The Dietary Intake of Wheat and other Cereal Grains and Their Role in Inflammation

Abstract
:1. Introduction
2. Wheat Grain, Gluten and Disease
2.1. Wheat Allergy and Intolerance
2.2. Wheat Grain and Gluten
2.3. Gluten, Gliadin and CD
2.4. Gliadin and Immunity
2.5. Gliadin and Intestinal Permeability
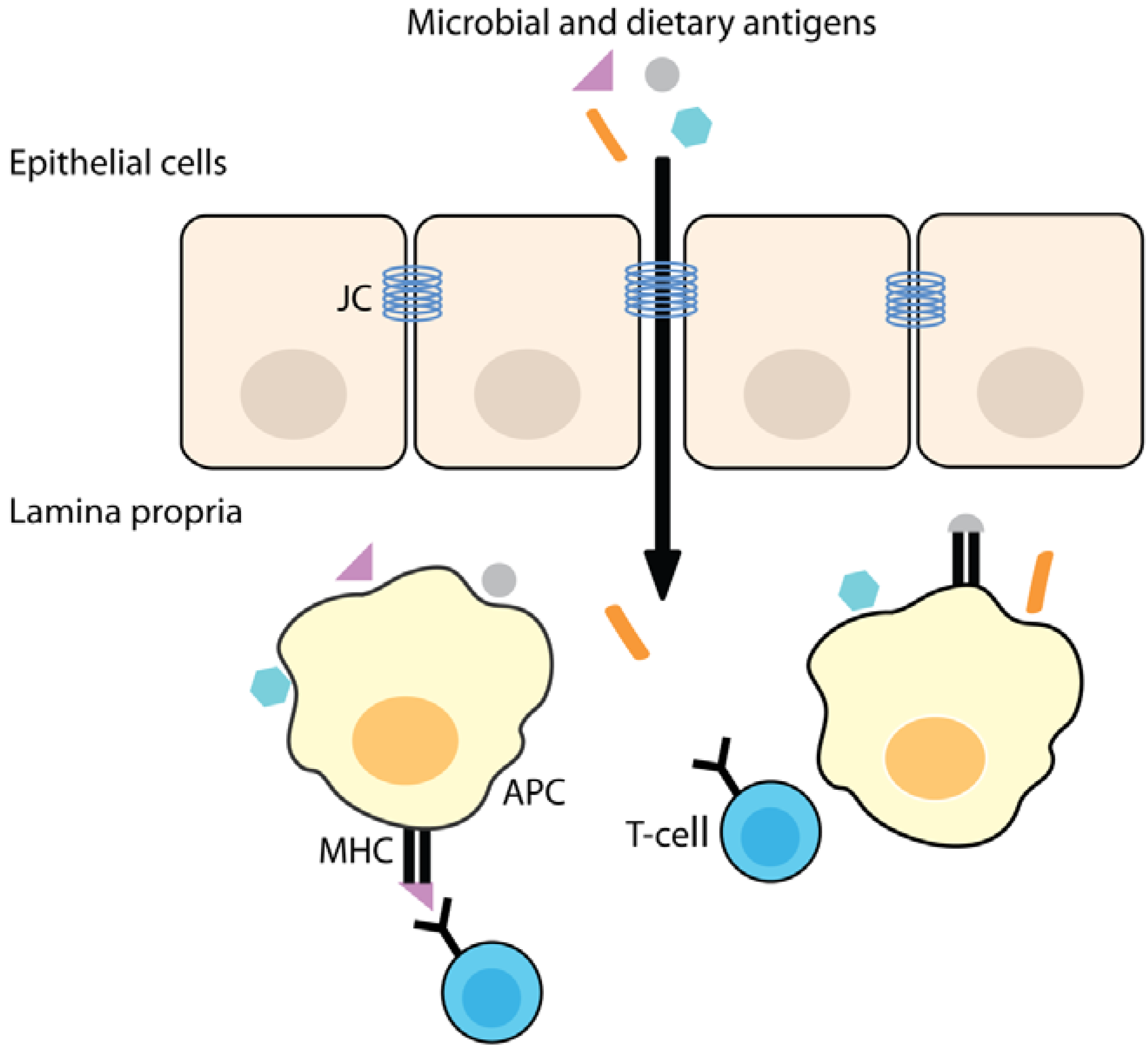
3. Increased Intestinal Permeability
3.1. Increased Intestinal Permeability is Associated with Disease
3.2. Intestinal Barrier Function and Inflammation
3.3. The Role of Zonulin Signaling on Intestinal Permeability
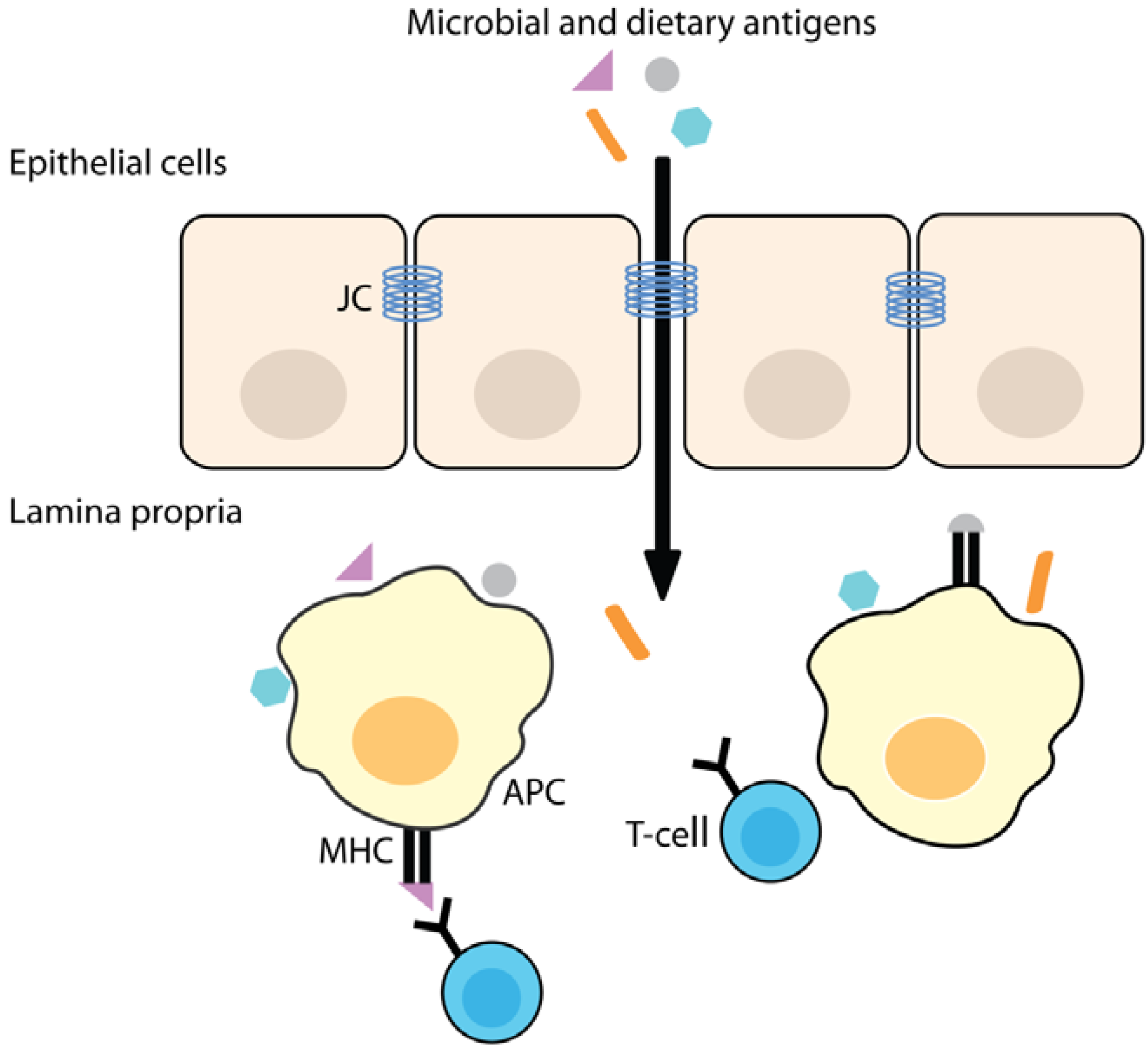
3.4. High Zonulin Levels are Observed in Auto-Immune and Inflammatory Diseases
4. Wheat Germ Agglutinin (WGA)
4.1. Dietary WGA

{kind=link}
| Wheat Derived Products | WGA μg/g (±SD) | Reference Source |
|---|---|---|
| Wheat germ | 300 (±35) | Vincenzi et al., 2000 [42] |
| Wheat germ | 100–500 | Peumans and Van Damme, 1996 [39] |
| Semolina a | 4.0 (±1.0)–10.7 (±1.5) | Matucci et al., 2004 [43] |
| Flour a | 4.3 (±0.7)–4.4 (±1.0) | |
| Wholemeal flour a | 29.5 (±2.5)–50 (±5.5) | |
| Pasta a | ≤0.4 (±0.2)–3.2 (±0.2) | |
| Pasta cooked a | ≤0.3 (±0.2) | |
| Wholemeal pasta (enriched with wheat germ) | 40 (±2.7) | |
| Wholemeal pasta (enriched with wheat germ) cooked | Not detectable | |
| Wholemeal pasta a | 0–5.7 (±0.2) | |
| Wholemeal pasta cooked a | Not detectable | |
| Breakfast cereals a | 13–53 | Ortega-Barria et al., 1994 [41] |
4.2. WGA Binds to Cell Surface Glycoconjugates
4.3. WGA and Immunity
4.4. WGA and Intestinal Permeability
5. Animal Data on Cereal Grain Intake
6. Human Studies on Cereal Grain Intake and Inflammation
6.1. Human Epidemiological Data on Cereal Grain Intake and Inflammation
6.2. Human Intervention Trials on Cereal Grain Intake and Inflammation
6.3. Health Effects of the Paleolithic Diet
6.4. Rechallenge Trial of Effects of Dietary Gluten
7. Conclusion
Acknowledgments
Conflict of Interest
References
- Barnes, P.J.; Adcock, I.M. Glucocorticoid resistance in inflammatory diseases. Lancet 2009, 373, 1905–1917. [Google Scholar] [CrossRef]
- Libby, P. Role of inflammation in atherosclerosis associated with rheumatoid arthritis. Am. J. Med. 2008, 121, S21–S31. [Google Scholar] [CrossRef]
- Raison, C.L.; Capuron, L.; Miller, A.H. Cytokines sing the blues: inflammation and the pathogenesis of depression. Trends Immunol. 2006, 27, 24–31. [Google Scholar] [CrossRef]
- Bosma-den Boer, M.M.; van Wetten, M.L.; Pruimboom, L. Chronic inflammatory diseases are stimulated by current lifestyle: How diet, stress levels and medication prevent our body from recovering. Nutr. Metab. (Lond.) 2012, 9. [Google Scholar] [CrossRef]
- Shelton, R.C.; Miller, A.H. Eating ourselves to death (and despair): The contribution of adiposity and inflammation to depression. Prog. Neurobiol. 2010, 91, 275–299. [Google Scholar] [CrossRef]
- Brown, C.M.; Dulloo, A.G.; Montani, J.P. Sugary drinks in the pathogenesis of obesity and cardiovascular diseases. Int. J. Obes. (Lond.) 2008, 32, S28–S34. [Google Scholar]
- Tatham, A.S.; Shewry, P.R. Allergens to wheat and related cereals. Clin. Exp. Allergy 2008, 38, 1712–1726. [Google Scholar]
- Sapone, A.; Bai, J.C.; Ciacci, C.; Dolinsek, J.; Green, P.H.; Hadjivassiliou, M.; Kaukinen, K.; Rostami, K.; Sanders, D.S.; Schumann, M.; et al. Spectrum of gluten-related disorders: Consensus on new nomenclature and classification. BMC Med. 2012, 10. [Google Scholar] [CrossRef]
- Shewry, P.R. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef]
- Troncone, R.; Jabri, B. Coeliac disease and gluten sensitivity. J. Intern. Med. 2011, 269, 582–590. [Google Scholar] [CrossRef]
- Neuhausen, S.L.; Steele, L.; Ryan, S.; Mousavi, M.; Pinto, M.; Osann, K.E.; Flodman, P.; Zone, J.J. Co-occurrence of celiac disease and other autoimmune diseases in celiacs and their first-degree relatives. J. Autoimmun. 2008, 31, 160–165. [Google Scholar] [CrossRef]
- Biesiekierski, J.R.; Newnham, E.D.; Irving, P.M.; Barrett, J.S.; Haines, M.; Doecke, J.D.; Shepherd, S.J.; Muir, J.G.; Gibson, P.R. Gluten causes gastrointestinal symptoms in subjects without celiac disease: A double-blind randomized placebo-controlled trial. Am. J. Gastroenterol. 2011, 106, 508–514. [Google Scholar] [CrossRef]
- Sapone, A.; Lammers, K.M.; Casolaro, V.; Cammarota, M.; Giuliano, M.T.; de Rosa, M.; Stefanile, R.; Mazzarella, G.; Tolone, C.; Russo, M.I.; et al. Divergence of gut permeability and mucosal immune gene expression in two gluten-associated conditions: Celiac disease and gluten sensitivity. BMC Med. 2011, 9. [Google Scholar] [CrossRef]
- Harris, K.M.; Fasano, A.; Mann, D.L. Cutting edge: IL-1 controls the IL-23 response induced by gliadin, the etiologic agent in celiac disease. J. Immunol. 2008, 181, 4457–4460. [Google Scholar]
- Lammers, K.M.; Khandelwal, S.; Chaudhry, F.; Kryszak, D.; Puppa, E.L.; Casolaro, V.; Fasano, A. Identification of a novel immunomodulatory gliadin peptide that causes interleukin-8 release in a chemokine receptor CXCR3-dependent manner only in patients with coeliac disease. Immunology 2011, 132, 432–440. [Google Scholar] [CrossRef]
- Sapone, A.; Lammers, K.M.; Mazzarella, G.; Mikhailenko, I.; Carteni, M.; Casolaro, V.; Fasano, A. Differential mucosal IL-17 expression in two gliadin-induced disorders: Gluten sensitivity and the autoimmune enteropathy celiac disease. Int. Arch. Allergy Immunol. 2010, 152, 75–80. [Google Scholar]
- Catassi, C.; Pierani, P.; Natalini, G.; Gabrielli, O.; Coppa, G.V.; Giorgi, P.L. Clinical application of a simple HPLC method for the sugar intestinal permeability test. J. Pediatr. Gastroenterol. Nutr. 1991, 12, 209–212. [Google Scholar] [CrossRef]
- Fasano, A. Leaky gut and autoimmune diseases. Clin. Rev. Allergy Immunol. 2012, 42, 71–78. [Google Scholar] [CrossRef]
- van Elburg, R.M.; Uil, J.J.; Mulder, C.J.; Heymans, H.S. Intestinal permeability in patients with coeliac disease and relatives of patients with coeliac disease. Gut 1993, 34, 354–357. [Google Scholar] [CrossRef]
- Sander, G.R.; Cummins, A.G.; Henshall, T.; Powell, B.C. Rapid disruption of intestinal barrier function by gliadin involves altered expression of apical junctional proteins. FEBS Lett. 2005, 579, 4851–4855. [Google Scholar] [CrossRef]
- Drago, S.; El Asmar, R.; Di Pierro, M.; Grazia Clemente, M.; Tripathi, A.; Sapone, A.; Thakar, M.; Iacono, G.; Carroccio, A.; D'Agate, C.; et al. Gliadin, zonulin and gut permeability: Effects on celiac and non-celiac intestinal mucosa and intestinal cell lines. Scand. J. Gastroenterol. 2006, 41, 408–419. [Google Scholar]
- Lammers, K.M.; Lu, R.; Brownley, J.; Lu, B.; Gerard, C.; Thomas, K.; Rallabhandi, P.; Shea-Donohue, T.; Tamiz, A.; Alkan, S.; et al. Gliadin induces an increase in intestinal permeability and zonulin release by binding to the chemokine receptor CXCR3. Gastroenterology 2008, 135, 194–204. [Google Scholar] [CrossRef]
- Cordain, L.; Toohey, L.; Smith, M.J.; Hickey, M.S. Modulation of immune function by dietary lectins in rheumatoid arthritis. Br. J. Nutr. 2000, 83, 207–217. [Google Scholar]
- Secondulfo, M.; Iafusco, D.; Carratu, R.; deMagistris, L.; Sapone, A.; Generoso, M.; Mezzogiomo, A.; Sasso, F.C.; Carteni, M.; de Rosa, R.; et al. Ultrastructural mucosal alterations and increased intestinal permeability in non-celiac, type I diabetic patients. Dig. Liver Dis. 2004, 36, 35–45. [Google Scholar] [CrossRef]
- Keita, A.V.; Soderholm, J.D. The intestinal barrier and its regulation by neuroimmune factors. Neurogastroenterol. Motil. 2010, 22, 718–733. [Google Scholar] [CrossRef]
- Hijazi, Z.; Molla, A.M.; Al-Habashi, H.; Muawad, W.M.; Molla, A.M.; Sharma, P.N. Intestinal permeability is increased in bronchial asthma. Arch. Dis. Child. 2004, 89, 227–229. [Google Scholar] [CrossRef]
- Maes, M. An intriguing and hitherto unexplained co-occurrence: Depression and chronic fatigue syndrome are manifestations of shared inflammatory, oxidative and nitrosative (IO&NS) pathways. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 784–794. [Google Scholar]
- Maes, M.; Kubera, M.; Leunis, J.C.; Berk, M. Increased IgA and IgM responses against gut commensals in chronic depression: Further evidence for increased bacterial translocation or leaky gut. J. Affect. Disord. 2012, 141, 55–62. [Google Scholar] [CrossRef]
- Maes, M.; Mihaylova, I.; Leunis, J.C. Increased serum IgA and IgM against LPS of enterobacteria in chronic fatigue syndrome (CFS): Indication for the involvement of gram-negative enterobacteria in the etiology of CFS and for the presence of an increased gut-intestinal permeability. J. Affect. Disord. 2007, 99, 237–240. [Google Scholar] [CrossRef]
- Sonier, B.; Patrick, C.; Ajjikuttira, P.; Scott, F.W. Intestinal immune regulation as a potential diet-modifiable feature of gut inflammation and autoimmunity. Int. Rev. Immunol. 2009, 28, 414–445. [Google Scholar] [CrossRef]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Fasano, A. Zonulin, regulation of tight junctions, and autoimmune diseases. Ann. N. Y. Acad. Sci. 2012, 1258, 25–33. [Google Scholar] [CrossRef]
- Papp, M.; Foldi, I.; Nemes, E.; Udvardy, M.; Harsfalvi, J.; Altorjay, I.; Mate, I.; Dinya, T.; Varvolgyi, C.; Barta, Z.; et al. Haptoglobin polymorphism: a novel genetic risk factor for celiac disease development and its clinical manifestations. Clin. Chem. 2008, 54, 697–704. [Google Scholar] [CrossRef]
- Carter, K.; Worwood, M. Haptoglobin: a review of the major allele frequencies worldwide and their association with diseases. Int. J. Lab. Hematol. 2007, 29, 92–110. [Google Scholar] [CrossRef]
- Fasano, A. Zonulin and its regulation of intestinal barrier function: The biological door to inflammation, autoimmunity, and cancer. Physiol. Rev. 2011, 91, 151–175. [Google Scholar] [CrossRef]
- Pusztai, A.; Ewen, S.W.; Grant, G.; Brown, D.S.; Stewart, J.C.; Peumans, W.J.; Van Damme, E.J.; Bardocz, S. Antinutritive effects of wheat-germ agglutinin and other N-acetylglucosamine-specific lectins. Br. J. Nutr. 1993, 70, 313–321. [Google Scholar] [CrossRef]
- Freed, D.L.J. Lectins in food: Their importance in health and disease. J. Nutr. Med. 1991, 2, 45–64. [Google Scholar] [CrossRef]
- Cordain, L. Cereal grains: Humanity’s double-edged sword. World Rev. Nutr. Diet. 1999, 84, 19–73. [Google Scholar] [CrossRef]
- Peumans, W.J.; Van Damme, E.J. Prevalence, biological activity and genetic manipulation of lectins in foods. Trends Food Sci. Technol. 1996, 7, 132–138. [Google Scholar] [CrossRef]
- Nachbar, M.S.; Oppenheim, J.D.; Thomas, J.O. Lectins in the U.S. Diet. Isolation and characterization of a lectin from the tomato (Lycopersicon esculentum). J. Biol. Chem. 1980, 255, 2056–2061. [Google Scholar]
- Ortega-Barria, E.; Ward, H.D.; Keusch, G.T.; Pereira, M.E. Growth inhibition of the intestinal parasite Giardia lamblia by a dietary lectin is associated with arrest of the cell cycle. J. Clin. Invest. 1994, 94, 2283–2288. [Google Scholar] [CrossRef]
- Vincenzi, S.; Zoccatelli, G.; Perbellini, F.; Rizzi, C.; Chignola, R.; Curioni, A.; Peruffo, A.D. Quantitative determination of dietary lectin activities by enzyme-linked immunosorbent assay using specific glycoproteins immobilized on microtiter plates. J. Agric. Food Chem. 2002, 50, 6266–6270. [Google Scholar] [CrossRef]
- Matucci, A.; Veneri, G.; Dalla Pellegrina, C.; Zoccatelli, G.; Vincenzi, S.; Chignola, R.; Peruffo, A.; Rizzi, C. Temperature-dependent decay of wheat germ agglutinin activity and its implications for food processing and analysis. Food Control 2004, 15, 391–395. [Google Scholar] [CrossRef]
- Shaw, L.; Yousefi, S.; Dennis, J.W.; Schauer, R. CMP-N-acetylneuraminic acid hydroxylase activity determines the wheat germ agglutinin-binding phenotype in two mutants of the lymphoma cell line MDAY-D2. Glycoconj. J. 1991, 8, 434–441. [Google Scholar] [CrossRef]
- Severi, E.; Hood, D.W.; Thomas, G.H. Sialic acid utilization by bacterial pathogens. Microbiology 2007, 153, 2817–2822. [Google Scholar] [CrossRef]
- Varki, A. Multiple changes in sialic acid biology during human evolution. Glycoconj. J. 2009, 26, 231–245. [Google Scholar] [CrossRef]
- Varki, A. Colloquium paper: Uniquely human evolution of sialic acid genetics and biology. Proc. Natl. Acad. Sci. USA 2010, 107, 8939–8946. [Google Scholar] [CrossRef]
- Lansman, J.B.; Cochrane, D.E. Wheat germ agglutinin stimulates exocytotic histamine secretion from rat mast cells in the absence of extracellular calcium. Biochem. Pharmacol. 1980, 29, 455–458. [Google Scholar] [CrossRef]
- Karlsson, A. Wheat germ agglutinin induces NADPH-oxidase activity in human neutrophils by interaction with mobilizable receptors. Infect. Immun. 1999, 67, 3461–3468. [Google Scholar]
- Haas, H.; Falcone, F.H.; Schramm, G.; Haisch, K.; Gibbs, B.F.; Klaucke, J.; Poppelmann, M.; Becker, W.M.; Gabius, H.J.; Schlaak, M. Dietary lectins can induce in vitro release of IL-4 and IL-13 from human basophils. Eur. J. Immunol. 1999, 29, 918–927. [Google Scholar] [CrossRef]
- Reed, J.C.; Robb, R.J.; Greene, W.C.; Nowell, P.C. Effect of wheat germ agglutinin on the interleukin pathway of human T lymphocyte activation. J. Immunol. 1985, 134, 314–323. [Google Scholar]
- Muraille, E.; Pajak, B.; Urbain, J.; Leo, O. Carbohydrate-bearing cell surface receptors involved in innate immunity: interleukin-12 induction by mitogenic and nonmitogenic lectins. Cell Immunol. 1999, 191, 1–9. [Google Scholar] [CrossRef]
- Sodhi, A.; Kesherwani, V. Production of TNF-alpha, IL-1beta, IL-12 and IFN-gamma in murine peritoneal macrophages on treatment with wheat germ agglutinin in vitro: involvement of tyrosine kinase pathways. Glycoconj. J. 2007, 24, 573–582. [Google Scholar] [CrossRef]
- Dalla Pellegrina, C.; Perbellini, O.; Scupoli, M.T.; Tomelleri, C.; Zanetti, C.; Zoccatelli, G.; Fusi, M.; Peruffo, A.; Rizzi, C.; Chignola, R. Effects of wheat germ agglutinin on human gastrointestinal epithelium: insights from an experimental model of immune/epithelial cell interaction. Toxicol. Appl. Pharmacol. 2009, 237, 146–153. [Google Scholar] [CrossRef]
- Schumacher, U.; Grafin von Armansperg, N.; Kreipe, H.; Welsch, U. Lectin binding and uptake in human (myelo)monocytic cell lines: HL60 and U937. Ultrastruct. Pathol. 1996, 20, 463–471. [Google Scholar] [CrossRef]
- Tchernychev, B.; Wilchek, M. Natural human antibodies to dietary lectins. FEBS Lett. 1996, 397, 139–142. [Google Scholar] [CrossRef]
- Sollid, L.M.; Kolberg, J.; Scott, H.; Ek, J.; Fausa, O.; Brandtzaeg, P. Antibodies to wheat germ agglutinin in coeliac disease. Clin. Exp. Immunol. 1986, 63, 95–100. [Google Scholar]
- Gabor, F.; Bogner, E.; Weissenboeck, A.; Wirth, M. The lectin-cell interaction and its implications to intestinal lectin-mediated drug delivery. Adv. Drug Deliv. Rev. 2004, 56, 459–480. [Google Scholar] [CrossRef]
- Clark, M.A.; Jepson, M.A.; Simmons, N.L.; Booth, T.A.; Hirst, B.H. Differential expression of lectin-binding sites defines mouse intestinal M-cells. J. Histochem. Cytochem. 1993, 41, 1679–1687. [Google Scholar] [CrossRef]
- Giannasca, P.J.; Giannasca, K.T.; Leichtner, A.M.; Neutra, M.R. Human intestinal M cells display the sialyl Lewis A antigen. Infect. Immun. 1999, 67, 946–953. [Google Scholar]
- Lochner, N.; Pittner, F.; Wirth, M.; Gabor, F. Wheat germ agglutinin binds to the epidermal growth factor receptor of artificial Caco-2 membranes as detected by silver nanoparticle enhanced fluorescence. Pharm. Res. 2003, 20, 833–839. [Google Scholar] [CrossRef]
- Flohe, S.B.; Wasmuth, H.E.; Kerad, J.B.; Beales, P.E.; Pozzilli, P.; Elliott, R.B.; Hill, J.P.; Scott, F.W.; Kolb, H. A wheat-based, diabetes-promoting diet induces a Th1-type cytokine bias in the gut of NOD mice. Cytokine 2003, 21, 149–154. [Google Scholar] [CrossRef]
- Chignola, R.; Rizzi, C.; Vincenzi, S.; Cestari, T.; Brutti, N.; Riviera, A.P.; Sartoris, S.; Peruffo, A.D.; Andrighetto, G. Effects of dietary wheat germ deprivation on the immune system in Wistar rats: a pilot study. Int. Immunopharmacol. 2002, 2, 1495–1501. [Google Scholar] [CrossRef]
- Jonnalagadda, S.S.; Harnack, L.; Liu, R.H.; McKeown, N.; Seal, C.; Liu, S.; Fahey, G.C. Putting the whole grain puzzle together: Health benefits associated with whole grains—summary of American Society for Nutrition 2010 Satellite Symposium. J. Nutr. 2011, 141, 1011S–1022S. [Google Scholar] [CrossRef]
- Lefevre, M.; Jonnalagadda, S. Effect of whole grains on markers of subclinical inflammation. Nutr. Rev. 2012, 70, 387–396. [Google Scholar] [CrossRef]
- Andersson, A.; Tengblad, S.; Karlstrom, B.; Kamal-Eldin, A.; Landberg, R.; Basu, S.; Aman, P.; Vessby, B. Whole-grain foods do not affect insulin sensitivity or markers of lipid peroxidation and inflammation in healthy, moderately overweight subjects. J. Nutr. 2007, 137, 1401–1407. [Google Scholar]
- Brownlee, I.A.; Moore, C.; Chatfield, M.; Richardson, D.P.; Ashby, P.; Kuznesof, S.A.; Jebb, S.A.; Seal, C.J. Markers of cardiovascular risk are not changed by increased whole-grain intake: the WHOLEheart study, a randomised, controlled dietary intervention. Br. J. Nutr. 2010, 104, 125–134. [Google Scholar] [CrossRef]
- Giacco, R.; Clemente, G.; Cipriano, D.; Luongo, D.; Viscovo, D.; Patti, L.; Di Marino, L.; Giacco, A.; Naviglio, D.; Bianchi, M.A.; et al. Effects of the regular consumption of wholemeal wheat foods on cardiovascular risk factors in healthy people. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 186–194. [Google Scholar] [CrossRef]
- Katcher, H.I.; Legro, R.S.; Kunselman, A.R.; Gillies, P.J.; Demers, L.M.; Bagshaw, D.M.; Kris-Etherton, P.M. The effects of a whole grain-enriched hypocaloric diet on cardiovascular disease risk factors in men and women with metabolic syndrome. Am. J. Clin. Nutr. 2008, 87, 79–90. [Google Scholar]
- Tighe, P.; Duthie, G.; Vaughan, N.; Brittenden, J.; Simpson, W.G.; Duthie, S.; Mutch, W.; Wahle, K.; Horgan, G.; Thies, F. Effect of increased consumption of whole-grain foods on blood pressure and other cardiovascular risk markers in healthy middle-aged persons: A randomized controlled trial. Am. J. Clin. Nutr. 2010, 92, 733–740. [Google Scholar] [CrossRef]
- Wolever, T.M.; Gibbs, A.L.; Mehling, C.; Chiasson, J.L.; Connelly, P.W.; Josse, R.G.; Leiter, L.A.; Maheux, P.; Rabasa-Lhoret, R.; Rodger, N.W.; et al. The Canadian Trial of Carbohydrates in Diabetes (CCD), a 1-y controlled trial of low-glycemic-index dietary carbohydrate in type 2 diabetes: No effect on glycated hemoglobin but reduction in C-reactive protein. Am. J. Clin. Nutr. 2008, 87, 114–125. [Google Scholar]
- Kyro, C.; Skeie, G.; Dragsted, L.O.; Christensen, J.; Overvad, K.; Hallmans, G.; Johansson, I.; Lund, E.; Slimani, N.; Johnsen, N.F.; et al. Intake of whole grain in Scandinavia: intake, sources and compliance with new national recommendations. Scand. J. Public Health 2012, 40, 76–84. [Google Scholar] [CrossRef]
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef]
- Jonsson, T.; Ahren, B.; Pacini, G.; Sundler, F.; Wierup, N.; Steen, S.; Sjoberg, T.; Ugander, M.; Frostegard, J.; Goransson, L.; et al. A Paleolithic diet confers higher insulin sensitivity, lower C-reactive protein and lower blood pressure than a cereal-based diet in domestic pigs. Nutr. Metab. (Lond.) 2006, 3. [Google Scholar] [CrossRef]
- Frassetto, L.A.; Schloetter, M.; Mietus-Synder, M.; Morris, R.C., Jr.; Sebastian, A. Metabolic and physiologic improvements from consuming a paleolithic, hunter-gatherer type diet. Eur. J. Clin. Nutr. 2009, 63, 947–955. [Google Scholar] [CrossRef]
- Lindeberg, S.; Jonsson, T.; Granfeldt, Y.; Borgstrand, E.; Soffman, J.; Sjostrom, K.; Ahren, B. A Palaeolithic diet improves glucose tolerance more than a Mediterranean-like diet in individuals with ischaemic heart disease. Diabetologia 2007, 50, 1795–1807. [Google Scholar] [CrossRef]
- Jonsson, T.; Granfeldt, Y.; Ahren, B.; Branell, U.C.; Palsson, G.; Hansson, A.; Soderstrom, M.; Lindeberg, S. Beneficial effects of a Paleolithic diet on cardiovascular risk factors in type 2 diabetes: A randomized cross-over pilot study. Cardiovasc. Diabetol. 2009, 8. [Google Scholar] [CrossRef]
- Matricon, J.; Meleine, M.; Gelot, A.; Piche, T.; Dapoigny, M.; Muller, E.; Ardid, D. Review article: Associations between immune activation, intestinal permeability and the irritable bowel syndrome. Aliment. Pharmacol. Ther. 2012, 36, 1009–1031. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
De Punder, K.; Pruimboom, L. The Dietary Intake of Wheat and other Cereal Grains and Their Role in Inflammation. Nutrients 2013, 5, 771-787. https://doi.org/10.3390/nu5030771
De Punder K, Pruimboom L. The Dietary Intake of Wheat and other Cereal Grains and Their Role in Inflammation. Nutrients. 2013; 5(3):771-787. https://doi.org/10.3390/nu5030771
Chicago/Turabian StyleDe Punder, Karin, and Leo Pruimboom. 2013. "The Dietary Intake of Wheat and other Cereal Grains and Their Role in Inflammation" Nutrients 5, no. 3: 771-787. https://doi.org/10.3390/nu5030771
APA StyleDe Punder, K., & Pruimboom, L. (2013). The Dietary Intake of Wheat and other Cereal Grains and Their Role in Inflammation. Nutrients, 5(3), 771-787. https://doi.org/10.3390/nu5030771