Chemistry and Biochemistry of Dietary Polyphenols

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Classification of Polyphenols
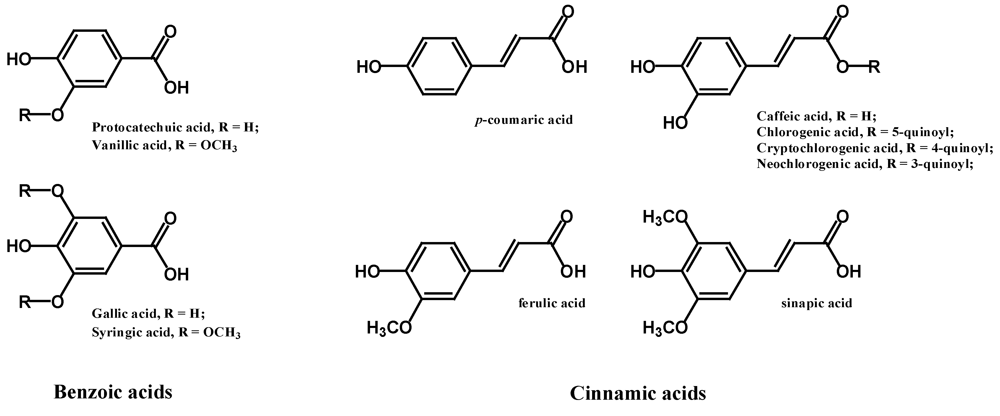
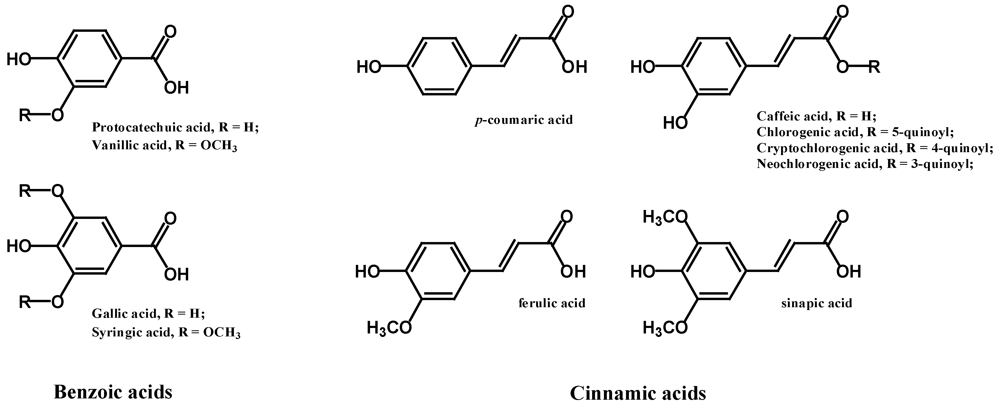
2.1. Phenolic Acids
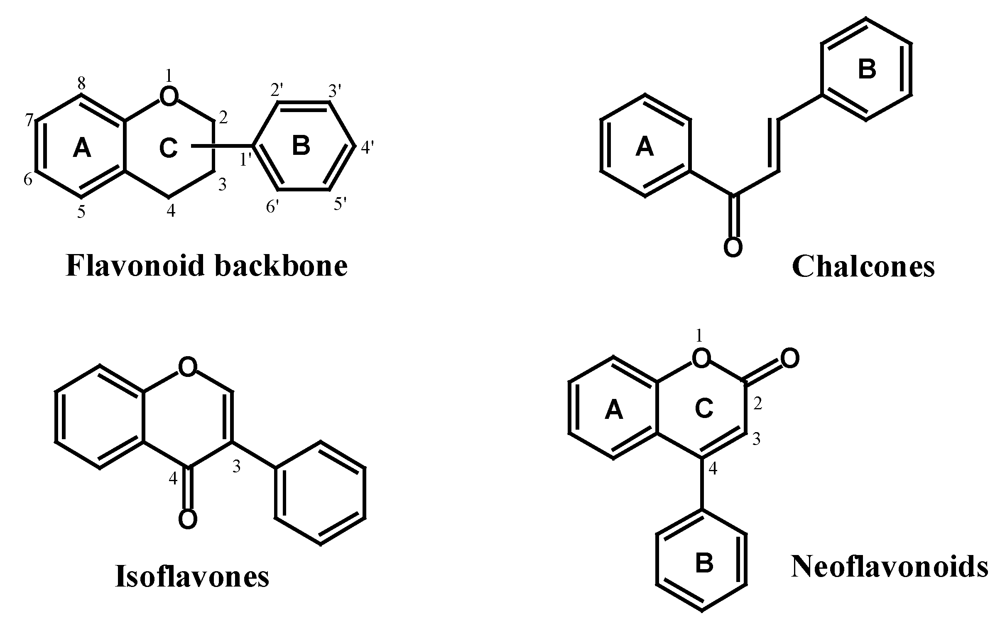
2.2. Flavonoids
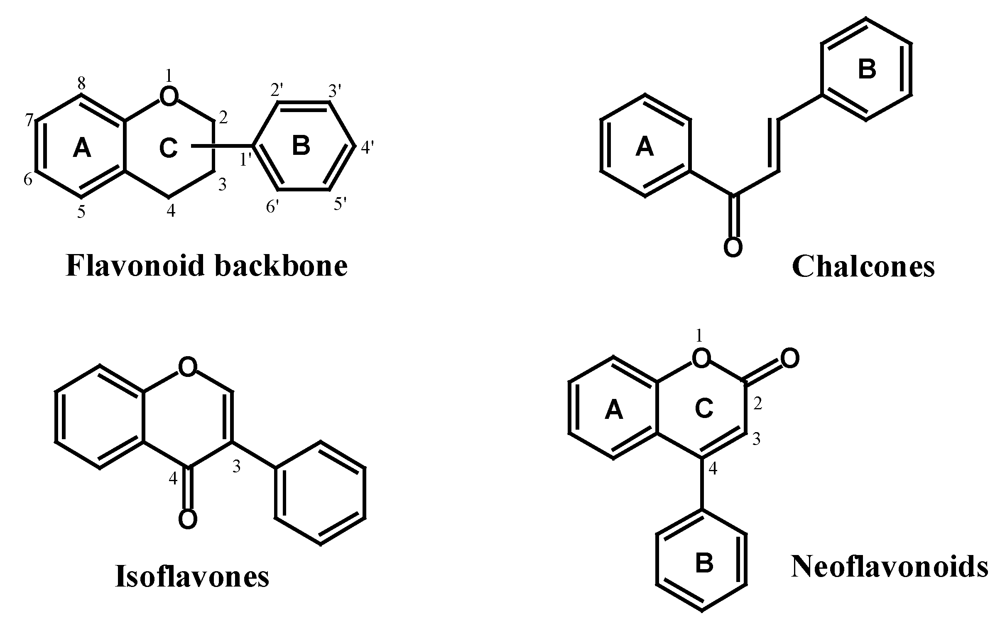
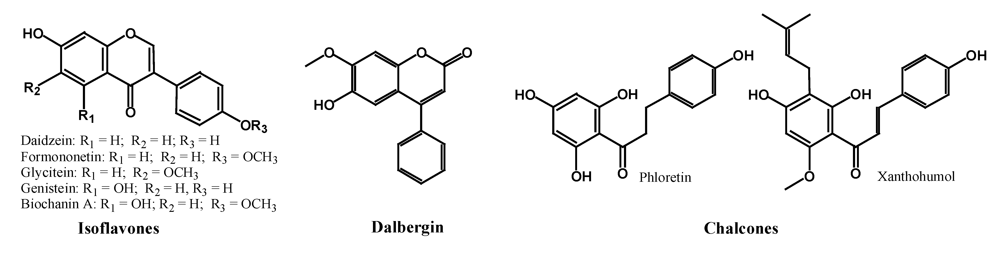
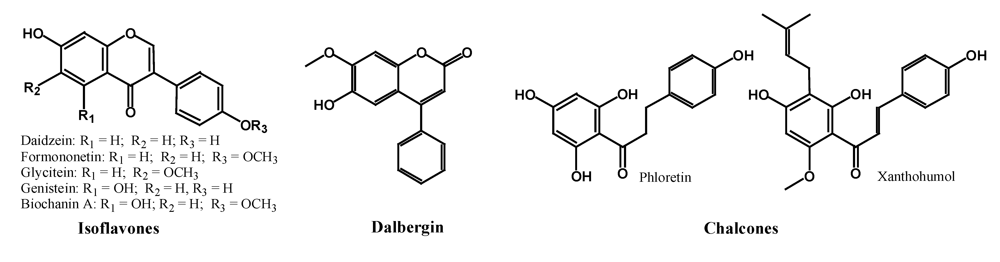
2.2.1. Isoflavones, Neoflavonoids and Chalcones
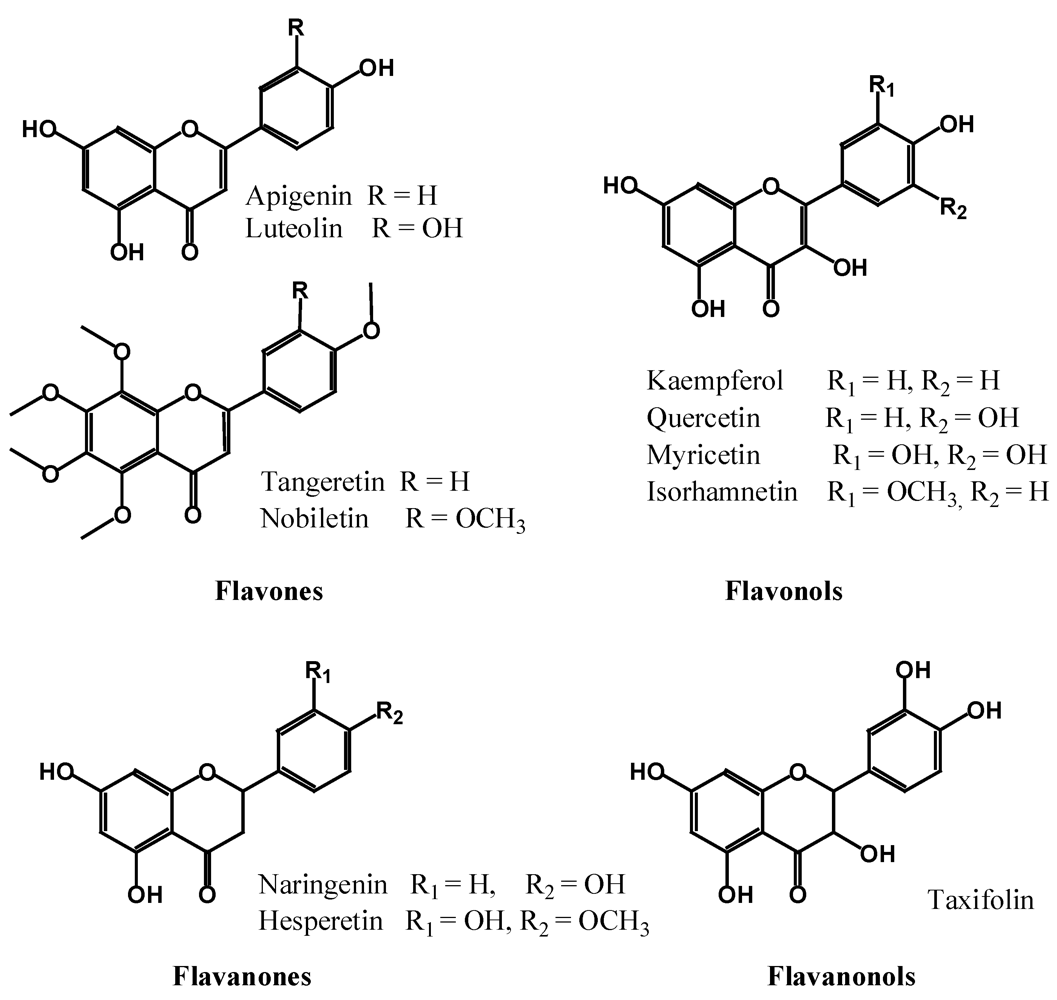
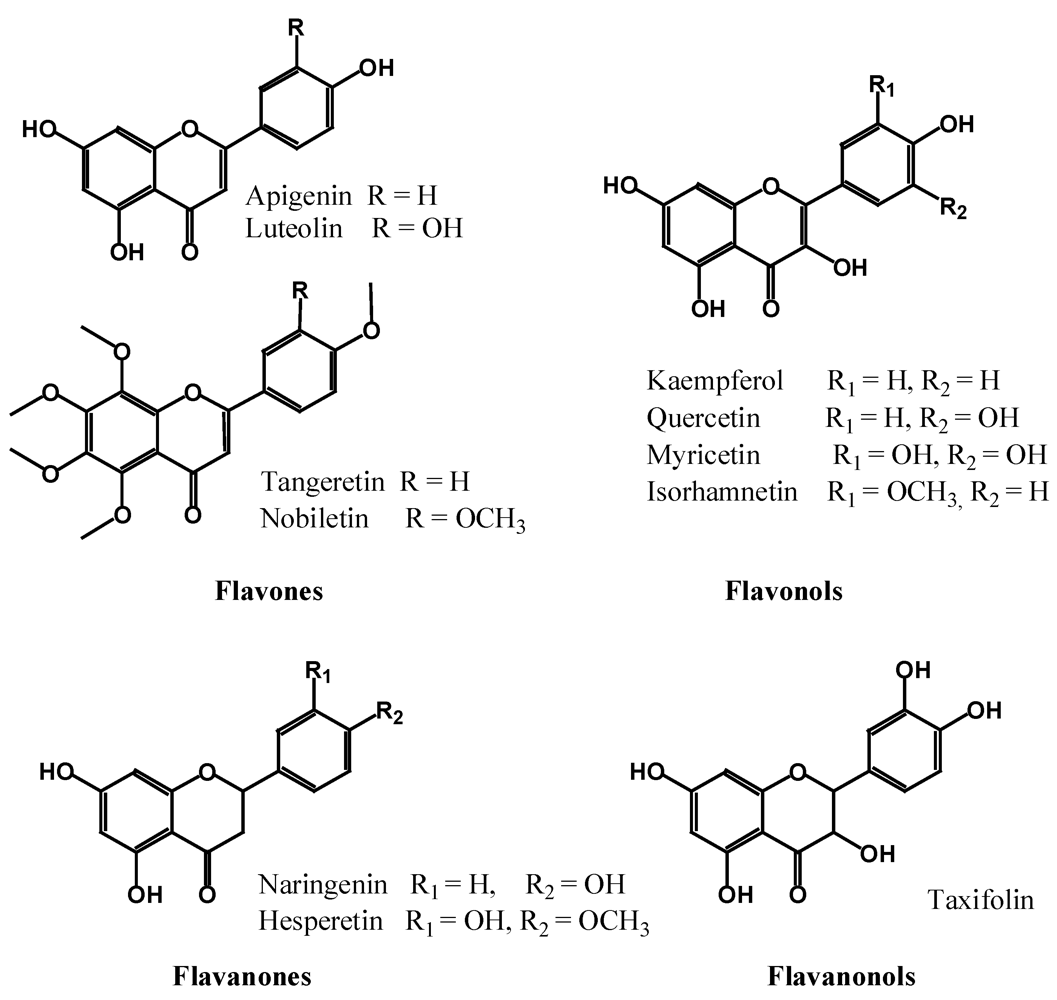
2.2.2. Flavones, Flavonols, Flavanones and Flavanonols
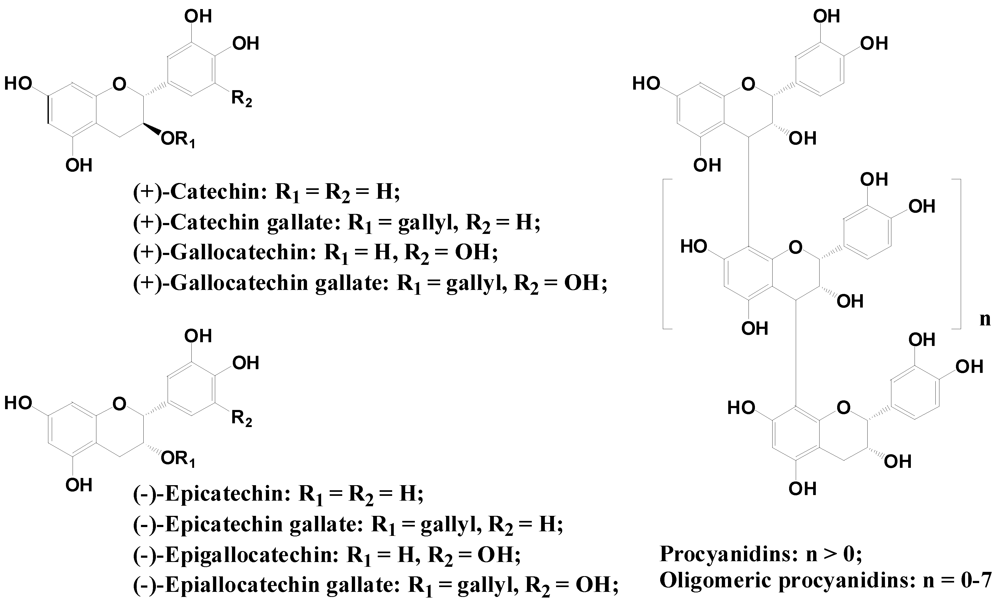
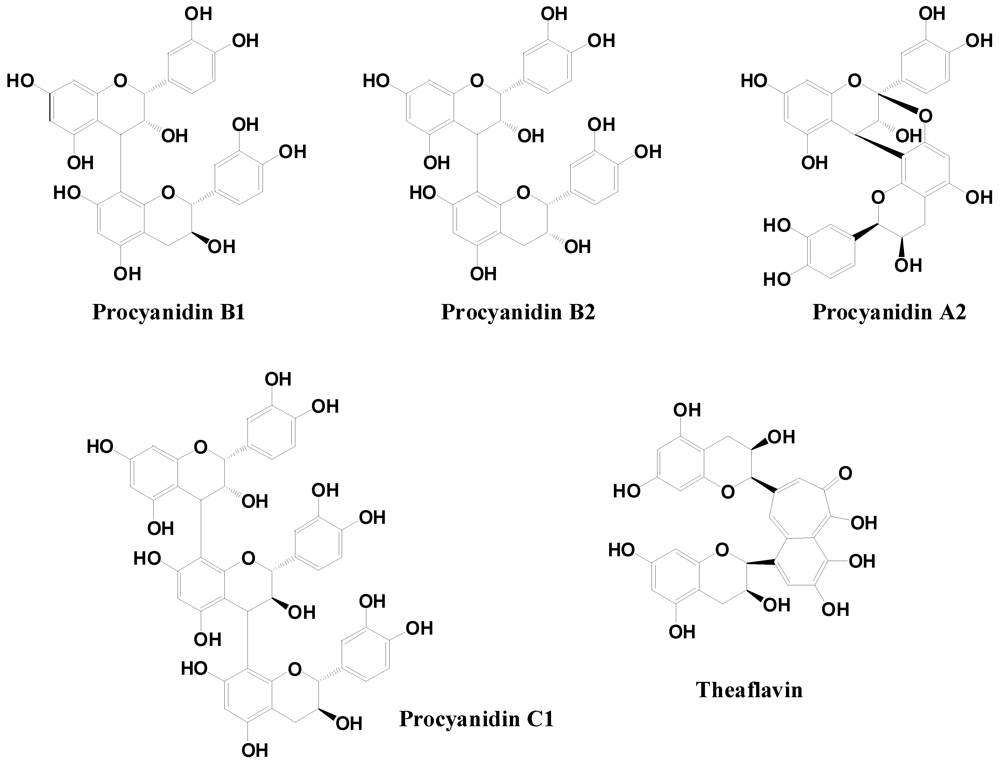
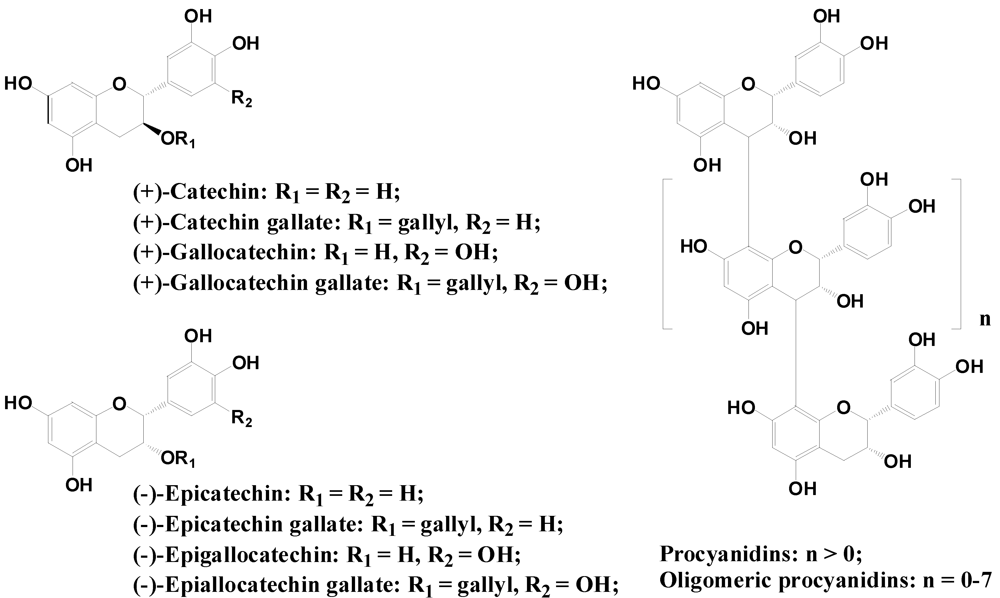
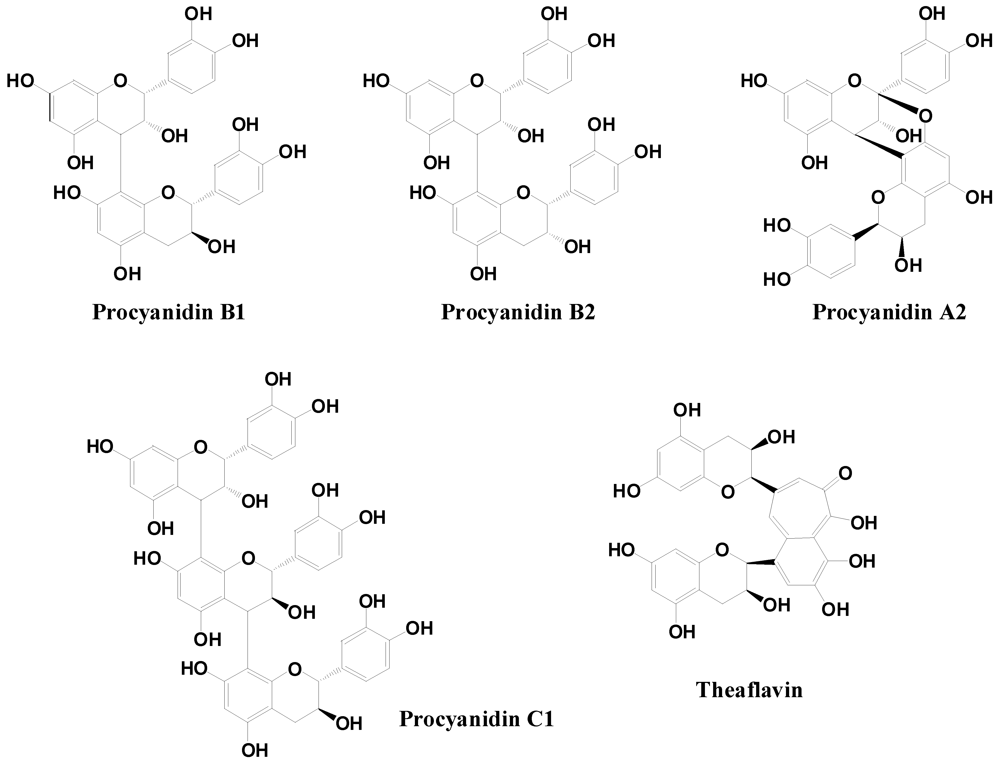
2.2.3. Flavanols and Proanthocyanidins
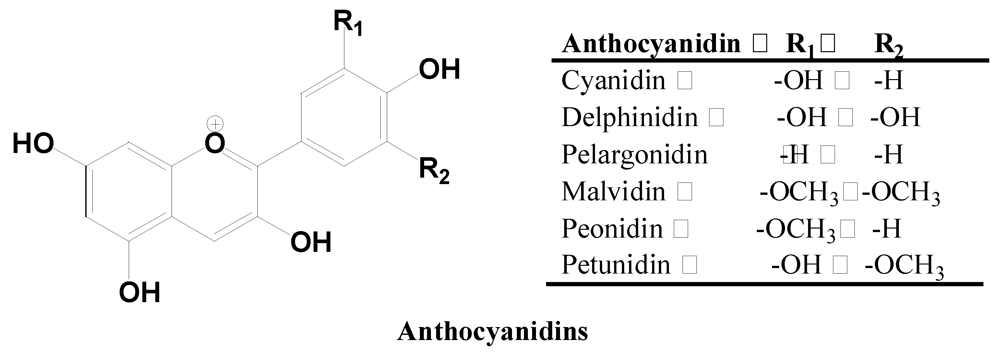
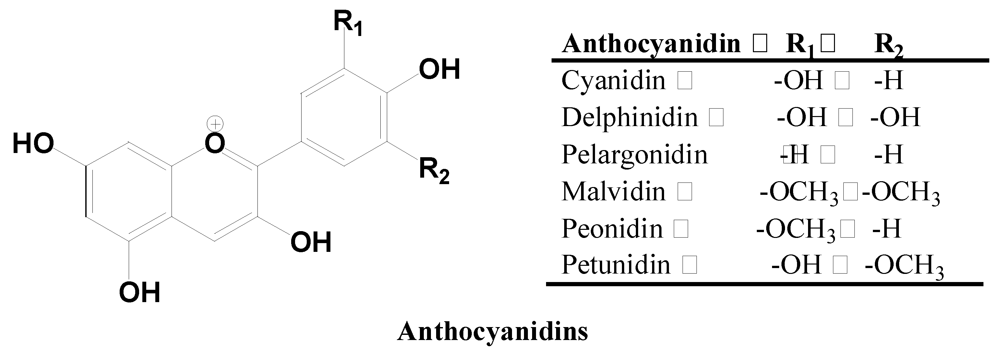
2.2.4. Anthocyanidins
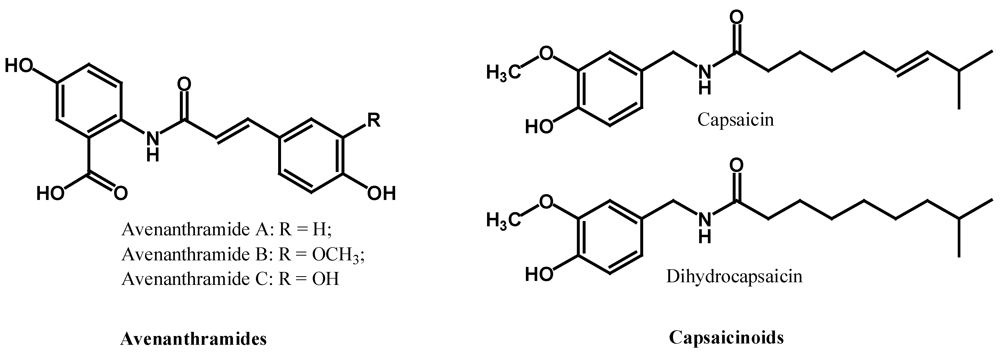
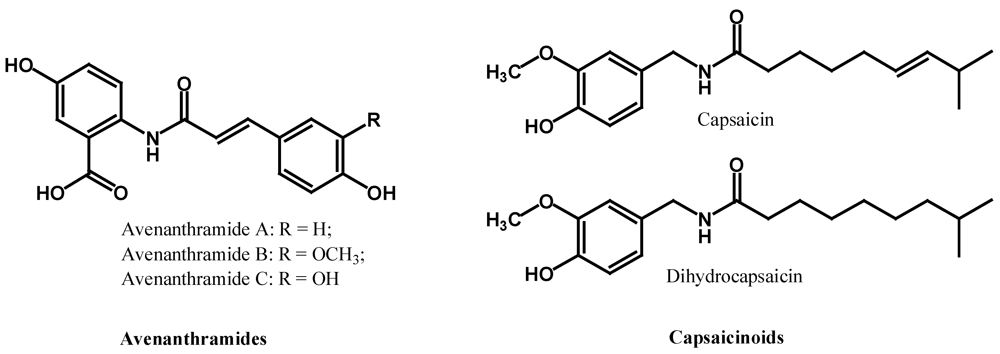
2.3. Polyphenolic Amides
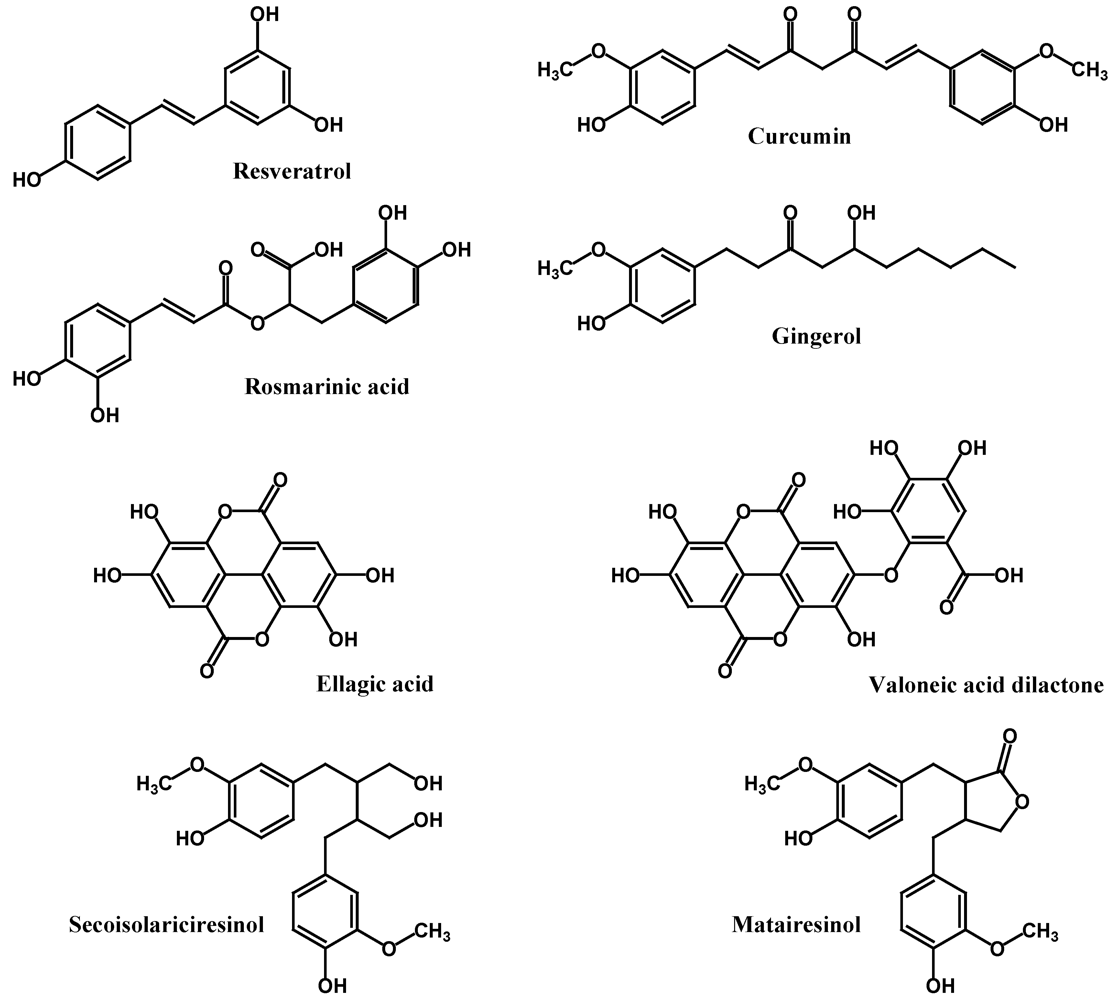
2.4. Other Polyphenols
3. Biosynthesis of Polyphenols
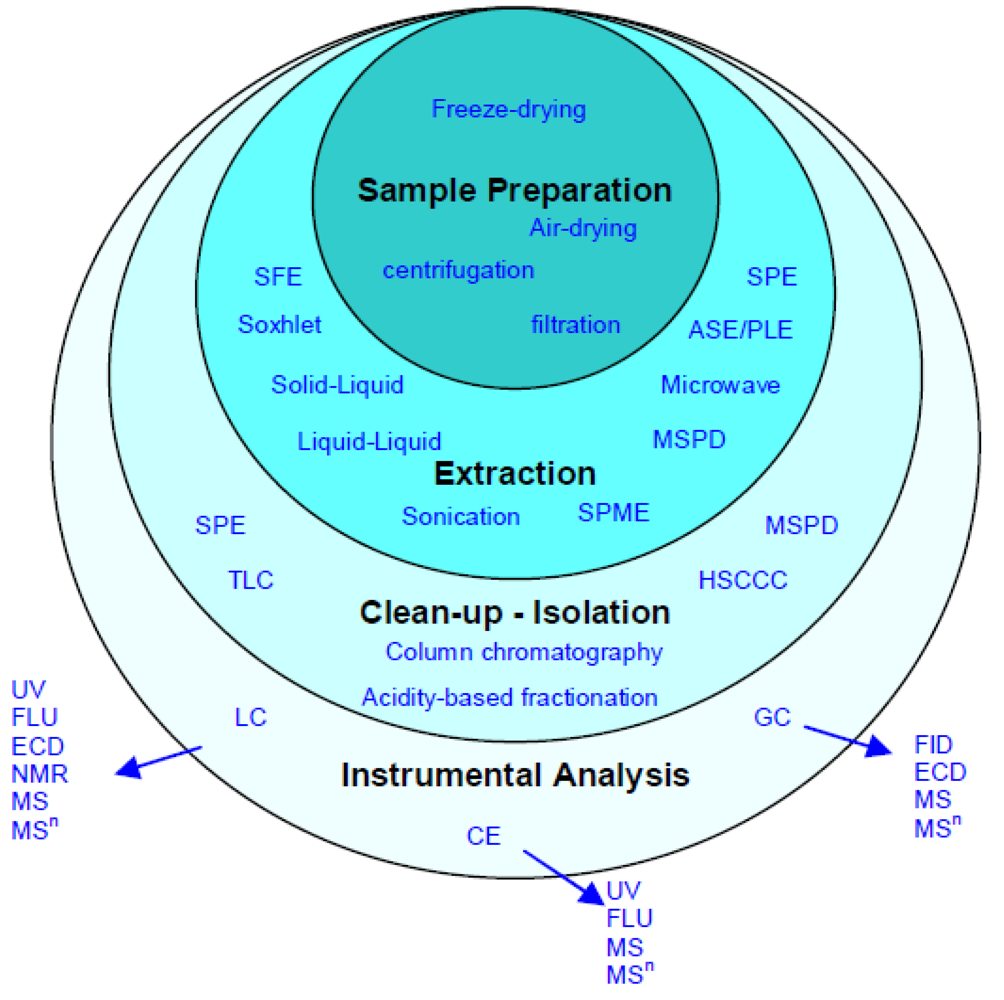
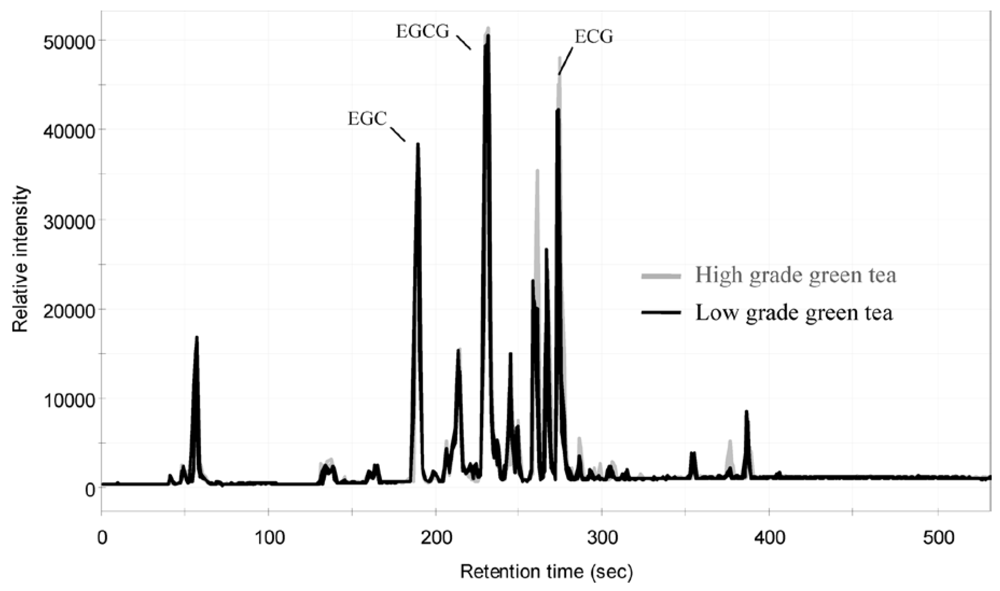
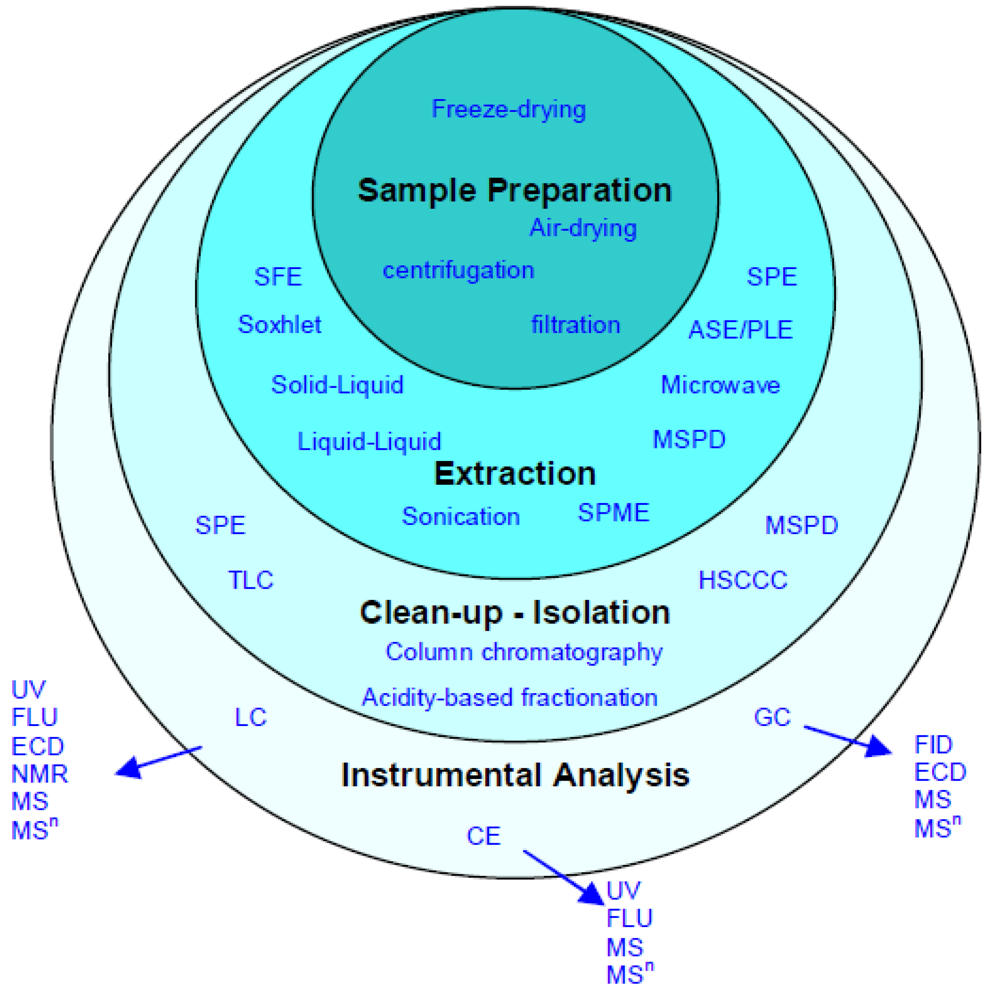
4. Separation, Identification and Analysis of Polyphenols
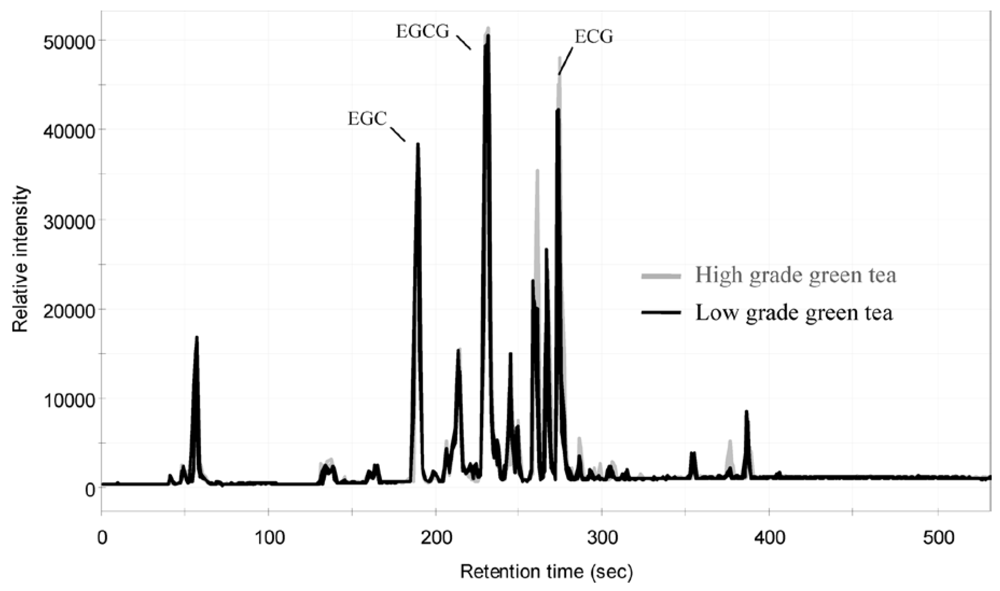
5. Biological Roles and Implications (Mainly on Antioxidant Activities)
5.1. Antioxidant Activity
5.2. Beyond the Usual Antioxidant Activity
References
- Milner, J.A. Reducing the risk of cancer. In Functional Foods: Designer Foods, Pharmafoods, Nutraceuticals; Goldberg, I., Ed.; Chapman & Hall: New York, NY, USA, 1994; pp. 39–70. [Google Scholar]
- Duthie, G.G.; Brown, K.M. Reducing the risk of cardiovascular disease. In Functional Foods: Designer Foods, Pharmafoods, Nutraceuticals; Goldberg, I., Ed.; Chapman & Hall: New York, NY, USA, 1994; pp. 19–38. [Google Scholar]
- Williams, R.J.; Spencer, J.P.; Rice-Evans, C. Flavonoids: Antioxidants or signaling molecules? Free Radic. Biol. Med. 2004, 36, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Surh, Y.-J.; Packer, L.; Cadenas, E. Dietary Modulation of Cell Signaling Pathways; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2009. [Google Scholar]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar]
- Bravo, L. Polyphenols: Chemistry, Dietary Sources, Metabolism, and Nutritional Significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Cheynier, V. Polyphenols in foods are more complex than often thought. Am. J. Clin. Nutr. 2005, 81, 223S–229S. [Google Scholar]
- Kim, K.-H.; Tsao, R.; Yang, R.; Cui, S.W. Phenolic acid profiles and antioxidant activities of wheat bran extracts and the effect of hydrolysis conditions. Food Chem. 2006, 95, 466–473. [Google Scholar] [CrossRef]
- Adom, K.K.; Liu, R.H. Antioxidant activity of grains. J. Agric. Food Chem. 2002, 50, 6182–6187. [Google Scholar]
- Chandrasekara, A.; Shahidi, F. Content of insoluble bound phenolics in millets and their contribution to antioxidant capacity. J. Agric. Food Chem. 2010, 58, 6706–6714. [Google Scholar]
- Wang, H.; Murphy, P.A. Isoflavone content in commercial soybean foods. J. Agric. Food Chem. 1994, 42, 1666–1673. [Google Scholar]
- Mazur, W.M.; Duke, J.A.; Wahala, K.; Rasku, S.; Adlercreutz, H. Isoflavonoids and lignans in legumes: Nutritional and health aspects in Humans. J. Nutr. Biochem. 1998, 9, 193–200. [Google Scholar]
- Tsao, R.; Papadopoulos, Y.; Yang, R.; Young, J.C.; McRae, K. Isoflavone profiles of red clovers and their distribution in different parts harvested at different growing stages. J. Agric. Food Chem. 2006, 54, 5797–5805. [Google Scholar]
- Garazd, M.M.; Garazd, Y.L.; Khilya, V.P. Neoflavones. 1. Natural distribution and spectral and biological properties. Chem. Nat. Comp. 2003, 39, 54–121. [Google Scholar] [CrossRef]
- Tsao, R.; Yang, R.; Young, J.C.; Zhu, H. Polyphenolic profiles in eight apple cultivars using high–performance liquid chromatography (HPLC). J. Agric. Food Chem. 2003, 51, 6347–6353. [Google Scholar]
- Zhao, F.; Watanabe, Y.; Nozawa, H.; Daikonnya, A.; Kondo, K.; Kitanaka, S. Prenylflavonoids and phloroglucinol derivatives from Hops (Humulus lupulus). J. Nat. Prod. 2005, 68, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R.; McCallum, J. Chemistry of Flavonoids. In Fruit and Vegetable Phytochemicals: Chemistry, Nutritional Value and Stability; de la Rosa, L.A., Alvarez-Parrilla, E., Gonzalez–Aguilar, G., Eds.; Blackwell Publishing: Ames, IA, USA, 2009; pp. 131–153. Chapter 5. [Google Scholar]
- Valant-Vetschera, K.M.; Wallenweber, E. Flavones and Flavonols. In Flavonoids: Chemistry, Biochemistry and Applications; Anderson, O.M., Markham, K.R., Eds.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2006; pp. 618–748. [Google Scholar]
- Williams, C.A. Flavone and flavonol O-glycosides. In Flavonoids: Chemistry, Biochemistry and Applications; Anderson, O.M., Markham, K.R., Eds.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2006; pp. 749–856. [Google Scholar]
- Grayer, R.J.; Veitch, N.C. Flavanones and dihydroflavonols. In Flavonoids: Chemistry, Biochemistry and Applications; Anderson, O.M., Markham, K.R., Eds.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2006; pp. 918–1002. [Google Scholar]
- Kawaii, S.; Tomono, Y.; Katase, E.; Ogawa, K.; Yano, M. Quantitation of flavonoid constituents in citrus fruits. J. Agric. Food Chem. 1999, 47, 3565–3571. [Google Scholar]
- Si, W.; Gong, J.; Tsao, R.; Kalab, M.; Yang, R.; Yin, Y. Bioassay-guided purification and identification of antimicrobial components in Chinese green tea extract. J. Chromatogr. A 2006, 1125, 204–210. [Google Scholar]
- Prior, R.L.; Lazarus, S.A.; Cao, G.; Muccitelli, H.; Hammerstone, J.F. Identification of procyanidins and anthocyanins in blueberries and cranberries (Vaccinium spp.) using high–performance liquid chromatography/mass spectrometry. J. Agric. Food Chem. 2001, 49, 1270–1276. [Google Scholar]
- Anderson, O.M.; Jordheim, M. The anthocyanins. In Flavonoids: Chemistry, Biochemistry and Applications; Anderson, O.M., Markham, K.R., Eds.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2006; pp. 472–551. [Google Scholar]
- McCallum, J.L.; Yang, R.; Young, J.C.; Strommer, J.N.; Tsao, R. Improved high performance liquid chromatographic separation of anthocyanin compounds from grapes using a novel mixed–mode ion-exchange reversed-phase column. J. Chromatogr. A 2007, 1148, 38–45. [Google Scholar]
- Davis, C.B.; Markey, C.E.; Busch, M.A.; Busch, K.W. Determination of capsaicinoids in habanero peppers by chemometric analysis of UV spectral data. J. Agric. Food Chem. 2007, 55, 5925–5933. [Google Scholar]
- Bratt, K.; Sunnerheim, K.; Bryngelsson, S.; Fagerlund, A.; Engman, L.; Andersson, R.E.; Dimberg, L.H. Avenanthramides in oats (Avena sativa L.) and structure-antioxidant activity relationships. J. Agric. Food Chem. 2003, 51, 594–600. [Google Scholar] [PubMed]
- Knaggs, A.R. The biosynthesis of shikimate metabolites. Nat. Prod. Rep. 2001, 18, 334–355. [Google Scholar]
- Fatland, B.L.; Ke, J.; Anderson, M.D.; Mentzen, W.I.; Cui, L.W.; Allred, C.C.; Johnston, J.L.; Nikolau, B.J.; Wurtele, E.S. Molecular characterization of a heteromeric ATP-citrate lyase that generates cytosolic acetyl-Coenzyme A in Arabidopsis. Plant Physiol. 2004, 130, 740–756. [Google Scholar]
- Bohm, B.A. Introduction to Flavonoids; Harwood Academic Publishers: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Tsao, R.; Khanizadeh, S.; Dale, A. Designer fruits and vegetables with enriched phytochemicals for human health. Can. J. Plant Sci. 2006, 86, 773–786. [Google Scholar]
- Tsao, R.; Deng, Z. Separation Procedures for naturally occurring antioxidant phytochemicals. J. Chromatogr. B 2004, 812, 85–99. [Google Scholar]
- Stalikas, C.D. Extraction, separation, and detection methods for phenolic acids and flavonoids. J. Sep. Sci. 2007, 30, 3268–3295. [Google Scholar]
- Stalikas, C.D. Phenolic acids and flavonoids: occurrence and analytical methods. Methods Mol. Biol. 2010, 610, 65–90. [Google Scholar]
- Santos-Buelga, C.; Williamson, G. Method in Polyphenol Analysis; The Royal Society of Chemistry: Cambridge, UK, 2003. [Google Scholar]
- Tsao, R.; Yang, R.; Xie, S.; Sockovie, E.; Khanizadeh, S. Which polyphenolic compounds contribute to the total antioxidant activities of apple? J. Agric. Food Chem. 2005, 53, 4989–4995. [Google Scholar]
- Chun, O.K.; Kim, D.O.; Moon, H.Y.; Kang, H.G.; Lee, C.Y. Contribution of individual polyphenolics to total antioxidant capacity of plums. J. Agric. Food Chem. 2003, 51, 7240–7245. [Google Scholar]
- Pongsuwan, W.; Bamba, T.; Harada, K.; Yonetani, T.; Kobayashi, A.; Fukusaki, E. High–throughput technique for comprehensive analysis of Japanese green tea quality assessment using ultra–performance liquid chromatography with time-of-flight mass spectrometry (UPLC/TOF MS). J. Agric. Food Chem. 2008, 56, 10705–10708. [Google Scholar]
- Oleszek, W.; Stochmal, A.; Janda, B. Concentration of isoflavones and other phenolics in the aerial parts of Trifolium species. J. Agric. Food Chem. 2007, 55, 8095–8100. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R.; Yang, R. Optimisation of a new mobile phase to know the complex and real polyphenolic composition: Towards a total phenolic index using HPLC. J. Chromatogr. A 2003, 1018, 29–40. [Google Scholar]
- Li, L.; Tsao, R.; Yang, R.; Liu, C.; Zhu, H.; Young, J.C. Polyphenolic profiles and antioxidant activities of heartnut (Juglans ailanthifolia Var. cordiformis) and Persian walnut (Juglans regia L.). J. Agric. Food Chem. 2006, 54, 8033–8040. [Google Scholar] [PubMed]
- Wang, H.; Cao, G.; Prior, R.L. Total antioxidant capacity of fruits. J. Agric. Food Chem. 1996, 44, 701–705. [Google Scholar]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar]
- Guo, J.-J.; Hsieh, H.-Y.; Hu, C.-H. Chain-breaking activity of carotenes in lipid peroxidation: A theoretical study. J. Phys. Chem. B 2009, 113, 15699–15708. [Google Scholar]
- Perron, N.R.; Brumaghim, J.L. A review of the antioxidant mechanisms of polyphenol compounds related to iron binding. Cell Biochem. Biophys. 2009, 53, 75–100. [Google Scholar]
- Zhou, B.; Wu, L.-M.; Yang, L.; Liu, Z.-L. Evidence for alpha-tocopherol regeneration reaction of green tea polyphenols in SDS micelles. Free Radic. Biol. Med. 2005, 38, 78–84. [Google Scholar]
- Prior, R.L.; Wu, X.; Schauch, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar]
- Halliwell, B. Are polyphenols antioxidants or pro-oxidants? What do we learn from cell culture and in vivo studies? Arch. Biochem. Biophys. 2008, 476, 107–112. [Google Scholar]
- Disilvestro, R.A. Flavonoids as Antioxidants. In Handbook of Nutraceuticals and Functional Foods; Wildman, R.E.C., Ed.; CRC Press: Boca Raton, FL, USA, 2001; pp. 127–142. Chapter 8. [Google Scholar]
- Du, Y.; Guo, H.; Lou, H. Grape seed polyphenols protect cardiac cells from apoptosis via induction of endogenous antioxidant enzymes. J. Agric. Food Chem. 2007, 55, 1695–1701. [Google Scholar]
- Manach, C.; Donovan, J.L. Pharmacokinetics and metabolism of dietary flavonoids in humans. Free Radic. Res. 2004, 38, 771–785. [Google Scholar]
- Williamson, G.; Barron, D.; Shimoi, K.; Terao, J. In vitro biological properties of flavonoid conjugates found in vivo. Free Radic. Res. 2005, 39, 457–469. [Google Scholar]
- Rechner, A.R.; Kuhnle, G.; Bremner, P.; Hubbard, G.P.; Moore, K.P.; Rice-Evans, C.A. The metabolic fate of dietary polyphenols in humans. Free Radic. Biol. Med. 2002, 33, 220–235. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231-1246. https://doi.org/10.3390/nu2121231
Tsao R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients. 2010; 2(12):1231-1246. https://doi.org/10.3390/nu2121231
Chicago/Turabian StyleTsao, Rong. 2010. "Chemistry and Biochemistry of Dietary Polyphenols" Nutrients 2, no. 12: 1231-1246. https://doi.org/10.3390/nu2121231
APA StyleTsao, R. (2010). Chemistry and Biochemistry of Dietary Polyphenols. Nutrients, 2(12), 1231-1246. https://doi.org/10.3390/nu2121231