1. Introduction
Given the variety and abundance of foods available in the marketplace, eating is an enjoyable life experience. For some individuals, however, consuming certain foods can be a pathological, possibly life-threatening event. This is why the Roman philosopher Lucretius said, many years ago, that one man’s food may be another man’s poison. Such is the case with food allergies and sensitivities [
1]. The discussion about food allergy and sensitivity, in particular with regards to gluten and casein, and associated health problems has grown over the last decade. Classical food allergies are becoming more prevalent and recognized in our society. It is estimated that 5%–6% of children and 3%–4% of adults may have IgE-mediated or immediate type hypersensitivity to various food antigens [
2,
3]. A meta-analysis of 57 articles from different countries examined the prevalence of food allergies by using different criteria. Depending on food antigens, location, and gender, the authors found marked heterogeneity between studies [
4,
5,
6,
7,
8].
The big eight; milk, wheat, eggs, peanuts, fish, crustacean, tree nuts, and soy beans, are thought to account for more than 90% of all IgE-mediated food allergies in the USA, as well as on a worldwide basis [
1,
5]. Estimates for food sensitivities that are not IgE-mediated are more difficult to determine, since percentages may vary from one food antigen to another, and even from gender to gender and location to location [
7,
8]. For example, reaction to various wheat proteomes could be involved in three distinct conditions called wheat allergy, celiac disease (CD) and non-celiac gluten sensitivity (NCGS) [
9,
10,
11]. In these conditions, the reaction to gluten is mediated by both cellular and humoral immune responses, resulting in the presentation of different symptomatologies. In fact, in wheat allergy a specific sequence of gliadin peptides cross-links two IgE molecules on the surface of mast cells and basophils that trigger the release of mediators, such as histamines and leukotrienes [
12].
Celiac disease is an autoimmune condition with known genetic makeup and environmental triggers, such as gliadin peptides. CD affects between 1% and 2% of the general population. Markers for confirming a diagnosis of this disorder are IgA against native, deamidated gliadin peptides, and IgA anti-tissue transglutaminase (tTg) autoantibody. In comparison with CD, NCGS may affect 6% to 7% of the population, [
13,
14]. According to two articles published in 2010 and 2011 by Sapone
et al. [
13,
14], symptoms in GS may resemble some of the gastrointestinal symptoms that are associated with CD or wheat allergy, but it is emphasized that objective diagnostic tests for nonceliac gluten sensitivity are currently missing [
13,
14].
Very recently, in a case series study conducted in our laboratory, we attempted to use ELISA methodology to differentiate between CD and NCGS [
15]. The results brought us to contemplate that continuous exposure to environmental factors, such as wheat antigen-induced inflammation for a prolonged period of time, may result in inflammatory bowel disease or Crohn’s disease.
Indeed, it has been shown that both CD and NCGS can result in many autoimmune conditions, including type 1 diabetes, arthritis, thyroiditis, and even neuroautoimmune conditions, such as gluten ataxia and multiple sclerosis (MS) [
15,
16,
17,
18].
Nutritionists through the years seem to agree that milk is one of the most basic necessities of a healthy diet. However, unfortunately, cow’s milk proteins are the most common food allergens affecting young infants and some children and adults [
19]. Major allergenic proteins of milk are α-casein, β-casein, κ-casein, and β-lactoglobulin. In addition to IgE-mediated cow’s milk allergy there is increasing awareness that early consumption of cow’s milk may be a risk for the development of autoimmune diseases such as celiac disease, Crohn’s disease, Behçets disease, MS, mild rheumatoid arthritis in rabbits, and type 1 diabetes in humans [
20,
21,
22,
23,
24,
25]. These findings are supported by the detection of significantly higher levels of IgG and IgA antibodies in disease sufferers compared to normal controls. In these studies it was concluded that active immune response against cow’s milk proteins plays a putative role in the pathogenesis of autoimmune disorders [
7,
20,
21,
22,
23,
24,
25]. Despite these elevations in the IgG and IgA antibodies against wheat and milk proteins in blood samples of patients with various autoimmune disorders, the immunopathological role of these antibodies were not examined in the context of celiac disease in the IgE and non-IgE mediated reactivities [
9,
10,
26,
27,
28,
29]. Furthermore, in the non-IgE-mediated delayed wheat and milk reactivities, only IgG and IgA were studied and the role of IgM was completely ignored [
15,
26,
30]. Therefore, the aims of this study were threefold: (1) to examine the frequency of IgG-, IgA- and IgM-specific antibodies produced against wheat and milk proteins; (2) to examine the co-occurrence of these antibodies in sera with antibody elevation against glutamic acid decarboxylase (GAD-65), myelin basic protein (MBP), cerebellar peptide, and myelin oligodendrocytes glycoprotein (MOG); and (3) to discuss the mechanism of action of these antibodies in various neuroautoimmune reactivities in a subgroup of healthy controls.
3. Results
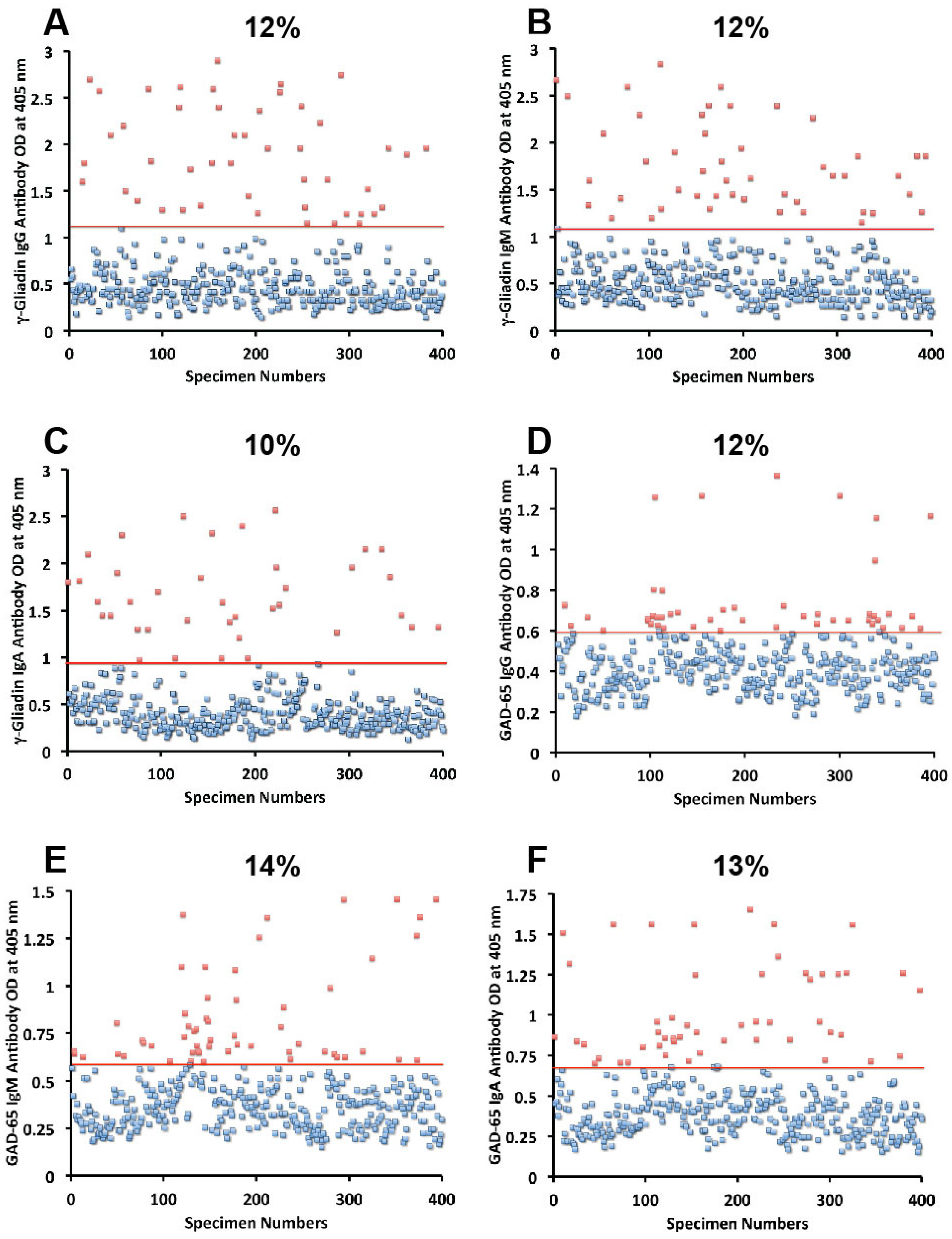
Sera from 400 blood donors were measured for the simultaneous presence of IgG, IgM, and IgA antibodies against wheat, α-gliadin 33-mer, γ-gliadin 15-mer and 18-mer, milk, α + β-casein, milk butyrophilin, GAD-65, cerebellar, MBP, and MOG. Results expressed as OD at 405 nm in the form of scattergrams are shown in
Figure 1,
Figure 2,
Figure 3,
Figure 4 and
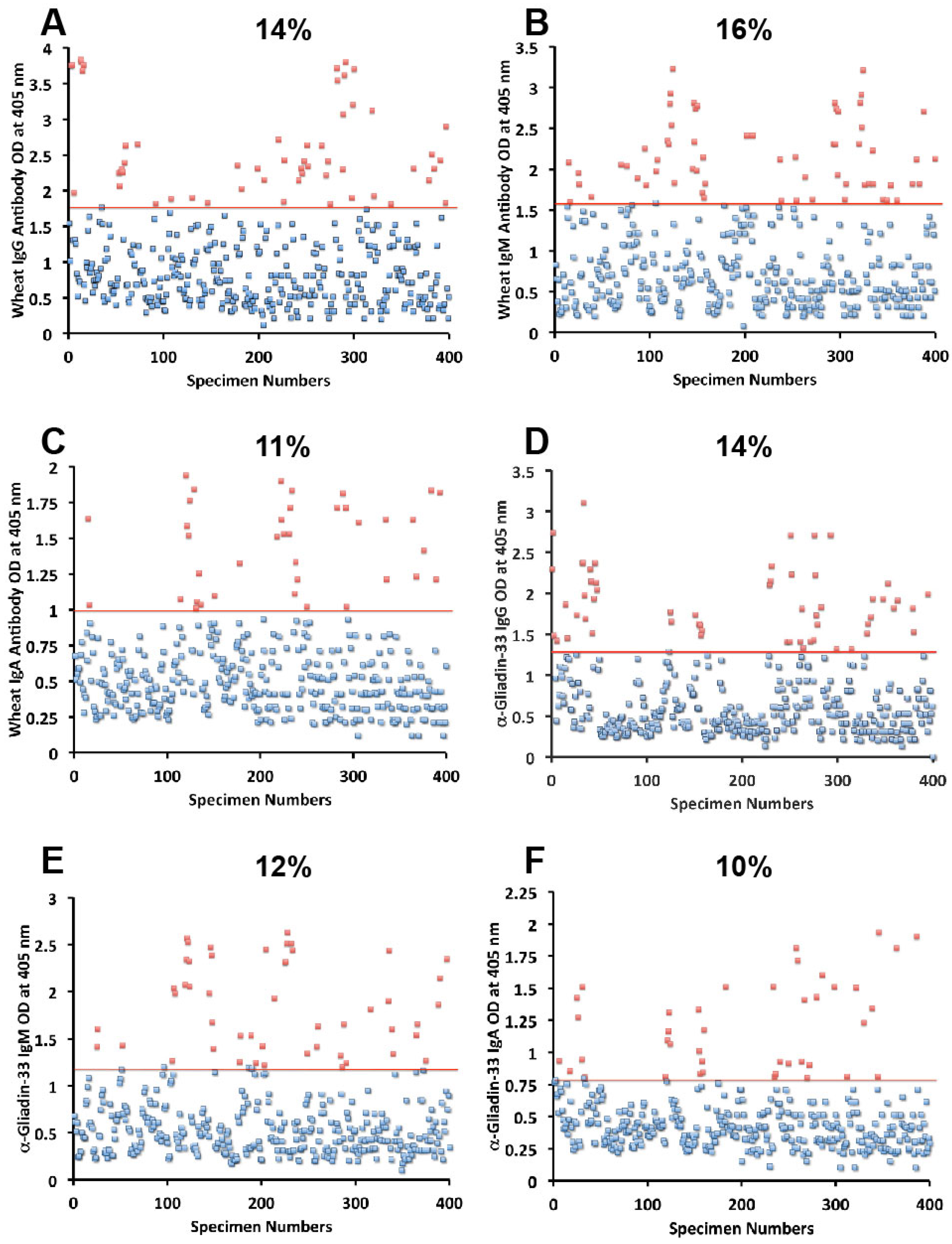
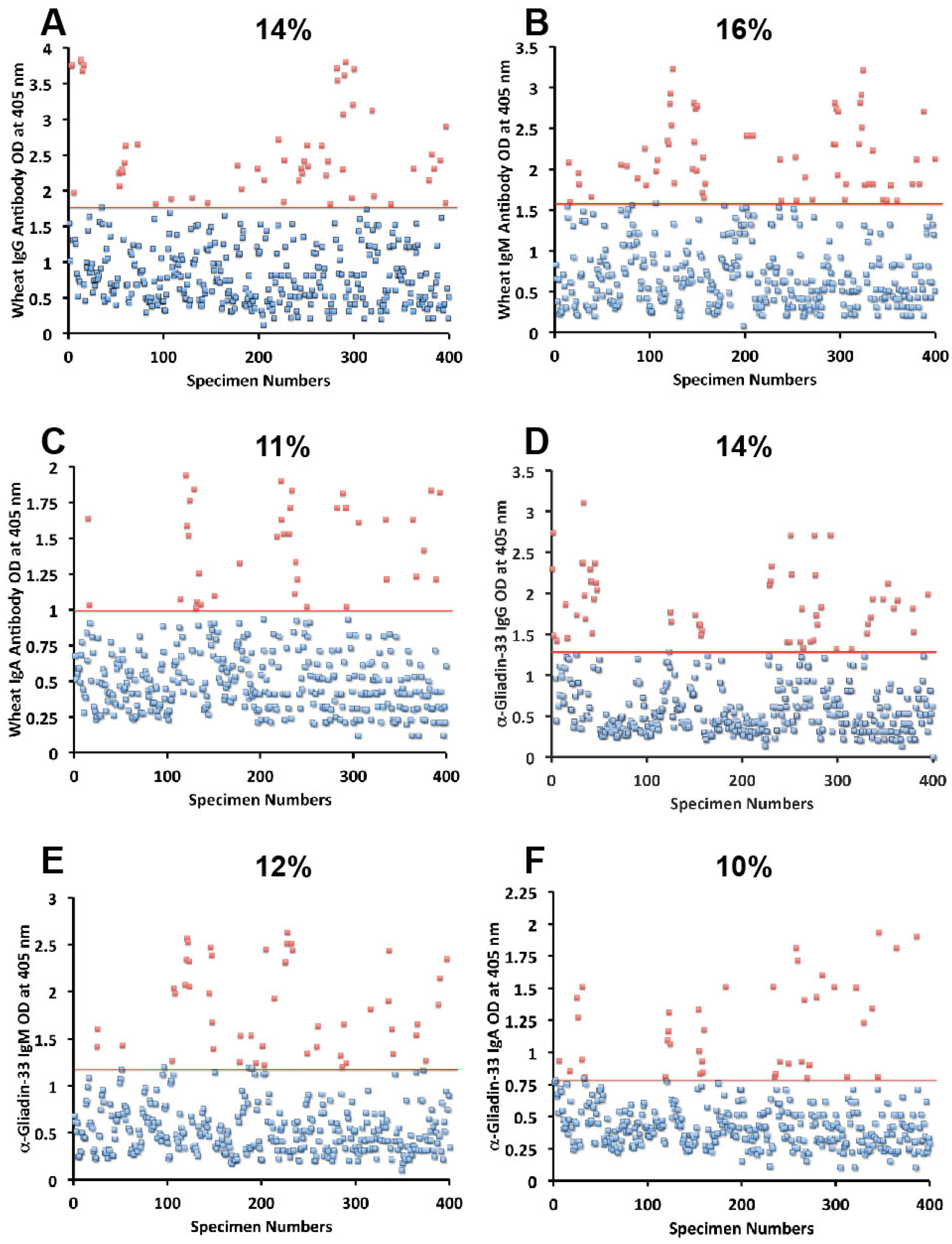
Figure 5. The OD for IgG antibody levels against wheat ranged from 0.1 to 3.8 with mean value of 1.0. At two standard deviations above the mean or OD of 1.8, 14% of individuals exhibited IgG antibody against wheat proteins (
Figure 1A). The mean OD of IgM anti-wheat was 0.92 and % elevation was 16 (
Figure 1B), while for wheat IgA antibody the mean OD was 0.6 with only 11% elevation (
Figure 1C). Levels of these antibodies against α-gliadin 33-mer peptide expressed by ODs were from 0.1 to 3.1 with mean 0.74 (
Figure 1D) for IgG, 0.1–2.6 with mean 0.68 for IgM (
Figure 1E), and from 0.1 to 1.9 with mean 0.49 for IgA (
Figure 1F). The percentage elevation of antibodies against α-gliadin 33-mer were 14, 12, and 10 for IgG, IgM, and IgA, respectively. Regarding the γ-gliadin antibody, while the ODs varied from 0.14 to 2.9 at 2 SD above the mean, 12% for IgG, 12% for IgM, and 10% for IgA of the specimens exhibited antibody elevation (
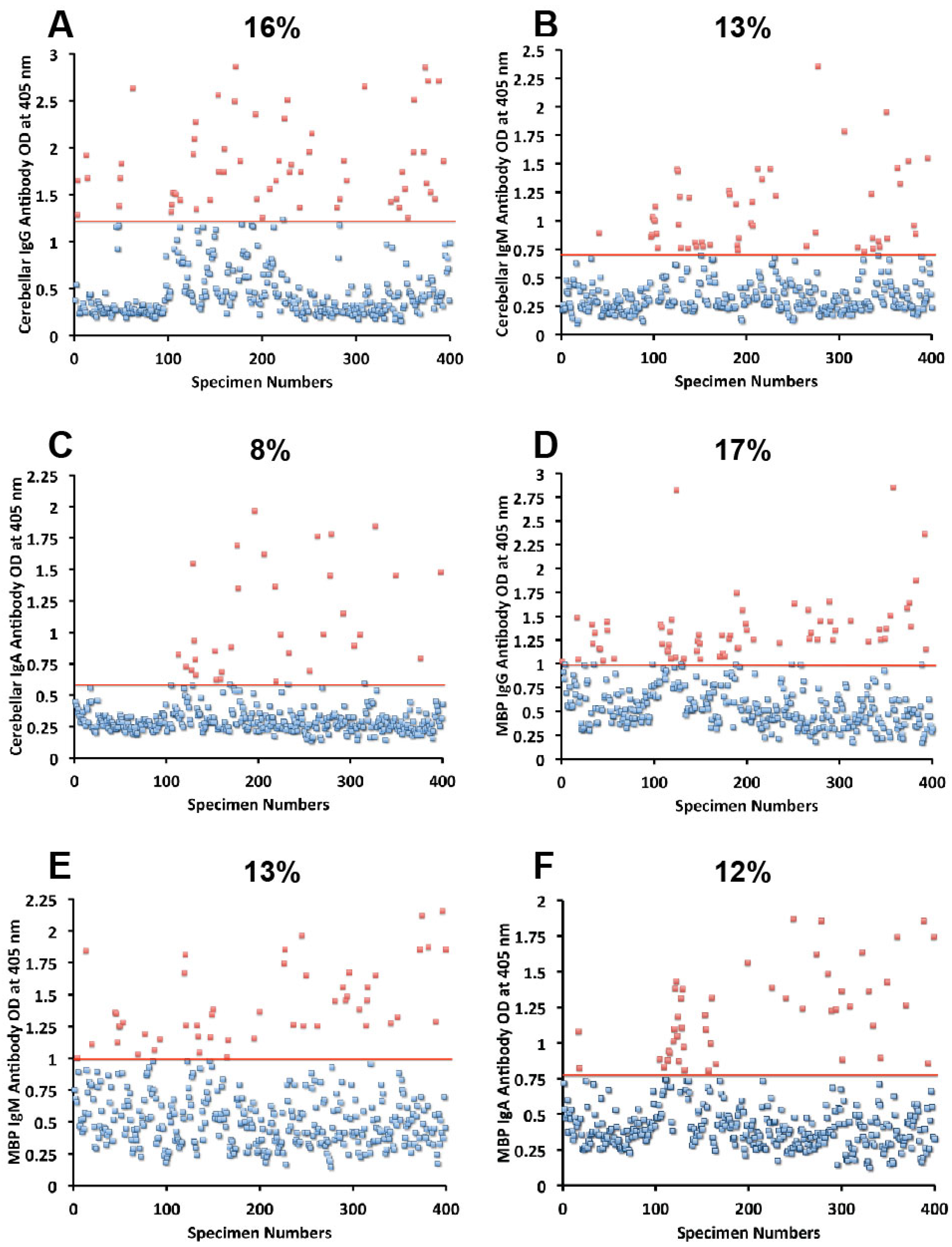
Figure 2A–C). The distribution of milk-, α + β-casein- and butyrophilin-specific antibody levels were as follows: for cow’s milk, elevations were IgG 13%, IgM 14%, and IgA 10%; (
Figure 4A–C). For α + β-casein, elevations for IgG and IgM were 10%, while IgA was 9% (
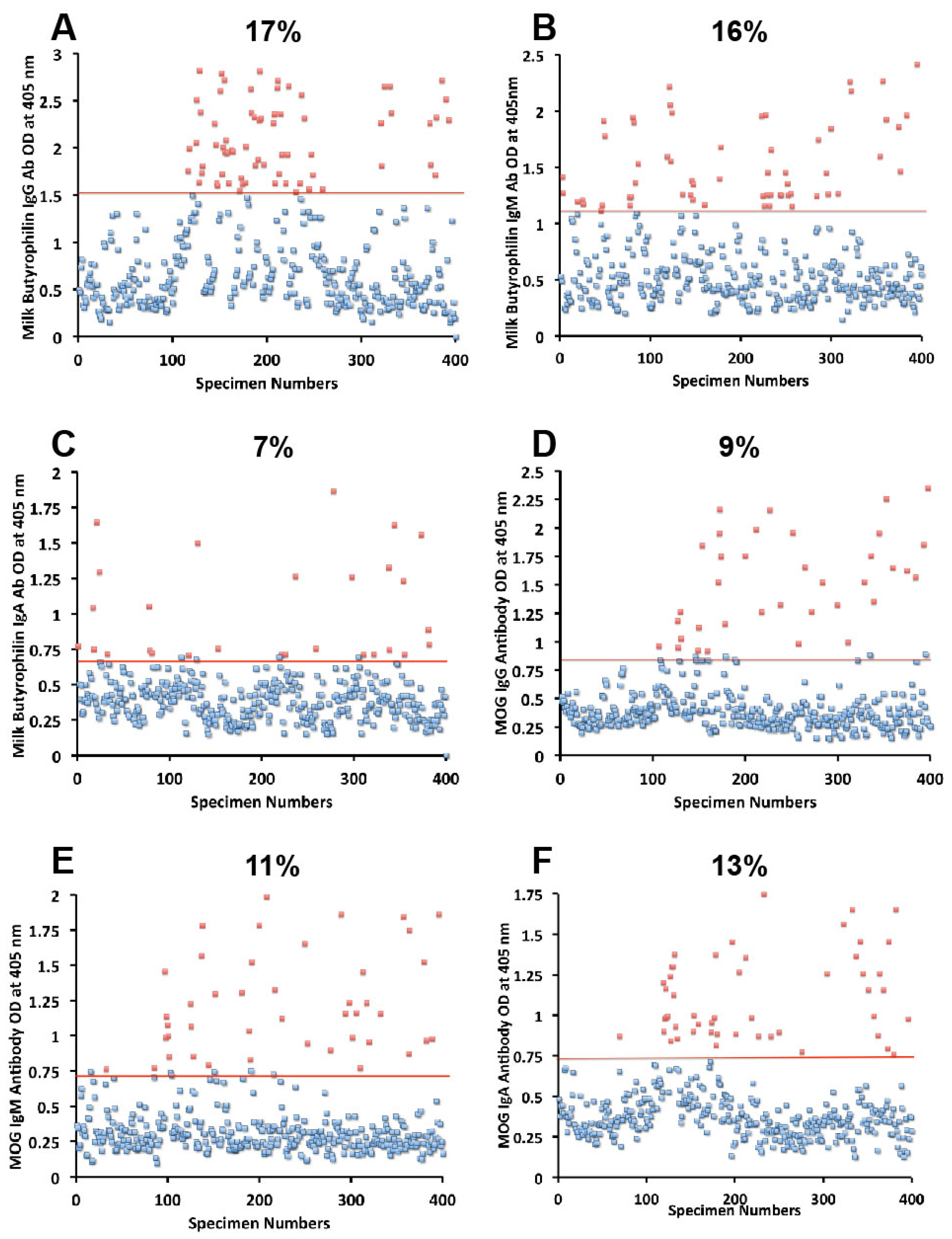
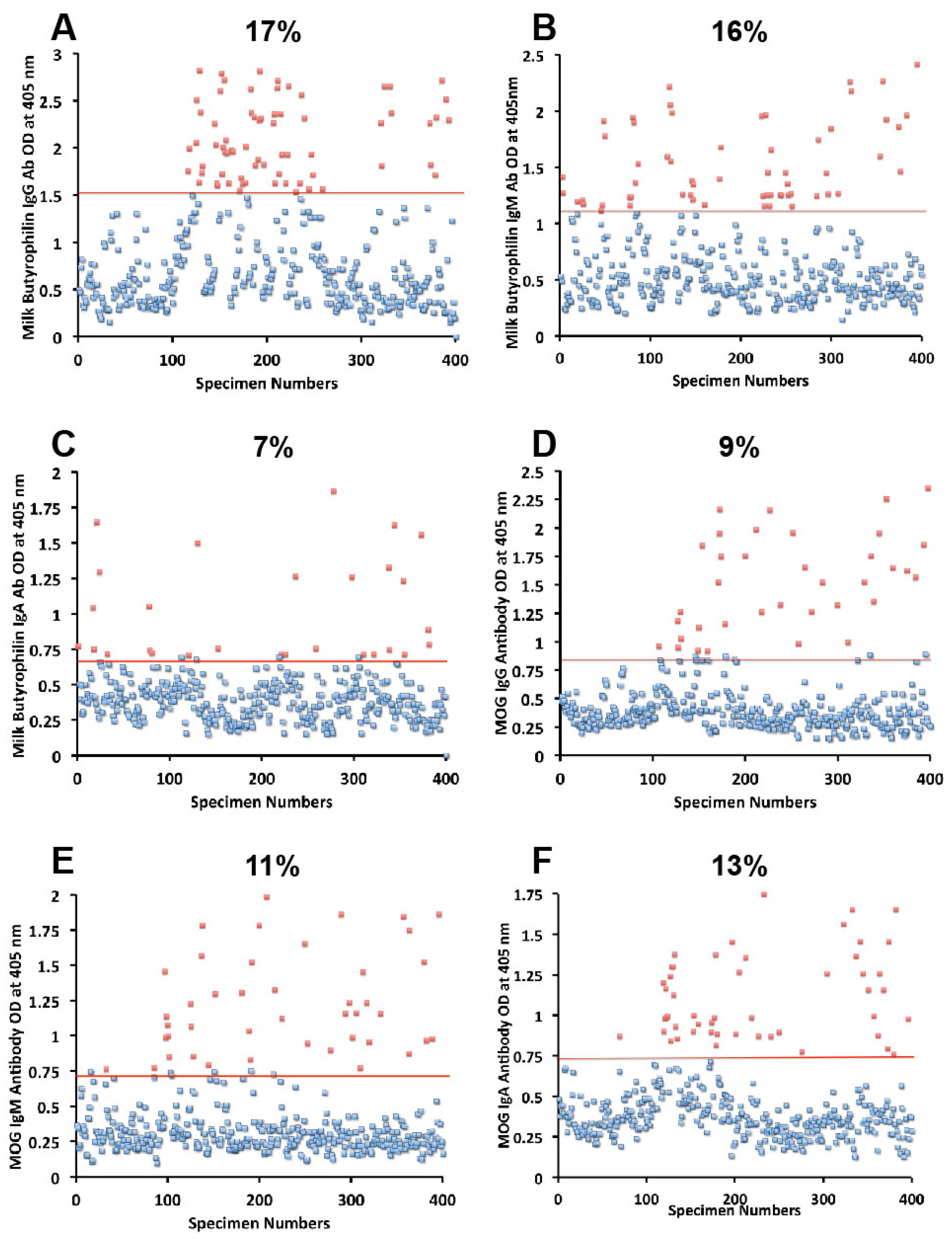
Figure 4D–F). And for milk butyrophilin, elevations were IgG 17%, IgM 16%, and IgA 7% (
Figure 5A–C).
Figure 1.
Results for wheat and α-gliadin-33 expressed as OD at 405 nm in the form of scattergrams with percentages of elevation. (A) Wheat IgG; (B) Wheat IgM; (C) Wheat IgA; (D) α-gliadin-33 IgG; (E) α-gliadin-33 IgM; (F) α-gliadin-33 IgA.
Figure 1.
Results for wheat and α-gliadin-33 expressed as OD at 405 nm in the form of scattergrams with percentages of elevation. (A) Wheat IgG; (B) Wheat IgM; (C) Wheat IgA; (D) α-gliadin-33 IgG; (E) α-gliadin-33 IgM; (F) α-gliadin-33 IgA.
Figure 2.
Results for γ-gliadin and GAD-65 expressed as OD at 405 nm in the form of scattergrams with percentages of elevation. (A) γ-gliadin IgG; (B) γ-gliadin IgM; (C) γ-gliadin IgA; (D) GAD-65 IgG; (E) GAD-65 IgM; (F) GAD-65 IgA.
Figure 2.
Results for γ-gliadin and GAD-65 expressed as OD at 405 nm in the form of scattergrams with percentages of elevation. (A) γ-gliadin IgG; (B) γ-gliadin IgM; (C) γ-gliadin IgA; (D) GAD-65 IgG; (E) GAD-65 IgM; (F) GAD-65 IgA.
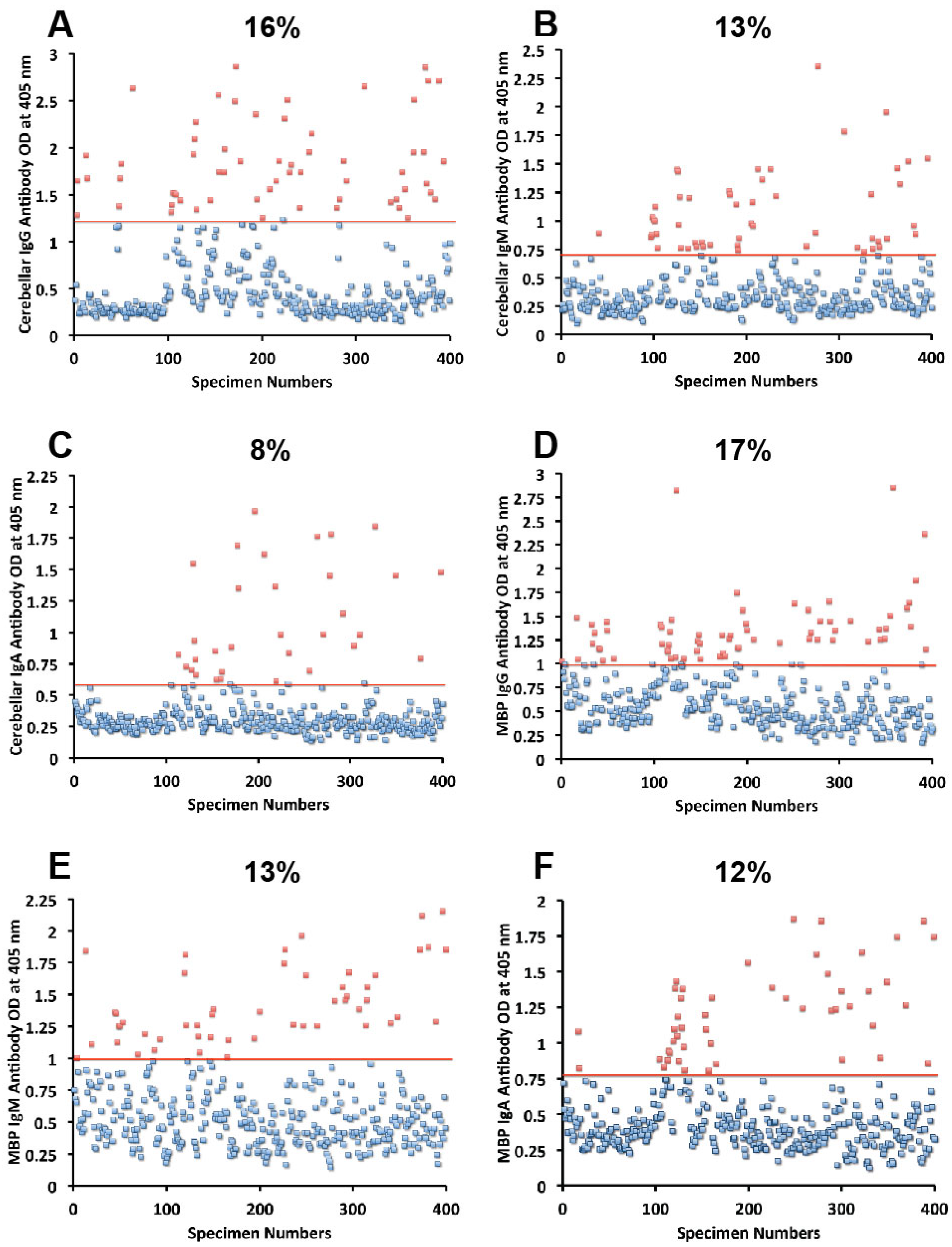
Figure 3.
Results for cerebellar and MBP expressed as OD at 405 nm in the form of scattergrams with percentages of elevation. (A) Cerebellar IgG; (B) Cerebellar IgM; (C) Cerebellar IgA; (D) MBP IgG; (E) MBP IgM; (F) MBP IgA.
Figure 3.
Results for cerebellar and MBP expressed as OD at 405 nm in the form of scattergrams with percentages of elevation. (A) Cerebellar IgG; (B) Cerebellar IgM; (C) Cerebellar IgA; (D) MBP IgG; (E) MBP IgM; (F) MBP IgA.
Figure 4.
Results for cow’s milk and α + β-casein expressed as OD at 405 nm in the form of scattergrams with percentages of elevation. (A) Cow’s milk IgG; (B) Cow’s milk IgM; (C) Cow’s milk IgA; (D) α + β-casein IgG; (E) α + β-casein IgM; (F) α + β-casein IgA.
Figure 4.
Results for cow’s milk and α + β-casein expressed as OD at 405 nm in the form of scattergrams with percentages of elevation. (A) Cow’s milk IgG; (B) Cow’s milk IgM; (C) Cow’s milk IgA; (D) α + β-casein IgG; (E) α + β-casein IgM; (F) α + β-casein IgA.
Figure 5.
Results for milk butyrophilin and MOG expressed as OD at 405 nm in the form of scattergrams with percentages of elevation. (A) Milk butyrophilin IgG; (B) Milk butyrophilin IgM; (C) Milk butyrophilin IgA; (D) MOG IgG; (E) MOG IgM; (F) MOG IgA.
Figure 5.
Results for milk butyrophilin and MOG expressed as OD at 405 nm in the form of scattergrams with percentages of elevation. (A) Milk butyrophilin IgG; (B) Milk butyrophilin IgM; (C) Milk butyrophilin IgA; (D) MOG IgG; (E) MOG IgM; (F) MOG IgA.
3.1. Measurement of Antibodies against Neural Antigens
Concomitant with the increase of IgG, IgM, and IgA antibodies against wheat, α- and γ-gliadin, milk, α + β-casein, and milk butyrophilin, in a small subgroup of individuals we observed a significant increase in GAD-65 (
Figure 2D–F), cerebellar peptide (
Figure 3A–C), MBP (
Figure 3D–F), and MOG antibodies (
Figure 5D–F). The percentage of elevation in antibodies against GAD-65, cerebellar, MBP and MOG varied from 9% to 17% for IgG, 11%–14% for IgM, and 8%–13% for IgA. Fifty-three out of 400 specimens (13%) showed IgG elevation against milk, and 68 or 17% had IgG elevation against MBP; about one third of these specimens positive for milk or MBP showed simultaneous elevation in IgG antibody against α + β-casein + MBP.
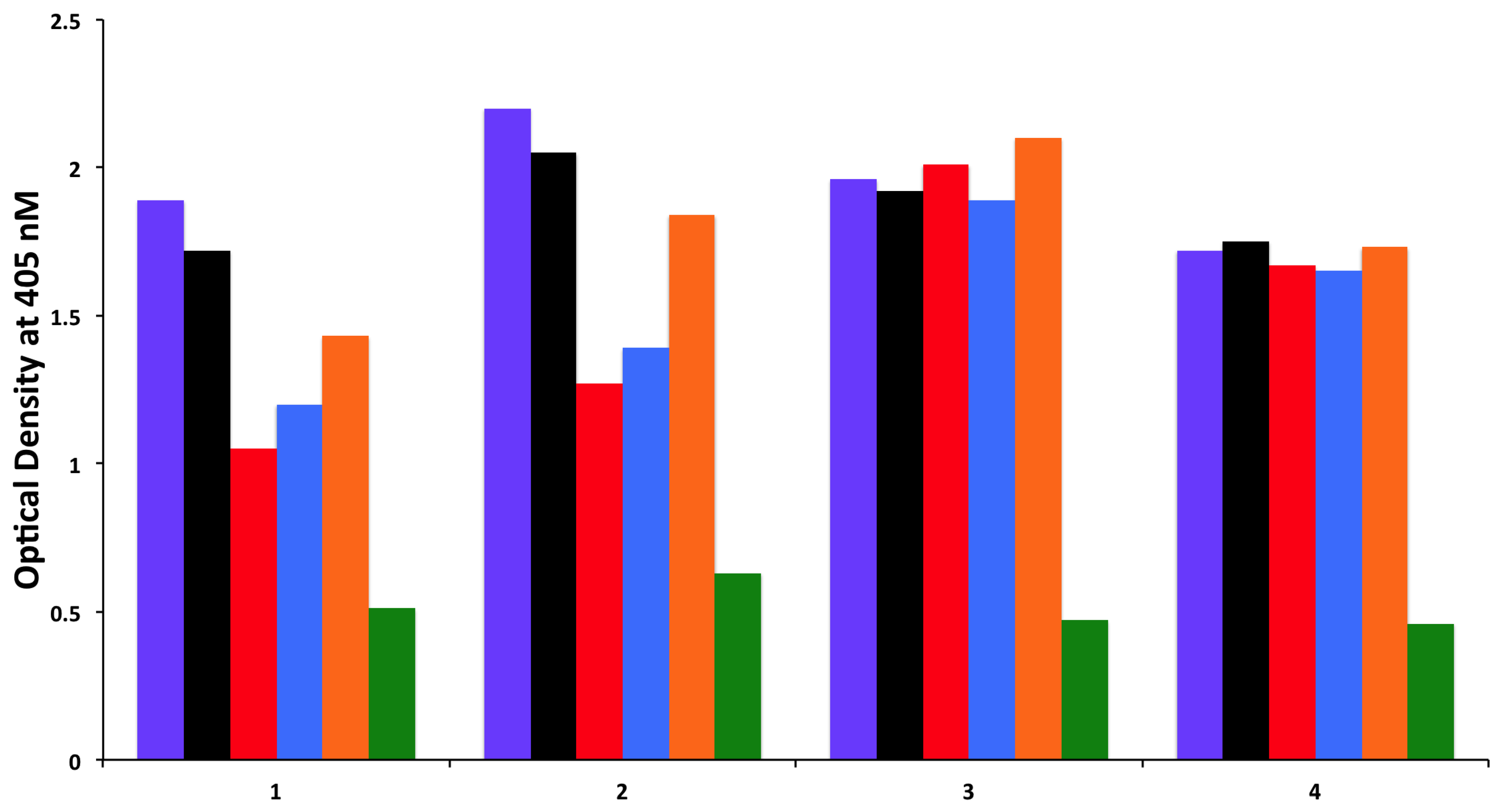
3.2. Absorption of Sera with High Titer of Antibody against Cerebellar or MOG with Specific and Non-Specific Antigens
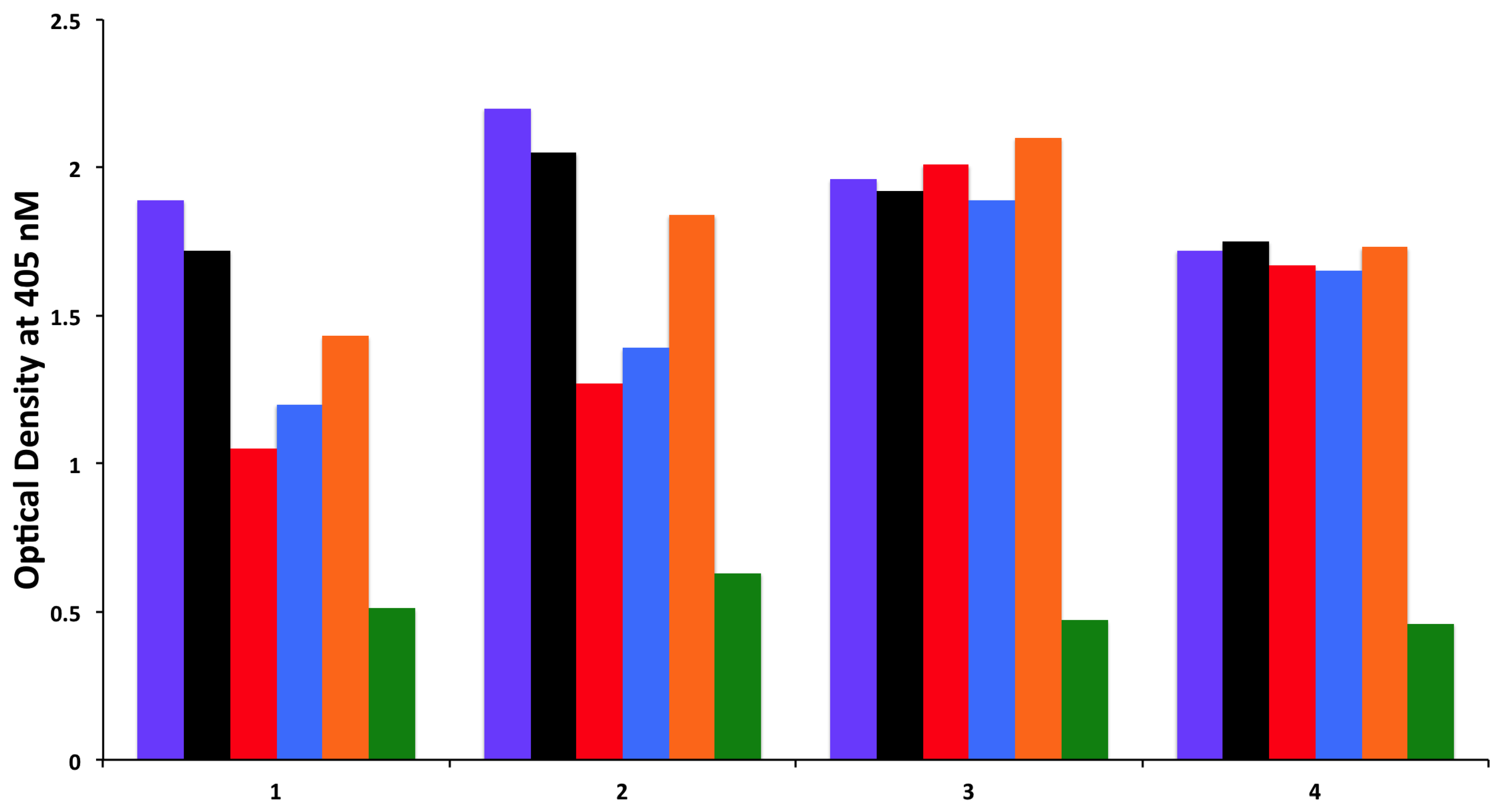
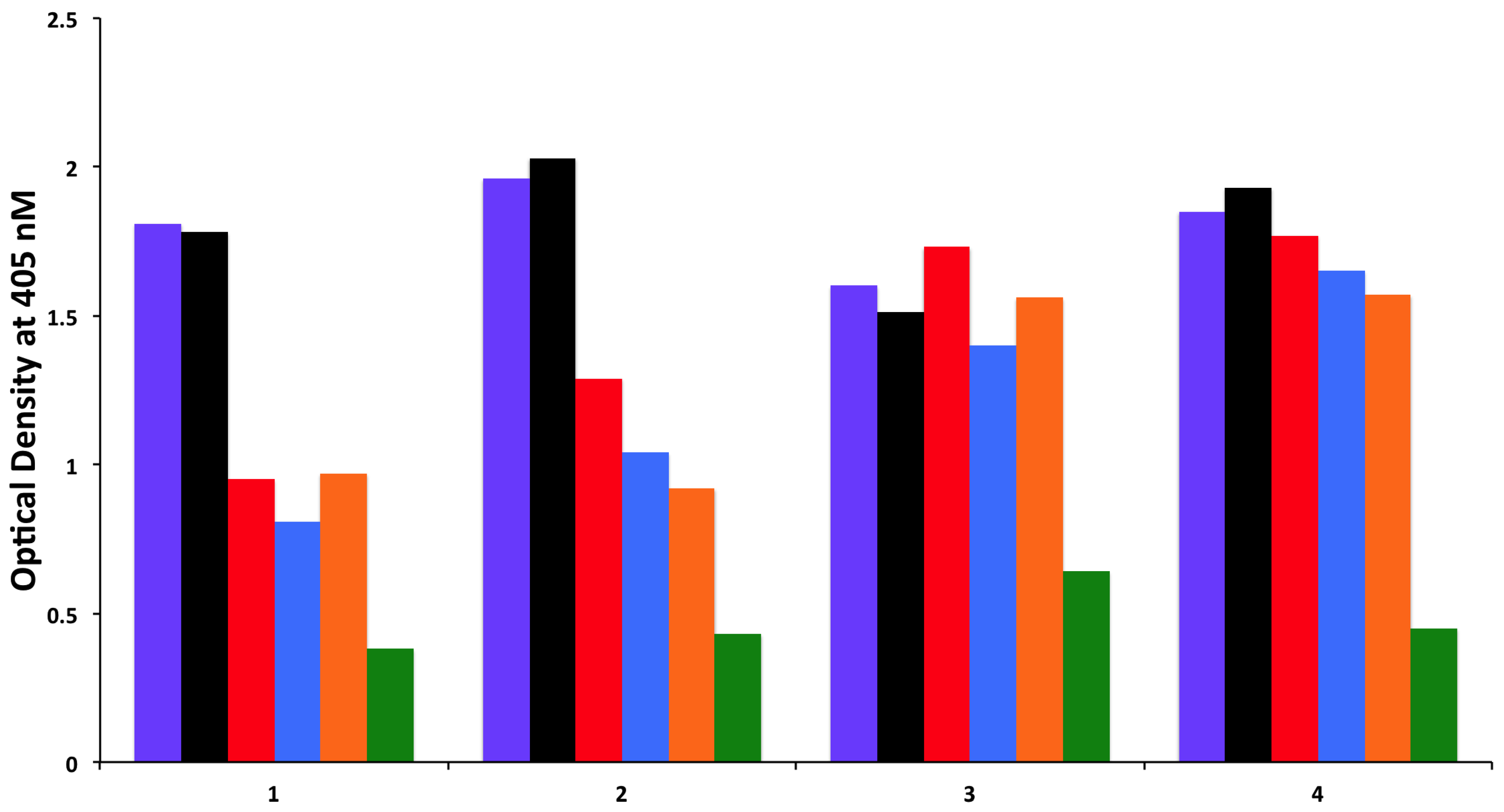
To examine whether antibodies detected against cerebellar are cross-reacting with wheat proteins and antibodies detected against MOG are cross-reacting with milk proteins, we performed an absorption study with non-specific antigens, specific antigens, and possible cross-reactive antigens. Four different sera with high levels of antibodies against cerebellar were subjected to the absorption study with HSA as a non-specific antigen, cerebellar peptide as a specific antigen, and wheat antigen, gliadin, and GAD-65 peptides as possible cross-reactive antigens. In all four of these sera, the high titers of cerebellar antibody were inhibited by more than 70% by the addition of cerebellar peptide. However, it was only in serum numbers 1 and 2 that the antibody levels were inhibited by 38%, 34%, and 23%, respectively, after the addition of wheat, gliadin, and GAD-65, resulting in
p values of 0.026 for wheat, 0.036 for gliadin, and 0.13 for GAD-65. Very similar results were obtained when sera with high levels of MOG antibody were subjected to absorption with HSA as a non-specific antigen, MOG peptide as a specific antigen, and milk, milk butyrophilin, and α + β-casein peptides as cross-reactive antigens. In all four of these sera, the high titers of MOG antibody were inhibited by more than 70% after the addition of MOG to the mixture. As was the case with the cerebellar sera, it was only in serum numbers 1 and 2 that the antibody levels were inhibited by about 40% by the addition of milk, α + β-casein and milk butyrophilin peptides to the mixture, resulting in
p values of 0.049 for milk, 0.014 for milk butyrophilin, and 0.016 for α + β-casein. Inhibition with the same antigens for the third serum was 20%–25%, and no inhibition at all was observed with the fourth serum (
Figure 6 and
Figure 7).
Figure 6.
Inhibition of immune reaction of sera containing high levels of IgG, IgM and IgA antibody against cerebellar before
![Nutrients 06 00015 i001]()
and after absorption with HSA
![Nutrients 06 00015 i002]()
, wheat
![Nutrients 06 00015 i003]()
, gliadin peptide
![Nutrients 06 00015 i004]()
, GAD-65
![Nutrients 06 00015 i005]()
and cerebellar peptide
![Nutrients 06 00015 i006]()
.
Figure 6.
Inhibition of immune reaction of sera containing high levels of IgG, IgM and IgA antibody against cerebellar before
![Nutrients 06 00015 i001]()
and after absorption with HSA
![Nutrients 06 00015 i002]()
, wheat
![Nutrients 06 00015 i003]()
, gliadin peptide
![Nutrients 06 00015 i004]()
, GAD-65
![Nutrients 06 00015 i005]()
and cerebellar peptide
![Nutrients 06 00015 i006]()
.
Sample numbers
Figure 7.
Inhibition of immune reaction of sera containing high levels of IgG, IgM and IgA antibody against MOG both before
![Nutrients 06 00015 i001]()
and after absorption with HSA
![Nutrients 06 00015 i002]()
, milk
![Nutrients 06 00015 i003]()
, milk butyrophilin
![Nutrients 06 00015 i004]()
, α + β-casein
![Nutrients 06 00015 i005]()
, and MOG
![Nutrients 06 00015 i006]()
.
Figure 7.
Inhibition of immune reaction of sera containing high levels of IgG, IgM and IgA antibody against MOG both before
![Nutrients 06 00015 i001]()
and after absorption with HSA
![Nutrients 06 00015 i002]()
, milk
![Nutrients 06 00015 i003]()
, milk butyrophilin
![Nutrients 06 00015 i004]()
, α + β-casein
![Nutrients 06 00015 i005]()
, and MOG
![Nutrients 06 00015 i006]()
.
Sample numbers
3.3. Statistical Analyses of the Data for Investigating Association between the Food Proteins and the Brain Proteins
Next, we tested whether there are significant associations between the elevations of lgG, lgA, and lgM isotypes of GAD-65, cerebellar, MBP, and MOG with similar isotypes of wheat, gliadin-33, γ-gliadin, cow’s milk, casein, and milk butyrophilin. We fitted simple linear regression models between each such pairs, and calculated the
R2 values and the
p-values. The summary of the results is presented in the
Table 1,
Table 2 and
Table 3. From the tables we see that several food proteins significantly elevates similar isotypes of some brain proteins. Specifically, considering statistically significant elevations only, we found that wheat lgG elevates MBP lgG, α-gliadin 33-mer lgG elevates GAD-65 lgG and MBP lgG, γ-gliadin lgG elevates GAD-65 lgG and MBP lgG, and milk butyrophilin lgG elevates cerebellar lgG, MBP lgG, and MOG lgG (
Table 3). We also found that wheat lgA elevates all four brain lgAs, α-gliadin 33-mer lgA elevates MBP lgA, cow’s milk lgA elevates GAD-65 lgA and MBP lgA, α + β-casein lgA elevates MBP lgA, milk butyrophilin lgA elevates GAD-65 lgA, cerebellar lgA, and MBP lgA (
Table 4). From
Table 5 we found that wheat lgM elevates GAD-65 lgM and MBP lgM, α-gliadin 33-mer lgM elevates GAD-65 lgM, cerebellar lgM and MBP lgM, γ-gliadin lgM elevates GAD-65 lgM, cow’s milk lgM elevates GAD-65 lgM and MBP lgM, α + β-casein lgM elevates cerebellar lgM and MOG lgM, while milk butyrophilin lgM elevates GAD-65 lgM and MBP lgM.
Table 1.
Results of the simple linear regression between lgG isotypes of food proteins and the brain proteins. The first number in each cell presents corresponding Pearson’s correlation coefficient and the second number presents its p-value. Small p-value (less than 0.05) indicates statistical significance. Note that R2 values of these regressions are the squares of the Pearson’s correlation coefficients.
Table 1.
Results of the simple linear regression between lgG isotypes of food proteins and the brain proteins. The first number in each cell presents corresponding Pearson’s correlation coefficient and the second number presents its p-value. Small p-value (less than 0.05) indicates statistical significance. Note that R2 values of these regressions are the squares of the Pearson’s correlation coefficients.
| Wheat & Milk Proteins | GAD-65 lgG | Cerebellar lgG | MBP lgG | MOG lgG |
|---|
| wheat lgG | 0.0172
0.7314 | −0.0078
0.8772 | 0.1025
0.0404
Significant | −0.0029
0.9534 |
| α-gliadin 33-mer lgG | 0.1116
0.0256
Significant | −0.0576
0.2500 | 0.1456
0.0035
Significant | −0.0024
0.9623 |
| γ-gliadin lgG | 0.1251
0.0123
Significant | 0.0323
0.5193 | 0.1479
0.0030
Significant | 0.0051
0.9192 |
| cow’s milk lgG | 0.0932
0.0626 | 0.0911
0.0689 | 0.0595
0.2350 | 0.0525
0.2949 |
| α + β-casein lgG | 0.0763
0.1280 | 0.0432
0.3897 | 0.0847
0.0912 | −0.0760
0.1296 |
| milk butyrophilin lgG | 0.0506
0.3128 | 0.2680
<0.0001
Significant | 0.1642
0.0010
Significant | 0.1618
0.0012
Significant |
Table 2.
Results of the simple linear regression between lgA isotypes of food proteins and the brain proteins. The first number in each cell presents corresponding Pearson’s correlation coefficient and the second number presents its p-value. Small p-value (less than 0.05) indicates statistical significance. Note that R2 values of these regressions are the squares of the Pearson’s correlation coefficients.
Table 2.
Results of the simple linear regression between lgA isotypes of food proteins and the brain proteins. The first number in each cell presents corresponding Pearson’s correlation coefficient and the second number presents its p-value. Small p-value (less than 0.05) indicates statistical significance. Note that R2 values of these regressions are the squares of the Pearson’s correlation coefficients.
| Wheat & Milk Proteins | GAD-65 lgA | Cerebellar lgA | MBP lgA | MOG lgA |
|---|
| wheat lgA | 0.2032
<0.0001
Significant | 0.1978
<0.0001
Significant | 0.2476
<0.0001
Significant | 0.2318
<0.0001
Significant |
| α-gliadin 33-mer lgA | 0.0874
0.0984 | 0.0446
0.3742 | 0.1757
0.0004
Significant | 0.0762
0.1279 |
| γ-gliadin lgA | 0.0517
0.3028 | −0.0044
0.9300 | 0.0381
0.4474 | 0.0166
0.7411 |
| cow’s milk lgA | 0.1401
0.0050
Significant | 0.0776
0.1213 | 0.1671
0.0008
Significant | 0.0912
0.0683 |
| α + β-casein lgA | 0.0346
0.4903 | 0.0226
0.6521 | 0.1157
0.0206
Significant | 0.0297
0.5537 |
| milk butyrophilin lgA | 0.1248
0.0125
Significant | 0.0994
0.0469
Significant | 0.1483
0.0029
Significant | 0.0710
0.1565 |
Table 3.
Results of the simple linear regression between lgM isotypes of food proteins and the brain proteins. The first number in each cell presents corresponding Pearson’s correlation coefficient and the second number presents its p-value. Small p-value (less than 0.05) indicates statistical significance. Note that R2 values of these regressions are the squares of the Pearson’s correlation coefficients.
Table 3.
Results of the simple linear regression between lgM isotypes of food proteins and the brain proteins. The first number in each cell presents corresponding Pearson’s correlation coefficient and the second number presents its p-value. Small p-value (less than 0.05) indicates statistical significance. Note that R2 values of these regressions are the squares of the Pearson’s correlation coefficients.
| Wheat & Milk Proteins | GAD-65 lgM | Cerebellar lgM | MBP lgM | MOG lgM |
|---|
| wheat lgM | 0.2032
<0.0001
Significant | −0.0196
0.6962 | 0.1931
0.0001
Significant | 0.0598
0.2328 |
| α-gliadin 33-mer lgM | 0.2181
<0.0001
Significant | 0.1244
0.0127
Significant | 0.2046
<0.0001
Significant | 0.0022
0.9653 |
| γ-gliadin lgM | 0.1133
0.0234
Significant | −0.0460
0.3587 | −0.0062
0.9017 | −0.0025
0.9610 |
| cow’s milk lgM | 0.2022
<0.0001
Significant | 0.0457
0.3620 | 0.2003
0.0001
Significant | 0.0619
0.2170 |
| α + β-casein lgM | −0.0174
0.7289 | 0.3445
<0.0001
Significant | −0.0211
0.6738 | 0.3178
<0.0001
Significant |
| milk butyrophilin lgM | 0.2759
<0.0001Significant | −0.0732
0.1442 | 0.2606
<0.0001
Significant | −0.0131
0.7946 |
Table 4.
Results of the simple linear regression between each pair of lgG isotype of the antigens. The first number in each cell presents corresponding Pearson’s correlation coefficient and the second number presents its p-value. Small p-value (less than 0.05) indicates statistical significance. Note that R2 values of these regressions are the squares of the Pearson’s correlation coefficients.
Table 4.
Results of the simple linear regression between each pair of lgG isotype of the antigens. The first number in each cell presents corresponding Pearson’s correlation coefficient and the second number presents its p-value. Small p-value (less than 0.05) indicates statistical significance. Note that R2 values of these regressions are the squares of the Pearson’s correlation coefficients.
| Wheat & Milk Proteins | Wheat lgG | α-Gliadin 33-Mer lgG | γ-Gliadin lgG | Cow’s Milk lgG | α + β-Casein lgG | Milk Butyrophilin lgG |
|---|
| wheat lgG | 1.0000
0.0000 | 0.0791
0.1141 | 0.1486
0.0029
Significant | 0.3408
<0.0001
Significant | 0.3203
<0.0001
Significant | −0.0683
0.1731 |
| α-gliadin 33-mer lgG | 0.0791
0.1141 | 1.0000
0.0000 | 0.4871
<0.0001
Significant | 0.2058
<0.0001
Significant | 0.1535
0.0021
Significant | 0.1002
0.0452
Significant |
| γ-gliadin lgG | 0.1486
0.0029
Significant | 0.4871
<0.0001
Significant | 1.0000
0.0000 | 0.2009
0.0001
Significant | 0.2165
<0.0001
Significant | 0.0954
0.0567 |
| cow’s milk lgG | 0.3408
<0.0001
Significant | 0.2058
<0.0001
Significant | 0.2009
0.0001
Significant | 1.0000
0.0000 | 0.4467
<0.0001
Significant | 0.1929
0.0001
Significant |
| α + β-casein lgG | 0.3203
<0.0001
Significant | 0.1535
0.0021
Significant | 0.2165
<0.0001
Significant | 0.4467
<0.0001
Significant | 1.0000
0.0000 | 0.0384
0.4449 |
| milk butyrophilin lgG | −0.0683
0.1731 | 0.1002
0.0452
Significant | 0.0954
0.0567 | 0.1929
0.0001
Significant | 0.0384
0.4449 | 1.0000
0.0000 |
Next, we performed simple regression analyses between each pair of antigens to check if there are tendencies that the same individuals have high titres to multiple antigens. The summary of the results is presented in
Table 4,
Table 5, and
Table 6. From the tables we see that several pairs of the antigens have significant correlations.
Table 5.
Results of the simple linear regression between each pair of lgA isotype of the antigens. The first number in each cell presents corresponding Pearson’s correlation coefficient and the second number presents its p-value. Small p-value (less than 0.05) indicates statistical significance. Note that R2 values of these regressions are the squares of the Pearson’s correlation coefficients.
Table 5.
Results of the simple linear regression between each pair of lgA isotype of the antigens. The first number in each cell presents corresponding Pearson’s correlation coefficient and the second number presents its p-value. Small p-value (less than 0.05) indicates statistical significance. Note that R2 values of these regressions are the squares of the Pearson’s correlation coefficients.
| Wheat & Milk Proteins | Wheat lgA | α-Gliadin 33-Mer lgA | γ-Gliadin lgA | Cow’s Milk lgA | α + β-Casein lgA | Milk Butyrophilin lgA |
|---|
| wheat lgA | 1.0000
0.0000 | 0.1451
0.0036
Significant | 0.1558
0.0018
Significant | 0.2994
<0.0001
Significant | 0.2667
<0.0001
Significant | 0.0701
0.1615 |
| α-gliadin 33-mer lgA | 0.1451
0.0036
Significant | 1.0000
0.0000 | 0.1602
0.0013
Significant | 0.0468
0.3507 | 0.0989
0.0482
Significant | 0.1067
0.0328
Significant |
| γ-gliadin lgA | 0.1558
0.0018
Significant | 0.1602
0.0013
Significant | 1.0000
0.0000 | 0.1037
0.0382
Significant | 0.2293
<0.0001
Significant | 0.1182
0.0180
Significant |
| cow’s milk lgA | 0.2994
<0.0001
Significant | 0.0468
0.3507 | 0.1037
0.0382
Significant | 1.0000
0.0000 | 0.4489
<0.0001
Significant | 0.1502
0.0026
Significant |
| α + β-casein lgA | 0.2667
<0.0001
Significant | 0.0989
0.0482
Significant | 0.2293
<0.0001
Significant | 0.4489
<0.0001
Significant | 1.0000
0.0000 | 0.1396
0.0052
Significant |
| milk butyrophilin lgA | 0.0701
0.1615 | 0.1067
0.0328
Significant | 0.1182
0.0180
Significant | 0.1502
0.0026
Significant | 0.1396
0.0052
Significant | 1.0000
0.0000 |
Table 6.
Results of the simple linear regression between each pair of lgM isotype of the antigens. The first number in each cell presents corresponding Pearson’s correlation coefficient and the second number presents its p-value. Small p-value (less than 0.05) indicates statistical significance. Note that R2 values of these regressions are the squares of the Pearson’s correlation coefficients.
Table 6.
Results of the simple linear regression between each pair of lgM isotype of the antigens. The first number in each cell presents corresponding Pearson’s correlation coefficient and the second number presents its p-value. Small p-value (less than 0.05) indicates statistical significance. Note that R2 values of these regressions are the squares of the Pearson’s correlation coefficients.
| Wheat & Milk Proteins | Wheat lgM | α-Gliadin 33-Mer lgM | γ-Gliadin lgM | Cow’s Milk lgM | α + β-Casein lgM | Milk Butyrophilin lgM |
|---|
| wheat lgM | 1.0000
0.0000 | 0.3712
<0.0001
Significant | 0.0712
0.1553 | 0.1625
0.0011
Significant | 0.0256
0.6094 | 0.3058
<0.0001
Significant |
| α-gliadin 33-mer lgM | 0.3712
<0.0001
Significant | 1.0000
0.0000 | 0.0099
0.8434 | 0.2339
<0.0001
Significant | 0.0124
0.8055 | 0.3027
<0.0001
Significant |
| γ-gliadin lgM | 0.0712
0.1553 | 0.0099
0.8434 | 1.0000
0.0000 | −0.0798
0.1109 | 0.0118
0.8146 | 0.0909
0.0694 |
| cow’s milk lgM | 0.1625
0.0011
Significant | 0.2339
<0.0001
Significant | −0.0798
0.1109 | 1.0000
0.0000 | −0.0147
0.7692 | 0.2035
<0.0001
Significant |
| α + β-casein lgM | 0.0256
0.6094 | 0.0124
0.8055 | 0.0118
0.8146 | −0.0147
0.7692 | 1.0000
0.0000 | −0.0376
0.4528 |
| milk butyrophilin lgM | 0.3058
<0.0001
Significant | 0.3027
<0.0001
Significant | 0.0909
0.0694 | 0.2035
<0.0001
Significant | −0.0376
0.4528 | 1.0000
0.0000 |
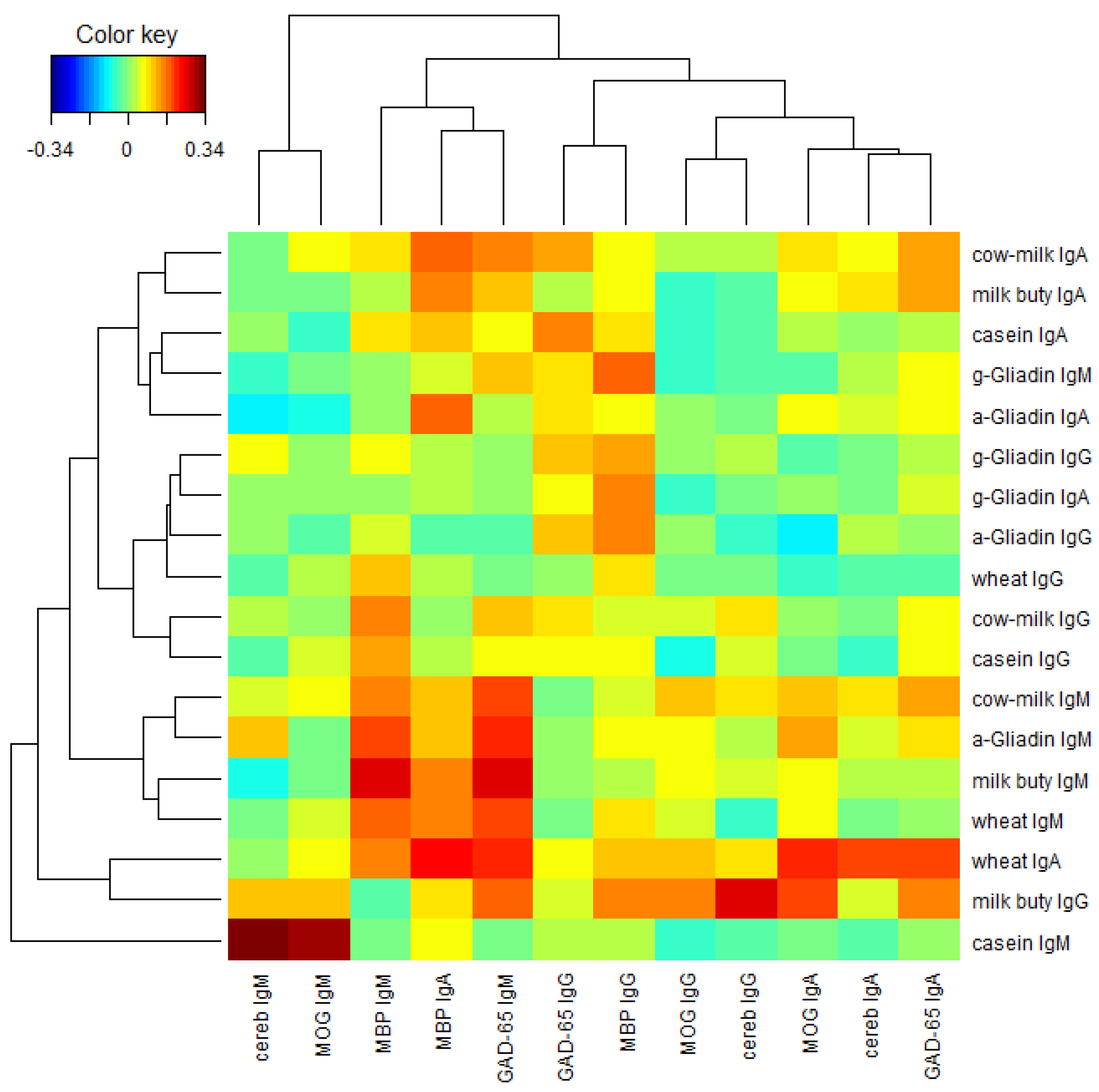
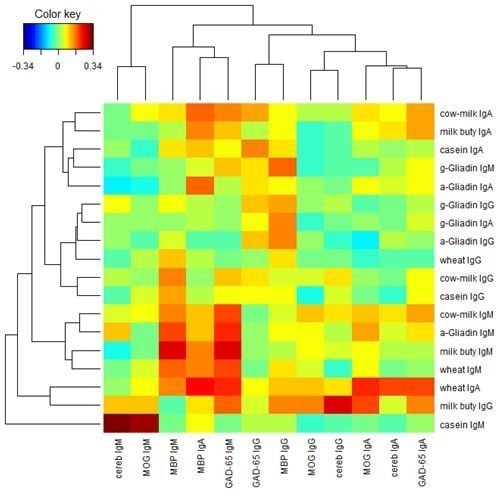
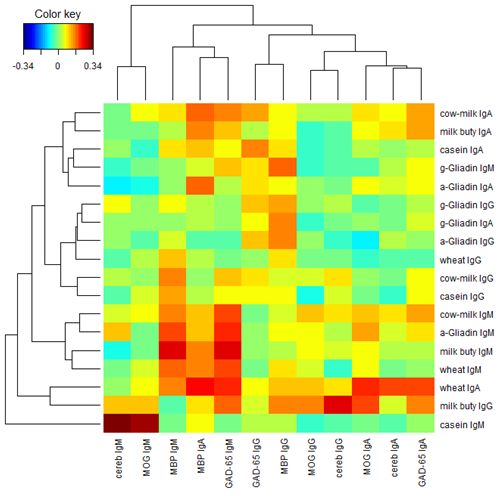
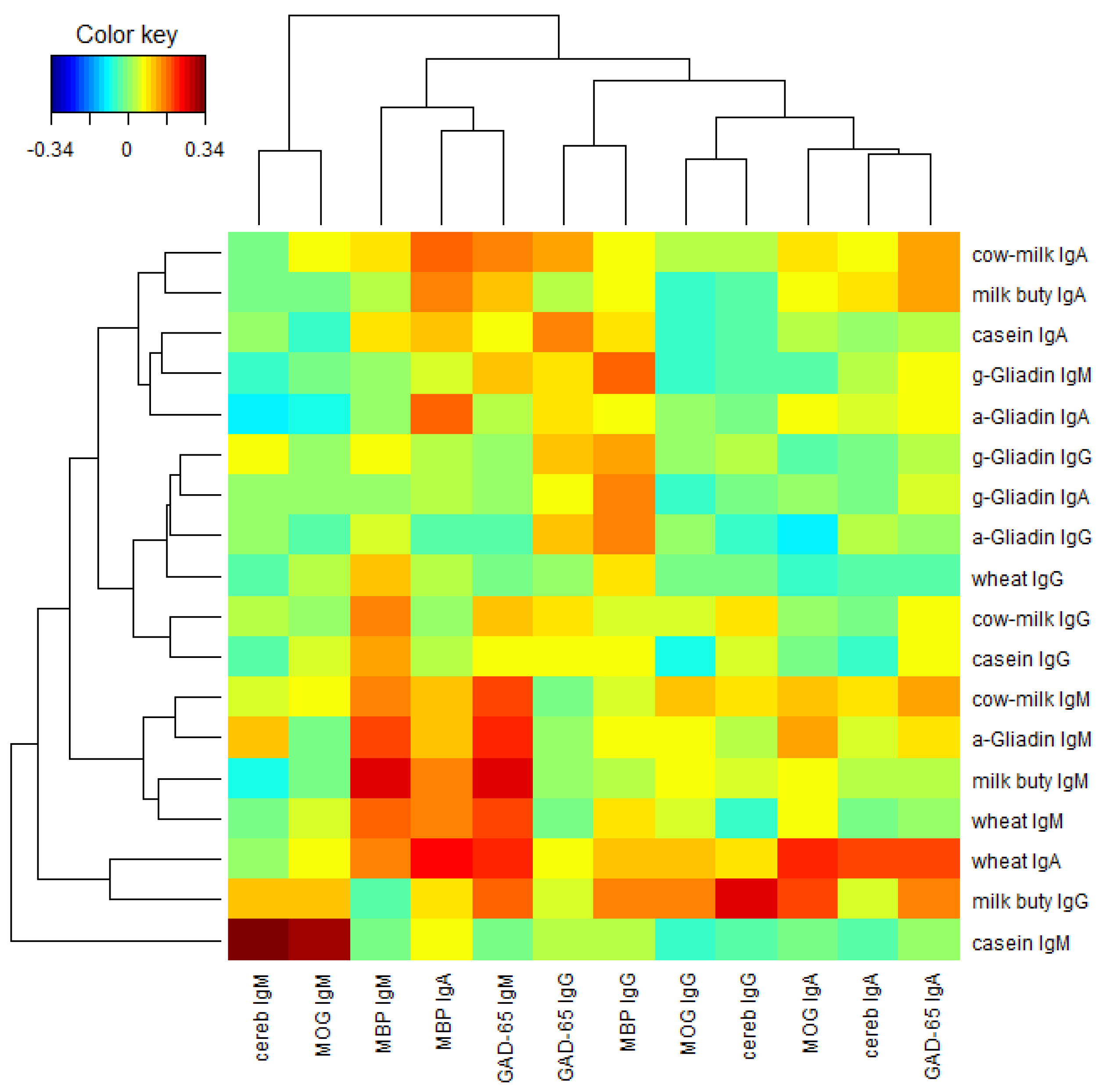
Finally, we performed a two-way cluster analysis of the Pearson’s correlation coefficients between the food proteins and the brain proteins.
Figure 8 presents the result. From
Figure 8 we see that lgG, lgA, and lgM isotypes of the brain proteins (columns) are clustered almost separately with the exception of MBP lgA, lgM, lgG, and lgA isotypes of the brain proteins are clustered near the left, middle and right portions respectively in
Figure 8. Similar phenomena are observed among the food proteins (rows) with a few exceptions. lgA, lgG, and lgM isotypes of the food proteins are clustered near the top, middle, and bottom portions, respectively, in
Figure 8. It shows that the Pearson’s correlation coefficients between the food proteins and the brain proteins are quite different among different isotypes.
Figure 8.
Two-way cluster analysis of the Pearson’s correlation coefficients between the food proteins and the brain proteins.
Figure 8.
Two-way cluster analysis of the Pearson’s correlation coefficients between the food proteins and the brain proteins.
4. Discussion
The goal of this study was to measure elevations in IgG, IgM, and IgA antibodies against wheat and milk and their major allergens such as α-gliadin, γ-gliadin, α + β-casein, and milk butyrophilin. As wheat and milk proteins and peptides share homology with each other and their antibodies cross-react with neural antigens [
13,
16,
18,
20], we wanted to investigate whether or not antibodies detected against gliadin, casein, and butyrophilin contribute to elevation in antibodies against GAD-65, cerebellar, MBP, and MOG.
In our study the serum concentrations and percentage of elevation in antibodies against wheat and cow’s milk in tested populations were very similar. The two-way cluster analysis of the Pearson’s correlation coefficient between the food proteins and brain proteins, as depicted in
Figure 8, shows cow’s milk, milk butyrophilin, α + β-casein, α-gliadin, and wheat clustering significantly when cross-referenced with MBP and GAD-65. This simultaneous elevation in wheat, α-gliadin, milk, and α + β-casein may be related to antigenic similarity or cross-reactivity between wheat and milk proteins. It has been shown that there is a high degree of homology or cross-reactivity between bovine α + β-casein and the α-gliadin 33-mer. This homology between milk proteins is demonstrated not only by IgA anti-gliadin antibody immune reactivity with milk proteins [
26,
31] but also by IgA reactivity to α + β-casein in celiac disease [
32] and the induction of a similar local inflammatory reaction after rectal challenge with wheat and milk proteins [
33]. This cross-reactivity between gliadin and casein was confirmed in our recent study after application of monoclonal and polyclonal antibodies made against α-gliadin 33-mer and exhibiting strong immune reactivity against milk proteins both by ELISA and dot blot assays [
16]. We also demonstrated that mouse or rabbit anti-α-gliadin 33-mer reacted significantly with GAD-65, MBP and cerebellar peptides [
16]. Therefore, it was important to look for co-occurrence of IgG, IgM, and IgA antibodies against wheat, milk protein, and neural antigens in human serum. Analysis of data showed that a significant number of specimens with elevation in antibodies against α-gliadin or γ-gliadin peptides exhibited simultaneous elevation in isotype-specific antibodies against GAD-65 and cerebellar peptides (
Figure 8). This indicates that cross reactivity of gliadin with GAD-65 and cerebellar contributes only partially to elevation in antibodies against GAD 65 and cerebellar.
In addition, earlier studies have shown that injection of bovine milk protein induced MS-like syndrome in animal model and concluded that consumption of milk products may modulate the pathogenic autoimmune response to MOG peptide sequence 76–87 (IGEGKVTLRIRN) [
23]. Furthermore, it was demonstrated that antibody cross-reactivity between the
N-terminal domain of bovine milk protein with MOG was responsible for the simultaneous detection of butyrophilin-specific antibody, as well as MOG peptide antibody in 34% of MS patients [
34]. It was suggested that exposure to common dietary antigens may influence the composition and function of the MOG-specific autoimmune repertoire during the course of MS. Our findings presented in
Table 2 showed that only half of the individuals reacting to milk proteins exhibit simultaneous elevations in milk protein as well as MOG- and MBP-specific antibodies (
Figure 8).
In the earlier study [
34] it was indicated that although the MOG-specific antibody repertoire cross-reacts with multiple butyrophilin peptide epitopes and this pattern of epitope recognition varies from individual to individual, the highest frequency of antibody responses was against peptides spanning amino acid sequences 76–100 of both butyrophilin and MOG, with a match of over 50% (shown below) [
34].
In fact, one of our earlier studies investigated the link between immune response to dietary proteins, gliadin, and cerebellar peptides in children with autism. The study showed epitope similarity between gliadin 8-mer (EQVPLVQQ) and cerebellar 8-mer (EDVPLLED) [
18]. Anti-gliadin epitopes or anti-cerebellar epitopes reacted almost equally against both gliadin and cerebellar peptides, indicating that this epitope may be responsible for cross-reactive antibody production between gliadin and cerebellar peptides [
18]. This difference in epitope recognition was confirmed in the current study by conducting inhibition assays, which demonstrated that while cerebellar peptide was capable of inhibiting cerebellar antibody in all four tested sera by 60%–70%, wheat antigens and α-gliadin inhibited these antibodies by more than 30% in only two out of four of the tested specimens (
Figure 6). Similar to these results all sera with high levels of MOG antibodies were inhibited by absorption with MOG, but in only two out of four sera could MOG-specific antibodies be inhibited by milk, milk butyrophilin, and α + β-casein, but not by HSA (
Figure 7). With these experiments we demonstrated that cerebellar and MOG peptides, which are known to be important autoantigens in gluten ataxia and MS, can cross-react with wheat and milk proteins. The demonstration of molecular mimicry between α-gliadin, cerebellar peptide, milk butyrophilin, and MOG, and the simultaneous detection of antibodies against these proteins in a small percentage of the general population may have broader implications in the induction of neuroimmune disorders. In these individuals, due to a regulatory defect in mucosal immunity, the consumption of wheat and milk products provides a source of α-gliadin, γ-gliadin, and milk butyrophilin-derived peptides that can cross the gut mucosa to stimulate antigen-specific immune responses both locally in the gut as well as in the periphery. In the majority of the population the cerebellar and MOG normally remains sequestered behind the blood brain barrier (BBB). However, CNS inflammation and BBB breakdown can render the neural tissue antigens accessible to the cross-reactive antibodies and auto-reactive lymphocytes, subsequently resulting in neuroimmune disorders [
34,
35,
36].
5. Conclusions
It has been demonstrated that wheat and milk proteins act as molecular mimics of cerebellar peptide and MOG.
This cross-reactivity between α-gliadin and cerebellar peptides, and between milk butyrophilin and MOG peptides, could be responsible for the simultaneous detection of antibodies against these molecules in a small percentage of tested blood samples.
The pathophysiological consequences of molecular mimicry involving wheat and milk with human tissue antigens are difficult to predict, as is the detection of antibodies against them in human sera. This is because they can be influenced by multiple factors, including an individual’s genotype, the timing and level of exposure, and the health of the gut and blood brain barriers, and as such these complex interactions deserve further study.






 and after absorption with HSA
and after absorption with HSA  , wheat
, wheat  , gliadin peptide
, gliadin peptide  , GAD-65
, GAD-65  and cerebellar peptide
and cerebellar peptide  .
.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}