.; Kim, S.R.; Sung, S.H.; Lim, D.; Kim, H.; Choi, H.; Park, H.K.; Je, S.; Ki, Y.C. Asiatic acid derivatives protect cultured cortical neurons from glutamate-induced excitotoxicity. Res. Commun. Mol. Pathol. Pharmacol. 2000, 108, 75–86. [Google Scholar]
- Mook-Jung, I.; Shin, J.E.; Yun, S.H.; Huh, K.; Koh, J.Y.; Park, H.K.; Jew, S.S.; Jung, M.W. Protective effects of asiaticoside derivatives against β-amyloid neurotoxicity. J. Neurosci. Res. 1999, 58, 417–425. [Google Scholar] [CrossRef]
- Huang, S.S.; Chiu, C.S.; Chen, H.J.; Hou, W.C.; Sheu, M.J.; Lin, Y.C.; Shie, P.H.; Huang, G.J. Antinociceptive activities and the mechanisms of anti-inflammation of asiatic acid in mice. Evid. Based Complement. Altern. Med. 2011, 2011, 895857. [Google Scholar]
- Ramachandran, V.; Saravanan, R. Efficacy of asiatic acid, a pentacyclic triterpene on attenuating the key enzymes activities of carbohydrate metabolism in streptozotocin-induced diabetic rats. Phytomedicine 2013, 20, 230–236. [Google Scholar] [CrossRef]
- Ramachandran, V.; Saravanan, R.; Senthilraja, P. Antidiabetic and antihyperlipidemic activity of asiatic acid in diabetic rats, role of HMG CoA: In vivo and in silico approaches. Phytomedicine 2013. [Google Scholar] [CrossRef]
- Poudyal, H.; Panchal, S.K.; Ward, L.C.; Waanders, J.; Brown, L. Chronic high-carbohydrate, high-fat feeding in rats induces reversible metabolic, cardiovascular, and liver changes. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1472–E1482. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Luangaram, S.; Kukongviriyapan, U.; Pakdeechote, P.; Kukongviriyapan, V.; Pannangpetch, P. Protective effects of quercetin against phenylhydrazine-induced vascular dysfunction and oxidative stress in rats. Food Chem. Toxicol. 2007, 45, 448–455. [Google Scholar] [CrossRef]
- Nakmareong, S.; Kukongviriyapan, U.; Pakdeechote, P.; Donpunha, W.; Kukongviriyapan, V.; Kongyingyoes, B.; Sompamit, K.; Phisalaphong, C. Antioxidant and vascular protective effects of curcumin and tetrahydrocurcumin in rats with l-NAME-induced hypertension. Naunyn Schmiedebergs Arch. Pharmacol. 2011, 383, 519–529. [Google Scholar] [CrossRef]
- Liu, J.; He, T.; Lu, Q.; Shang, J.; Sun, H.; Zhang, L. Asiatic acid preserves β cell mass and mitigates hyperglycemia in streptozocin-induced diabetic rats. Diabetes Metab. Res. Rev. 2010, 26, 448–454. [Google Scholar] [CrossRef]
- Basu, A.; Sanchez, K.; Leyva, M.J.; Wu, M.; Betts, N.M.; Aston, C.E.; Lyons, T.J. Green tea supplementation affects body weight, lipids, and lipid peroxidation in obese subjects with metabolic syndrome. J. Am. Coll. Nutr. 2010, 29, 31–40. [Google Scholar] [CrossRef]
- Meng, R.; Zhu, D.L.; Bi, Y.; Yang, D.H.; Wang, Y.P. Anti-oxidative effect of apocynin on insulin resistance in high-fat diet mice. Ann. Clin. Lab. Sci. 2011, 41, 236–243. [Google Scholar]
- Alam, M.A.; Kauter, K.; Withers, K.; Sernia, C.; Brown, L. Chronic l-arginine treatment improves metabolic, cardiovascular and liver complications in diet-induced obesity in rats. Food Funct. 2013, 4, 83–91. [Google Scholar] [CrossRef]
- Wei, J.; Huang, Q.; Huang, R.; Chen, Y.; Lv, S.; Wei, L.; Liang, C.; Liang, S.; Zhuo, L.; Lin, X. Asiatic acid from Potentilla chinensis attenuate ethanol-induced hepatic injury via suppression of oxidative stress and Kupffer cell activation. Biol. Pharm. Bull. 2013, 36, 1980–1989. [Google Scholar] [CrossRef]
- Hansen, K.; Nyman, U.; Smitt, U.W.; Adsersen, A.; Gudiksen, L.; Rajasekharan, S.; Pushpangadan, P. In vitro screening of traditional medicines for anti-hypertensive effect based on inhibition of the angiotensin converting enzyme (ACE). J. Ethnopharmacol. 1995, 48, 43–51. [Google Scholar] [CrossRef]
- Diwan, V.; Poudyal, H.; Brown, L. Piperine attenuates cardiovascular, liver and metabolic changes in high carbohydrate, high fat-fed rats. Cell Biochem. Biophys. 2013, 67, 297–304. [Google Scholar] [CrossRef]
- Hwang, I.S.; Ho, H.; Hoffman, B.B.; Reaven, G.M. Fructose-induced insulin resistance and hypertension in rats. Hypertension 1987, 10, 512–516. [Google Scholar] [CrossRef]
- Zhang, X.; Yan, S.; Zheng, H.; Hu, D.; Zhang, Y.; Guan, Q.; Ding, Q. A mechanism underlying hypertensive occurrence in the metabolic syndrome: Cooperative effect of oxidative stress and calcium accumulation in vascular smooth muscle cells. Horm. Metab. Res. 2013. [Google Scholar] [CrossRef]
- Yang, R.L.; Li, W.; Shi, Y.H.; Le, G.W. Lipoic acid prevents high-fat diet-induced dyslipidemia and oxidative stress: A microarray analysis. Nutrition 2008, 24, 582–588. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar]
- Ihm, S.H.; Jang, S.W.; Kim, O.R.; Chang, K.; Oak, M.H.; Lee, J.O.; Lim, D.Y.; Kim, J.H. Decaffeinated green tea extract improves hypertension and insulin resistance in a rat model of metabolic syndrome. Atherosclerosis 2012, 224, 377–383. [Google Scholar] [CrossRef]
- Montefusco-Pereira, C.V.; de Carvalho, M.J.; de Araujo Boleti, A.P.; Teixeira, L.S.; Matos, H.R.; Lima, E.S. Antioxidant, anti-inflammatory, and hypoglycemic effects of the leaf extract from Passiflora nitida Kunth. Appl. Biochem. Biotechnol. 2013, 170, 1367–1378. [Google Scholar] [CrossRef]
- Bradley, J.R. TNF-mediated inflammatory disease. J. Pathol. 2008, 214, 149–160. [Google Scholar]
- Huang, H.Y.; Korivi, M.; Tsai, C.H.; Yang, J.H.; Tsai, Y.C. Supplementation of lactobacillus plantarum K68 and fruit-vegetable ferment along with high fat-fructose diet attenuates metabolic syndrome in rats with insulin resistance. Evid. Based Complement. Altern. Med. 2013, 2013. [Google Scholar] [CrossRef]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Investig. 2003, 112, 1821–1830. [Google Scholar]
- Kroncke, K.D.; Fehsel, K.; Kolb-Bachofen, V. Inducible nitric oxide synthase in human diseases. Clin. Exp. Immunol. 1998, 113, 147–156. [Google Scholar] [CrossRef]
- Desai, N.; Roman, A.; Rochelson, B.; Gupta, M.; Xue, X.; Chatterjee, P.K.; Tam Tam, H.; Metz, C.N. Maternal metformin treatment decreases fetal inflammation in a rat model of obesity and metabolic syndrome. Am. J. Obstet. Gynecol. 2013, 209, e1–e9. [Google Scholar]
- Brevetti, G.; Schiano, V.; Sirico, G.; Giugliano, G.; Laurenzano, E.; Chiariello, M. Metabolic syndrome in peripheral arterial disease: Relationship with severity of peripheral circulatory insufficiency, inflammatory status, and cardiovascular comorbidity. J. Vasc. Surg. 2006, 44, 101–107. [Google Scholar] [CrossRef]
- Richard, C.; Couture, P.; Desroches, S.; Lamarche, B. Effect of the Mediterranean diet with and without weight loss on markers of inflammation in men with metabolic syndrome. Obesity 2013, 21, 51–57. [Google Scholar] [CrossRef]
- Yun, K.J.; Kim, J.Y.; Kim, J.B.; Lee, K.W.; Jeong, S.Y.; Park, H.J.; Jung, H.J.; Cho, Y.W.; Yun, K.; Lee, K.T. Inhibition of LPS-induced NO and PGE2 production by asiatic acid via NF-κB inactivation in RAW 264.7 macrophages: Possible involvement of the IKK and MAPK pathways. Int. Immunopharmacol. 2008, 8, 431–441. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pakdeechote, P.; Bunbupha, S.; Kukongviriyapan, U.; Prachaney, P.; Khrisanapant, W.; Kukongviriyapan, V. Asiatic Acid Alleviates Hemodynamic and Metabolic Alterations via Restoring eNOS/iNOS Expression, Oxidative Stress, and Inflammation in Diet-Induced Metabolic Syndrome Rats. Nutrients 2014, 6, 355-370. https://doi.org/10.3390/nu6010355
Pakdeechote P, Bunbupha S, Kukongviriyapan U, Prachaney P, Khrisanapant W, Kukongviriyapan V. Asiatic Acid Alleviates Hemodynamic and Metabolic Alterations via Restoring eNOS/iNOS Expression, Oxidative Stress, and Inflammation in Diet-Induced Metabolic Syndrome Rats. Nutrients. 2014; 6(1):355-370. https://doi.org/10.3390/nu6010355
Chicago/Turabian StylePakdeechote, Poungrat, Sarawoot Bunbupha, Upa Kukongviriyapan, Parichat Prachaney, Wilaiwan Khrisanapant, and Veerapol Kukongviriyapan. 2014. "Asiatic Acid Alleviates Hemodynamic and Metabolic Alterations via Restoring eNOS/iNOS Expression, Oxidative Stress, and Inflammation in Diet-Induced Metabolic Syndrome Rats" Nutrients 6, no. 1: 355-370. https://doi.org/10.3390/nu6010355