
Glycomacropeptide Safety and Its Effect on Gut Microbiota in Patients with Phenylketonuria: A Pilot Study

 , ,
, ,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cohort
2.2. Clinical Data Collection
2.2.1. Nutritional Assessment
2.2.2. Biochemistry
2.3. Short-Chain Fatty Acids Quantification
2.4. Gut Microbiota Sequencing and Analysis
2.5. Statistical Analysis
3. Results
3.1. Cohort Description
3.1.1. Body Composition Analysis
3.1.2. Dietary Assessment and Biochemical Analysis
3.1.3. Short-Chain Fatty Acid Quantification
3.2. Microbiota Characterization
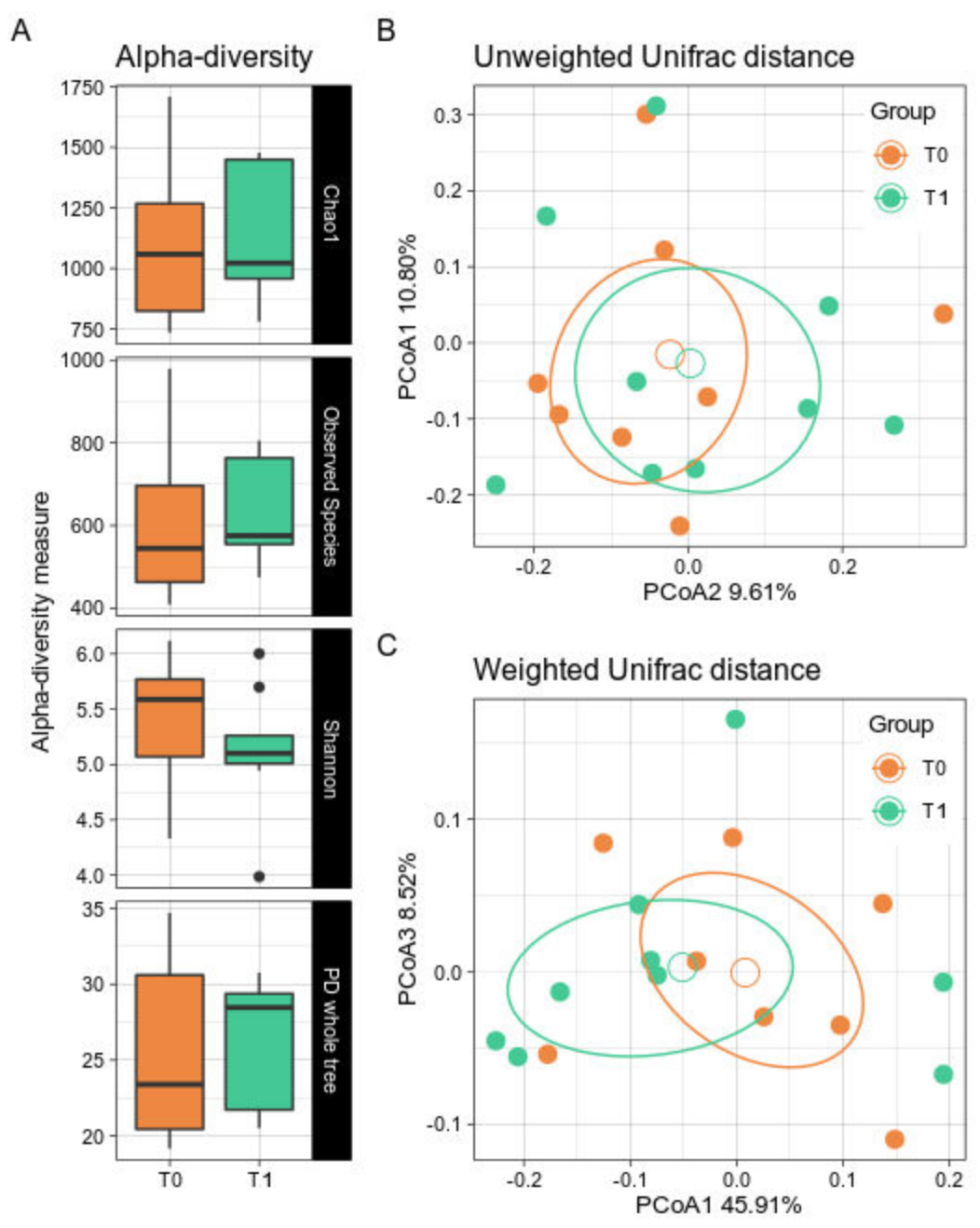
3.2.1. Biodiversity Assessment
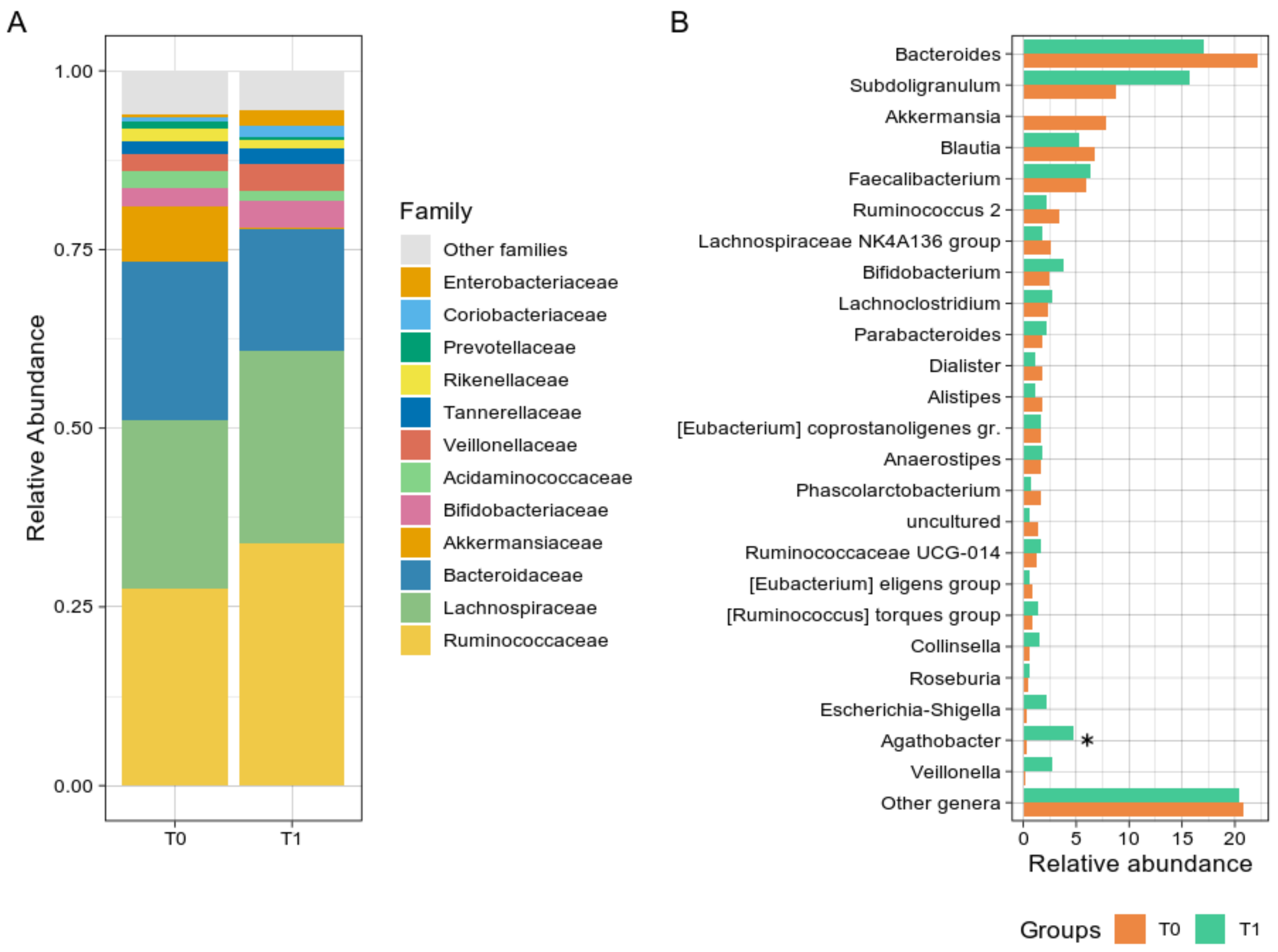
3.2.2. Taxonomic Comparisons
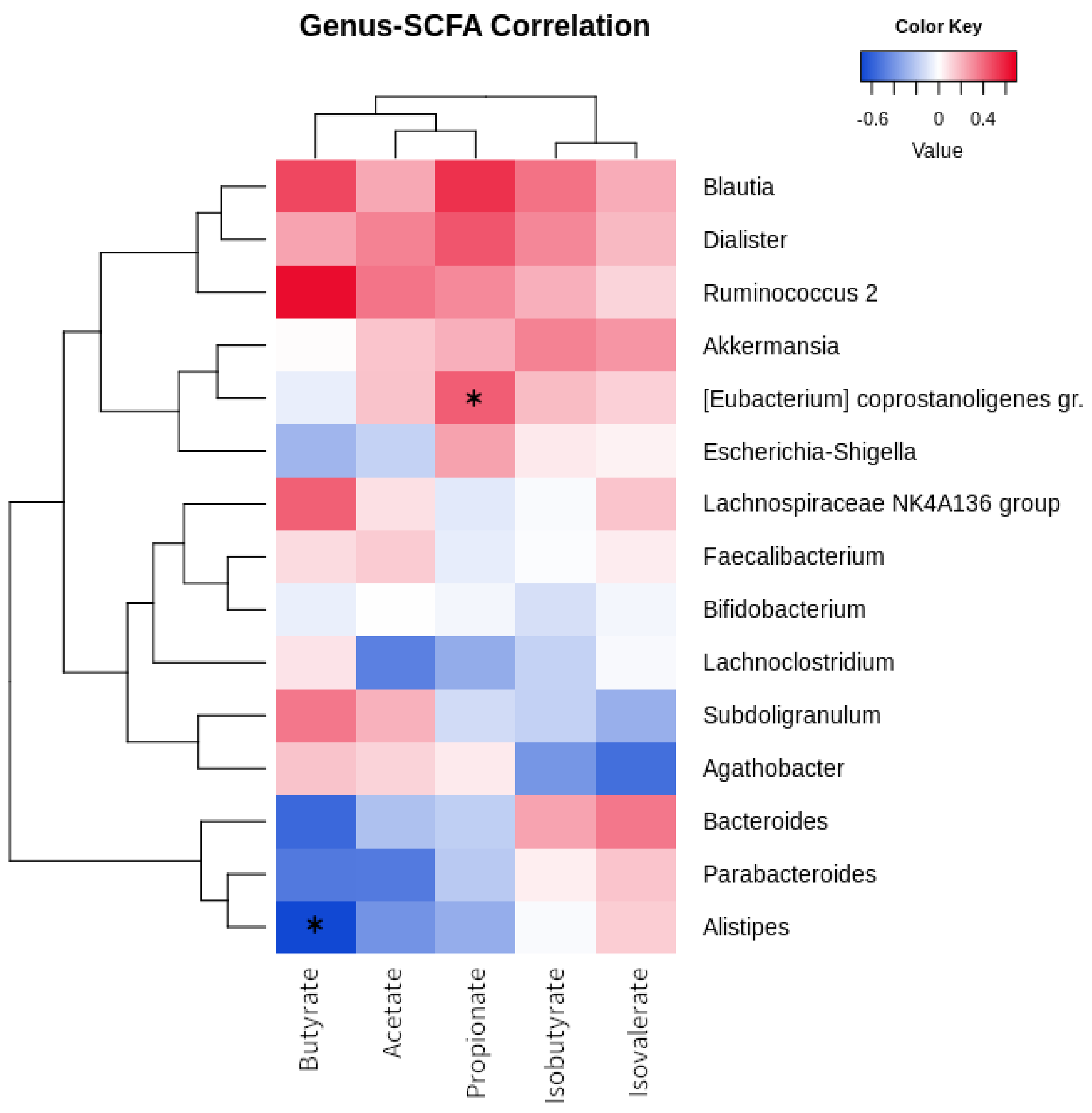
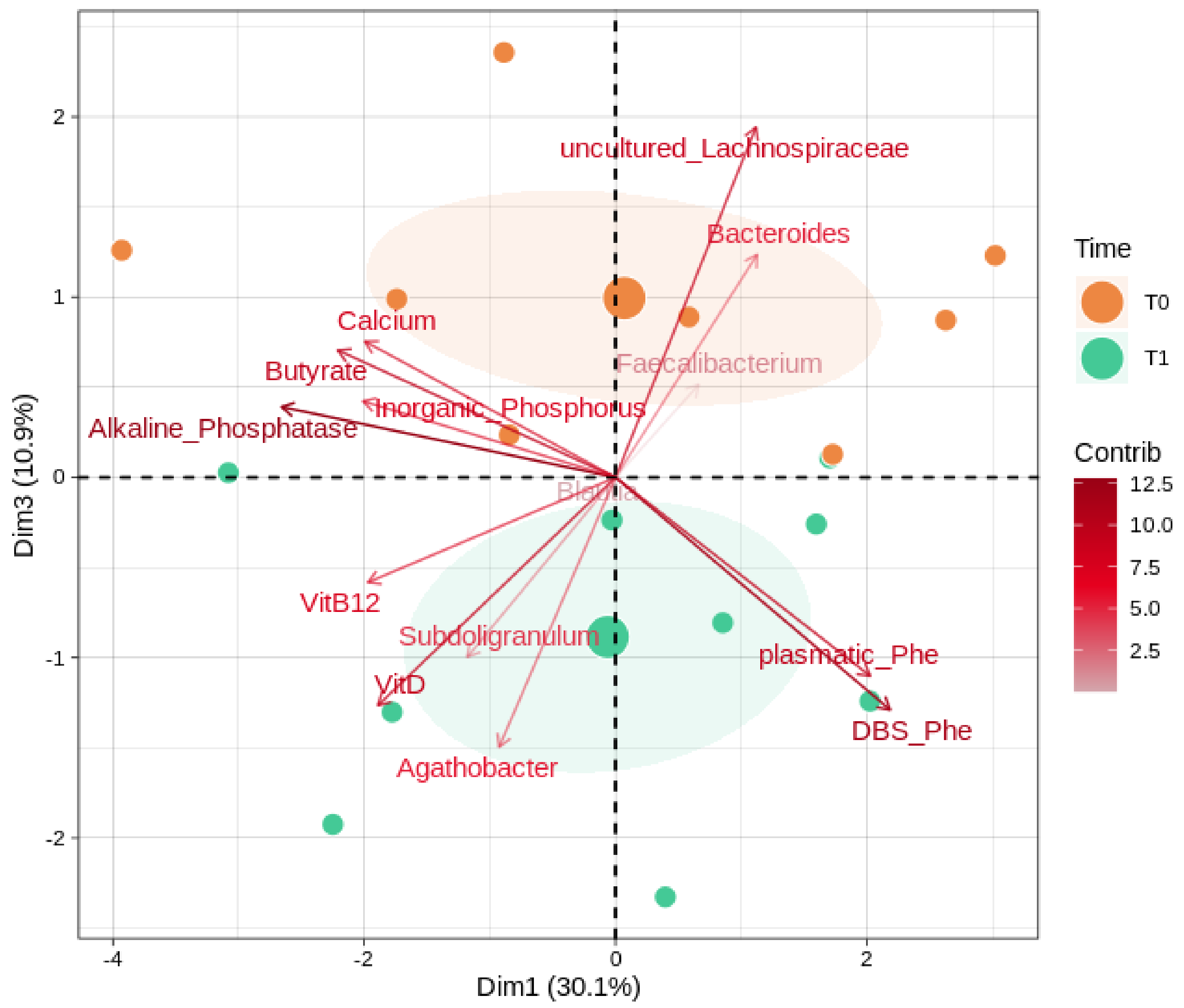
3.3. Correlations between Microbiota, SCFA, and Biochemical Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, R.H.; Rohr, F.; Frazier, D.; Cunningham, A.; Mofidi, S.; Ogata, B.; Splett, P.L.; Moseley, K.; Huntington, K.; Acosta, P.B.; et al. Recommendations for the Nutrition Management of Phenylalanine Hydroxylase Deficiency. Genet. Med. 2014, 16, 121–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Spronsen, F.J.; van Wegberg, A.M.; Ahring, K.; Bélanger-Quintana, A.; Blau, N.; Bosch, A.M.; Burlina, A.; Campistol, J.; Feillet, F.; Giżewska, M.; et al. Key European Guidelines for the Diagnosis and Management of Patients with Phenylketonuria. Lancet Diabetes Endocrinol. 2017, 5, 743–756. [Google Scholar] [CrossRef] [Green Version]
- Dominguez-Bello, M.G.; Godoy-Vitorino, F.; Knight, R.; Blaser, M.J. Role of the Microbiome in Human Development. Gut 2019, 68, 1108–1114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, H.; Tracey, K.J. Regulation of Macrophage Activation and Inflammation by Spermine: A New Chapter in an Old Story. Crit. Care Med. 2000, 28, N60–N66. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, J.; Lange, B.; Frick, J.-S.; Sauer, H.; Zimmermann, K.; Schwiertz, A.; Rusch, K.; Klosterhalfen, S.; Enck, P. A Vegan or Vegetarian Diet Substantially Alters the Human Colonic Faecal Microbiota. Eur. J. Clin. Nutr. 2012, 66, 53–60. [Google Scholar] [CrossRef]
- Harmsen, H.J.M.; Wildeboer-Veloo, A.C.M.; Grijpstra, J.; Knol, J.; Degener, J.E.; Welling, G.W. Development of 16S RRNA-Based Probes for the Coriobacterium Group and the Atopobium Cluster and Their Application for Enumeration of Coriobacteriaceae in Human Feces from Volunteers of Different Age Groups. Appl. Environ. Microbiol. 2000, 66, 4523–4527. [Google Scholar] [CrossRef] [Green Version]
- Verduci, E.; Moretti, F.; Bassanini, G.; Banderali, G.; Rovelli, V.; Casiraghi, M.C.; Morace, G.; Borgo, F.; Borghi, E. Phenylketonuric Diet Negatively Impacts on Butyrate Production. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 385–392. [Google Scholar] [CrossRef]
- Scott, K.P.; Gratz, S.W.; Sheridan, P.O.; Flint, H.J.; Duncan, S.H. The Influence of Diet on the Gut Microbiota. Pharmacol. Res. 2013, 69, 52–60. [Google Scholar] [CrossRef]
- Mancilla, V.J.; Mann, A.E.; Zhang, Y.; Allen, M.S. The Adult Phenylketonuria (PKU) Gut Microbiome. Microorganisms 2021, 9, 530. [Google Scholar] [CrossRef]
- Pinheiro de Oliveira, F.; Mendes, R.H.; Dobbler, P.T.; Mai, V.; Pylro, V.S.; Waugh, S.G.; Vairo, F.; Refosco, L.F.; Roesch, L.F.W.; Schwartz, I.V.D. Phenylketonuria and Gut Microbiota: A Controlled Study Based on Next-Generation Sequencing. PLoS ONE 2016, 11, e0157513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosca, A.; Leclerc, M.; Hugot, J.P. Gut Microbiota Diversity and Human Diseases: Should We Reintroduce Key Predators in Our Ecosystem? Front. Microbiol. 2016, 7, 455. [Google Scholar] [CrossRef] [Green Version]
- Verduci, E.; Carbone, M.T.; Borghi, E.; Ottaviano, E.; Burlina, A.; Biasucci, G. Nutrition, Microbiota and Role of Gut-Brain Axis in Subjects with Phenylketonuria (PKU): A Review. Nutrients 2020, 12, 3319. [Google Scholar] [CrossRef] [PubMed]
- Sawin, E.A.; De Wolfe, T.J.; Aktas, B.; Stroup, B.M.; Murali, S.G.; Steele, J.L.; Ney, D.M. Glycomacropeptide Is a Prebiotic That Reduces Desulfovibrio Bacteria, Increases Cecal Short-Chain Fatty Acids, and Is Anti-Inflammatory in Mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G590–G601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Le Roy, C.I. Effect of Diet on the Gut Microbiota: Rethinking Intervention Duration. Nutrients 2019, 11, E2862. [Google Scholar] [CrossRef] [Green Version]
- Lupori, L.; Cornuti, S.; Mazziotti, R.; Borghi, E.; Ottaviano, E.; Cas, M.D.; Sagona, G.; Pizzorusso, T.; Tognini, P. The Gut Microbiota of Environmentally Enriched Mice Regulates Visual Cortical Plasticity. Cell Rep. 2022, 38, 110212. [Google Scholar] [CrossRef]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-End Assembler for Illumina Sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of RRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A Comprehensive Online Resource for Quality Checked and Aligned Ribosomal RNA Sequence Data Compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘Vegan’. Community Ecol. Package Version 2013, 2, 1–295. [Google Scholar]
- Kuczmarski, R.J.; Ogden, C.L.; Grummer-Strawn, L.M.; Flegal, K.M.; Guo, S.S.; Wei, R.; Mei, Z.; Curtin, L.R.; Roche, A.F.; Johnson, C.L. CDC Growth Charts: United States. Adv. Data 2000, 314, 1–27. [Google Scholar]
- Ofenheimer, A.; Breyer-Kohansal, R.; Hartl, S.; Burghuber, O.C.; Krach, F.; Schrott, A.; Franssen, F.M.E.; Wouters, E.F.M.; Breyer, M.-K. Reference Charts for Body Composition Parameters by Dual-Energy X-Ray Absorptiometry in European Children and Adolescents Aged 6 to 18 Years-Results from the Austrian LEAD (Lung, HEart, SociAl, BoDy) Cohort. Pediatr. Obes. 2021, 16, e12695. [Google Scholar] [CrossRef]
- Kyle, U.G.; Genton, L.; Slosman, D.O.; Pichard, C. Fat-Free and Fat Mass Percentiles in 5225 Healthy Subjects Aged 15 to 98 Years. Nutrition 2001, 17, 534–541. [Google Scholar] [CrossRef]
- van Wegberg, A.M.J.; MacDonald, A.; Ahring, K.; Bélanger-Quintana, A.; Blau, N.; Bosch, A.M.; Burlina, A.; Campistol, J.; Feillet, F.; Giżewska, M.; et al. The Complete European Guidelines on Phenylketonuria: Diagnosis and Treatment. Orphanet J. Rare Dis. 2017, 12, 162. [Google Scholar] [CrossRef] [Green Version]
- Ntemiri, A.; Chonchúir, F.N.; O’Callaghan, T.F.; Stanton, C.; Ross, R.P.; O’Toole, P.W. Glycomacropeptide Sustains Microbiota Diversity and Promotes Specific Taxa in an Artificial Colon Model of Elderly Gut Microbiota. J. Agric. Food Chem. 2017, 65, 1836–1846. [Google Scholar] [CrossRef]
- Ahring, K.K.; Dagnæs-Hansen, F.; Brüel, A.; Christensen, M.; Jensen, E.; Jensen, T.G.; Johannsen, M.; Johansen, K.S.; Lund, A.M.; Madsen, J.G.; et al. The Effect of Casein Glycomacropeptide versus Free Synthetic Amino Acids for Early Treatment of Phenylketonuria in a Mice Model. PLoS ONE 2022, 17, e0261150. [Google Scholar] [CrossRef]
- Gustavo Hermes, R.; Molist, F.; Francisco Pérez, J.; Gómez de Segura, A.; Ywazaki, M.; Davin, R.; Nofrarías, M.; Korhonen, T.K.; Virkola, R.; Martín-Orúe, S.M. Casein Glycomacropeptide in the Diet May Reduce Escherichia Coli Attachment to the Intestinal Mucosa and Increase the Intestinal Lactobacilli of Early Weaned Piglets after an Enterotoxigenic E. Coli K88 Challenge. Br. J. Nutr. 2013, 109, 1001–1012. [Google Scholar] [CrossRef] [Green Version]
- Wernlund, P.G.; Hvas, C.L.; Dahlerup, J.F.; Bahl, M.I.; Licht, T.R.; Knudsen, K.E.B.; Agnholt, J.S. Casein Glycomacropeptide Is Well Tolerated in Healthy Adults and Changes Neither High-Sensitive C-Reactive Protein, Gut Microbiota nor Faecal Butyrate: A Restricted Randomised Trial. Br. J. Nutr. 2021, 125, 1374–1385. [Google Scholar] [CrossRef]
- Bassanini, G.; Ceccarani, C.; Borgo, F.; Severgnini, M.; Rovelli, V.; Morace, G.; Verduci, E.; Borghi, E. Phenylketonuria Diet Promotes Shifts in Firmicutes Populations. Front. Cell. Infect. Microbiol. 2019, 9, 101. [Google Scholar] [CrossRef]
- Moretti, F.; Pellegrini, N.; Salvatici, E.; Rovelli, V.; Banderali, G.; Radaelli, G.; Scazzina, F.; Giovannini, M.; Verduci, E. Dietary Glycemic Index, Glycemic Load and Metabolic Profile in Children with Phenylketonuria. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 176–182. [Google Scholar] [CrossRef]
- Scott, K.P.; Martin, J.C.; Duncan, S.H.; Flint, H.J. Prebiotic Stimulation of Human Colonic Butyrate-Producing Bacteria and Bifidobacteria, in Vitro. FEMS Microbiol. Ecol. 2014, 87, 30–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Shadike, Q.; Wang, M.; Jiang, H.; Liu, W.; Liu, J.; Tuerdi, R.; Zhou, W.; Li, L. A Low Abundance of Genus Bacteroides in Gut Microbiota Is Negatively Correlated with Blood Phenylalanine Levels in Uygur Patients with Phenylketonuria. Transl. Pediatr. 2021, 10, 2521–2532. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Lazarevic, V.; Derrien, M.; Girard, M.; Muccioli, G.G.; Muccioli, G.M.; Neyrinck, A.M.; Possemiers, S.; Van Holle, A.; François, P.; et al. Responses of Gut Microbiota and Glucose and Lipid Metabolism to Prebiotics in Genetic Obese and Diet-Induced Leptin-Resistant Mice. Diabetes 2011, 60, 2775–2786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hul, M.; Le Roy, T.; Prifti, E.; Dao, M.C.; Paquot, A.; Zucker, J.-D.; Delzenne, N.M.; Muccioli, G.G.; Clément, K.; Cani, P.D. From Correlation to Causality: The Case of Subdoligranulum. Gut Microbes 2020, 12, 1849998. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Thomas, R.L.; Jiang, L.; Adams, J.S.; Xu, Z.Z.; Shen, J.; Janssen, S.; Ackermann, G.; Vanderschueren, D.; Pauwels, S.; Knight, R.; et al. Vitamin D Metabolites and the Gut Microbiome in Older Men. Nat. Commun. 2020, 11, 5997. [Google Scholar] [CrossRef]
- Gaschott, T.; Werz, O.; Steinmeyer, A.; Steinhilber, D.; Stein, J. Butyrate-Induced Differentiation of Caco-2 Cells Is Mediated by Vitamin D Receptor. Biochem. Biophys. Res. Commun. 2001, 288, 690–696. [Google Scholar] [CrossRef]
- Cantorna, M.T. Vitamin D, Multiple Sclerosis and Inflammatory Bowel Disease. Arch. Biochem. Biophys. 2012, 523, 103–106. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Rawat, A.; Alwakeel, M.; Sharif, E.; Al Khodor, S. The Potential Role of Vitamin D Supplementation as a Gut Microbiota Modifier in Healthy Individuals. Sci. Rep. 2020, 10, 21641. [Google Scholar] [CrossRef]
- Adamczyk, P.; Morawiec-Knysak, A.; Płudowski, P.; Banaszak, B.; Karpe, J.; Pluskiewicz, W. Bone Metabolism and the Muscle-Bone Relationship in Children, Adolescents and Young Adults with Phenylketonuria. J. Bone Miner. Metab. 2011, 29, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Mendes, A.B.; Martins, F.F.; Cruz, W.M.S.; da Silva, L.E.; Abadesso, C.B.M.; Boaventura, G.T. Bone Development in Children and Adolescents with PKU. J. Inherit. Metab. Dis. 2012, 35, 425–430. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Dietary Reference Values for Vitamin D. EFSA J. 2016, 14, e04547. [Google Scholar] [CrossRef]
- Saggese, G.; Vierucci, F.; Prodam, F.; Cardinale, F.; Cetin, I.; Chiappini, E.; De’ Angelis, G.L.; Massari, M.; Miraglia Del Giudice, E.; Miraglia Del Giudice, M.; et al. Vitamin D in Pediatric Age: Consensus of the Italian Pediatric Society and the Italian Society of Preventive and Social Pediatrics, Jointly with the Italian Federation of Pediatricians. Ital. J. Pediatr. 2018, 44, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solverson, P.; Murali, S.G.; Litscher, S.J.; Blank, R.D.; Ney, D.M. Low Bone Strength Is a Manifestation of Phenylketonuria in Mice and Is Attenuated by a Glycomacropeptide Diet. PLoS ONE 2012, 7, e45165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stroup, B.M.; Sawin, E.A.; Murali, S.G.; Binkley, N.; Hansen, K.E.; Ney, D.M. Amino Acid Medical Foods Provide a High Dietary Acid Load and Increase Urinary Excretion of Renal Net Acid, Calcium, and Magnesium Compared with Glycomacropeptide Medical Foods in Phenylketonuria. J. Nutr. Metab. 2017, 2017, 1909101. [Google Scholar] [CrossRef] [Green Version]
- Daly, A.; Högler, W.; Crabtree, N.; Shaw, N.; Evans, S.; Pinto, A.; Jackson, R.; Strauss, B.J.; Wilcox, G.; Rocha, J.C.; et al. Growth and Body Composition in PKU Children-A Three-Year Prospective Study Comparing the Effects of L-Amino Acid to Glycomacropeptide Protein Substitutes. Nutrients 2021, 13, 1323. [Google Scholar] [CrossRef]
- Pena, M.J.; Pinto, A.; Daly, A.; MacDonald, A.; Azevedo, L.; Rocha, J.C.; Borges, N. The Use of Glycomacropeptide in Patients with Phenylketonuria: A Systematic Review and Meta-Analysis. Nutrients 2018, 10, E1794. [Google Scholar] [CrossRef] [Green Version]
- Daly, A.; Pinto, A.; Evans, S.; MacDonald, A. Glycomacropeptide in PKU-Does It Live Up to Its Potential? Nutrients 2022, 14, 807. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Median Value at T0; IQR (n = 9) | Median Value at T1; IQR (n = 9) | p-Value |
|---|---|---|---|
| Protein nutritional status | |||
| Albumine (g/dL) | 4.6; 0.3 | 4.5; 0.5 | 0.078 |
| Transthyretin (prealbumin) (mg/dL) | 24.8; 3.3 | 24.2; 5.1 | 0.426 |
| Total protein (mg/dL) | 7.5; 0.3 | 7.5; 0.8 | 0.068 |
| Vitamin and mineral status | |||
| Vitamin D (25-OH) (ng/mL) | 32.2; 11.7 | 44.7; 21.9 | 0.027 |
| Calcium (mg/dL) | 9.9; 0.3 | 9.7; 0.4 | 0.138 |
| Phosphorus (mg/dL) | 4; 0.6 | 3.7; 0.5 | 0.024 |
| Vitamin B12 (pg/mL) | 854; 255 | 846; 74 | 0.359 |
| Folate (ng/mL) | 9.7; 7.2 | 15; 6.1 | 0.059 |
| Alkaline phosphatase (U/L) | 102; 115 | 72; 52 | 0.027 |
| Glucose metabolism | |||
| Glucose (mg/dL) | 83; 11 | 86; 8 | 0.905 |
| Insulin (µU/mL) | 5.6; 4.1 | 5.1; 4.7 | 0.297 |
| Lipid profile | |||
| Total Cholesterol (mg/dL) | 160; 14 | 163; 11 | 0.570 |
| LDL Cholesterol (mg/dL) | 96; 33 | 89; 26 | 0.820 |
| HDL Cholesterol (mg/dL) | 52; 16 | 54; 21 | 0.105 |
| Triglycerides (mg/dL) | 66; 41 | 89; 50 | 0.635 |
| Iron status | |||
| Plasmatic iron (µg/dL) | 90; 38 | 117; 74 | 0.250 |
| Ferritin (ng/mL) | 45.2; 23.9 | 46.3; 8.9 | 0.652 |
| Transferrin (mg/dL) | 258; 42.0 | 243; 32 | 0.075 |
| Metabolic control | |||
| Blood Phe values (µmol/L) a | 300; 222.6 | 391.8; 94.8 | 0.496 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montanari, C.; Ceccarani, C.; Corsello, A.; Zuvadelli, J.; Ottaviano, E.; Dei Cas, M.; Banderali, G.; Zuccotti, G.; Borghi, E.; Verduci, E. Glycomacropeptide Safety and Its Effect on Gut Microbiota in Patients with Phenylketonuria: A Pilot Study. Nutrients 2022, 14, 1883. https://doi.org/10.3390/nu14091883
Montanari C, Ceccarani C, Corsello A, Zuvadelli J, Ottaviano E, Dei Cas M, Banderali G, Zuccotti G, Borghi E, Verduci E. Glycomacropeptide Safety and Its Effect on Gut Microbiota in Patients with Phenylketonuria: A Pilot Study. Nutrients. 2022; 14(9):1883. https://doi.org/10.3390/nu14091883
Chicago/Turabian StyleMontanari, Chiara, Camilla Ceccarani, Antonio Corsello, Juri Zuvadelli, Emerenziana Ottaviano, Michele Dei Cas, Giuseppe Banderali, Gianvincenzo Zuccotti, Elisa Borghi, and Elvira Verduci. 2022. "Glycomacropeptide Safety and Its Effect on Gut Microbiota in Patients with Phenylketonuria: A Pilot Study" Nutrients 14, no. 9: 1883. https://doi.org/10.3390/nu14091883