Differential Mammary Gland Development in FVB and C57Bl/6 Mice: Implications for Breast Cancer Research


Abstract
:1. Introduction
2. Material and Methods
3. Results
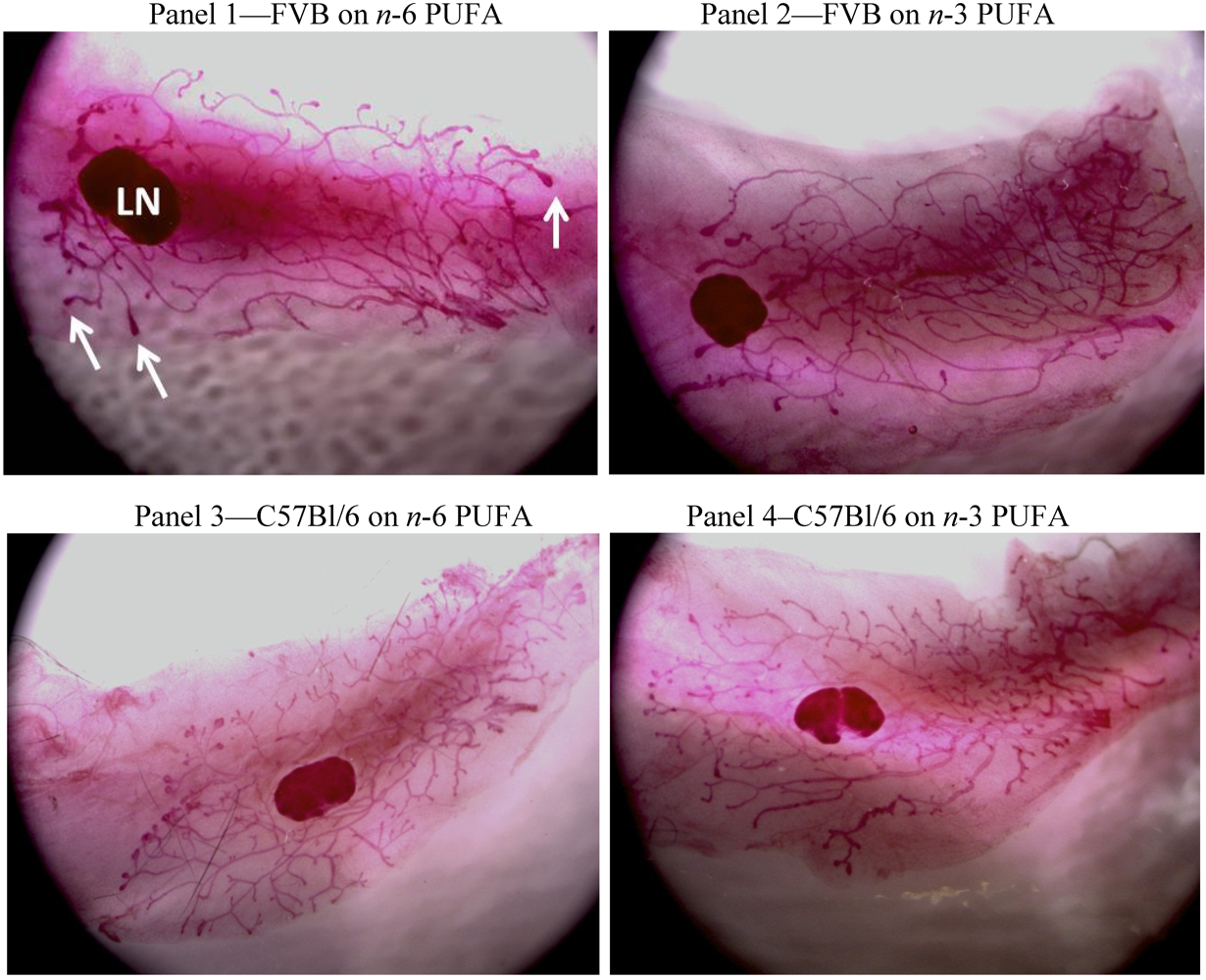
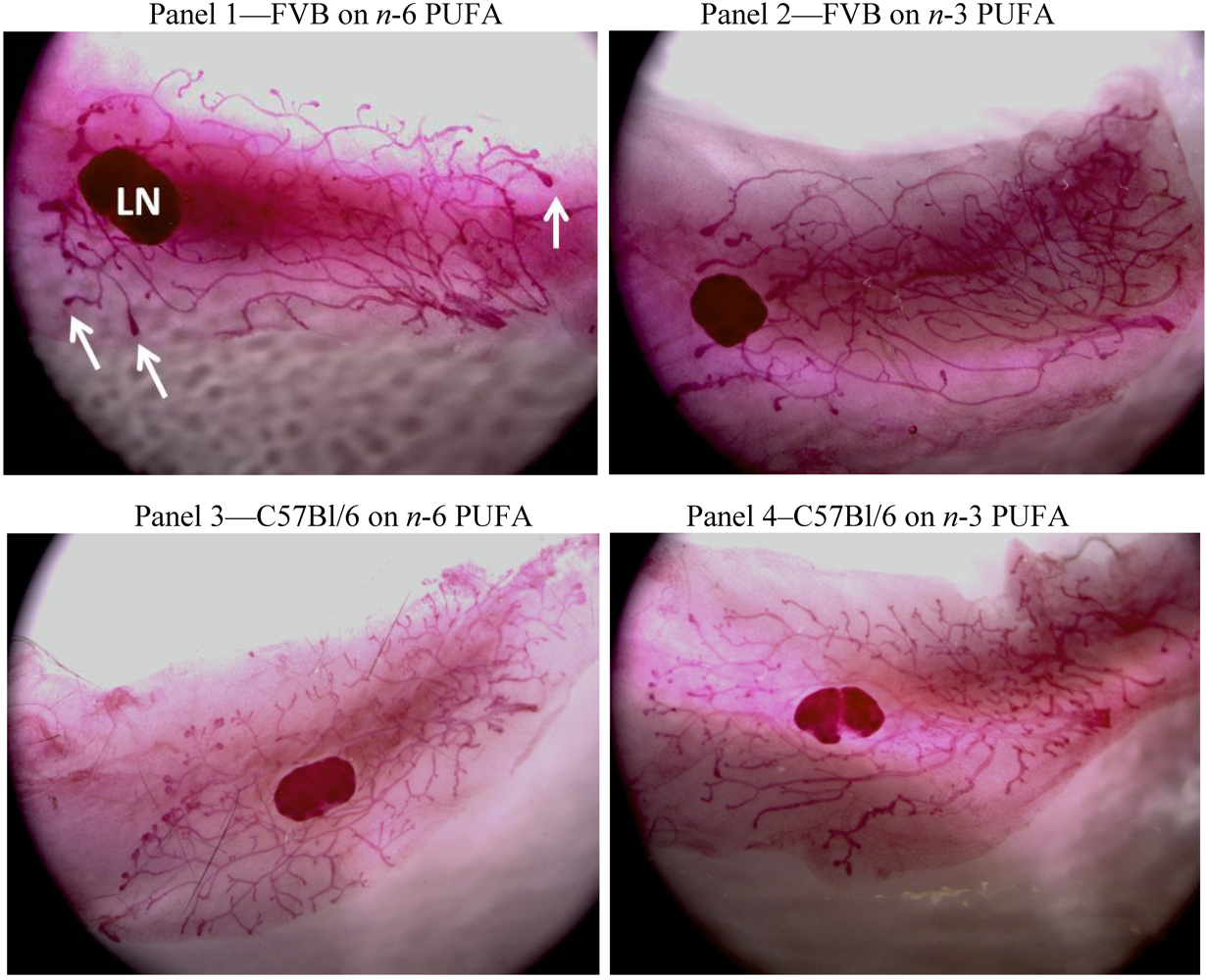
3.1. FVB Mice

{kind=link}
{kind=link}
| n-6 PUFA | n-3 PUFA | ||
|---|---|---|---|
| 6 weeks | FVB | 18.7 ± 4.0 (15) | * 14.9 ± 3.3 (13) |
| C57Bl/6 | 19.9 ± 8.5 (7) | * 11.3 ± 8.7 (18) |
3.2. C57Bl/6 Mice
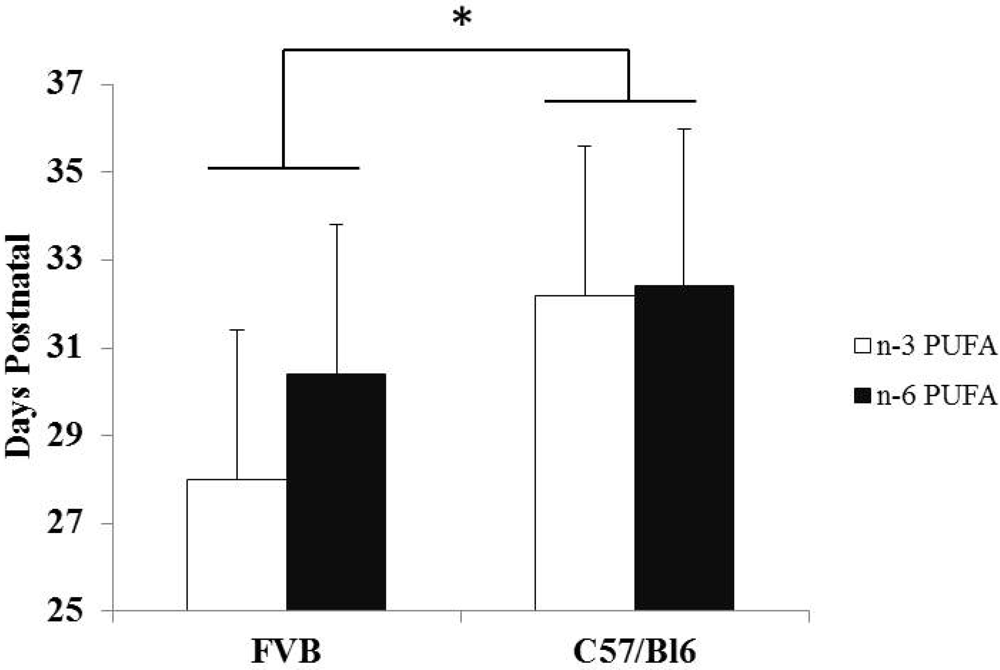
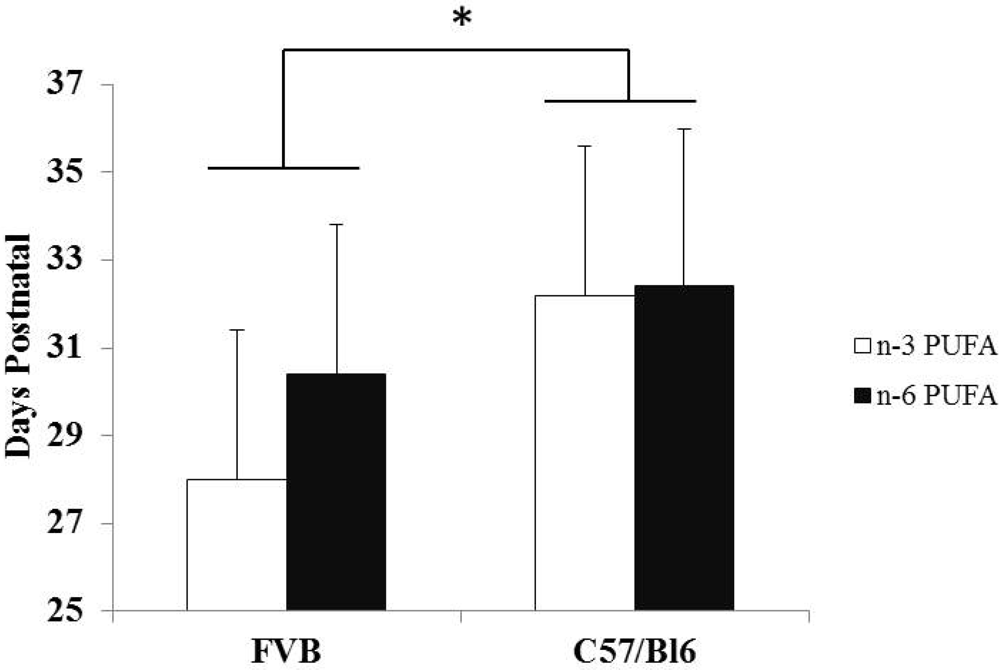
3.3. Strain Differences between FVB and C57Bl/6 Mice
4. Discussion
5. Implications
Acknowledgements
Conflict of Interest
References
- Haluzik, M.; Colombo, C.; Gavrilova, O.; Chua, S.; Wolf, N.; Chen, M.; Stannard, B.; Dietz, K.R.; Le Roith, D.; Reitman, M.L. Genetic background (C57BL/6J versus FVB/N) strongly influences the severity of diabetes and insulin resistance in ob/ob mice. Endocrinology 2004, 145, 3258–3264. [Google Scholar] [PubMed]
- Aupperlee, M.D.; Drolet, A.A.; Durairaj, S.; Wang, W.; Schwartz, R.C.; Haslam, S.Z. Strain-specific differences in the mechanisms of progesterone regulation of murine mammary gland development. Endocrinology 2009, 150, 1485–1494. [Google Scholar]
- Yang, C.; Tan, Y.S.; Harkema, J.R.; Haslam, S.Z. Differential effects of peripubertal exposure to perfluorooctanoic acid on mammary gland development in C57Bl/6 and Balb/c mouse strains. Reprod. Toxicol. 2009, 27, 299–306. [Google Scholar]
- Olson, L.K.; Tan, Y.; Zhao, Y.; Aupperlee, M.D.; Haslam, S.Z. Pubertal exposure to high fat diet causes mouse strain-dependent alterations in mammary gland development and estrogen responsiveness. Int. J. Obes. (Lond.) 2010, 34, 1415–1426. [Google Scholar] [CrossRef] [PubMed]
- Russo, J.; Gusterson, B.A.; Rogers, A.E.; Russo, I.H.; Wellings, S.R.; van Zwieten, M.J. Comparative study of human and rat mammary tumorigenesis. Lab. Invest. 1990, 62, 244–278. [Google Scholar]
- Russo, J.; Russo, I.H. Molecular Basis of Breast Cancer: Prevention and Treatment; Springer: Berlin, Germany, 2004. [Google Scholar]
- MacLennan, M.; Ma, D.W. Role of dietary fatty acids in mammary gland development and breast cancer. Breast Cancer Res. 2010, 12, 211. [Google Scholar]
- Ziegler, R.G.; Hoover, H.N.; Pike, M.C.; Hildesheim, A.; Nomura, A.M.; West, D.W.; Wu-Williams, A.H.; Kolonel, L.N.; Horn-Ross, P.L.; Rosenthal, J.F.; et al. Migration patterns and breast cancer risk in Asian-American women. J. Natl. Cancer Inst. 1993, 85, 1819–1827. [Google Scholar] [CrossRef] [PubMed]
- Hilakivi-Clarke, L.; Clarke, R.; Onojafe, I.; Raygada, M.; Cho, E.; Lippman, M. A maternal diet high in n-6 polyunsaturated fats alters mammary gland development, puberty onset, and breast cancer risk among female rat offspring. Proc. Natl. Acad. Sci. USA 1997, 94, 9372–9377. [Google Scholar]
- Hilakivi-Clarke, L.; Olivo, S.E.; Shajahan, A.; Khan, G.; Zhu, Y.; Zwart, A.; Cho, E.; Clarke, R. Mechanisms mediating the effects of prepubertal (n-3) polyunsaturated fatty acid diet on breast cancer risk in rats. J. Nutr. 2005, 135, 2946–2952. [Google Scholar] [PubMed]
- Davie, S.A.; Maglione, J.E.; Manner, C.K.; Young, D.; Cardiff, R.D.; MacLeod, C.L.; Ellies, L.G. Effects of FVB/NJ and C57Bl/6J strain backgrounds on mammary tumor phenotype in inducible nitric oxide synthase deficient mice. Transgenic Res. 2007, 16, 193–201. [Google Scholar]
- Kuperwasser, C.; Hurlbut, G.D.; Kittrell, F.S.; Dickinson, E.S.; Laucirica, R.; Medina, D.; Naber, S.P.; Jerry, D.J. Development of spontaneous mammary tumors in BALB/c p53 heterozygous mice: a model for Li-Fraumeni syndrome. Am. J. Pathol. 2000, 157, 2151–2159. [Google Scholar]
- Tou, J.C.; Thompson, L.U. Exposure to flaxseed or its lignan component during different developmental stages influences rat mammary gland structures. Carcinogenesis 1999, 20, 1831–1835. [Google Scholar]
- Fata, J.E.; Chaudhary, V.; Khokha, R. Cellular turnover in the mammary gland is correlated with systemic levels of progesterone and not 17β-estradiol during the estrous cycle. Biol. Reprod. 2001, 65, 680–688. [Google Scholar]
- Richert, M.M.; Schwertfeger, K.L.; Ryder, J.W.; Anderson, S.M. An atlas of mouse mammary gland development. J. Mammary Gland. Biol. Neoplasia 2000, 5, 227–241. [Google Scholar]
- Wechselberger, C.; Strizzi, L.; Kenney, N.; Hirota, M.; Sun, Y.; Ebert, A.; Orozco, O.; Bianco, C.; Khan, N.I.; Wallace-Jones, B.; et al. Human Cripto-1 overexpression in the mouse mammary gland results in the development of hyperplasia and adenocarcinoma. Oncogene 2005, 24, 4094–4105. [Google Scholar] [PubMed]
- Clavel-Chapelon, F. Differential effects of reproductive factors on the risk of pre- and postmenopausal breast cancer. Results from a large cohort of French women. Br. J. Cancer 2002, 86, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.W.; Mak, J.; Cheung, P.S.; Epstein, R.J. Evidence for a programming effect of early menarche on the rise of breast cancer incidence in Hong Kong. Cancer Detect. Prev. 2008, 32, 156–161. [Google Scholar]
- Velie, E.M.; Nechuta, S.; Osuch, J.R. Lifetime reproductive and anthropometric risk factors for breast cancer in postmenopausal women. Breast Dis. 2005, 24, 17–35. [Google Scholar]
- Kelsey, J.L.; Gammon, M.D.; John, E.M. Reproductive factors and breast cancer. Epidemiol. Rev. 1993, 15, 36–47. [Google Scholar]
- Michels, K.B.; Trichopoulos, D.; Robins, J.M.; Rosner, B.A.; Manson, J.E.; Hunter, D.J.; Colditz, G.A.; Hankinson, S.E.; Speizer, F.E.; Willett, W.C. Birthweight as a risk factor for breast cancer. Lancet 1996, 348, 1542–1546. [Google Scholar]
- van den Brandt, P.A.; Spiegelman, D.; Yaun, S.S.; Adami, H.O.; Beeson, L.; Folsom, A.R.; Fraser, G.; Goldbohm, R.A.; Graham, S.; Kushi, L.; et al. Pooled analysis of prospective cohort studies on height, weight, and breast cancer risk. Am. J. Epidemiol. 2000, 152, 514–527. [Google Scholar] [CrossRef] [PubMed]
- Gago-Dominguez, M.; Yuan, J.M.; Sun, C.L.; Lee, H.P.; Yu, M.C. Opposing effects of dietary n-3 and n-6 fatty acids on mammary carcinogenesis: The Singapore Chinese Health Study. Br. J. Cancer 2003, 89, 1686–1692. [Google Scholar] [CrossRef] [PubMed]
- Begin, M.E.; Ells, G.; Horrobin, D.F. Polyunsaturated fatty acid-induced cytotoxicity against tumor cells and its relationship to lipid peroxidation. J. Natl. Cancer Inst. 1988, 80, 188–194. [Google Scholar]
- Gonzalez, M.J.; Schemmel, R.A.; Dugan, L., Jr.; Gray, J.I.; Welsch, C.W. Dietary fish oil inhibits human breast carcinoma growth: a function of increased lipid peroxidation. Lipids 1993, 28, 827–832. [Google Scholar]
- Hardman, W.E.; Munoz, J., Jr.; Cameron, I.L. Role of lipid peroxidation and antioxidant enzymes in omega 3 fatty acids induced suppression of breast cancer xenograft growth in mice. Cancer Cell Int. 2002, 2, 10. [Google Scholar]
- Olivo, S.E.; Hilakivi-Clarke, L. Opposing effects of prepubertal low- and high-fat n-3 polyunsaturated fatty acid diets on rat mammary tumorigenesis. Carcinogenesis 2005, 26, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MacLennan, M.B.; Anderson, B.M.; Ma, D.W.L. Differential Mammary Gland Development in FVB and C57Bl/6 Mice: Implications for Breast Cancer Research. Nutrients 2011, 3, 929-936. https://doi.org/10.3390/nu3110929
MacLennan MB, Anderson BM, Ma DWL. Differential Mammary Gland Development in FVB and C57Bl/6 Mice: Implications for Breast Cancer Research. Nutrients. 2011; 3(11):929-936. https://doi.org/10.3390/nu3110929
Chicago/Turabian StyleMacLennan, Mira B., Breanne M. Anderson, and David W.L. Ma. 2011. "Differential Mammary Gland Development in FVB and C57Bl/6 Mice: Implications for Breast Cancer Research" Nutrients 3, no. 11: 929-936. https://doi.org/10.3390/nu3110929