Alternative Sources of Omega-3 Fats: Can We Find a Sustainable Substitute for Fish?



Abstract
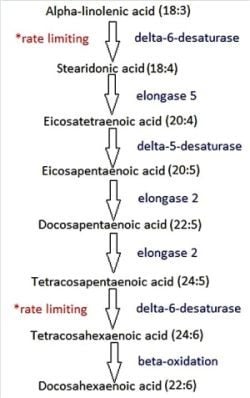
:
1. Introduction
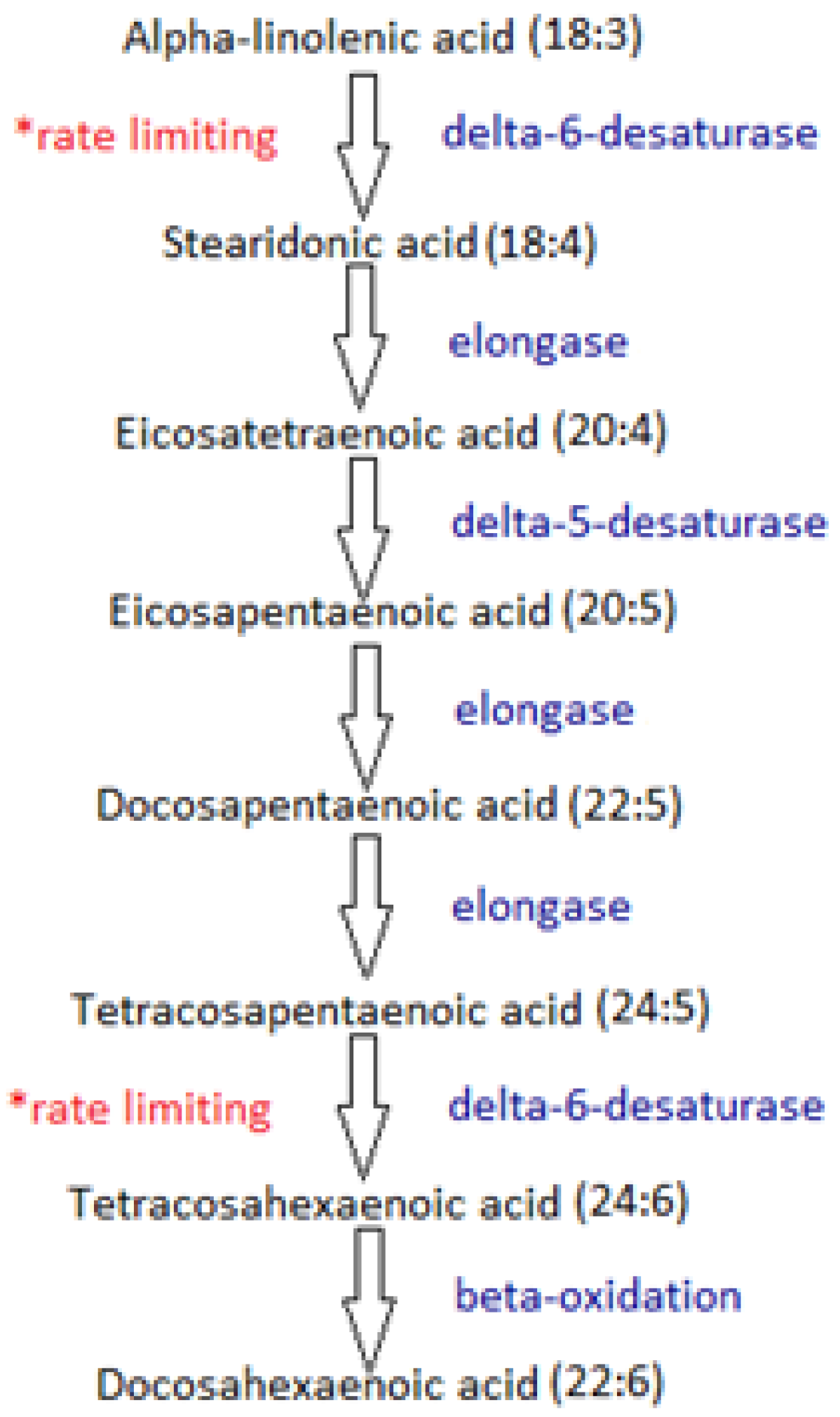
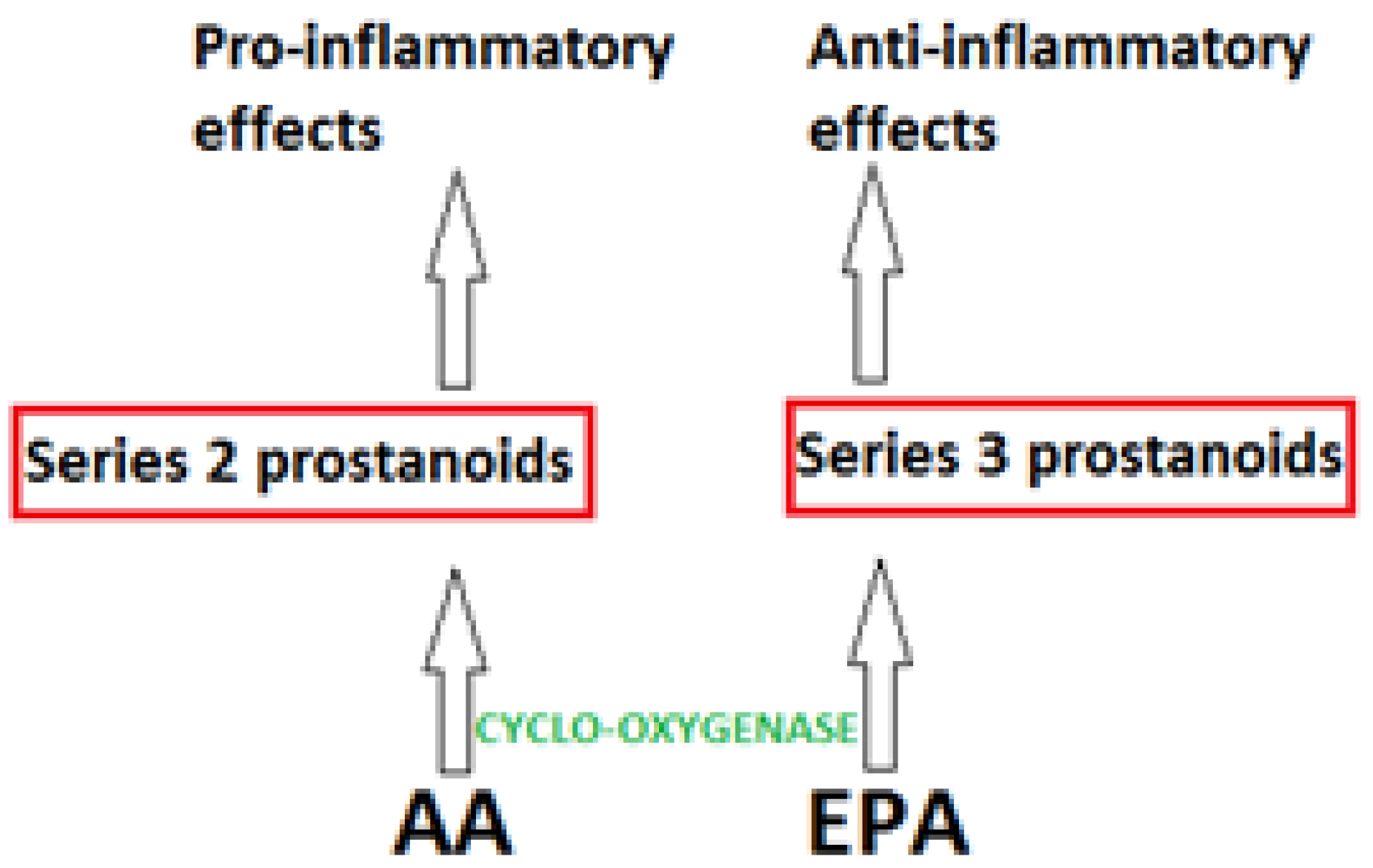
2. Mechanisms of Fatty Acids in Inflammation
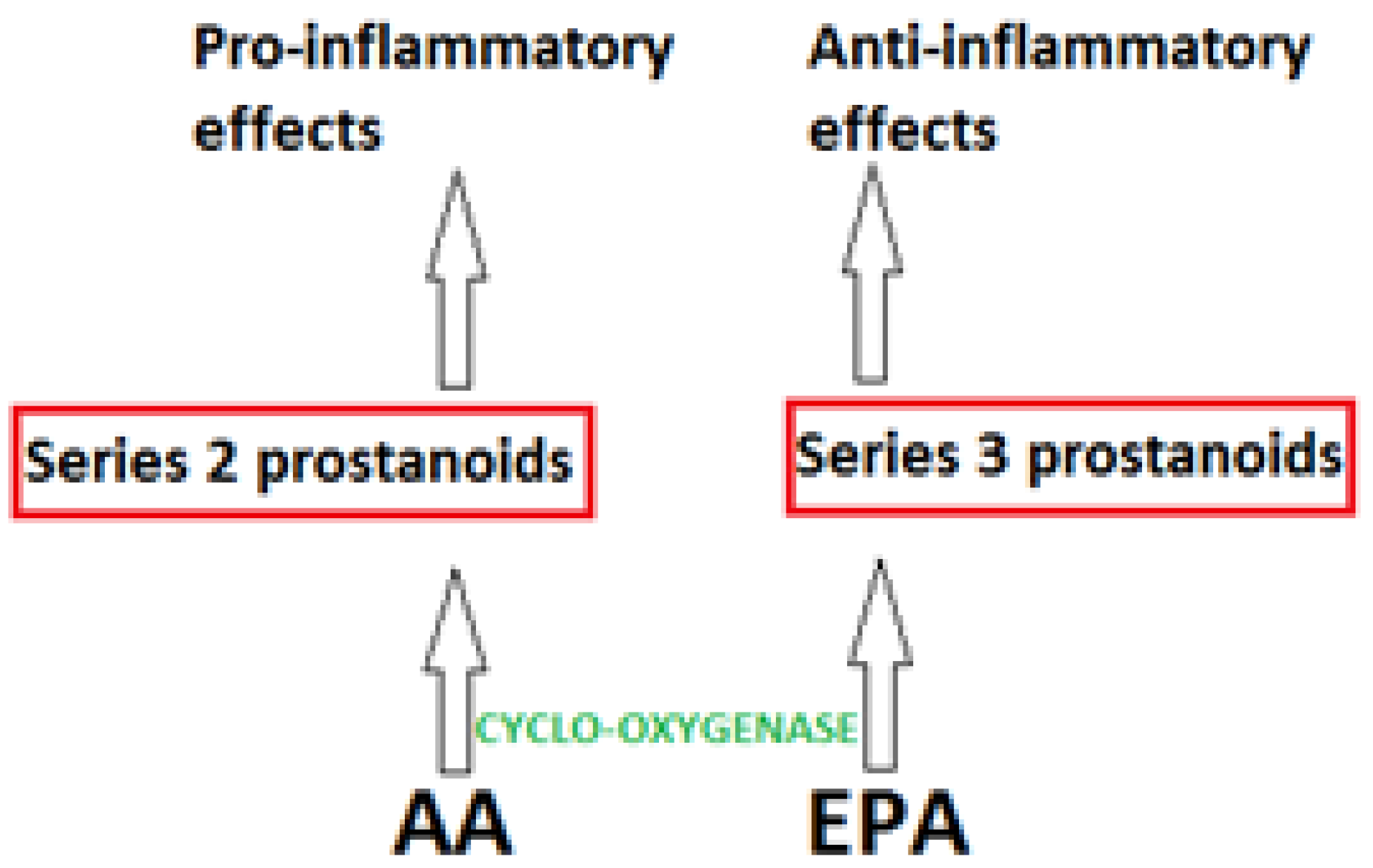
3. Inflammation, Omega-3 Pufa and Health Benefits
4. Do Plant Oils Offer an Alternative to Fish Oils?
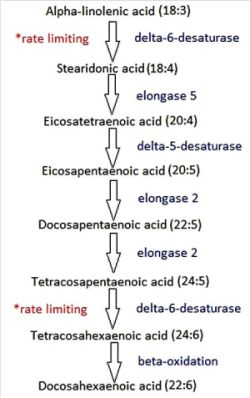
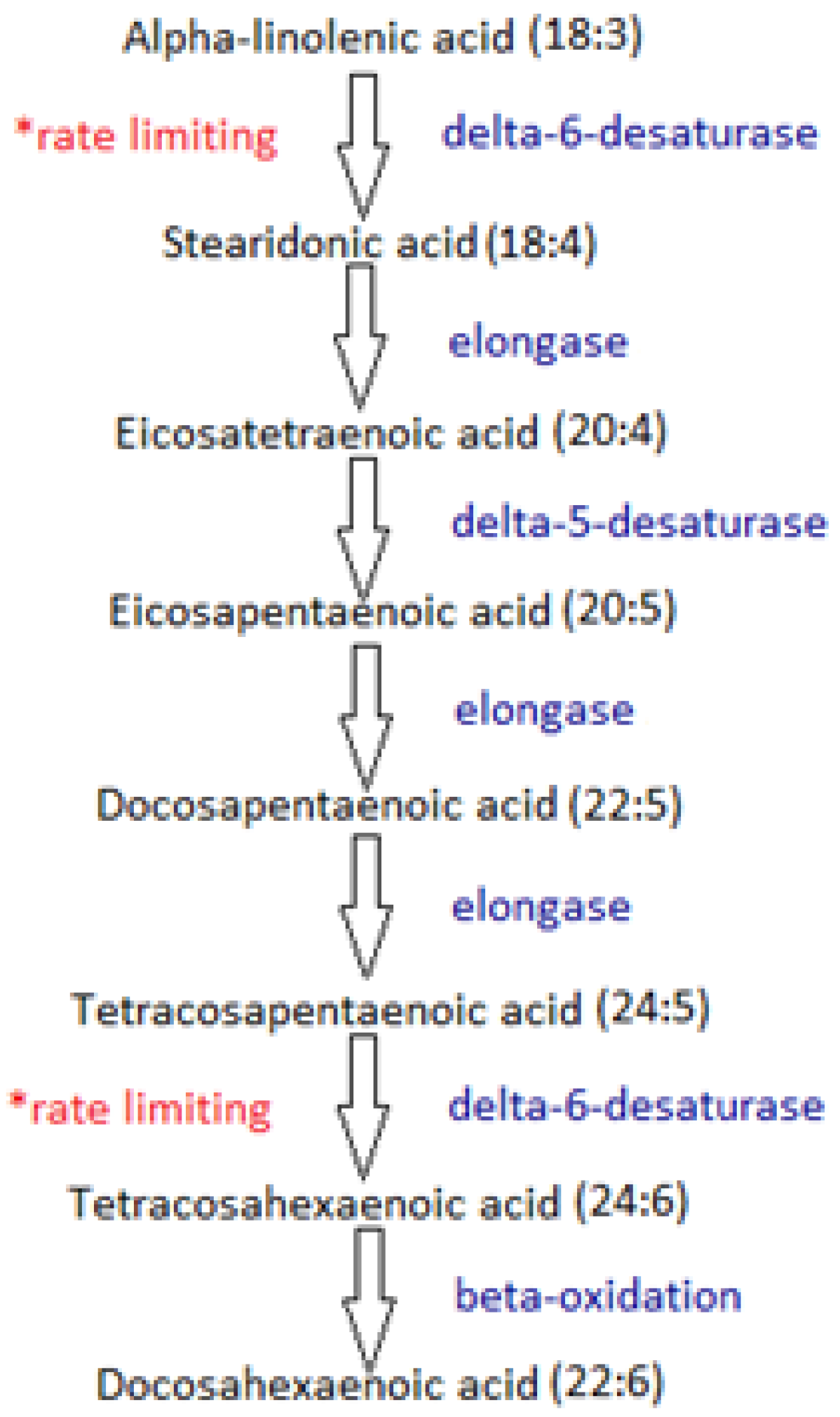
5. Stearidonic Acid: Can We Bypass the Rate-Limiting Step?
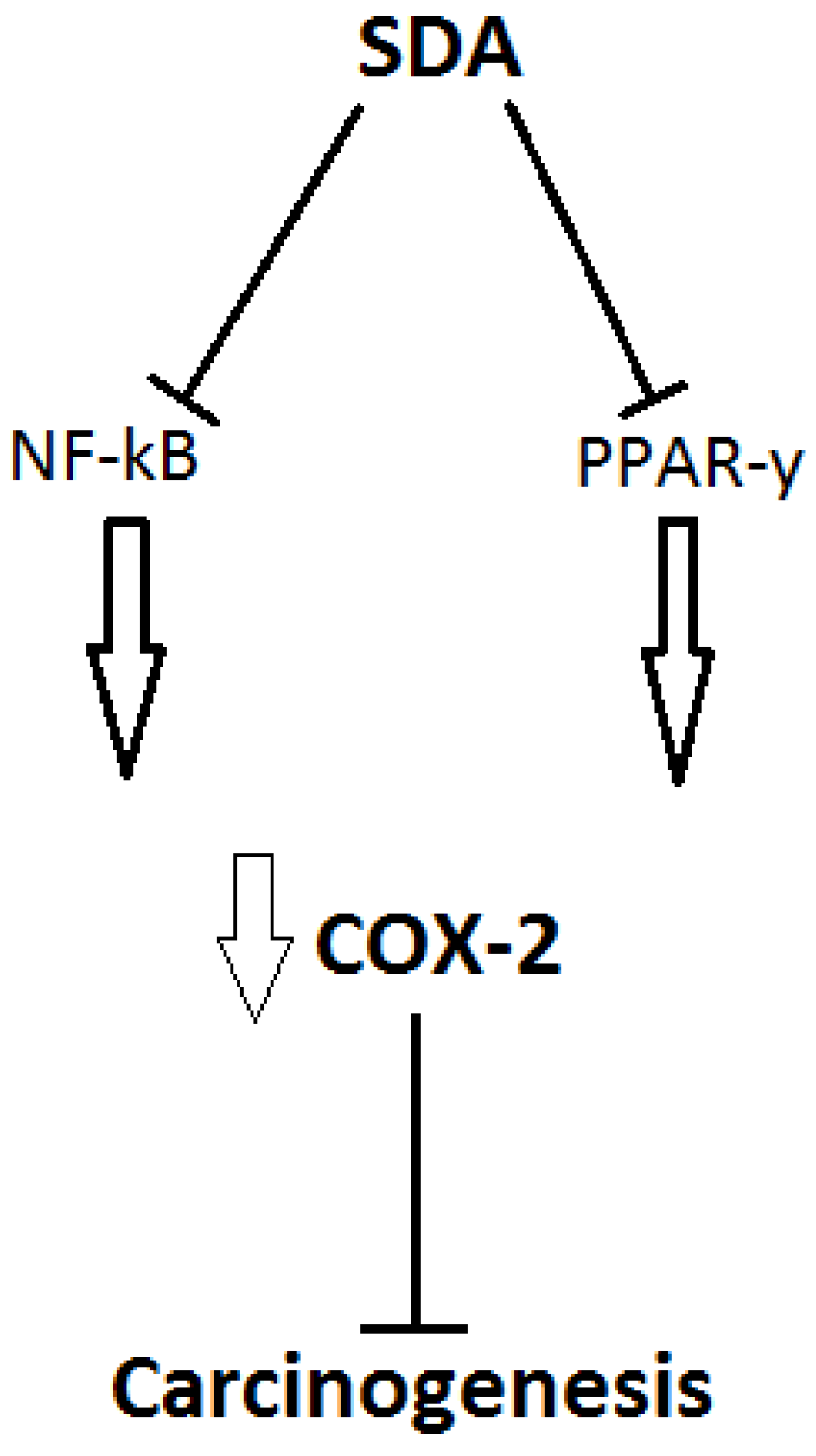
5.1. The Relationship of SDA Supplementation with Health and Disease
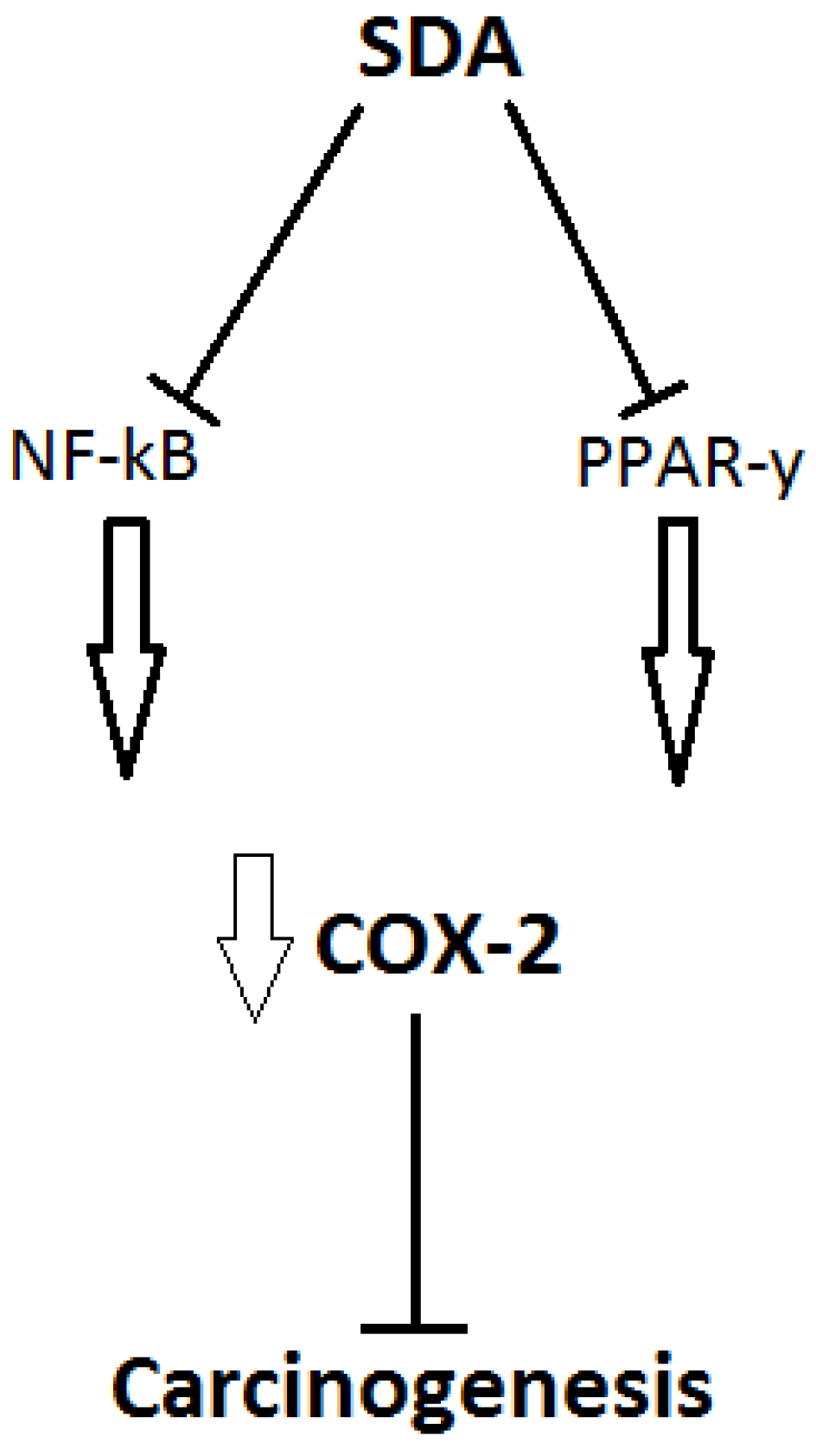
{kind=link}
{kind=link}
{kind=link}
{kind=link}
5.2. Can SDA Substitute as an Animal Feed?
5.3. The Differential Effects of EPA and DHA in Inflammation
6. Algal Oils as a Source of EPA and DHA
7. Conclusions
Acknowledgments
Conflict of Interest
References
- Calder, P. n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83, 1505–1519. [Google Scholar]
- Guil-Guerrero, J.L. Stearidonic acid (18:4n-3): Metabolism, nutritional importance, medical uses and natural sources. Eur. J. Lipid. Sci. Technol. 2007, 109, 1226–1236. [Google Scholar] [CrossRef]
- Simopoulos, A. Omega-3 fatty acids in inflammation and autoimmune diseases. J. Am. Coll. Nutr. 2002, 21, 495–505. [Google Scholar]
- Strobel, C.; Jahreis, G.; Kuhnt, K. Survey of n-3 and n-6 polyunsaturated fatty acids in fish and fish products. Lipids Health Dis. 2012, 11, 144. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Sadovy, Y.; Reynolds, J.D. Extinction vulnerability in marine populations. Fish Fish. 2003, 4, 25–64. [Google Scholar] [CrossRef]
- Calder, P. Polyunsaturated fatty acids, inflammatory processes and inflammatory bowel diseases. Mol. Nutr. Food Res. 2008, 52, 885–897. [Google Scholar] [CrossRef]
- Burdge, G.; Calder, P. Conversion of alpha-linolenic acid to longer-chain polyunsaturated fatty acids in human adults. Reprod. Nutr. Dev. 2005, 45, 581–597. [Google Scholar] [CrossRef]
- Surette, M.; Edens, M.; Chilton, F.; Tramposch, K. Dietary echium oil increases plasma and neutrophil long-chain (n-3) fatty acids and lowers serum triacylglycerols in hypertriglyceridemic humans. J. Nutr. 2004, 134, 1406–1411. [Google Scholar]
- Calder, P. Mechanisms of action of (n-3) fatty acids. J. Nutr. 2012, 142, 592–599. [Google Scholar] [CrossRef]
- Nie, D.; Che, M.; Grignon, D.; Tang, K.; Honn, K. Role of eicosanoids in prostate cancer progression. Cancer Metast. Rev. 2001, 20, 195–206. [Google Scholar] [CrossRef]
- Berridge, M. Inositol trisphosphate and diacylglycerol as second messengers. Biochem. J. 1984, 220, 345–360. [Google Scholar]
- Egert, S. Influence of three rapeseed oil-rich diets, fortified with α-linolenic acid, eicosapentaenoic acid or docosahexaenoic acid on the composition and oxidizability of low-density lipoproteins: Results of a controlled study in healthy volunteers. Eur. J. Clin. Nutr. 2007, 61, 314–325. [Google Scholar] [CrossRef]
- Miles, E.A.; Banerjee, T.; Dooper, M.M.B.W.; M’Rabet, L.; Graus, Y.M.F.; Calder, P.C. The influence of different combinations of γ-linolenic acid, stearidonic acid and epa on immune function in healthy young male subjects. Br. J. Nutr. 2004, 91, 893. [Google Scholar] [CrossRef]
- Vendramini-Costa, D.; Carvalho, J. Molecular link mechanisms between inflammation and cancer. Curr. Pharm. Des. 2012, 18, 3831–3852. [Google Scholar] [CrossRef]
- Calder, P. Polyunsaturated fatty acids, inflammation, and immunity. Lipids 2001, 36, 1007–1024. [Google Scholar] [CrossRef]
- Sharma, R.; Zucknick, M.; London, R.; Kacevska, M.; Liddle, C.; Clarke, S. Systemic inflammatory response predicts prognosis in patients with advanced-stage colorectal cancer. Clin. Colorectal Cancer 2008, 7, 331–337. [Google Scholar] [CrossRef]
- Robert, G.; Descazeaud, A.; Nicolaïew, N.; Terry, S.; Sirab, N.; Vacherot, F.; Maillé, P.; Allory, Y.; de la Taille, A. Inflammation in benign prostatic hyperplasia: A 282 patients’ immunohistochemical analysis. Prostate 2009, 69, 1774–1780. [Google Scholar] [CrossRef]
- Yen, D.; Cheung, J.; Scheerens, H.; Poulet, F.; McClanahan, T.; McKenzie, B.; Kleinschek, M.; Owyang, A.; Mattson, J.; Blumenschein, W.; et al. IL-23 is essential for T cell-mediated colitis and promotes inflammation via IL-17 and IL-6. J. Clin. Invest. 2006, 116, 1310–1316. [Google Scholar] [CrossRef]
- Elson, C.; Cong, Y.; Weaver, C.; Schoeb, T.; McClanahan, T.; Fick, R.; Kastelein, R. Monoclonal anti-interleukin 23 reverses active colitis in a T cell-mediated model in mice. Gastroenterology 2007, 132, 2359–2370. [Google Scholar] [CrossRef]
- Panayi, G.; Lanchbury, J.; Kingsley, G. The importance of the T cell in initiating and maintaining the chronic synovitis of rheumatoid arthritis. Arthritis Rheum. 1992, 35, 729–735. [Google Scholar] [CrossRef]
- Krutmann, J.; Grewe, M. Sequential activation of Th1 and Th2 cells in the immunopathogenesis of atopic eczema—The 2-phase model. Allergologie 1996, 19, 449–451. [Google Scholar]
- Hansson, G. Inflammation, atherosclerosis, and coronary artery disease. N. Engl. J. Med. 2005, 352, 1685–1695. [Google Scholar] [CrossRef]
- Ishihara, K.; Komatsu, W.; Saito, H.; Shinohara, K. Comparison of the effects of dietary alpha-linolenic, stearidonic, and eicosapentaenoic acids on production of inflammatory mediators in mice. Lipids 2002, 37, 481–486. [Google Scholar] [CrossRef]
- Van Horssen, R.; Ten Hagen, T.; Eggermont, A. TNF-alpha in cancer treatment: Molecular insights, antitumor effects, and clinical utility. Oncologist 2006, 11, 397–408. [Google Scholar] [CrossRef]
- Mohammed, A.; Janakiram, N.; Brewer, M.; Duff, A.; Lightfoot, S.; Brush, R.; Anderson, R.; Rao, C. Endogenous n-3 polyunsaturated fatty acids delay progression of pancreatic ductal adenocarcinoma in Fat-1-p48Cre/+-LSL-KrasG12D/+ mice. Neoplasia 2012, 14, 1249–1259. [Google Scholar]
- Chua, M.; Sio, M.; Sorongon, M.; Dy, J. Relationship of dietary intake of omega-3 and omega-6 fatty acids with risk of prostate cancer development: A meta-analysis of prospective studies and review of literature. Prostate Cancer 2012, 2012, 826254. [Google Scholar]
- Williams, C.D.; Whitley, B.; Hoyo, C.; Grant, D.; Iraggi, J.; Newman, K.; Gerber, L.; Taylor, L.; McKeever, M.; Freedland, S. A High ratio of dietary n-6/n-3 polyunsaturated fatty acids is associated with increased risk of prostate cancer. Nutr. Res. 2011, 31, 1–8. [Google Scholar]
- Oh, K.; Willett, W.; Fuchs, C.; Giovannucci, E. Dietary marine n-3 fatty acids in relation to risk of distal colorectal adenoma in women. Cancer Epidemiol. Biomarkers Prev. 2005, 14, 835–841. [Google Scholar] [CrossRef]
- Daniel, C.; McCullough, M.; Patel, R.; Jacobs, E.; Flanders, W.; Thun, M.; Calle, E. Dietary intake of omega-6 and omega-3 fatty acids and risk of colorectal cancer in a prospective cohort of U.S. men and women. Cancer Epidemiol. Biomarkers Prev. 2009, 18, 516–525. [Google Scholar] [CrossRef]
- Kobayashi, N.; Barnard, J.; Henning, S.; Elashoff, D.; Reddy, S.; Cohen, P.; Leung, P.; Hong-Gonzalez, J.; Freedland, S.; Said, J.; et al. Effect of altering dietary w-6/w-3 fatty acid ratios on prostate cancer membrane composition, cyclooxygenase-2, and prostaglandin E2. Clin. Cancer Res. 2006, 12, 4670. [Google Scholar]
- Murff, H.; Shu, X.; Li, H.; Yang, G.; Wu, X.; Cai, H.; Wen, W.; Gao, Y.; Zheng, W. Dietary polyunsaturated fatty acids and breast cancer risk in chinese women, a prospective cohort study. Int. J. Cancer Suppl. 2011, 128, 1434–1441. [Google Scholar] [CrossRef]
- Musa-Veloso, K.; Binns, M.; Kocenas, A.; Chung, C.; Rice, H.; Oppedal-Olsen, H.; Lloyd, H.; Lemke, S. Impact of low v. moderate intakes of long-chain n-3 fatty acids on risk of coronary heart disease. Br. J. Nutr. 2011, 106, 1129–1141. [Google Scholar] [CrossRef]
- Jiang, W.; Oken, H.; Fiuzat, M.; Shaw, L.; Martsberger, C.; Kuchibhatla, M.; Kaddurah-Daouk, R.; Steffens, D.; Baillie, R.; Cuffe, M.; et al. Plasma omega-3 polyunsaturated fatty acids and survival in patients with chronic heart failure and major depressive disorder. J. Cardiovasc. Trans. Res. 2012, 5, 92–99. [Google Scholar] [CrossRef]
- Ibrahim, A.; Mbodji, K.; Hassan, A.; Aziz, M.; Boukhettala, N.; Coeffier, M.; Savoye, G.; Dechelotte, P.; Marion-Letellier, R. Anti-inflammatory and anti-angiogenic effect of long chain n-3 polyunsaturated fatty acids in intestinal microvascular endothelium. Clin. Nutr. 2011, 30, 678–687. [Google Scholar] [CrossRef]
- Uchiyama, K.; Nakamura, M.; Odahara, S.; Koido, S.; Katahira, K.; Shiraishi, H.; Ohkusa, T.; Fujise, K.; Tajiri, H. n-3 polyunsaturated fatty acid diet therapy for patients with inflammatory bowel disease. Inflamm. Bowel. Dis. 2010, 16, 1696–1707. [Google Scholar] [CrossRef]
- Miller, M.; Nichols, P.; Carter, C. n-3 oil sources for use in aquaculture—Alternatives to the unsustainable harvest of wild fish. Nutr. Res. Rev. 2008, 21, 85–96. [Google Scholar] [CrossRef]
- Kuhnt, K.; Degen, C.; Jaudszus, A.; Jahreis, G. Searching for health beneficial n-3 and n-6 fatty acids in plant seeds. Eur. J. Lipid. Sci. Technol. 2012, 114, 153–160. [Google Scholar] [CrossRef]
- Bell, J.; Tocher, D.; Henderson, R.; Dick, J.; Crampton, V. Altered fatty acid compositions in atlantic salmon (Salmo Salar L.) fed diets containing linseed and rapeseed oils can be partially restored by a subsequent fish oil finishing diet. J. Nutr. 2003, 133, 2793–2801. [Google Scholar]
- Torstensen, B.E.; Lie, Ø.; Frøyland, L. Lipid metabolism and tissue composition in atlantic salmon (Salmo Salar L.)—Effects of capelin oil, palm oil, and oleic acid-enriched sunflower oil as dietary lipid sources. Lipids 2000, 35, 653–664. [Google Scholar] [CrossRef]
- Seierstad, S.; Seljeflot, I.; Johansen, O.; Hansen, R.; Haugen, M.; Rosenlund, G.; Froyland, L.; Arnesen, H. Dietary intake of differently fed salmon; the influence on markers of human atherosclerosis. Eur. J. Clin. Nutr. 2005, 35, 52–59. [Google Scholar]
- Stulc, T.; Ceska, R. Cholesterol lowering and the vessel wall: New insights and future perspectives. Physiol. Res. 2001, 50, 461–471. [Google Scholar]
- Bell, J.G.; Henderson, R.J.; Tocher, D.R.; Sargent, J.R. Replacement of dietary fish oil with increasing levels of linseed oil: Modification of flesh fatty acid compositions in Atlantic salmon (Salmo Salar L.) using a fish oil finishing diet. Lipids 2004, 39, 223–232. [Google Scholar] [CrossRef]
- Codabaccus, M.; Bridle, A.; Nichols, P.; Carter, C. Restoration of fillet n-3 long-chain polyunsaturated fatty acid is improved by a modified fish oil finishing diet strategy for Atlantic salmon (Salmo Salar L.) smolts fed palm fatty acid distillate. J. Agric. Food Chem. 2012, 60, 458–466. [Google Scholar] [CrossRef]
- James, M.; Ursin, V.; Cleland, L. Metabolism of stearidonic acid in human subjects: Comparison with the metabolism of other n-3 fatty acids. Am. J. Clin. Nutr. 2003, 77, 1140–1145. [Google Scholar]
- Surette, M. Dietary omega-3 PUFA and health: Stearidonic acid-containing seed oils as effective and sustainable alternatives to traditional marine oils. Mol. Nutr. Food Res. 2013, 57, 748–759. [Google Scholar]
- Lemke, S.; Vicini, J.; Su, H.; Goldstein, D.; Nemeth, M.; Krul, E.; Harris, W. Dietary intake of stearidonic acid-enriched soybean oil increases the omega-3 index: Randomized, double-blind clinical study of efficacy and safety. Am. J. Clin. Nutr. 2010, 92, 766–775. [Google Scholar] [CrossRef]
- Harris, W.; DiRienzo, M.; Sands, S.; George, C.; Jones, P.; Eapen, A. Stearidonic acid increases the red blood cell and heart eicosapentaenoic acid content in dogs. Lipids 2007, 42, 325–333. [Google Scholar] [CrossRef]
- Hammond, B.; Lemen, J.; Ahmed, G.; Miller, K.; Kirkpatrick, J.; Fleeman, T. Safety assessment of SDA soybean oil: Results of a 28-day gavage study and a 90-day/one generation reproduction feeding study in rats. Regul. Toxicol. Pharmacol. 2008, 52, 311–323. [Google Scholar] [CrossRef]
- Krul, E.; Lemke, S.; Mukherjea, R.; Taylor, M.; Goldstein, D.; Su, H.; Liu, P.; Lawless, A.; Harris, W.; Maki, K. Effects of duration of treatment and dosage of eicosapentaenoic acid and stearidonic acid on red blood cell eicosapentaenoic acid content. Prostaglandins Leukot. Essent. Fat. Acids 2012, 86, 51–59. [Google Scholar] [CrossRef]
- Von Schacky, C. A review of omega-3 ethyl esters for cardiovascular prevention and treatment of increased blood triacylglyceride levels. Vasc. Health Risk Manag. 2006, 2, 251–262. [Google Scholar] [CrossRef]
- Walker, C.; Jebb, S.; Calder, P. Stearidonic acid as a supplemental source of w-3 polyunsaturated fatty acids to enhance status for improved human health. Nutrition 2013, 29, 363–369. [Google Scholar] [CrossRef]
- Forrest, L.; Boudyguina, E.; Wilson, M.; Parks, J. Echium oil reduces atherosclerosis in apoB100-only LDLrKO mice. Atherosclerosis 2012, 220, 118–121. [Google Scholar] [CrossRef]
- Howard, B.; Robbins, D.; Sievers, M.; Lee, E.; Rhoades, D.; Devereux, R.; Cowan, L.; Gray, R.; Welty, T.; Go, O.; et al. LDL cholesterol as a strong predictor of coronary heart disease in diabetic individuals with insulin resistance and low LDL: The strong heart study. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 830–835. [Google Scholar] [CrossRef]
- Banz, W.; Davis, J.; Clough, R.; Cheatwood, J. Stearidonic acid: Is there a role in the prevention and management of type 2 diabetes mellitus? J. Nutr. 2012, 142, 635S–640S. [Google Scholar] [CrossRef]
- Wu, D.; Meydani, M.; Leka, L.; Nightinggale, Z.; Handelman, G.; Blumberg, J.; Meydani, S. Effect of dietary supplementation with black currant seed oil on the immune response of healthy elderly subjects. Am. J. Clin. Nutr. 1999, 70, 536–543. [Google Scholar]
- Whelan, J.; Gouffon, J.; Zhao, Y. Effects of dietary stearidonic acid on biomarkers of lipid metabolism. J. Nutr. 2012, 142, 630S–634S. [Google Scholar] [CrossRef]
- Horia, E.; Watkins, B. Comparison of stearidonic acid and α-linoleic acid on PGE2 production and COX-2 protein levels in MDA-MB-231 breast cancer cell cultures. J. Nutr. Biochem. 2005, 16, 184–192. [Google Scholar] [CrossRef]
- Ferrandina, G.; Legge, F.; Ranelletti, F.; Zannoni, G.; Maggiano, N.; Evangelisti, A.; Mancuso, S.; Scambia, G.; Lauriola, L. Cyclooxygenase-2 expression in endometrial carcinoma: Correlation with clinicopathologic parameters and clinical outcome. Cancer 2002, 95, 801–807. [Google Scholar] [CrossRef]
- Kelavkar, U.; Hutzley, J.; Dhir, R.; Kim, P.; Allen, K.; McHugh, K. Prostate tumour growth and recurrence can be modulated by the ω-6:ω-3 ratio in diet: Athymic mouse xenograft model simulating radical prostatectomy. Neoplasia 2006, 8, 112–124. [Google Scholar] [CrossRef]
- Kitessa, S.M.; Young, P. Echium oil is better than rapeseed oil in enriching poultry meat with n-3 polyunsaturated fatty acids, including eicosapentaenoic acid and docosapentaenoic acid. Br. J. Nutr. 2009, 101, 709. [Google Scholar] [CrossRef]
- Cleveland, B.; Francis, D.; Turchini, G. Echium oil provides no benefit over linseed oil for (n-3) long-chain PUFA biosynthesis in rainbow trout. J. Nutr. 2012, 142, 1449–1455. [Google Scholar] [CrossRef]
- Miller, M.; Nichols, P.; Carter, C. Replacement of dietary fish oil for Atlantic salmon parr (Salmo Salar L.) with a stearidonic acid containing oil has no effect on omega-3 long-chain polyunsaturated fatty acid concentrations. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 146, 197–206. [Google Scholar] [CrossRef]
- Kitessa, S.M.; Young, P.; Nattrass, G.; Gardner, G.; Pearce, K.; Pethick, D.W. When balanced for precursor fatty acid supply echium oil is not superior to linseed oil in enriching lamb tissues with long-chain n-3 PUFA. Br. J. Nutr. 2012, 108, 71. [Google Scholar] [CrossRef]
- Verlengia, R.; Gorjao, R.; Kanunfre, C.; Bordin, S.; de Lima, T.; Martins, E.; Curi, R. Comparative effects of eicosapentaenoic acid and docosahexaenoic acid on proliferation, cytokine production and pleiotropic gene expression in Jurkat cells. J. Nutr. Biochem. 2004, 15, 657–665. [Google Scholar] [CrossRef]
- Moore, K.; de Waal Malefyt, R.; Coffman, R.; O-Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef]
- Weldon, S.; Mullen, A.; Loscher, C.; Hurley, L.; Roche, H. Docosahexaenoic acid induces an anti-inflammatory profile in lipopolysaccharide-stimulated human THP-1 macrophages more effectively than eicosapentaenoic acid. J. Nutr. Biochem. 2007, 18, 250–258. [Google Scholar] [CrossRef]
- Lewis, A.; Varghese, S.; Xu, H.; Alexander, H. Interleukin-1 and cancer progression: The emerging role of interleukin-1 receptor antagonist as a novel therapeutic agent in cancer treatment. J. Transl. Med. 2006, 4, 48–60. [Google Scholar] [CrossRef]
- Naugler, W.; Karin, M. The wolf in sheep’s clothing: The role of interleukin-6 in immunity, inflammation and cancer. Trends Mol. Med. 2008, 14, 109–119. [Google Scholar] [CrossRef]
- Adarme-Vega, T.; Lim, D.; Timmins, M.; Vernen, F.; Li, Y.; Schenk, P. Microalgal biofactories: A promising approach towards sustainable omega-3 fatty acid production. Microb. Cell Fact. 2012, 11. [Google Scholar] [CrossRef]
- Subhadra, B.; Grinson-George. Algal Biorefinery-Based Industry: An Approach to Address Fuel and Food Insecurity for a Carbon-Smart World. J. Sci. Food. Agric. 2011, 91, 2–13. [Google Scholar] [CrossRef]
- Doughman, D.; Krupanidhi, S.; Sanjeeve, C. Omega-3 fatty acids for nutrition and medicine considering microalgae oil as a vegetarian source of EPA and DHA. Curr. Diabetes Rev. 2007, 3, 198–203. [Google Scholar] [CrossRef]
- Kyle, D. The large-scale production and use of a single-cell oil highly enriched in docosahexaenoic acid. ACS Symp. Ser. 2001, 788, 92–107. [Google Scholar]
- Sijtsma, L.; de Swaaf, M. Biotechnological production and applications of the ω-3 polyunsaturated fatty acid docosahexaenoic acid. Appl. Microbiol. Biotechnol. 2004, 64, 146–153. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lenihan-Geels, G.; Bishop, K.S.; Ferguson, L.R. Alternative Sources of Omega-3 Fats: Can We Find a Sustainable Substitute for Fish? Nutrients 2013, 5, 1301-1315. https://doi.org/10.3390/nu5041301
Lenihan-Geels G, Bishop KS, Ferguson LR. Alternative Sources of Omega-3 Fats: Can We Find a Sustainable Substitute for Fish? Nutrients. 2013; 5(4):1301-1315. https://doi.org/10.3390/nu5041301
Chicago/Turabian StyleLenihan-Geels, Georgia, Karen S. Bishop, and Lynnette R. Ferguson. 2013. "Alternative Sources of Omega-3 Fats: Can We Find a Sustainable Substitute for Fish?" Nutrients 5, no. 4: 1301-1315. https://doi.org/10.3390/nu5041301
APA StyleLenihan-Geels, G., Bishop, K. S., & Ferguson, L. R. (2013). Alternative Sources of Omega-3 Fats: Can We Find a Sustainable Substitute for Fish? Nutrients, 5(4), 1301-1315. https://doi.org/10.3390/nu5041301