1. Introduction
Branched-chain amino acids (BCAAs; valine, leucine, isoleucine) have been linked to the development of type 2 diabetes, insulin resistance, and obesity in both rodents and humans [
1,
2,
3,
4,
5]. It is unclear whether this association is a direct reflection of the diet, or the consequence of systemic metabolic derangement [
3,
6,
7]. In mice, increasing intake of proteins and BCAAs is associated with both elevated circulating BCAA levels and changes in body composition [
3,
4,
6,
8]. In humans, however, the link between dietary BCAAs and health is less clear. While some studies linking high circulating levels of BCAAs with metabolic dysregulation also show increases in total protein and/or BCAA intake [
1,
2,
9,
10,
11], other observational studies report conflicting evidence, with some showing positive, negative, or neutral effects of dietary protein and/or BCAAs on health [
12,
13].
Therefore, the question of how closely the relationship between diet, BCAAs, and body composition in rodents reflect the relationship in humans is an important issue to resolve. In animal studies, elucidating the relationship between diet, circulating BCAAs, and body composition can be uniquely informative, controlling for various factors that could not otherwise be achieved in a free-living human population. However, translating these findings in order to humans to better understand the role of BCAAs in health requires that key outcomes in both rodents and humans respond similarly.
Here, we directly compare the relationship between circulating BCAAs, energy and macronutrient intake, and body composition in a group of men aged over 75 years with a cohort of older mice at 15 months of age. We show that the relationship between circulating BCAAs, intake, and body composition were comparable across both free-living humans and mice under tightly controlled dietary manipulations, suggesting that preclinical findings in mice may reliably inform studies examining the link between BCAAs and body composition in humans.
2. Materials and Methods
2.1. Human Cohort
2.1.1. Study Design
The Concord Health and Ageing in Men Project (CHAMP) is a longitudinal cohort study investigating the health of older men based in Sydney, Australia. The recruitment, which is described elsewhere [
14], involved a defined urban region (the Local Government Areas of Burwood, Canada Bay, and Strathfield near Concord Hospital in Sydney, Australia) sampled using the New South Wales Electoral Roll. A total of 1705 men participated in the initial study wave in 2005–2006 [
14]. Dietary data were assessed through diet history interviews at the third wave (five-year follow-up, 2012). At the five-year follow-up, 954 participants completed questionnaires and clinical assessments, and of those, 794 completed diet history interviews. CHAMP was approved by the Sydney South West Area Health Service Human Ethics Committee, Concord Repatriation General Hospital, Sydney, Australia (Protocol number HREC/10/CRGH/28). Written consent was obtained from all participants and the study was conducted in accordance with the World Medical Association Helsinki Declaration.
2.1.2. Dietary Data
Details about CHAMP dietary data collection and validation have been previously described [
15]. Briefly, usual dietary intake (past 3 months) was assessed through diet history interviews [
16]; those interviews followed a standardised protocol and were conducted at participants’ residences by a trained dietitian. Validation of this method was obtained through comparison with a 4-day weighed food record collected in a subgroup (
n = 56) of participants [
17].
2.1.3. Outcome Variables
Body weight was measured according to a standardised protocol. All fasting blood tests were performed at the Diagnostic Pathology Unit of Concord RG Hospital, which is a National Australian Testing Authority (NATA) accredited pathology service, using a MODULAR Analytics system (Roche Diagnostics, Castle Hill, Australia). Amino acids were analysed at the Australian Proteome Analysis Facility, Macquarie University, using the Waters AccQ-Tag Ultra Chemistry Kit (Waters Corporation, Milford, MA, USA). Dual X-ray absorptiometry (DXA) using the fan beam Discovery-W scanner (Hologic Inc., Bedford, MA, USA) was used to obtain individuals’ whole-body scans. Men removed jewellery and wore a light cotton gown free from metal. The head was excluded from all measures and lean mass was calculated as the difference between non-fat mass and bone mineral content.
2.1.4. Covariates
Information on socio-demographic, lifestyle, and economic factors were obtained through a self-completed questionnaire. Activity level was determined through the Physical Activity Scale for the Elderly (PASE) [
18]. Source of income was categorised as means-tested age pension only and other (repatriation pension, veteran’s pension, superannuation or other private income, own business/farm/partnership, wage or salary, other or any source of income combination). Source of income was used as a proxy of personal income, assuming that age pensioners had the lowest income. Participants self-reported whether they have been diagnosed (by a doctor or a health care provider) with diabetes, thyroid problems, osteoporosis, Paget’s disease, stroke, Parkinson’s disease, kidney stones, dementia, depression, epilepsy, hypertension, myocardial infarction, angina, heart failure, intermittent claudication, chronic obstructive lung disease, liver disease, chronic kidney disease, arthritis, or cancer (excluding non-melanotic skin cancer and benign tumours such as bowel polyps and meningioma), and number of morbidities was calculated as the sum of diagnosed conditions.
2.2. Mouse Cohort
2.2.1. Animals and Husbandry
A total of 384 C57BL/6J mice of both sexes were purchased from the Animal Resource Centre (Perth, WA) at 3 weeks old. Mice were group-housed in cages of four on a 12 h light–dark cycle at the Charles Perkins Centre at the University of Sydney. Mice were housed at 22–24 °C and were given ad libitum access to food and water. At 12 weeks of age, mice were randomly allocated to one of four experimental diets, monitored weekly, with food and body weights recorded bi-weekly until 6 months of age, and monthly thereafter. Animals remained on allocated diets throughout their lifetime. At 15 months of age, one mouse from each cage was sacrificed with tissues and blood collected for analysis (
n = 12 males and 12 females). For comparisons between BCAAs and body composition, 19–23 mice per diet treatment were analysed. Data relating to cardiometabolic health and brain ageing have been published previously [
19]. Experiments were approved by the University of Sydney’s Animal Ethics Committee (protocol no. 2014/752).
2.2.2. Experimental Diets
Four experimental diets were designed and manufactured in dry-pelleted form by Specialty Feeds (Perth, Australia). Diets had the same energy density (14.4 kJ/g), were matched in net metabolizable energy from fat (18%), but varied in protein (P) and carbohydrate (C) content. Diets consisted of either 5% P and 77% C, 10% P and 72% C, 15% P and 67% C, or 19% P and 63% C. Diets were based on the rodent diet AIN-93G and formulated to contain all essential vitamins, minerals, and amino acids for growth in mice. Casein was the primary protein component, the main carbohydrate component was wheatstarch, and the main fat component was soy oil.
2.2.3. Outcome Variables
Body composition and plasma amino acids were assessed in each mouse at 15 months of age. Body composition was assessed by EchoMRI (EchoMRI 900, Houston, TX, USA). Fat mass (g) and lean mass (g) were measured for each mouse and % body fat and % body lean calculated. Free amino acids were quantified in plasma. Targeted LC-QQQ-MS analysis was employed to detect metabolites. Metabolomic analysis was conducted using a tandem liquid chromatography–mass spectrometry (LC-MS/MS) system (Agilent 1260 Infinity liquid chromatography coupled to a QTRAP 5500 mass spectrometer (AB SCIEX, Framingham, USA). A hydrophilic interaction chromatography (HILIC) column was used for the detection of BCAAs. All raw data files (Analyst software, version 1.6.2; AB Sciex, Foster City, CA, USA) were imported into Multi-QuantTM 3.0 Software for MRM Q1/Q3 peak integration. A detailed description of metabolite profiling is described by O’Sullivan, et al. [
6,
20].
2.2.4. Statistical Analyses
Data were analysed in R (v.3.4.1, R Foundation for Statistical Computing, Vienna, Austria) and significance considered when
p < 0.05. Data from males and females were combined to analyse various outcomes in mice. Regression analyses were performed using the
lm function in the base package [
21,
22]. Univariate analyses were performed to investigate the associations between circulating BCAAs, energy and macronutrient intake, and body composition variables. Two sample
t-tests were performed to assess the mean difference between categorical variables (income and smoking status). Linear regressions were used to investigate the association between BCAA, age, physical activity level, and number of comorbidities. Subsequently, multivariate analyses were performed to adjust firstly for age and then for all factors commonly associated with dietary intake and circulating BCAA levels in humans. Correlation matrices were generated in R using the ”
corrplot” package [
23].
3. Results
The human cohort characteristics have been described elsewhere [
15,
23]. Briefly, the mean age was 81.3 years (SD = 4.6), body weight was 78.0 kg (SD = 12.78), fat mass 21.6 kg (SD = 7.22), percentage fat 29.5% (SD = 6.06), lean mass 47.8 kg (SD = 6.24), percentage lean 67.6% (SD = 5.90), and the majority of participants were non-smokers (96%), did not rely on the age pension as sole source of income (59%), and had at least two comorbidities. A total of 83 mice were analysed to determine the relationship of BCAAs with body composition. Mean body weight was 40.5 g (SD = 7.81), fat mass 14.0 g (SD = 5.59), percentage fat 33.4% (SD = 9.37), lean mass 24.1 g (SD = 3.74) and percentage lean 60.5% (SD = 8.79).
3.1. Circulating BCAAs and Body Composition
In humans, circulating BCAA levels significantly decreased with age (
p < 0.001,
Supplementary Materials, Figure S1). Body weight, fat, and lean mass as well as percentage body fat were positively associated with circulating BCAAs, whereas percentage body lean mass was negatively associated with BCAAs in both the human cohort and mice model, with stronger correlation in mice (
Figure 1). After adjustment for age, physical activity, number of morbidities, smoking status, and source of income, the association between circulating BCAAs and all body composition measures remained statistically significant in the human cohort (
Table 1). Individual BCAAs were also correlated with body composition variables in
Figure 1F,L. We found that all outcomes, except for % body lean, were positively correlated with valine, isoleucine, and leucine—a result consistent in both mice and men.
3.2. Circulating BCAAs and Diet
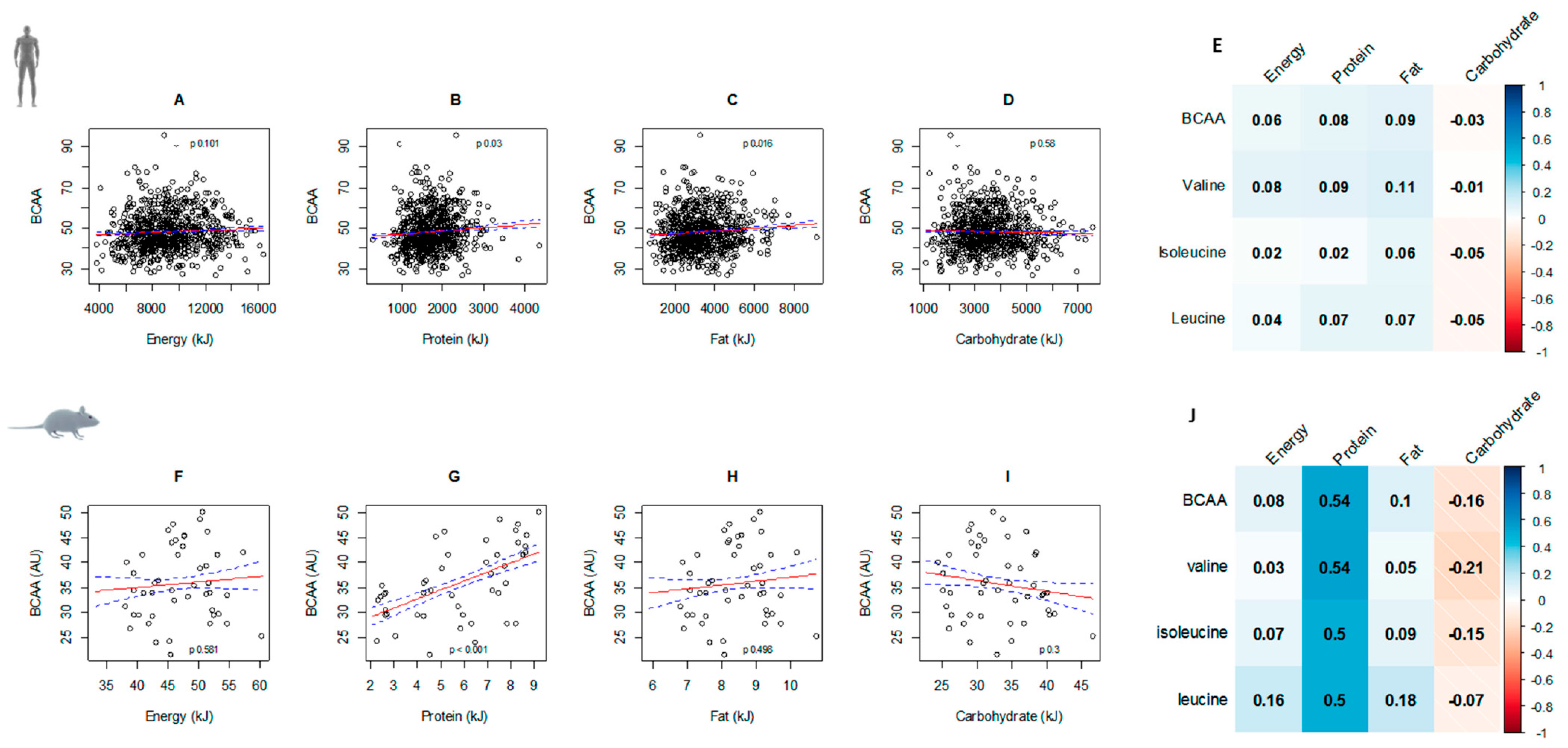
The association between energy, macronutrient intakes, and circulating BCAAs were similar in both mice and humans. Higher intakes of protein were significantly associated with higher circulating BCAAs and while similar trends occurred for energy intake, this was not statistically significant. Similarly, the negative relationship between BCAAs and carbohydrates showed a similar pattern in both species, with the correlation stronger in mice compared to humans. Fat intake was positively associated with circulating BCAAs in humans (
p = 0.016), with a similar—albeit non-significant—trend in mice (
Figure 2). Correlation matrices of individual BCAAs show similar relationships with energy and macronutrient intakes in both mice and men. With the exception of carbohydrate intake, all other nutrient intakes were positively correlated with valine, isoleucine, and leucine. As with total BCAA levels, individual amino acids were most strongly correlated with protein intake in mice, a result consistent with our previous findings [
4].
4. Discussion
There has been limited research into the relationship between blood levels of BCAAs and aging. Studies have suggested that BCAAs decline with old age [
24,
25,
26]. There have also been studies of the relationship between BCAAs and frailty or sarcopenia, which are classic age-related syndromes, some of which have shown an association with low BCAA levels [
27,
28,
29,
30]. Although it is unclear exactly why BCAAs decline with age, age-related changes in muscle protein metabolism, such as an imbalance in the rate of protein synthesis and breakdown, increased resistance to anabolic factors, changes in levels of physical activity, and an overall reduction in dietary intake of protein may be contributing factors [
31,
32]. Here, we demonstrate that total BCAA levels and individual levels of isoleucine, leucine, and valine, not only declined with age in humans, but were also linked with body composition, with those with higher circulating BCAA levels more likely to be carrying more weight and fat mass. Because the increase in fat mass outweighed the increase in lean mass, there was a corresponding increase in body fat percentage and a decrease in body lean percentage. Strikingly, we found that the association between BCAA and body composition markers were similar in mice models and in the human cohort of older males.
In terms of the associations between dietary intakes and circulating BCAA levels, as expected, a clear association was demonstrated between protein intake and circulating BCAA levels in mice. This association was not as clear in older men, though it followed the same trend. Plasma BCAA levels are likely to reflect long-term protein intake [
33], yet high protein intake can increase circulating concentrations of BCAAs, with up to 80% of consumed BCAAs reaching circulation [
34,
35]. A number of factors, such as insulin resistance [
36], cardiovascular disease [
37], body composition [
7], and protein sources [
5], can also influence circulating BCAA levels. In the current study, we investigated the correlation between protein and BCAA levels in mice kept in a well-controlled environment for their whole life, whereas for the human cohort, dietary intake was estimated through diet history interviews, which may also explain the weaker association between protein intake and circulating BCAAs in the human cohort. Another potential explanation for the difference in the strength of association between protein intake and BCAA levels in mice and humans was the source of protein: mice were provided with casein protein, whereas protein sources were varied in the human cohort [
15].
In recent years, BCAAs have received increased attention for their role in muscle synthesis and as a potential intervention option to improve muscle quality; a number of clinical studies have pointed to a crucial role of BCAAs in tackling muscle loss in older individuals. It has been suggested that the increase of protein or amino acids alone (as opposed to combining high protein or amino acids with high carbohydrate and/or energy) should result in better outcomes in older individuals [
31,
38]. The current study indicates that carbohydrate intakes have a negative association with circulating BCAA levels in both mice and humans.
The current study has several strengths worth mentioning: (1) data from a large sample of community-dwelling men and a large sample of mice on a well-defined diet and tightly controlled environmental conditions were used in our analyses; (2) the use of gold-standard methodology to obtain body-composition measures in humans (DXA) and in mice (EchoMRI); and (3) the dietary assessment method used (diet history) and frequent food intake measurements in mice. It is also worth mentioning the limitations of the current study: firstly, causal association cannot be established from the human cohort results; secondly, there may be other factors that play an important role in the relationship between dietary intake, BCAA levels, and body composition, such as genetic background and long-term dietary intake. Finally, the results may not be applicable to older females.
5. Conclusions
This is the first study to investigate the role of BCAA levels in large cohorts of mice and humans. Higher protein intakes were associated with higher BCAA levels and higher body weight and fat mass in both species. Further studies are required to elucidate the association between BCAA levels and body composition in older age, but this study adds to the body of evidence showing that mouse models can provide valuable insight into the relationship between nutrition and health, which can be translated to humans.
Author Contributions
R.V.R., S.M.S.-B. and D.G.L.C. developed the concepts for this paper. The CHAMP study was designed by R.G.C., V.N., D.J.H., M.J.S., D.G.L.C., L.M.W. and F.M.B. S.M.S.-B., D.G.L.C., and S.J.S. designed the mouse study. S.M.S.-B., T.P., V.C.C., X.C., J.O., V.C.C. and Y.C.K. performed mouse experiments. V.H., A.M.S., A.G., R.G.C., F.M.B., V.N., L.M.W., D.J.H., M.J.S., S.J.S., F.S. and D.G.L.C. collaborated in writing. All authors reviewed and approved the final version of the manuscript. All authors had primary responsibility for final content.
Funding
This research was funded by National Health and Medical Research Council (NHMRC) grant number 301916 and GNT1084267.
Acknowledgments
We thank all the CHAMP’s staff and participants for their contributions, Sydney Medical School Foundation and the Ageing, Alzheimer’s Institute and Jennie Mackenzie for their support. SSB is supported by the NHMRC Peter Doherty Biomedical Fellowship (GNT1110098) and the University of Sydney SOAR fellowship. RVR was supported by a Charles Perkins Centre Early Career Fellowship from Jennie Mackenzie.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fontana, L.; Cummings, N.E.; Apelo, S.I.A.; Neuman, J.C.; Kasza, I.; Schmidt, B.A.; Cava, E.; Spelta, F.; Tosti, V.; Syed, F.A.; et al. Decreased consumption of branched chain amino acids improves metabolic health. Cell Rep. 2016, 16, 520–530. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A Branched-Chain Amino Acid-Related Metabolic Signature that Differentiates Obese and Lean Humans and Contributes to Insulin Resistance. Cell Metab. 2009, 9, 565–566. [Google Scholar] [CrossRef] [Green Version]
- Maida, A.; Chan, J.S.; Sjøberg, K.A.; Zota, A.; Schmoll, D.; Kiens, B.; Herzig, S.; Rose, A.J. Repletion of branched chain amino acids reverses mTORC1 signaling but not improved metabolism during dietary protein dilution. Mol. Metab. 2017, 6, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Solon-Biet, S.M.; McMahon, A.C.; Ballard, J.W.O.; Ruohonen, K.; Wu, L.E.; Cogger, V.C.; Warren, A.; Huang, X.; Pichaud, N.; Melvin, R.G.; et al. The Ratio of Macronutrients, Not Caloric Intake, Dictates Cardiometabolic Health, Aging, and Longevity in Ad Libitum-Fed Mice. Cell Metab. 2014, 19, 418–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousseau, M.; Guénard, F.; Garneau, V.; Allam-Ndoul, B.; Lemieux, S.; Pérusse, L.; Vohl, M.C. Associations Between Dietary Protein Sources, Plasma BCAA and Short-Chain Acylcarnitine Levels in Adults. Nutrients 2019, 11, 173. [Google Scholar] [CrossRef] [PubMed]
- Solon-Biet, S.M.; Cogger, V.C.; Pulpitel, T.; Wahl, D.; Clark, X.; Bagley, E.E.; Gregoriou, G.C.; Senior, A.M.; Wang, Q.P.; Brandon, A.E.; et al. Branched-chain amino acids impact health and lifespan indirectly via amino acid balance and appetite control. Nat. Metab. 2019, 1, 532–545. [Google Scholar] [CrossRef]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef] [Green Version]
- Cummings, N.E.; Williams, E.M.; Kasza, I.; Konon, E.N.; Schaid, M.D.; Schmidt, B.A.; Poudel, C.; Sherman, D.S.; Yu, D.; Apelo, S.I.A.; et al. Restoration of metabolic health by decreased consumption of branched-chain amino acids. J. Physiol. 2017, 596, 623–645. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.J.; Larson, M.G.; Vasan, R.S.; Cheng, S.; Rhee, E.P.; McCabe, E.; Lewis, G.D.; Fox, C.S.; Jacques, P.F.; Fernandez, C.; et al. Metabolite Profiles and the Risk of Developing Diabetes. Nat. Med. 2011, 17, 448–453. [Google Scholar] [CrossRef]
- Shah, S.H.; Crosslin, D.R.; Haynes, C.S.; Nelson, S.; Turer, C.B.; Stevens, R.D.; Muehlbauer, M.J.; Wenner, B.R.; Bain, J.R.; Laferrere, B.; et al. Branched-chain amino acid levels are associated with improvement in insulin resistance with weight loss. Diabetologia 2012, 55, 321–330. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, Y.; Qi, Q.; Hruby, A.; Manson, J.E.; Willett, W.C.; Wolpin, B.M.; Hu, F.B.; Qi, L. Cumulative consumption of branched-chain amino acids and incidence of type 2 diabetes. Int. J. Epidemiol. 2016, 45, 1482–1492. [Google Scholar] [CrossRef] [Green Version]
- Bifari, F.; Nisoli, E. Branched-chain amino acids differently modulate catabolic and anabolic states in mammals: A pharmacological point of view. Br. J. Pharmacol. 2017, 174, 1366–1377. [Google Scholar] [CrossRef]
- D’Antona, G.; Ragni, M.; Cardile, A.; Tedesco, L.; Dossena, M.; Bruttini, F.; Caliaro, F.; Corsetti, G.; Bottinelli, R.; Carruba, M.O.; et al. Branched-Chain Amino Acid Supplementation Promotes Survival and Supports Cardiac and Skeletal Muscle Mitochondrial Biogenesis in Middle-Aged Mice. Cell Metab. 2010, 12, 362–372. [Google Scholar] [CrossRef] [Green Version]
- Cumming, R.G.; Handelsman, D.; Seibel, M.J.; Creasey, H.; Sambrook, P.; Waite, L.; Naganathan, V.; Le Couteur, D.; Litchfield, M. Cohort Profile: The Concord Health and Ageing in Men Project (CHAMP). Int. J. Epidemiol. 2009, 38, 374–378. [Google Scholar] [CrossRef]
- Waern, R.V.; Cumming, R.G.; Blyth, F.; Naganathan, V.; Allman-Farinelli, M.; Le Couteur, D.; Simpson, S.J.; Kendig, H.; Hirani, V. Adequacy of nutritional intake among older men living in Sydney, Australia: Findings from the Concord Health and Ageing in Men Project (CHAMP). Br. J. Nutr. 2015, 114, 812–821. [Google Scholar] [CrossRef]
- Burke, B. The dietary history as a tool in research. J. Am. Diet. Assoc. 1947, 23, 1041–1046. [Google Scholar]
- Rosilene, W.; Cumming, R.; Travison, T.; Blyth, F.; Naganathan, V.; Allman-Farinelli, M.; Hirani, V. Relative validity of a diet history questionnaire against a four-day weighed food record among older men in Australia: The Concord Health and Ageing in Men Project (CHAMP). J. Nutr. Health Aging 2015, 19, 603–610. [Google Scholar] [CrossRef]
- Washburn, R.A.; Smith, K.W.; Jette, A.M.; Janney, C.A. The physical activity scale for the elderly (PASE): Development and evaluation. J. Clin. Epidemiol. 1993, 46, 153–162. [Google Scholar] [CrossRef]
- Wahl, D.; Solon-Biet, S.M.; Wang, Q.P.; Wali, J.A.; Pulpitel, T.; Clark, X.; Raubenheimer, D.; Senior, A.M.; Sinclair, D.A.; Cooney, G.J.; et al. Comparing the Effects of Low-Protein and High-Carbohydrate Diets and Caloric Restriction on Brain Aging in Mice. Cell Rep. 2018, 25, 2234–2243. [Google Scholar] [CrossRef]
- Patsch, C.; Challet-Meylan, L.; Thoma, E.C.; Urich, E.; Heckel, T.; O’Sullivan, J.F.; Grainger, S.J.; Kapp, F.G.; Sun, L.; Christensen, K.; et al. Generation of vascular endothelial and smooth muscle cells from human pluripotent stem cells. Nat. Cell Biol. 2015, 17, 994. [Google Scholar] [CrossRef]
- Chambers, J.M. Chapter 4 of Statistical Models in S. In Linear Models; Wadsworth & Brooks/Cole: Pacific Grove, CA, USA, 1992. [Google Scholar]
- Wilkinson GNaR, C.E. Symbolic descriptions of factorial models for analysis of variance. Appl. Stat. 1973, 22, 392–399. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.84); R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Chaleckis, R.; Murakami, I.; Takada, J.; Kondoh, H.; Yanagida, M. Individual variability in human blood metabolites identifies age-related differences. Proc. Natl. Acad. Sci. USA 2016, 113, 4252–4259. [Google Scholar] [CrossRef] [Green Version]
- Kouchiwa, T.; Wada, K.; Uchiyama, M.; Kasezawa, N.; Niisato, M.; Murakami, H.; Fukuyama, K.; Yokogoshi, H. Age-related changes in serum amino acids concentrations in healthy individuals. Clin. Chem. Lab. Med. 2012, 50, 861–870. [Google Scholar] [CrossRef]
- Oja, S.S.; Kemppainen, K.; Mero, A.A.; Pitkänen, H.T.; Seppä, J.M. Serum amino acid concentrations in aging men and women. Amino Acids 2003, 24, 413–421. [Google Scholar]
- Calvani, R.; Picca, A.; Marini, F.; Biancolillo, A.; Gervasoni, J.; Persichilli, S.; Primiano, A.; Coelho-Junior, H.; Bossola, M.; Urbani, A.; et al. A Distinct Pattern of Circulating Amino Acids Characterizes Older Persons with Physical Frailty and Sarcopenia: Results from the Biosphere Study. Nutrients 2018, 10, 1691. [Google Scholar] [CrossRef]
- Adachi, Y.; Ono, N.; Imaizumi, A.; Muramatsu, T.; Andou, T.; Shimodaira, Y.; Nagao, K.; Kageyama, Y.; Mori, M.; Noguchi, Y.; et al. Plasma amino acid profile in severely frail elderly patients in Japan. Int. J. Gerontol. 2018, 12, 290–293. [Google Scholar] [CrossRef]
- Moaddel, R.; Fabbri, E.; Khadeer, M.A.; Carlson, O.D.; Gonzalez-Freire, M.; Zhang, P.; Semba, R.D.; Ferrucci, L. Plasma Biomarkers of Poor Muscle Quality in Older Men and Women from the Baltimore Longitudinal Study of Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 1266–1272. [Google Scholar] [CrossRef] [Green Version]
- Ter Borg, S.; Luiking, Y.C.; van Helvoort, A.; Boirie, Y.; Schols, J.; de Groot, C. Low Levels of Branched Chain Amino Acids, Eicosapentaenoic Acid and Micronutrients Are Associated with Low Muscle Mass, Strength and Function in Community-Dwelling Older Adults. J. Nutr. Health Aging 2019, 23, 27–34. [Google Scholar] [CrossRef]
- Fujita, S.; Volpi, E. Amino acids and muscle loss with aging. J. Nutr. 2006, 136 (Suppl. 1), 277S–280S. [Google Scholar] [CrossRef]
- Ottestad, I.; Ulven, S.M.; Øyri, L.K.L.; Sandvei, K.S.; Gjevestad, G.O.; Bye, A.; Sheikh, N.A.; Biong, A.S.; Andersen, L.F.; Holven, K.B. Reduced plasma concentration of branched-chain amino acids in sarcopenic older subjects: A cross-sectional study. Br. J. Nutr. 2018, 120, 445–453. [Google Scholar] [CrossRef]
- Würtz, P.; Mäkinen, V.P.; Soininen, P.; Kangas, A.J.; Tukiainen, T.; Kettunen, J.; Savolainen, M.J.; Tammelin, T.; Viikari, J.S.; Rönnemaa, T.; et al. Metabolic Signatures of Insulin Resistance in 7098 Young Adults. Diabetes 2012, 61, 1372–1380. [Google Scholar]
- Layman, D.K.; Baum, J.I. Dietary Protein Impact on Glycemic Control during Weight Loss. J. Nutr. 2004, 134, 968S–973S. [Google Scholar] [CrossRef]
- Rietman, A.; Schwarz, J.; Tomé, D.; Kok, F.J.; Mensink, M. High dietary protein intake, reducing or eliciting insulin resistance? Eur. J. Clin. Nutr. 2014, 68, 973. [Google Scholar] [CrossRef]
- Gannon, N.P.; Schnuck, J.K.; Vaughan, R.A. BCAA Metabolism and Insulin Sensitivity—Dysregulated by Metabolic Status? Mol. Nutr. Food Res. 2018, 62, 1700756. [Google Scholar] [CrossRef]
- Du, X.; You, H.; Li, Y.; Wang, Y.; Hui, P.; Qiao, B.; Lu, J.; Zhang, W.; Zhou, S.; Zheng, Y.; et al. Relationships between circulating branched chain amino acid concentrations and risk of adverse cardiovascular events in patients with STEMI treated with PCI. Sci. Rep. 2018, 8, 15809. [Google Scholar] [CrossRef]
- Wolfe, R.R. Branched-chain amino acids and muscle protein synthesis in humans: Myth or reality? J. Int. Soc. Sports Nutr. 2017, 14, 30. [Google Scholar] [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).

 ,
, 


{kind=link}
{kind=link}