Space Flight Calcium: Implications for Astronaut Health, Spacecraft Operations, and Earth

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Bone Loss
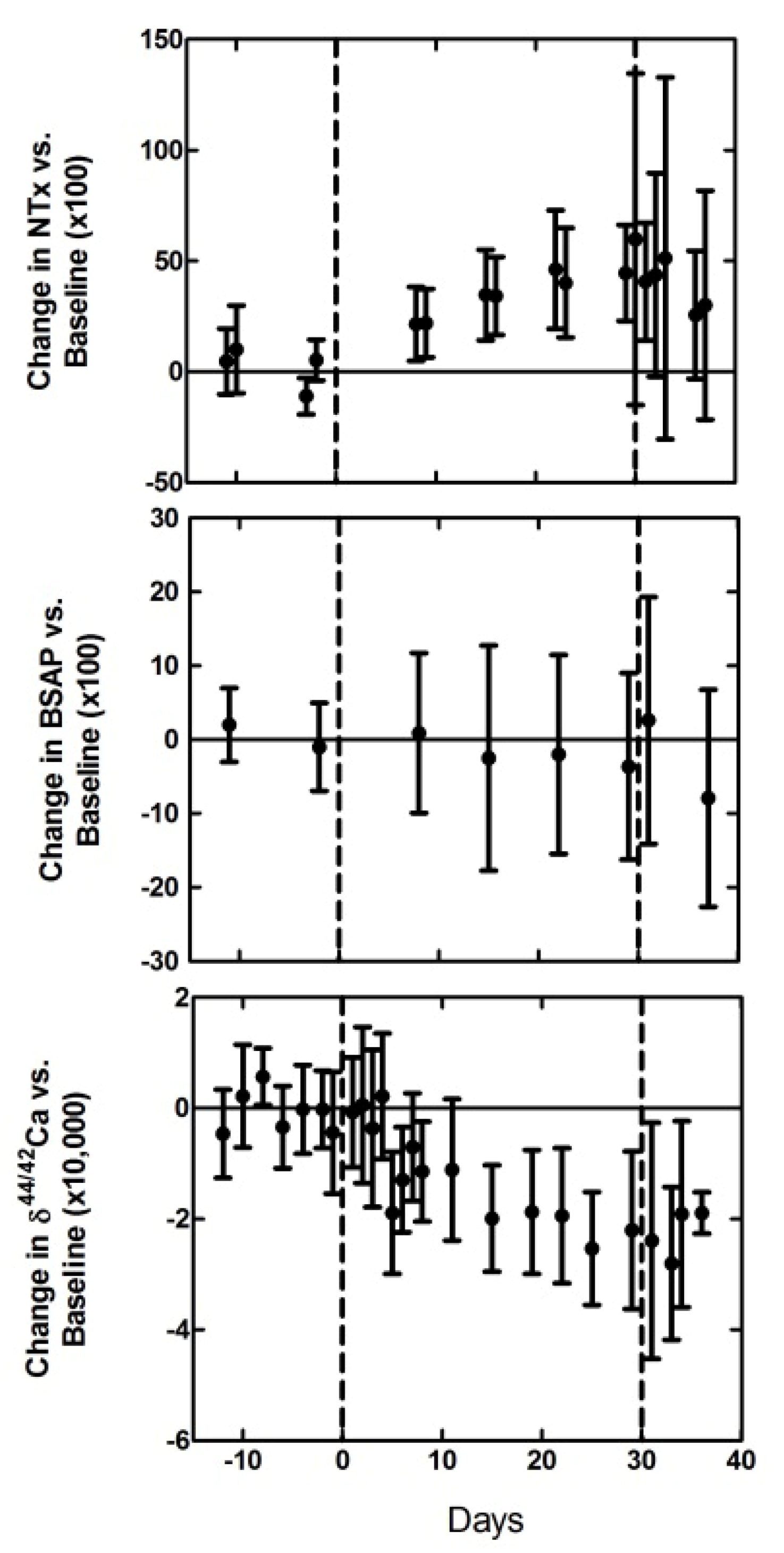
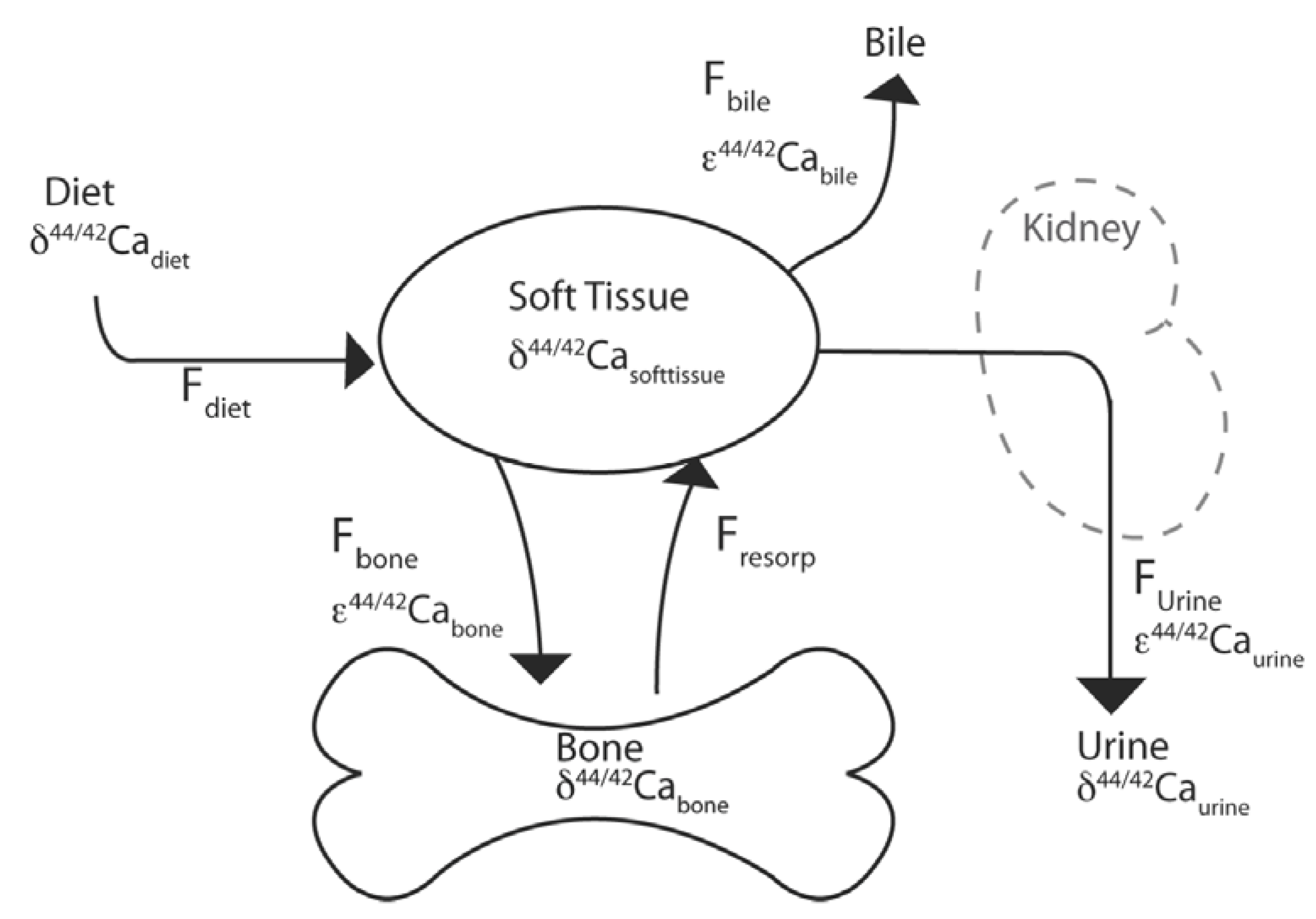
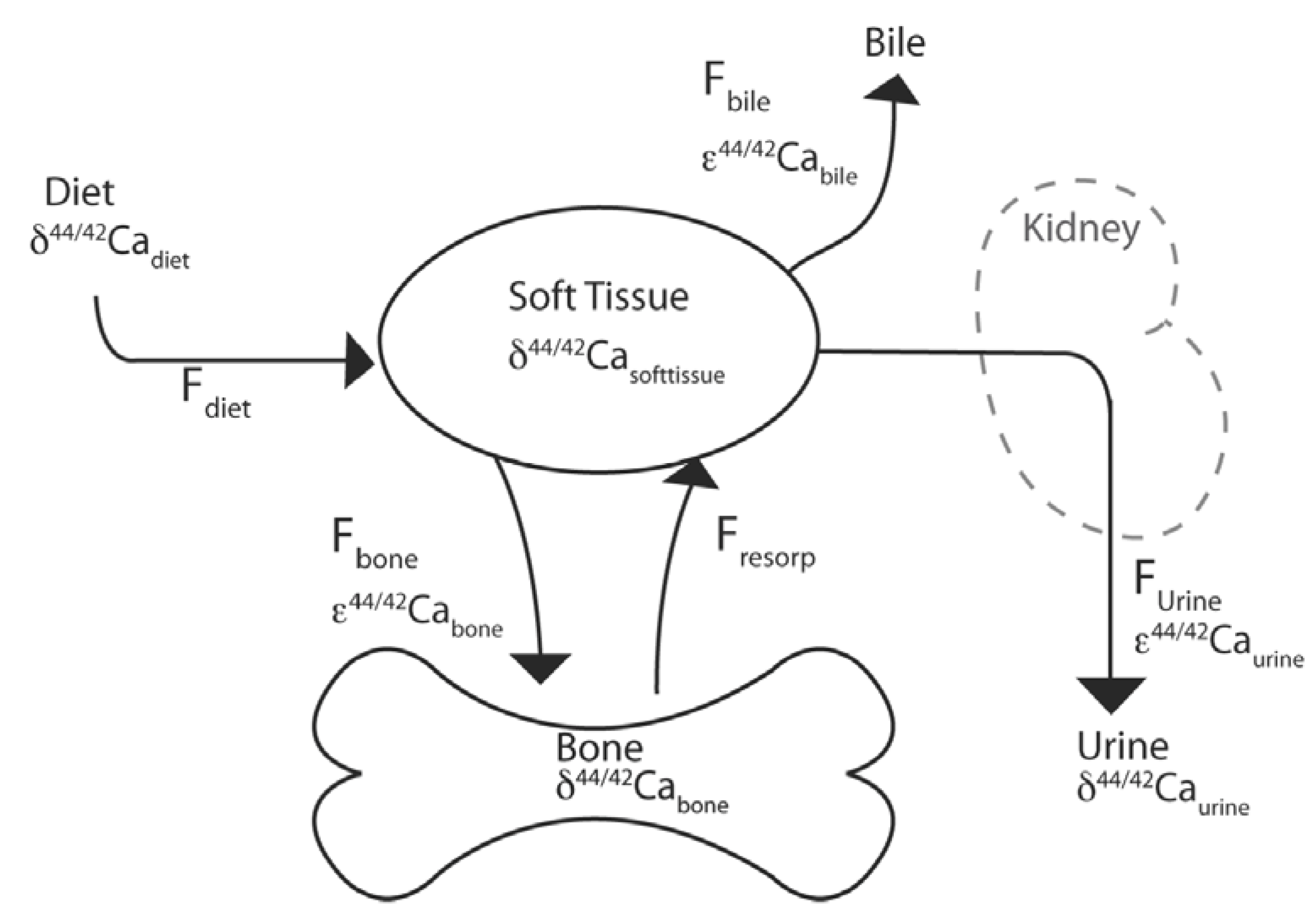
3. Calcium Isotopes and Relevance for Bone Turnover

4. Dietary Calcium
5. Calcium Absorption, Metabolism, and Excretion
6. Bone Loss Countermeasures
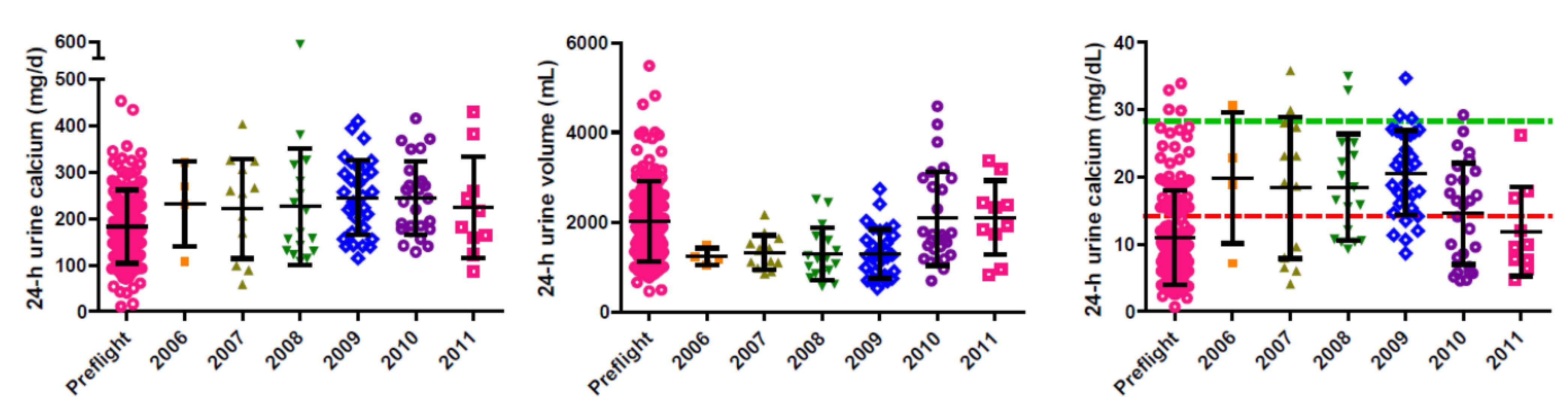
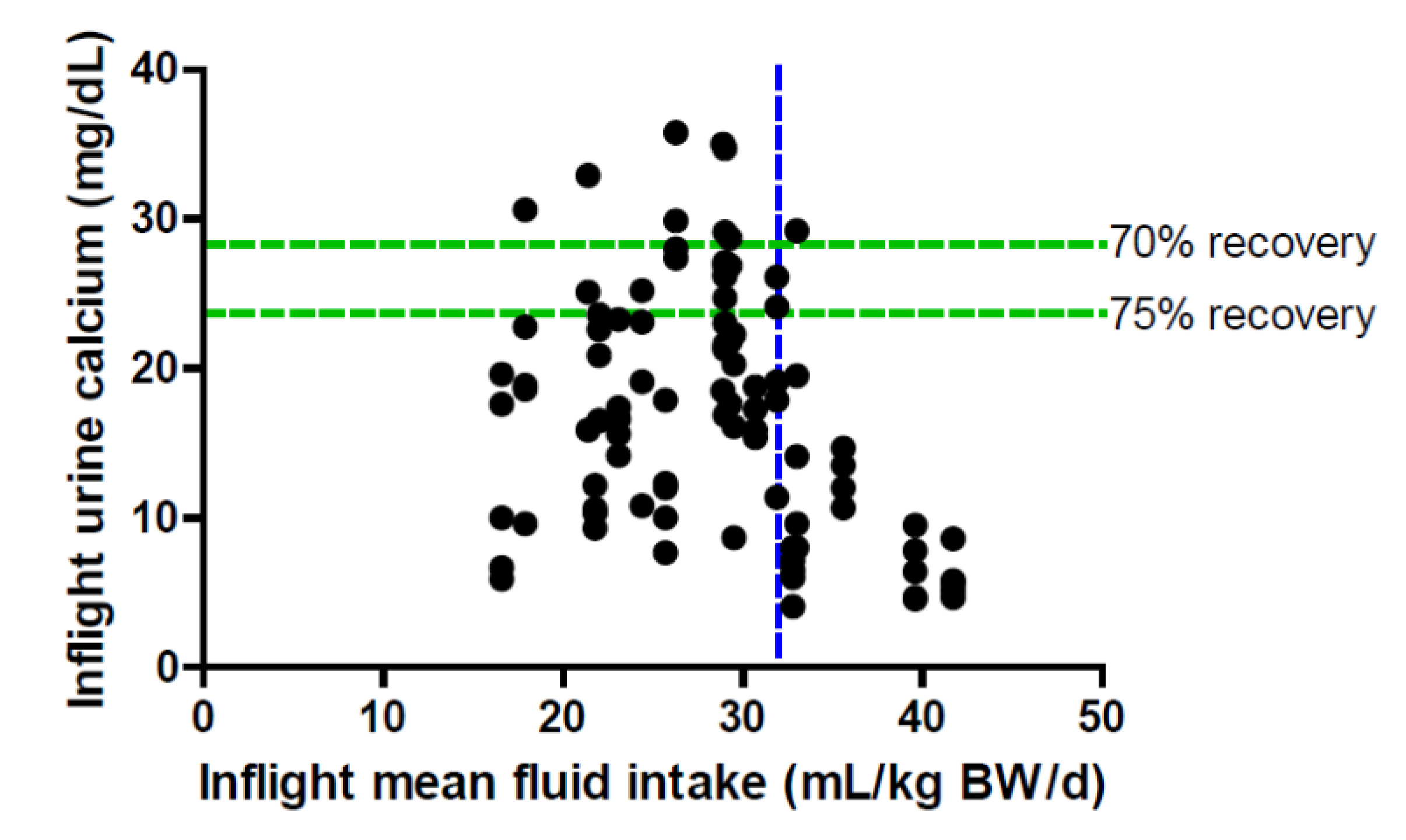
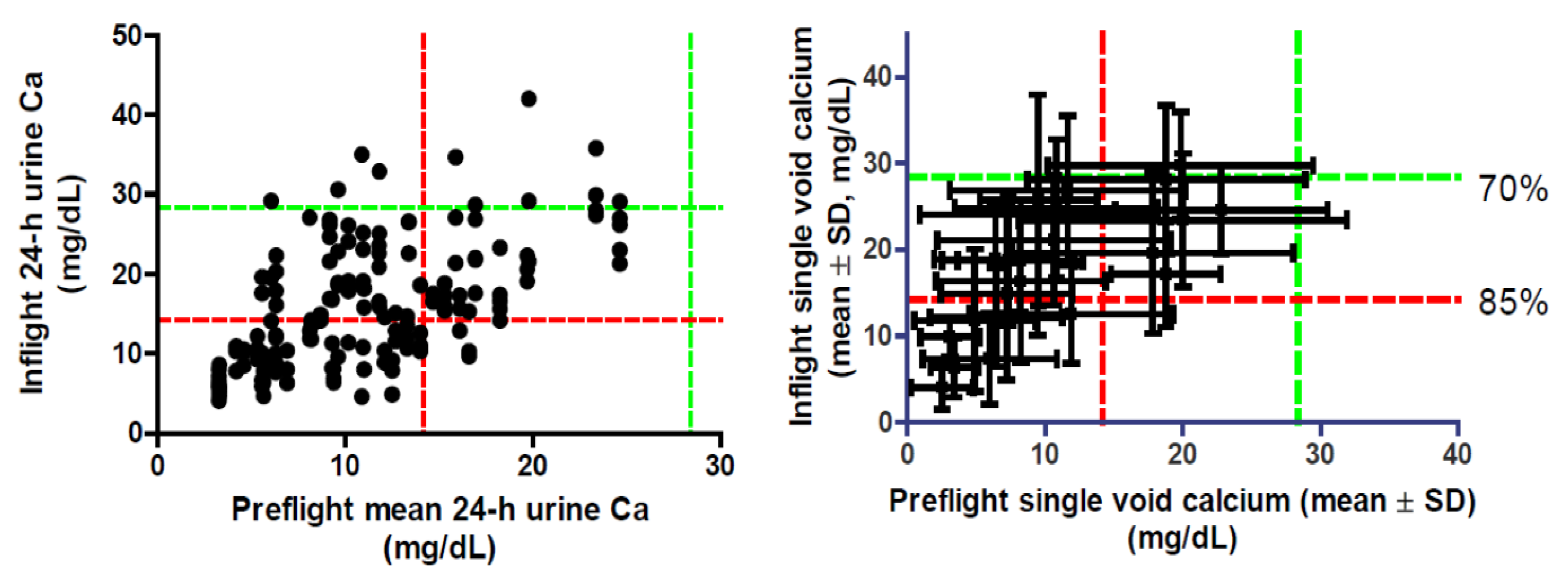
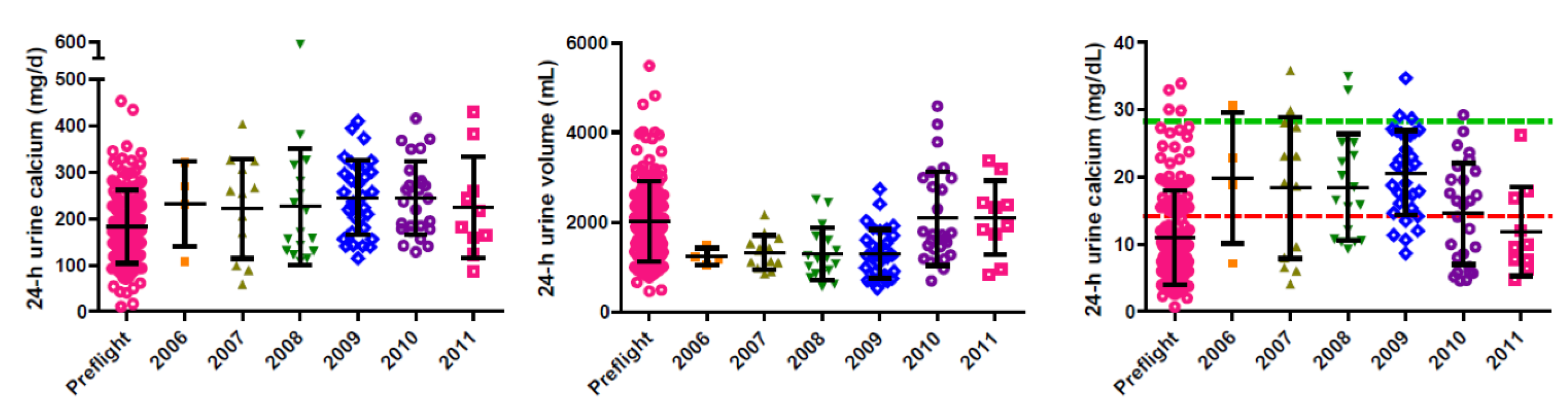
7. Urinary Calcium and Renal Stone Risk
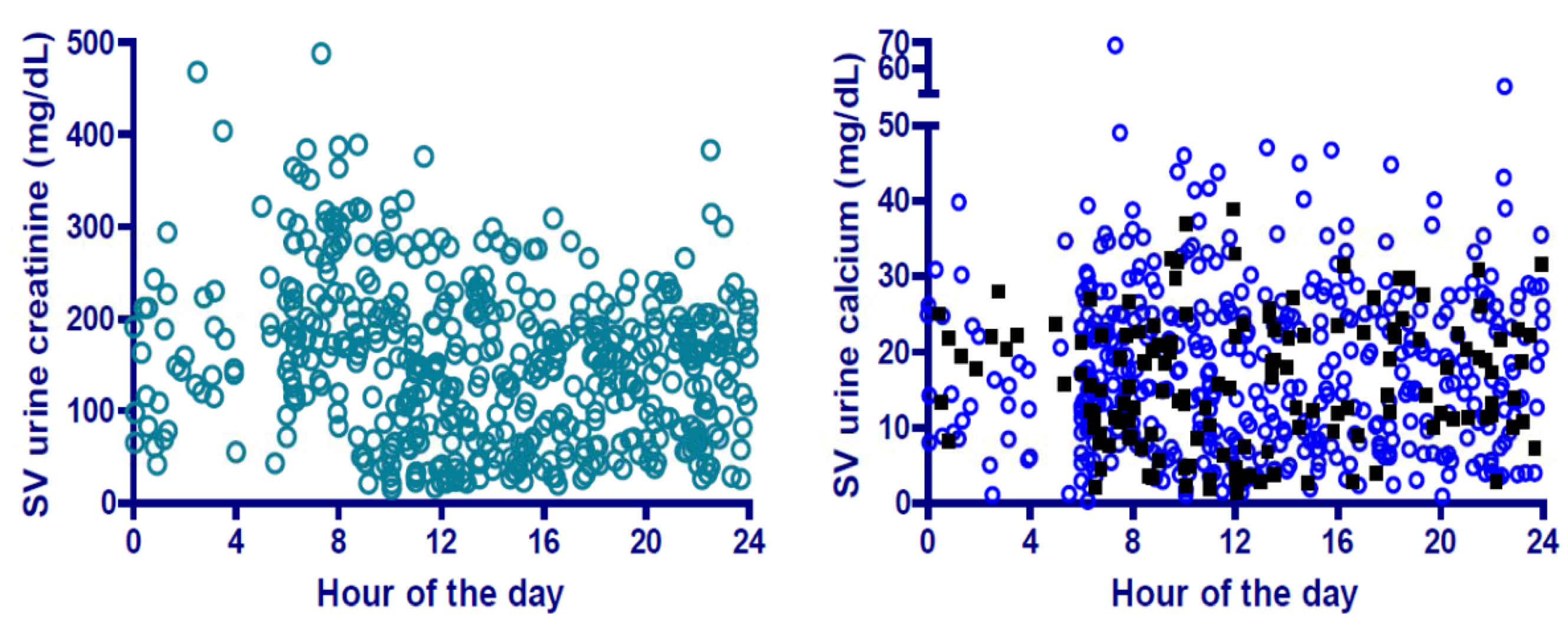
8. Urinary Calcium and Spacecraft Water Reclamation

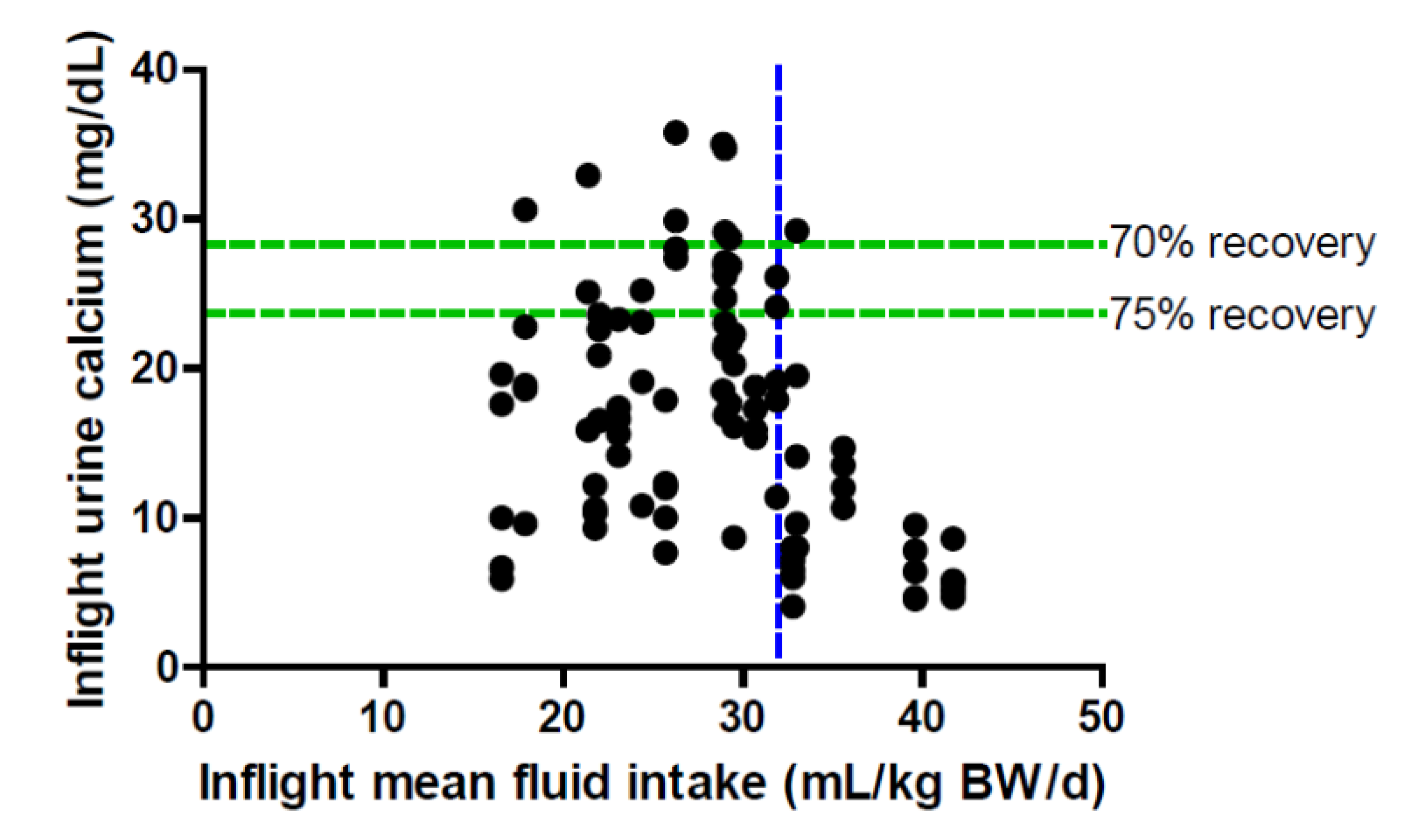
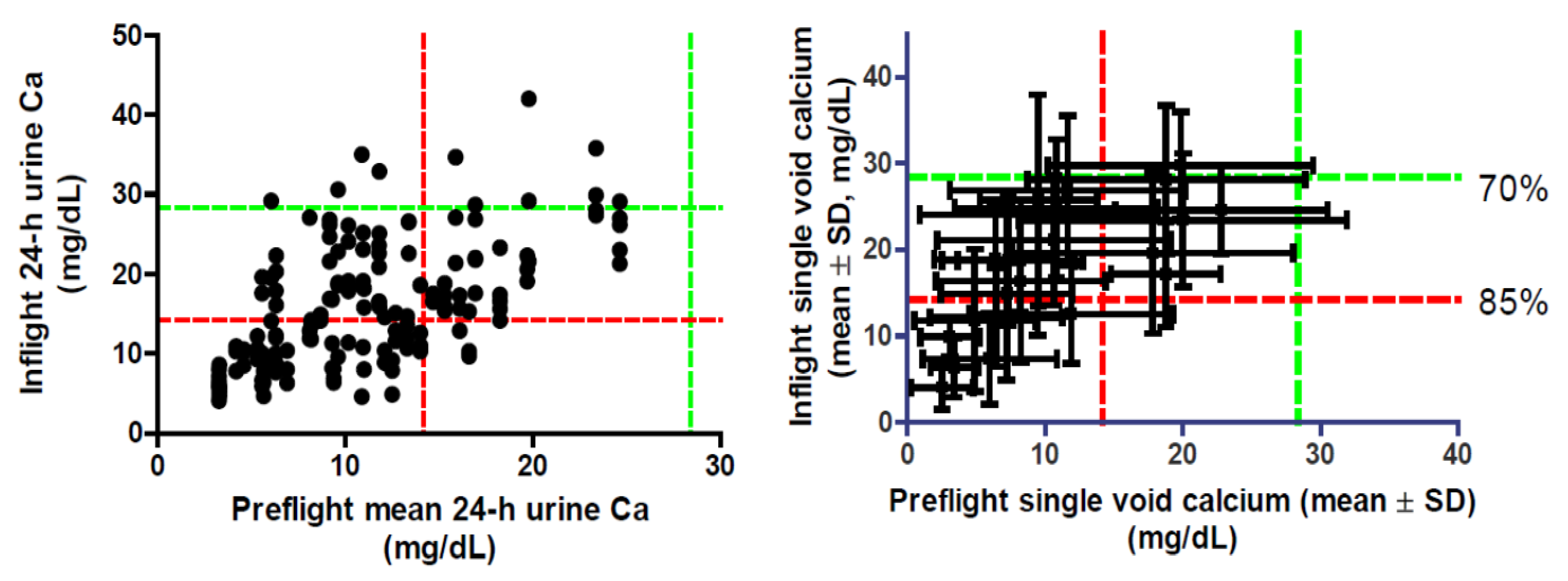

9. Conclusions
Conflict of Interest
References
- Sibonga, J.D.; Cavanagh, P.R.; Lang, T.F.; LeBlanc, A.D.; Schneider, V.S.; Shackelford, L.C.; Smith, S.M.; Vico, L. Adaptation of the skeletal system during long-duration spaceflight. Clin. Rev. Bone Miner. Metab. 2008, 5, 249–261. [Google Scholar]
- LeBlanc, A.; Schneider, V.; Shackelford, L.; West, S.; Oganov, V.; Bakulin, A.; Voronin, L. Bone mineral and lean tissue loss after long duration space flight. J. Musculoskelet. Neuronal Interact. 2000, 1, 157–160. [Google Scholar]
- Alexandre, C.; Vico, L. Pathophysiology of bone loss in disuse osteoporosis. Joint Bone Spine 2011, 78, 572–576. [Google Scholar] [CrossRef]
- Whedon, G.D.; Rambaut, P.C. Effects of long-duration spaceflight on calcium metabolism: Review of human studies from Skylab to the present. Acta Astronaut. 2006, 58, 59–81. [Google Scholar]
- Heer, M.; Kamps, N.; Biener, C.; Korr, C.; Boerger, A.; Zittermann, A.; Stehle, P.; Drummer, C. Calcium metabolism in microgravity. Eur. J. Med. Res. 1999, 4, 357–360. [Google Scholar]
- LeBlanc, A.D.; Spector, E.R.; Evans, H.J.; Sibonga, J.D. Skeletal responses to space flight and the bed rest analog: A review. J. Musculoskelet. Neuronal Interact. 2007, 7, 33–47. [Google Scholar]
- Pavy-Le Traon, A.; Heer, M.; Narici, M.V.; Rittweger, J.; Vernikos, J. From space to Earth: Advances in human physiology from 20 years of bed rest studies (1986–2006). Eur. J. Appl. Physiol. 2007, 101, 143–194. [Google Scholar] [CrossRef]
- Smith, S.M.; Zwart, S.R.; Kloeris, V.; Heer, M. Nutritional Biochemistry of Space Flight; Nova Science Publishers: New York, NY, USA, 2009. [Google Scholar]
- Smith, S.M.; Zwart, S.R. Nutritional biochemistry of spaceflight. Adv. Clin. Chem. 2008, 46, 87–130. [Google Scholar]
- Lutwak, L.; Whedon, G.D.; Lachance, P.A.; Reid, J.M.; Lipscomb, H.S. Mineral, electrolyte and nitrogen balance studies of the Gemini-VII fourteen-day orbital space flight. J. Clin. Endocrinol. Metab. 1969, 29, 1140–1156. [Google Scholar] [CrossRef]
- Rambaut, P.C.; Leach, C.S.; Johnson, P.C. Calcium and phosphorus change of the Apollo 17 crew members. Nutr. Metab. 1975, 18, 62–69. [Google Scholar]
- Deitrick, J.E.; Whedon, G.D.; Shorr, E. Effects of immobilization upon various metabolic and physiologic functions of normal men. Am. J. Med. 1948, 4, 3–36. [Google Scholar] [CrossRef]
- Whedon, G.; Lutwak, L.; Rambaut, P.; Whittle, M.; Leach, C.; Reid, J.; Smith, M. Effect of weightlessness on mineral metabolism; metabolic studies on Skylab orbital flights. Calcif. Tissue Res. 1976, 21, 423–430. [Google Scholar]
- Whedon, G.D.; Lutwak, L.; Rambaut, P.C.; Whittle, M.W.; Smith, M.C.; Reid, J.; Leach, C.; Stadler, C.R.; Sanford, D.D. Mineral and Nitrogen Metabolic Studies, Experiment M071. In Biomedical Results from Skylab (NASA Sp-377); Johnston, R.S., Dietlein, L.F., Eds.; National Aeronautics and Space Administration: Washington, DC, USA, 1977; pp. 164–174. [Google Scholar]
- Oganov, V.S.; Grigoriev, A.I.; Voronin, L.I.; Rakhmanov, A.S.; Bakulin, A.V.; Schneider, V.S.; LeBlanc, A.D. Bone mineral density in cosmonauts after flights lasting 4.5–6 months on the Mir orbital station. Aviakosm. Ekolog. Med. 1992, 26, 20–24. [Google Scholar]
- Vico, L.; Collet, P.; Guignandon, A.; Lafage-Proust, M.H.; Thomas, T.; Rehaillia, M.; Alexandre, C. Effects of long-term microgravity exposure on cancellous and cortical weight-bearing bones of cosmonauts. Lancet 2000, 355, 1607–1611. [Google Scholar]
- Collet, P.; Uebelhart, D.; Vico, L.; Moro, L.; Hartmann, D.; Roth, M.; Alexandre, C. Effects of 1- and 6-month spaceflight on bone mass and biochemistry in two humans. Bone 1997, 20, 547–551. [Google Scholar] [CrossRef]
- Caillot-Augusseau, A.; Lafage-Proust, M.H.; Soler, C.; Pernod, J.; Dubois, F.; Alexandre, C. Bone formation and resorption biological markers in cosmonauts during and after a 180-day space flight (Euromir 95). Clin. Chem. 1998, 44, 578–585. [Google Scholar]
- Smith, S.M.; Nillen, J.L.; LeBlanc, A.; Lipton, A.; Demers, L.M.; Lane, H.W.; Leach, C.S. Collagen cross-link excretion during space flight and bed rest. J. Clin. Endocrinol. Metab. 1998, 83, 3584–3591. [Google Scholar] [CrossRef]
- Lueken, S.A.; Arnaud, S.B.; Taylor, A.K.; Baylink, D.J. Changes in markers of bone formation and resorption in a bed rest model of weightlessness. J. Bone Miner. Res. 1993, 8, 1433–1438. [Google Scholar]
- Smith, S.M.; Wastney, M.E.; Morukov, B.V.; Larina, I.M.; Nyquist, L.E.; Abrams, S.A.; Taran, E.N.; Shih, C.Y.; Nillen, J.L.; Davis-Street, J.E.; et al. Calcium metabolism before, during, and after a 3-mo spaceflight: kinetic and biochemical changes. Am. J. Physiol. 1999, 277, R1–R10. [Google Scholar]
- Smith, S.M.; Wastney, M.E.; O’Brien, K.O.; Morukov, B.V.; Larina, I.M.; Abrams, S.A.; Davis-Street, J.E.; Oganov, V.; Shackelford, L.C. Bone markers, calcium metabolism, and calcium kinetics during extended-duration space flight on the Mir space station. J. Bone Miner. Res. 2005, 20, 208–218. [Google Scholar]
- Smith, B.J.; King, J.B.; Lucas, E.A.; Akhter, M.P.; Arjmandi, B.H.; Stoecker, B.J. Skeletal unloading and dietary copper depletion are detrimental to bone quality of mature rats. J. Nutr. 2002, 132, 190–196. [Google Scholar]
- Lang, T.; LeBlanc, A.; Evans, H.; Lu, Y.; Genant, H.; Yu, A. Cortical and trabecular bone mineral loss from the spine and hip in long-duration spaceflight. J. Bone Miner. Res. 2004, 19, 1006–1012. [Google Scholar]
- Pierre, M.C.; Genc, K.O.; Litow, M.; Humphreys, B.; Rice, A.J.; Maender, C.C.; Cavanagh, P.R. Comparison of knee motion on Earth and in space: An observational study. J. Neuroengineering Rehabil. 2006, 3, 8. [Google Scholar] [CrossRef]
- Smith, S.M.; Heer, M.A.; Shackelford, L.; Sibonga, J.D.; Ploutz-Snyder, L.; Zwart, S.R. Benefits for bone from resistance exercise and nutrition in long-duration spaceflight: Evidence from biochemistry and densitometry. J. Bone Miner. Res. 2012, 27, 1896–1906. [Google Scholar]
- Morgan, J.L.; Skulan, J.L.; Gordon, G.W.; Romaniello, S.J.; Smith, S.M.; Anbar, A.D. Rapidly assessing changes in bone mineral balance using natural stable calcium isotopes. Proc. Natl. Acad. Sci. USA 2012, 109, 9989–9994. [Google Scholar]
- Skulan, J.; Bullen, T.; Anbar, A.D.; Puzas, J.E.; Shackelford, L.; LeBlanc, A.; Smith, S.M. Natural calcium isotopic composition of urine as a marker of bone mineral balance. Clin. Chem. 2007, 53, 1155–1158. [Google Scholar] [CrossRef]
- Russell, W.A.; Papanastassiou, D.A.; Tombrello, T.A. Ca isotope fractionation on earth and other solar-system materials. Geochim. Cosmochim. Acta 1978, 42, 1075–1090. [Google Scholar]
- DePaolo, D.J. Geochemistry of Non-Traditional Stable Isotopes; Johnson, C., Beard, B., Albarede, F., Eds.; Mineralogical Society of America: Washington, DC, USA, 2004. [Google Scholar]
- Criss, R.E. Principles of Stable Isotope Distribution; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Morgan, J.L.; Gordon, G.W.; Arrua, R.C.; Skulan, J.L.; Anbar, A.D.; Bullen, T.D. High-precision measurement of variations in calcium isotope ratios in urine by multiple collector inductively coupled plasma mass spectrometry. Anal. Chem. 2011, 83, 6956–6962. [Google Scholar]
- Skulan, J.L.; DePaolo, D.J. Calcium isotope fractionation between soft and mineralized tissue as a monitor of calcium use in vertebrates. Proc. Natl. Acad. Sci. USA 1999, 96, 13709–13713. [Google Scholar]
- Spector, E.R.; Smith, S.M.; Sibonga, J.D. Skeletal effects of long-duration head-down bed rest. Aviat. Space Environ. Med. 2009, 80, A23–A28. [Google Scholar] [CrossRef]
- LeBlanc, A.D.; Driscol, T.B.; Shackelford, L.C.; Evans, H.J.; Rianon, N.J.; Smith, S.M.; Feeback, D.L.; Lai, D. Alendronate as an effective countermeasure to disuse induced bone loss. J. Musculoskelet. Neuronal Interact. 2002, 2, 335–343. [Google Scholar]
- Shackelford, L.C.; LeBlanc, A.D.; Driscoll, T.B.; Evans, H.J.; Rianon, N.J.; Smith, S.M.; Spector, E.; Feeback, D.L.; Lai, D. Resistance exercise as a countermeasure to disuse-induced bone loss. J. Appl. Physiol. 2004, 97, 119–129. [Google Scholar] [CrossRef]
- Whedon, G.D.; Lutwak, L.; Reid, J.; Rambaut, P.; Whittle, M.; Smith, M.; Leach, C. Mineral and nitrogen balance study: Results of metabolic observations on Skylab II 28-day orbital mission. Acta Astronaut. 1975, 2, 297–309. [Google Scholar] [CrossRef]
- Smith, S.M.; Zwart, S.R.; Block, G.; Rice, B.L.; Davis-Street, J.E. The nutritional status of astronauts is altered after long-term space flight aboard the International Space Station. J. Nutr. 2005, 135, 437–443. [Google Scholar]
- Caillot-Augusseau, A.; Lafage-Proust, M.H.; Margaillan, P.; Vergely, N.; Faure, S.; Paillet, S.; Lang, F.; Alexandre, C.; Estour, B. Weight gain reverses bone turnover and restores circadian variation of bone resorption in anorexic patients. Clin. Endocrinol. (Oxf.) 2000, 52, 113–121. [Google Scholar] [CrossRef]
- Smith, S.M.; Davis-Street, J.E.; Fontenot, T.B.; Lane, H.W. Assessment of a portable clinical blood analyzer during space flight. Clin. Chem. 1997, 43, 1056–1065. [Google Scholar]
- Zittermann, A.; Heer, M.; Caillot-Augusso, A.; Rettberg, P.; Scheld, K.; Drummer, C.; Alexandre, C.; Horneck, G.; Vorobiev, D. Microgravity inhibits intestinal calcium absorption as shown by a stable strontium test. Eur. J. Clin. Invest. 2000, 30, 1036–1043. [Google Scholar] [CrossRef]
- LeBlanc, A.; Schneider, V.; Spector, E.; Evans, H.; Rowe, R.; Lane, H.; Demers, L.; Lipton, A. Calcium absorption, endogenous excretion, and endocrine changes during and after long-term bed rest. Bone 1995, 16, 301S–304S. [Google Scholar]
- Baecker, N.; Frings-Meuthen, P.; Smith, S.M.; Heer, M. Short-term high dietary calcium intake during bedrest has no effect on markers of bone turnover in healthy men. Nutrition 2010, 26, 522–527. [Google Scholar] [CrossRef]
- Smith, S.M.; Zwart, S.R.; Heer, M.A.; Baecker, N.; Evans, H.J.; Feiveson, A.H.; Shackelford, L.C.; Leblanc, A.D. Effects of artificial gravity during bed rest on bone metabolism in humans. J. Appl. Physiol. 2009, 107, 47–53. [Google Scholar]
- Morgan, J.L.; Zwart, S.R.; Heer, M.; Ploutz-Snyder, R.; Ericson, K.; Smith, S.M. Bone metabolism and nutritional status during 30-day head-down tilt bed rest. J. Appl. Physiol. 2012, in press.. [Google Scholar]
- Smith, S.M.; Davis-Street, J.E.; Fesperman, J.V.; Calkins, D.S.; Bawa, M.; Macias, B.R.; Meyer, R.S.; Hargens, A.R. Evaluation of treadmill exercise in a lower body negative pressure chamber as a countermeasure for weightlessness-induced bone loss: A bed rest study with identical twins. J. Bone Miner. Res. 2003, 18, 2223–2230. [Google Scholar] [CrossRef]
- Smith, S.M.; Zwart, S.R.; Heer, M.; Lee, S.M.C.; Baecker, N.; Meuche, S.; Macias, B.R.; Shackelford, L.C.; Schneider, S.; Hargens, A.R. WISE-2005: Supine treadmill exercise within lower body negative pressure and flywheel resistive exercise as a countermeasure to bed rest-induced bone loss in women during 60-day simulated microgravity. Bone 2008, 42, 572–581. [Google Scholar]
- Armbrecht, G.; Belavy, D.L.; Gast, U.; Bongrazio, M.; Touby, F.; Beller, G.; Roth, H.J.; Perschel, F.H.; Rittweger, J.; Felsenberg, D. Resistive vibration exercise attenuates bone and muscle atrophy in 56 days of bed rest: Biochemical markers of bone metabolism. Osteoporos. Int. 2010, 21, 597–607. [Google Scholar] [CrossRef]
- Baecker, N.; Tomic, A.; Mika, C.; Gotzmann, A.; Platen, P.; Gerzer, R.; Heer, M. Bone resorption is induced on the second day of bed rest: results of a controlled crossover trial. J. Appl. Physiol. 2003, 95, 977–982. [Google Scholar]
- Holguin, N.; Uzer, G.; Chiang, F.P.; Rubin, C.; Judex, S. Brief daily exposure to low-intensity vibration mitigates the degradation of the intervertebral disc in a frequency-specific manner. J. Appl. Physiol. 2011, 111, 1846–1853. [Google Scholar] [CrossRef]
- Ozcivici, E.; Luu, Y.K.; Rubin, C.T.; Judex, S. Low-level vibrations retain bone marrow’s osteogenic potential and augment recovery of trabecular bone during reambulation. PLoS One 2010, 5, e11178. [Google Scholar]
- Holguin, N.; Muir, J.; Rubin, C.; Judex, S. Short applications of very low-magnitude vibrations attenuate expansion of the intervertebral disc during extended bed rest. Spine J. 2009, 9, 470–477. [Google Scholar]
- Baecker, N.; Frings-Meuthen, P.; Heer, M.; Mester, J.; Liphardt, A.M. Effects of vibration training on bone metabolism: Results from a short-term bed rest study. Eur. J. Appl. Physiol. 2012, 112, 1741–1750. [Google Scholar] [CrossRef]
- Belavy, D.L.; Armbrecht, G.; Gast, U.; Richardson, C.A.; Hides, J.A.; Felsenberg, D. Countermeasures against lumbar spine deconditioning in prolonged bed rest: Resistive exercise with and without whole body vibration. J. Appl. Physiol. 2010, 109, 1801–1811. [Google Scholar] [CrossRef]
- Belavy, D.L.; Beller, G.; Armbrecht, G.; Perschel, F.H.; Fitzner, R.; Bock, O.; Borst, H.; Degner, C.; Gast, U.; Felsenberg, D. Evidence for an additional effect of whole-body vibration above resistive exercise alone in preventing bone loss during prolonged bed rest. Osteoporos. Int. 2010, 22, 1581–1591. [Google Scholar]
- Belavy, D.L.; Miokovic, T.; Armbrecht, G.; Rittweger, J.; Felsenberg, D. Resistive vibration exercise reduces lower limb muscle atrophy during 56-day bed-rest. J. Musculoskelet. Neuronal Interact. 2009, 9, 225–235. [Google Scholar]
- Belavy, D.L.; Wilson, S.J.; Armbrecht, G.; Rittweger, J.; Felsenberg, D.; Richardson, C.A. Resistive vibration exercise during bed-rest reduces motor control changes in the lumbo-pelvic musculature. J. Electromyogr. Kinesiol. 2012, 22, 21–30. [Google Scholar] [CrossRef]
- Blottner, D.; Salanova, M.; Puttmann, B.; Schiffl, G.; Felsenberg, D.; Buehring, B.; Rittweger, J. Human skeletal muscle structure and function preserved by vibration muscle exercise following 55 days of bed rest. Eur. J. Appl. Physiol. 2006, 97, 261–271. [Google Scholar]
- Warren, L.E.; Reinertson, R.; Camacho, M.E.; Paloski, W.H. Implementation of the NASA Artificial Gravity Bed Rest Pilot Study. J. Gravit. Physiol. 2007, 14, P1–P4. [Google Scholar]
- Clément, G.; Bukley, A.P. Artificial Gravity; Springer: New York, NY, USA, 2007; p. 364. [Google Scholar]
- Vernikos, J. Artificial gravity intermittent centrifugation as a space flight countermeasure. J. Gravit. Physiol. 1997, 4, P13–P16. [Google Scholar]
- Zwart, S.R.; Crawford, G.E.; Gillman, P.L.; Kala, G.; Rodgers, A.S.; Rogers, A.; Inniss, A.M.; Rice, B.L.; Ericson, K.; Coburn, S.; et al. Effects of 21 days of bed rest, with or without artificial gravity, on nutritional status of humans. J. Appl. Physiol. 2009, 107, 54–62. [Google Scholar] [CrossRef]
- Yang, Y.; Baker, M.; Graf, S.; Larson, J.; Caiozzo, V.J. Hypergravity resistance exercise: The use of artificial gravity as potential countermeasure to microgravity. J. Appl. Physiol. 2007, 103, 1879–1887. [Google Scholar] [CrossRef]
- Zwart, S.R.; Davis-Street, J.E.; Paddon-Jones, D.; Ferrando, A.A.; Wolfe, R.R.; Smith, S.M. Amino acid supplementation alters bone metabolism during simulated weightlessness. J. Appl. Physiol. 2005, 99, 134–140. [Google Scholar]
- Zwart, S.R.; Hargens, A.R.; Lee, S.M.; Macias, B.R.; Watenpaugh, D.E.; Tse, K.; Smith, S.M. Lower body negative pressure treadmill exercise as a countermeasure for bed rest-induced bone loss in female identical twins. Bone 2007, 40, 529–537. [Google Scholar] [CrossRef]
- Zwart, S.R.; Hargens, A.R.; Smith, S.M. The ratio of animal protein intake to potassium intake is a predictor of bone resorption in space flight analogues and in ambulatory subjects. Am. J. Clin. Nutr. 2004, 80, 1058–1065. [Google Scholar]
- Zwart, S.R.; Smith, S.M. The impact of space flight on the human skeletal system and potential nutritional countermeasures. Int. SportMed J. 2005, 6, 199–214. [Google Scholar]
- Whiting, S.J.; Boyle, J.L.; Thompson, A.; Mirwald, R.L.; Faulkner, R.A. Dietary protein, phosphorus and potassium are beneficial to bone mineral density in adult men consuming adequate dietary calcium. J. Am. Coll. Nutr. 2002, 21, 402–409. [Google Scholar]
- Heer, M.; Boerger, A.; Kamps, N.; Mika, C.; Korr, C.; Drummer, C. Nutrient supply during recent European missions. Pflugers Arch. 2000, 441, R8–R14. [Google Scholar]
- Smith, S.M.; Davis-Street, J.; Rice, B.L.; Lane, H.W. Nutrition in space. Nutr. Today 1997, 32, 6–12. [Google Scholar]
- Goulding, A. Fasting urinary sodium/creatinine in relation to calcium/creatinine and hydroxyproline/creatinine in a general population of women. N. Z. Med. J. 1981, 93, 294–297. [Google Scholar]
- Arnaud, S.B.; Wolinsky, I.; Fung, P.; Vernikos, J. Dietary salt and urinary calcium excretion in a human bed rest spaceflight model. Aviat. Space Environ. Med. 2000, 71, 1115–1119. [Google Scholar]
- Nordin, B.E.; Need, A.G.; Morris, H.A.; Horowitz, M. The nature and significance of the relationship between urinary sodium and urinary calcium in women. J. Nutr. 1993, 123, 1615–1622. [Google Scholar]
- Carbone, L.D.; Bush, A.J.; Barrow, K.D.; Kang, A.H. The relationship of sodium intake to calcium and sodium excretion and bone mineral density of the hip in postmenopausal African-American and Caucasian women. J. Bone Miner. Metab. 2003, 21, 415–420. [Google Scholar] [CrossRef]
- Chan, A.Y.; Poon, P.; Chan, E.L.; Fung, S.L.; Swaminathan, R. The effect of high sodium intake on bone mineral content in rats fed a normal calcium or a low calcium diet. Osteoporos. Int. 1993, 3, 341–344. [Google Scholar] [CrossRef]
- Chan, E.L.; Ho, C.S.; MacDonald, D.; Ho, S.C.; Chan, T.Y.; Swaminathan, R. Interrelationships between urinary sodium, calcium, hydroxyproline and serum PTH in healthy subjects. Acta Endocrinol. (Copenh.) 1992, 127, 242–245. [Google Scholar]
- Massey, L.K.; Whiting, S.J. Dietary salt, urinary calcium, and bone loss. J. Bone Miner. Res. 1996, 11, 731–736. [Google Scholar]
- Massey, L.K.; Whiting, S.J. Dietary salt, urinary calcium, and stone risk. Nutr. Rev. 1995, 53, 131–139. [Google Scholar]
- Heer, M.; Frings-Meuthen, P.; Titze, J.; Boschmann, M.; Frisch, S.; Baecker, N.; Beck, L. Increasing sodium intake from a previous low or high intake affects water, electrolyte and acid-base differently. Br. J. Nutr. 2009, 101, 1286–1294. [Google Scholar]
- Frings-Meuthen, P.; Baecker, N.; Heer, M. Low-grade metabolic acidosis may be the cause of sodium chloride-induced exaggerated bone resorption. J. Bone Miner. Res. 2008, 23, 517–524. [Google Scholar] [CrossRef]
- Arnett, T. Regulation of bone cell function by acid-base balance. Proc. Nutr. Soc. 2003, 62, 511–520. [Google Scholar] [CrossRef]
- Arnett, T.R. Extracellular pH regulates bone cell function. J. Nutr. 2008, 138, 415S–418S. [Google Scholar]
- Arnett, T.R.; Dempster, D.W. Effect of pH on bone resorption by rat osteoclasts in vitro. Endocrinology 1986, 119, 119–124. [Google Scholar] [CrossRef]
- Frings-Meuthen, P.; Buehlmeier, J.; Baecker, N.; Stehle, P.; Fimmers, R.; May, F.; Kluge, G.; Heer, M. High sodium chloride intake exacerbates immobilization-induced bone resorption and protein losses. J. Appl. Physiol. 2011, 111, 537–542. [Google Scholar]
- Zwart, S.R.; Pierson, D.; Mehta, S.; Gonda, S.; Smith, S.M. Capacity of omega-3 fatty acids or eicosapentaenoic acid to counteract weightlessness-induced bone loss by inhibiting NF-kappaB activation: From cells to bed rest to astronauts. J. Bone Miner. Res. 2010, 25, 1049–1057. [Google Scholar]
- Griel, A.E.; Kris-Etherton, P.M.; Hilpert, K.F.; Zhao, G.; West, S.G.; Corwin, R.L. An increase in dietary n-3 fatty acids decreases a marker of bone resorption in humans. Nutr. J. 2007, 6, 2–10. [Google Scholar] [CrossRef]
- Bonnet, N.; Ferrari, S.L. Effects of long-term supplementation with omega-3 fatty acids on longitudinal changes in bone mass and microstructure in mice. J. Nutr. Biochem. 2011, 22, 665–672. [Google Scholar] [CrossRef]
- Rousseau, J.H.; Kleppinger, A.; Kenny, A.M. Self-reported dietary intake of omega-3 fatty acids and association with bone and lower extremity function. J. Am. Geriatr. Soc. 2009, 57, 1781–1788. [Google Scholar]
- Tartibian, B.; Hajizadeh Maleki, B.; Kanaley, J.; Sadeghi, K. Long-term aerobic exercise and omega-3 supplementation modulate osteoporosis through inflammatory mechanisms in post-menopausal women: A randomized, repeated measures study. Nutr. Metab. 2011, 8, 71. [Google Scholar] [CrossRef]
- Watkins, B.A.; Li, Y.; Lippman, H.E.; Feng, S. Modulatory effect of omega-3 polyunsaturated fatty acids on osteoblast function and bone metabolism. Prostaglandins Leukot. Essent. Fatty Acids 2003, 68, 387–398. [Google Scholar] [CrossRef]
- Zwart, S.R.; Mehta, S.K.; Ploutz-Snyder, R.; Bourbeau, Y.; Locke, J.P.; Pierson, D.L.; Smith, S.M. Response to vitamin D supplementation during Antarctic winter is related to BMI, and supplementation can mitigate Epstein-Barr virus reactivation. J. Nutr. 2011, 141, 692–697. [Google Scholar] [CrossRef]
- Smith, S.M.; Gardner, K.K.; Locke, J.; Zwart, S.R. Vitamin D supplementation during Antarctic winter. Am. J. Clin. Nutr. 2009, 89, 1092–1098. [Google Scholar]
- Vermeer, C.; Wolf, J.; Craciun, A.M.; Knapen, M.H. Bone markers during a 6-month space flight: Effects of vitamin K supplementation. J. Gravit. Physiol. 1998, 5, 65–69. [Google Scholar]
- Vermeer, C.; Wolf, J.; Knapen, M.H. Microgravity-induced changes of bone markers: Effects of vitamin K-supplementation. Bone 1997, 20, 16S. [Google Scholar]
- Zwart, S.R.; Booth, S.L.; Peterson, J.W.; Wang, Z.; Smith, S.M. Vitamin K status in spaceflight and ground-based models of spaceflight. J. Bone Miner. Res. 2011, 26, 948–954. [Google Scholar] [CrossRef]
- Lockwood, D.R.; Vogel, J.M.; Schneider, V.S.; Hulley, S.B. Effect of the diphosphonate EHDP on bone mineral metabolism during prolonged bed rest. J. Clin. Endocrinol. Metab. 1975, 41, 533–541. [Google Scholar] [CrossRef]
- Rittweger, J.; Frost, H.M.; Schiessl, H.; Ohshima, H.; Alkner, B.; Tesch, P.; Felsenberg, D. Muscle atrophy and bone loss after 90 days’ bed rest and the effects of flywheel resistive exercise and pamidronate: Results from the LTBR study. Bone 2005, 36, 1019–1029. [Google Scholar]
- Schultheis, L.; Ruff, C.B.; Rastogi, S.; Bloomfield, S.; Hogan, H.A.; Fedarko, N.; Thierry-Palmer, M.; Ruiz, J.; Bauss, F.; Shapiro, J.R. Disuse bone loss in hindquarter suspended rats: Partial weightbearing, exercise and ibandronate treatment as countermeasures. J. Gravit. Physiol. 2000, 7, P13–P14. [Google Scholar]
- Shapiro, J.; Smith, B.; Beck, T.; Ballard, P.; Dapthary, M.; Brintzenhofeszoc, K.; Caminis, J. Treatment with zoledronic acid ameliorates negative geometric changes in the proximal femur following acute spinal cord injury. Calcif. Tissue Int. 2007, 80, 316–322. [Google Scholar] [CrossRef]
- LeBlanc, A.; Matsumoto, T.; Jones, J.; Shapiro, J.; Lang, T.; Shackelford, L.; Smith, S.M.; Evans, H.; Spector, E.; Ploutz-Snyder, L.; et al. Bisphosphonates as a supplement to exercise to protect bone during long-duration spaceflight. Osteoporos. Int. 2012, in press.. [Google Scholar]
- Strollo, F.; Strollo, G.; More, M.; Bollanti, L.; Ciarmatori, A.; Longo, E.; Quintiliani, R.; Mambro, A.; Mangrossa, N.; Ferretti, C. Hormonal adaptation to real and simulated microgravity. J. Gravit. Physiol. 1998, 5, P89–P92. [Google Scholar]
- Smith, S.M.; Heer, M.; Wang, Z.; Huntoon, C.L.; Zwart, S.R. Long-duration space flight and bed rest effects on testosterone and other steroids. J. Clin. Endocrinol. Metab. 2012, 97, 270–278. [Google Scholar] [CrossRef]
- Whitson, P.A.; Pietrzyk, R.A.; Morukov, B.V.; Sams, C.F. The risk of renal stone formation during and after long duration space flight. Nephron 2001, 89, 264–270. [Google Scholar]
- Whitson, P.A.; Pietrzyk, R.A.; Pak, C.Y. Renal stone risk assessment during Space Shuttle flights. J. Urol. 1997, 158, 2305–2310. [Google Scholar] [CrossRef]
- Okada, A.; Ichikawa, J.; Tozawa, K. Kidney stone formation during space flight and long-term bed rest. Clin. Calcium 2011, 21, 1505–1510. [Google Scholar]
- Hwang, T.I.; Hill, K.; Schneider, V.; Pak, C.Y. Effect of prolonged bedrest on the propensity for renal stone formation. J. Clin. Endocrinol. Metab. 1988, 66, 109–112. [Google Scholar] [CrossRef]
- Monga, M.; Macias, B.; Groppo, E.; Kostelec, M.; Hargens, A. Renal stone risk in a simulated microgravity environment: Impact of treadmill exercise with lower body negative pressure. J. Urol. 2006, 176, 127–131. [Google Scholar]
- Okada, A.; Ohshima, H.; Itoh, Y.; Yasui, T.; Tozawa, K.; Kohri, K. Risk of renal stone formation induced by long-term bed rest could be decreased by premedication with bisphosphonate and increased by resistive exercise. Int. J. Urol. 2008, 15, 630–635. [Google Scholar] [CrossRef]
- Watanabe, Y.; Ohshima, H.; Mizuno, K.; Sekiguchi, C.; Fukunaga, M.; Kohri, K.; Rittweger, J.; Felsenberg, D.; Matsumoto, T.; Nakamura, T. Intravenous pamidronate prevents femoral bone loss and renal stone formation during 90-day bed rest. J. Bone Miner. Res. 2004, 19, 1771–1778. [Google Scholar] [CrossRef]
- Zerwekh, J.E.; Odvina, C.V.; Wuermser, L.A.; Pak, C.Y. Reduction of renal stone risk by potassium-magnesium citrate during 5 weeks of bed rest. J. Urol. 2007, 177, 2179–2184. [Google Scholar]
- Pak, C.Y.; Sakhaee, K.; Crowther, C.; Brinkley, L. Evidence justifying a high fluid intake in treatment of nephrolithiasis. Ann. Intern. Med. 1980, 93, 36–39. [Google Scholar]
- Whitson, P.; Pietrzyk, R.; Pak, C.; Cintron, N. Alterations in renal stone risk factors after space flight. J. Urol. 1993, 150, 803–807. [Google Scholar]
- Whitson, P.A.; Pietrzyk, R.A.; Sams, C.F. Urine volume and its effects on renal stone risk in astronauts. Aviat. Space Environ. Med. 2001, 72, 368–372. [Google Scholar]
- Whitson, P.A.; Pietrzyk, R.A.; Jones, J.A.; Nelman-Gonzalez, M.; Hudson, E.K.; Sams, C.F. Effect of potassium citrate therapy on the risk of renal stone formation during spaceflight. J. Urol. 2009, 182, 2490–2496. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Smith, S.M.; McCoy, T.; Gazda, D.; Morgan, J.L.L.; Heer, M.; Zwart, S.R. Space Flight Calcium: Implications for Astronaut Health, Spacecraft Operations, and Earth. Nutrients 2012, 4, 2047-2068. https://doi.org/10.3390/nu4122047
Smith SM, McCoy T, Gazda D, Morgan JLL, Heer M, Zwart SR. Space Flight Calcium: Implications for Astronaut Health, Spacecraft Operations, and Earth. Nutrients. 2012; 4(12):2047-2068. https://doi.org/10.3390/nu4122047
Chicago/Turabian StyleSmith, Scott M., Torin McCoy, Daniel Gazda, Jennifer L. L. Morgan, Martina Heer, and Sara R. Zwart. 2012. "Space Flight Calcium: Implications for Astronaut Health, Spacecraft Operations, and Earth" Nutrients 4, no. 12: 2047-2068. https://doi.org/10.3390/nu4122047
APA StyleSmith, S. M., McCoy, T., Gazda, D., Morgan, J. L. L., Heer, M., & Zwart, S. R. (2012). Space Flight Calcium: Implications for Astronaut Health, Spacecraft Operations, and Earth. Nutrients, 4(12), 2047-2068. https://doi.org/10.3390/nu4122047